Fisher's theory predicts equal sex ratios at the end of parental care if the cost associated with raising offspring of each sex is equal. However, sex ratios have important evolutionary consequences and are often biased for many factors. Reported sex ratios are often biased in raptors, which display various degrees of reversed sexual dimorphism, but there seems no consistent pattern in their offspring sex ratios. In this study, we investigated the offspring sex ratio of the Eurasian Kestrel (Falco tinnunculus) and tested whether the patterns of biased sex ratios were related to laying order, egg mass, hatching order, laying date or clutch size. The brood sex ratio of the Eurasian Kestrel (male) in eggs was 47.0%, not statistically biased from 0.5, but in fledglings it was 40.8%, significantly biased from 0.5 (p=0.029). At population level, both primary and secondary sex ratios did not depart from parity. We found that clutch size and egg mass affected the secondary brood sex ratio, i.e., the larger the clutch size, the larger the number of males and eggs producing sons were heavier than eggs producing daughters. Laying date affected both the primary and secondary sex ratios, and laying earlier is associated with a greater proportion of males.
Over the past 40 years there have been intensive theoretical and experimental investigations of how natural selection moulds the sex ratio. Fisher's (1930) classic frequency-dependent model of sex allocation predicts that the offspring sex ratio should not deviate from parity as long as the costs of producing males and females are equal. Modern empirical sex ratio research began when Hamilton (1967) observed that many insects and mites have highly female-biased sex ratios and that this trait is associated with high levels of brother-sister mating. Hamilton explained this result by his theory of local mate competition, which has been extended to other cases of interactions between siblings. Briefly, if one sex suffers a greater reduction in fitness through competition with siblings of the same sex, the mother is programmed in her selection of the sex ratio to favor the sex that suffers less competition. Fisher's frequency-dependent argument is thus modified in populations where relatives compete. A second major development was by Trivers and Willard (1973) who originally proposed that maternal quality should affect the direction of this investment for species, exhibiting high variation in male reproductive success. Modern classic sex allocation theory predicts that selection should act on parents to sex-specifically vary the level of investment in offspring when the fitness returns differ for the two sexes (Charnov, 1982; Frank, 1990; Hardy, 2002).
Birds have long been considered incapable of facultatively shifting their primary sex ratio because their sex determination is chromosomal (Clutton-Brock, 1986). However, recent studies have suggested that sex ratio adjustment in birds is not as constrained as thought earlier (West and Sheldon, 2002). In birds, the sex-ratio adjustment can occur at laying period (primary sex-ratio adjustment) and during the period of provisioning for the young (secondary sex-ratio adjustment) (Ellegren and Sheldon, 1997; Kilner, 1998).
Mechanisms for both primary and secondary sex-ratio adjustment in birds remain largely unknown (Emlen, 1997; Sheldon, 1998; Krackow, 1999; Martins, 2004), but preovulatory mechanisms have been suggested as the most efficient means of primary sex-ratio adjustment for some species (Komdeur et al., 2002). Pike and Petrie (2003) presented an excellent review of the potential mechanisms of avian sex manipulation, such as steroid hormones, paternal attractiveness, maternal condition, photoperiod and phenotypic sex and others. In addition, much new experimental and relevant evidence have appeared since then, providing new and interesting perspectives (Pike and Petrie, 2006; Rutkowska and Badyaev, 2008).
Sex allocation patterns among raptor populations have attracted considerable research attention because most raptorial birds exhibit reversed sexual size dimorphism (females are the larger sex) (Newton and Marquiss, 1979; Olsen and Cockburn, 1991; Appleby et al., 1997; Brommer et al., 2003; McDonald et al., 2005). One consequence of this size difference is a disparity in food requirements: daughters require more food than sons do during the nestling period (Anderson et al., 1993a; Krijgsveld et al., 1998; Riedstra et al., 1998; Laaksonen et al., 2004). Rearing larger daughters is more costly since they require more parental investment in the form of food requirements as opposed to sons. Theoretically, the offspring sex ratio in raptors should be male-biased. However, there seems no consistent pattern in the offspring sex ratios of raptors; the proportion of males is either increasing (Newton and Marquiss, 1979; Olsent and Cockburn, 1991; Korpimäki et al., 2000) or decreasing (Tella et al., 1996) and no clear adaptive explanations for these trends have been found so far.
The Eurasian Kestrel (Falco tinnunculus), a sexually size-dimorphic raptor (females ca. 20% heavier than males; Village, 1990), widespread in open country throughout the Palearctic, Afrotropical and Oriental regions (Cramp and Simmons, 1980). A total of 12 subspecies have been identified (Dickinson, 2003). The subspecies in our study is F. tinnunculus interstinctus. The female Eurasian Kestrel might be able to adjust her reproductive effort by promoting sex differences in the mass of first-laid eggs (Blanco et al., 2003), where sexual differences in the size of first-laid eggs defines the pattern of within-brood mass hierarchies of fledglings. In addition, some researchers have documented that females have the potential ability to adjust resources allocated to different-sex eggs laid in varying order (Anderson et al., 1997; Blanco et al., 2003; Martínez-Padilla and Fargallo, 2007).
We investigated the primary and secondary offspring sex ratio of the Eurasian Kestrel at the Zuojia Nature Reserve in northeastern China. Our objectives were: 1) to test whether its offspring sex ratio demonstrates a general deviation from a balanced sex ratio and 2) whether a pattern of biased sex ratio exists in relation to laying order, egg mass, hatching order, laying date and clutch size.
Study area and methods
Study area
We conducted the study in the Zuojia Nature Reserve in Jilin Province, northeastern China during the breeding seasons (from late March to late July) in 2007 and 2008. The Zuojia Nature Reserve stretches from the eastern Changbai mountains to the western plains (126°1′–127°2′N, 44°6′–45°5′E), where the elevation ranges from 200 m to 530 m. The region is subject to an eastern monsoon climate, characterized by hot, dry summers and cold, snowy winters; the area is covered by approximately 35% open habitat and 65% forests. Vegetation in the area is diverse and the existing forests are secondary.
Data collection
The Eurasian Kestrels do not build their own nests, but rely mainly on the stick nests of the Magpie (Pica pica) in our study area (Xiang et al., 1991; Deng et al., 2006; Zhou et al., 2009). Most magpie nests used by the kestrels were inaccessible. We therefore installed nest boxes to develop the study and most kestrels then shifted to breed in these boxes.
The nest boxes we used in the study were constructed from rough-cut boards and put together with exterior nails or deck screws. The internal dimensions of the boxes were all the same: 50 cm deep, a 35 cm × 35 cm bottom area and with a 12.5 cm or 15 cm diameter entrance hole near the top. These nest boxes were placed randomly about 8–13 m above ground in various tree species with distances ranging from 50 m to 150 m between the nearest two boxes. The number of boxes monitored was 68 in 2007 and 77 in 2008.
We monitored nest boxes used by the Eurasian Kestrels every day to record the laying date and marked the laying sequence of each egg accurately during the egg laying period. We used an electronic balance to weigh eggs to the nearest 0.01 g on the day of being laid. In order to assign all marked eggs to their corresponding nestlings, and record the hatching order, we checked nests hourly (between 08:00 and 19:00 hours) on the days of expected hatching. We distinguished nestlings by drawing some simple symbols on their head with indelible ink. We removed unhatched eggs from the nest after several days when the rest of the clutch had hatched. We discarded eggs containing no visible embryos, and stored dead embryos in 100% ethanol. After hatching, we visited nests at an interval of two days to collect body tissues of the dead nestlings as a source of DNA and to monitor fledging success. We collected a blood sample of each nestling (20–50 μL) from brachial veins with a sterilized hypodermic needle, 9 to 11 days after hatching. We stored blood samples in EDTA for determination of sex in the laboratory.
Sex determination
We sexed the nestlings using a polymerase chain reaction (PCR) amplification based on the technique used by Fridolfsson and Ellegren (1999). We isolated DNA from the red blood cells using proteinase K digestion and phenol-chloroform extraction. We ran PCR amplification using a particular set of primers (2550F and 2718R), as suggested by Fridolfsson and Ellegren (1999). The temperature profile in PCR was as follows: hotstart at 94℃, initial denaturation at 94℃ for 5 min, followed by a "touch-down" scheme (Don et al., 1991) where the annealing temperature was lowered 1℃ per cycle, starting from 58℃, until a temperature of 48℃ was reached. Then 25–35 additional cycles were run at a constant annealing temperature of 48℃. Denaturation was at 94℃ for 30 s, annealing for 45 s and extension at 72℃ for 1 min. A final extension step of 5 min was added after the last cycle. PCR products, separated by electrophoresis on 2% agarose gels containing ethidium bromide, were visualized under UV light. We assigned the sex of individuals from five adult pairs correctly using this technique, so we could be confident the method worked successfully in this species.
Statistics analysis
We used two data sets to analyze the offspring sex ratio. The data set for analyzing the primary sex ratio included broods that all laid eggs had hatched successfully, including dead embryos. We sampled all nestlings (both blood and tissue samples) for a total of 81 nestlings in 15 broods. The data set for analyzing the secondary sex ratio included 105 nestlings in 23 successfully fledged broods. We examined the brood sex ratio (hereafter referred to as the proportion of males) using the binomial test (Zar, 1984). We used the Wilcoxon signed rank test in S-PLUS 6.0 to test the primary and secondary sex ratios at population level. In order to study the relationship between primary sex ratio, laying order, clutch size and egg mass, we performed linear mixed model (LMM) in SPSS 14.0 (SPSS Inc., Chicago, Illinois, USA). In this model, we entered the sex ratio as the response variable, clutch size and laying order as fixed factors. The kestrel hatches asynchronously with a hatching spread of 3–5 days and therefore we divided the laying order into three categories: initial, middle and last. The initial category corresponded to the first eggs and the last category to the last eggs. The other eggs were defined as the middle category (Hardy, 2002). Thus, the sex ratio used in the model stood for the sex ratio of each category. Moreover, egg mass in the model also represented the egg mass of each category. We encoded the laying dates into five stages: March, early April (April Ⅰ), middle April (April Ⅱ), late April (April Ⅲ) and May and introduced it as a random factor, with egg mass as covariate. By introducing the laying date as a higher level random effect in addition to the residual error term, our LMM permitted an estimation of the fixed effects and interaction parameters while accounting for random variation in the sex ratio between broods. We analyzed the relationship between secondary sex ratio, laying order, clutch size, hatching order and egg mass using the same model. For all statistical tests, we used a significance level of 0.05. We presented values as means ± SD.
Results
Sex ratio at brood and population level
The brood sex ratio of the Eurasian Kestrel in eggs was 47.0±4.3%, not statistically biased from 0.5 (Wilcoxon signed rank test, Z15 = –0.402, p = 0.342). However, the brood sex ratio in fledglings was 40.8±4.5%, significantly different from 0.5 (Wilcoxon signed rank test, Z23 = –1.882, p = 0.029).
The primary sex ratio at population level was 48.2%, which did not depart from parity (binomial test, n = 81, p = 0.824). The sex ratio in eggs for the two study years showed a similar pattern (binomial test, 48.9%, n = 45, p = 1.000 in 2007; 47.2%, n = 36, p = 0.868 in 2008). The secondary sex ratio at population level was 42.9%, which did not depart significantly from parity (binomial test, n = 105, p = 0.172). Moreover, we detected the same tendency in nestlings (binomial test, 41.9%, n = 62, p = 0.253 in 2007; 44.2%, n = 43, p = 0.542 in 2008).
Effect of laying order, egg mass, clutch size on primary brood sex ratio
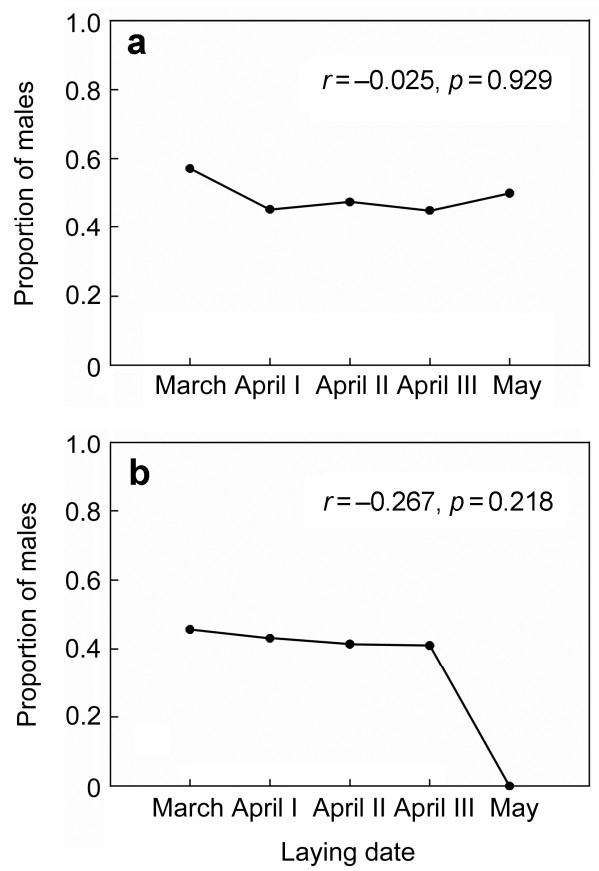
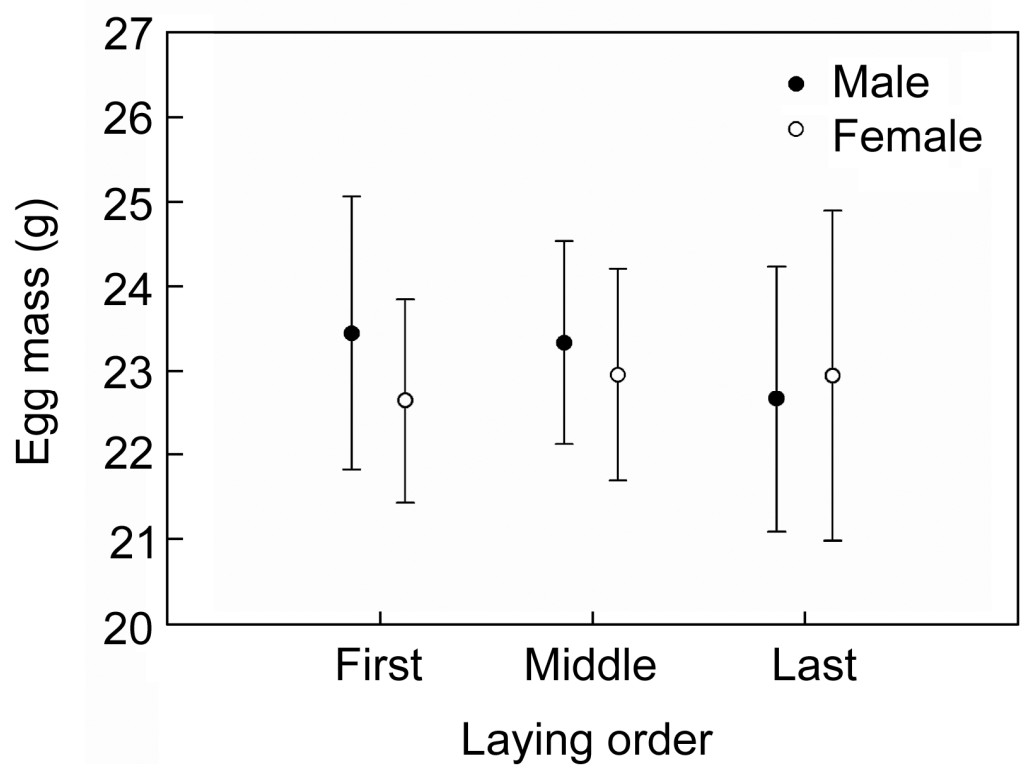
The sex ratio of eggs for different categories of laying order did not differ from parity (binomial tests: initial, 40.0%, n = 15, p = 0.607; middle, 43.1%, n = 51, p = 0.401; last, 73.3%, n = 15, p = 0.118). Results of LMM showed that the sex ratio in eggs was not associated with laying order, clutch size and their interaction (Table 1). Our results indicate that the laying date had a significant impact on primary offspring sex (Table 1), but there was no significant correlation (r = –0.025, p = 0.929) (Fig. 1a). Egg mass did not have a significant effect on primary sex ratio, but the egg mass of males (23.15±1.36, n = 42) was slightly heavier than that of females (22.89±1.28, n = 38), although the difference was not statistically significant (Mann-Whitney test U = 712, p = 0.407). In addition, egg mass was not associated linearly with laying order; the eggs in the middle position in the laying order were the heaviest (Fig. 2).
Table
1.
LMM analyses of the effects of clutch size and laying order on the sex of the eggs in Eurasian Kestrels
Source
NDF
DDF
FNDF, DDF
p
Clutch size
3
8
1.570
0.271
Laying order
1
8
0.004
0.951
Egg mass
1
8
1.796
0.217
Clutch size × Laying order
1
8
0.000
0.983
Covariance parameter estimates:
Estimate
SE
Z
p
Laying date
0.262
0.130
2.000
0.046
The model accounts for random variation between laying date. NDF represents the degrees of freedom of the numerator and DDF that of the denominator. FNDF, DDF stands for Fisher's test statistics, p represents the significance level, "estimate" represents the estimated covariance parameters, SE stands for standard error, Z represents Z statistic value of the Wald test.
Effects of laying order, egg mass, clutch size and hatching order on secondary brood sex ratio
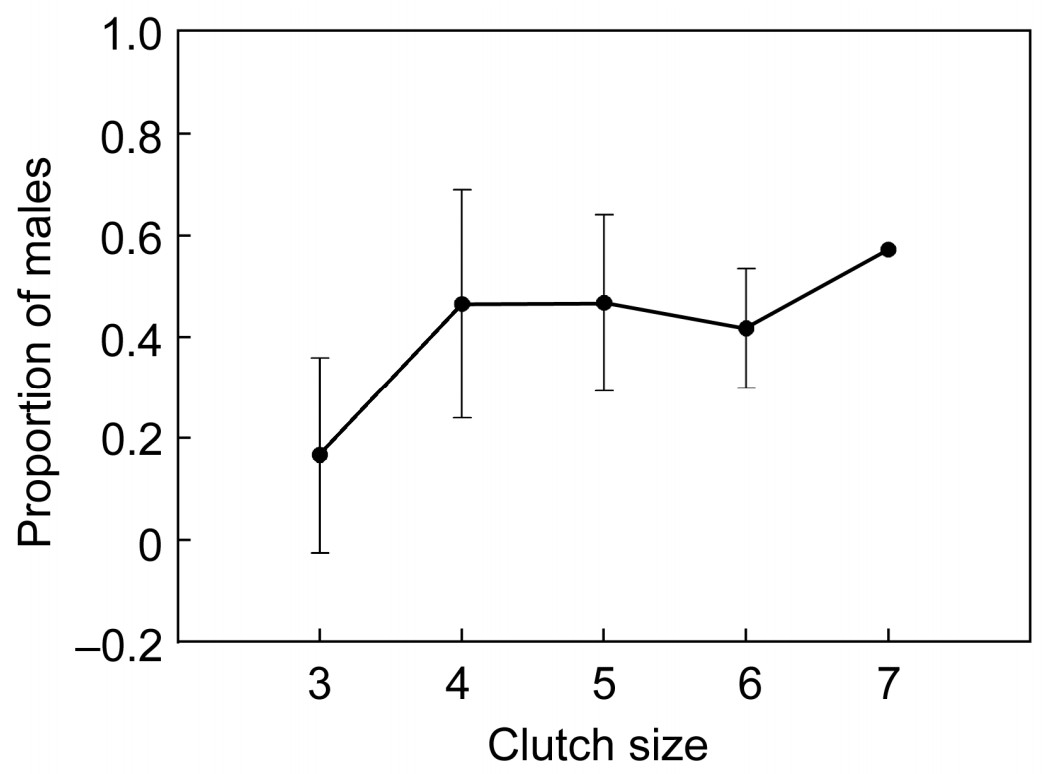
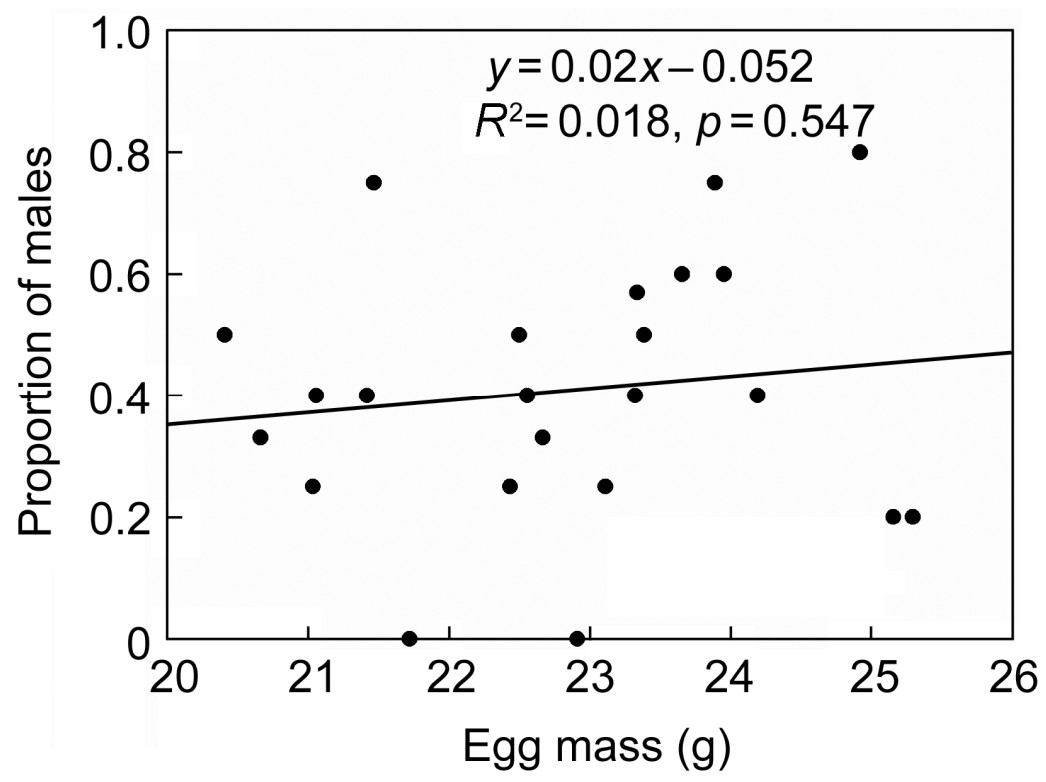
The secondary brood sex ratio was significantly related to clutch size (F = 5.297, p = 0.017) and egg mass (F = 8.099, p = 0.016). With increasing clutch size and egg mass, the proportion of males became gradually higher, but there were no significantly positive correlations (Figs. 3 and 4). Laying order and hatching order did not have a significant effect on the proportion of males in fledglings; however, when they interacted with clutch size, both factors had a significant effect on the secondary brood sex ratio (Table 2). In addition, the LMM showed that laying date had a significant effect on secondary offspring sex (Table 1), but there was no significant correlation (r = –0.267, p = 0.218) (Fig. 1b).
Figure
3.
Relationship between secondary brood sex ratio and clutch size in Eurasian Kestrel in northeastern China
Table
2.
LMM analyses of the effects of clutch size, laying order and hatching order on the sex of the fledgings in Eurasian Kestrel
Source
NDF
DDF
FNDF, DDF
p
Clutch size
3
11
5.297
0.017
Laying order
1
11
0.851
0.376
Hatching order
1
11
0.128
0.728
Egg mass
1
11
8.099
0.016
Clutch size × Laying order
1
11
6.447
0.028
Clutch size × Hatching order
1
11
5.508
0.039
Covariance parameter estimates:
Estimate
SE
Z
p
Laying date
0.194
0.008
2.345
0.019
The model accounts for random variation between laying date. NDF is the degree of freedom of the numerator and DDF of the denominator, FNDF, DDF is Fisher's test statistic, p represents significance level, "estimate" represents the estimated covariance parameters, SE stands for standard error, and Z represents the Z statistic value of the Wald test.
In this study, the sex ratio at population level and the primary brood sex ratio did not depart from parity, which was similar to the result obtained by Ankney (1982). We found that the secondary brood sex ratio of Eurasian Kestrels was consistently female-biased across the two-year study. A number of studies have shown that females can adjust sex ratio of their offspring by adjusting the laying and hatching orders (Ankney, 1982; Kilner, 1998; Cichoń et al., 2003; Ležalová et al., 2005). However, our results, as another study (Martínez-Padilla and Fargallo, 2007) on the species, indicated that the laying order did not affect the sex ratio of Eurasian Kestrels. Interestingly, our results showed that clutch size and egg mass only affected secondary brood sex ratio, while the laying date affected both primary and secondary sex ratios.
For many bird species, there is no a consistent relationship between egg mass and sex ratio, because females might adjust egg mass either positively or negatively to correlate with the laying order in assigning resources to different sexual offspring (Fiala, 1981; Ankney, 1982; Bancroft, 1984). Anderson et al. (1997) suggested that the sex ratio of offspring was significantly associated with the laying order and that male eggs were significant larger than female eggs in the American Kestrel (F. sparverius). In our study, egg mass had a significant effect on the secondary brood sex ratio and showed the same pattern as the American Kestrel in spite of a lack of significant difference between female and male eggs. Therefore, we speculate that the sexually size-dimorphic Eurasian Kestrel was able to adjust the sex ratio of its offspring by adopting different strategies of egg mass provisioning according to egg sex and laying order.
Some studies showed that the fledgling sex ratios of several raptor species are skewed toward males early in the season while late broods had an excess of females (Dijkstra et al., 1990; Daan et al., 1996; Tella et al., 1996; Griggio et al., 2002). In our study, the laying date had a significant effect on the primary and secondary brood sex ratios. Moreover, both sex ratios showed a slightly decreasing trend as the season progressed in spite of a lack of significant shift. Eurasian Kestrel nestlings are sexually dimorphic, with daughters larger than sons. The larger daughters may need more foods and have an advantage during sibling competition for food (Anderson et al., 1993a, 1993b). Sex-specific characteristics of nestlings may affect the reproductive biology of the species, while nestling mortality should fall more heavily on sons than on daughters. We suggest that the seasonal variation in the sex ratio of the Eurasian Kestrel might be associated with favouritism for the male function on the part of the parents, both to counter the risk of excess mortality of sons and because sons require less parental care per capita during earlier breeding seasons.
A large number of studies show that clutch size does not affect the sex ratio of birds, except for the study by Wegge (1980), who indicated that in a declining population of Capercaillie (Tetrao urogallus), the sex ratio of fledglings declined with clutch size, but that this was probably a consequence of differential mortality between hatchings and fledgings. In our study, clutch size had a significant effect on the secondary sex ratio in spite of a lack of significant correlation. The highest male proportion occurred with a clutch size of seven, with the highest probability of mortality of hatchings from the seventh egg. Two potential reasons may explain this conclusion. First, we found that the Eurasian Kestrel, bred in an artificial nest box, had a larger clutch size than kestrels bred in natural nests (5.77±0.15 vs 4.86± 0.26), where clutch size of the species was not more than 6 in natural nests (W. Gao unpublished data). As a result, it was difficult for the parents to rear all the nestlings in this condition. We therefore considered that the kestrel may adopt a "brood reduction strategy" in order to adjust the secondary brood sex ratio (Slagsvold et al., 1984). Secondly, food availability might also constrain the survival rate of nestlings. Kestrels feed mainly on small mammals, especially voles, but birds, frogs and insects also form part of their diet during the nestling period, which might reflect a reduced availability of rodents due to their being preyed upon continuously by the kestrels and several other raptor species in the study area (Geng et al., 2009).
Our results revealed relatively complex patterns of sex allocation in the Eurasian Kestrel that appears to be associated with both maternal ability to modify the sex of the progeny by adapting to its environment. Future work on the kestrel should include experiments that evaluate the ultimate mechanism of sex ratio allocation and parental provisioning patterns to different sex eggs with egg mass, adjusting for reproductive efforts.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (Grant No. 30400047), the Training Fund of Northeast Normal University Scientific Innovation Project (Grant No. 07013) and the Jilin Provincial Science and Technology Department (Grant No. 20070558 and 20090575). The authors thank Shi Jiao, Bo Qin and Yufang Qin for assistance in the field and anonymous reviewers for providing comments to improve the manuscript.
Anderson DJ, Budde C, Apanius V, Gomez JEM, Bird DM, Weathers WW. 1993a. Prey size influences female competitive dominance in nestling American kestrels. Ecology, 74: 367–376
Anderson DJ, Reeve J, Bird DM. 1997. Sexually dimorphic eggs, nestling growth and sibling competition in American Kestrels (Falco sparverius). Funct Ecol, 11: 331–335
Anderson DJ, Reeve J, Gomez JEM, Weathers WW, Hutson S, Cunningham HV, Bird DM. 1993b. Sexual size dimorphism and food requirements of nestling birds. Can J Zool, 71: 2541–2545
Ankney CD. 1982. Sex ratio varies with egg sequence in Lesser Snow Geese. Auk, 99: 662–666
Appleby BM, Petty SJ, Blakey JK, Raieny P, Macdonald DW. 1997. Does variation of sex ratio enhance reproductive success of offspring in Tawny Owls (Strix aluco). Proc R Soc B, 264: 1111–1116
Bancroft GT. 1984. Patterns of variation in size of Boat-tailed Grackle (Quiscalus major) eggs. Ibis, 126: 496–509
Bize P, Roulin A, Telia JL, Richner H. 2005. Female-biased mortality in experimentally parasitized Alpine Swift (Apus melba) nestlings. Funct Ecol, 19: 405–413
Blanco G, Martínez-Padilla J, Serrano D, Dávila JA, Viñuela J. 2003. Mass provisioning to different-sex eggs within the laying sequence: Consequences for adjustment of reproductive effort in a sexually dimorphic bird. J Anim Ecol, 72: 831–838
Brommer JE, Karell P, Pihlaja T, Painter JN, Primmer CR, Pietiäinen H. 2003. Ural Owl sex allocation and parental investment under poor food conditions. Oecologia, 137: 140–147
Burley N. 1981. Sex ratio manipulation and selection for attractiveness. Science, 211: 721–722
Cassey P, Ewen JG, Møller AP. 2006. Revised evidence for facultative sex ratio adjustment in birds: a correction. Proc R Soc B, 273: 3129–3130
Charnov EL. 1982. The theory of sex allocation. Monogr Popul Biol, 18: 1–355
Cichoń M, Dubiec A, Stoczke M. 2003. Laying order and offspring sex in blue tits Parus caeruleus. J Avian Biol, 34: 355–359
Clutton-Brock TH. 1986. Sex ratio variation in birds. Ibis, 128: 317–329
Cramp S, Simmons KEL. 1980. Handbook of the Birds of Europe, the Middle East and North Africa. Oxford University Press, Oxford
Daan S, Dijkstra C, Weissing FJ. 1996. An evolutionary explanation for seasonal trends in avian sex ratios. Behav Ecol, 7: 426–430
Deng QX, Gao W, Yang YL, Zhou T. 2006. The role of magpie in formed bird community organism in secondary forest. J Northeast Normal Univ, 38: 101–104 (in Chinese)
Dickinson EC. 2003. The Howard and Moor Complete Checklist of the Birds of the World. 3rd ed. Princeton University Press, Princeton
Dijkstra C, Daan S, Buker JB. 1990. Adaptive seasonal variation in the sex ratio of kestrel broods. Funct Ecol, 4: 143–147
Don RH, Cox PT, Wainwright BJ, Baker K, Mattick JS. 1991. 'Touchdown' PCR to circumvent spurious priming during gene amplification. Nucl Acids Res, 19: 4008
Ellegren H, Gustafsson L, Sheldon BC. 1996. Sex ratio adjustment in relation to paternal attractiveness in a wild bird population. Proc Natl Acad Sci USA, 15: 11723–11728
Ellegren H, Sheldon BC. 1997. New tools for sex identification and the study of sex allocation in birds. Trends Ecol Evol, 12: 255–259
Emlen ST. 1997. When mothers prefer daughters over sons. Trends Ecol Evol, 12: 291–292
Fargallo JA, Polo V, De Neve L, Martin J, Davila JA, Soler M. 2006. Hatching order and size-dependent mortality in relation to brood sex ratio composition in chinstrap penguins. Behav Ecol, 17: 772–778
Ferree ED. 2007. White tail plumage and brood sex ratio in Dark-eyed Juncos (Junco hyemalis thurberi). Behav Ecol Sociobiol, 62: 109–117
Fiala KL. 1981. Sex ratio constancy in the red-winged blackbird. Evolution, 35: 898–910
Fisher R. 1930. The Genetical Theory of Natural Selection. Oxford University Press, London
Frank SA. 1990. Sex allocation theory for birds and mammals. Annu Rev Ecol Syst, 21: 13–56
Fridolfsson AK, Ellegren H. 1999. A simple and universal method for molecular sexing of non-ratite birds. J Avian Biol, 30: 116–121
Geng R, Zhang XJ, Ou W, Sun HM, Lei FM, Gao W, Wang HT. 2009. Diet and prey consumption of breeding Common Kestrel (Falco tinnunculus) in Northeast China. Prog Nat Sci, 19: 1501–1507
Griffith SC, Örnborg J, Russell AF, Andersson S, Sheldon BC. 2003. Correlations between ultraviolet coloration, overwinter survival and offspring sex ratio in the Blue Tit. J Evol Biol, 16: 1045–1054
Griggio M, Hamerstrom F, Rosenfield R N, Tavecchia G. 2002. Seasonal variation in sex ratio of fledgling American Kestrels: A long term study. Wilson Bull, 114: 474–478
Hamilton WD. 1967. Extraordinary sex ratios. Science, 156: 477–488
Hardy ICW. 2002. Sex Ratios: Concepts and Methods. Cambridge University Press, Cambridge
Heinsohn R, Legge S, Barry S. 1997. Extreme bias in sex allocation in Eclectus parrots. Proc R Soc Lond B, 264: 1325–1329
Howe HF. 1977. Sex ratio adjustment in the Common Grackle. Science, 198: 744–745
Kalmbach E, Nager RG, Griffiths R, Furenss RW. 2001. Increased reproductive effort results in male-biased offspring sex ratio: an experimental study in a species with reversed sexual size dimorphism. Proc R Soc B, 268: 2175–2179
Kilner R. 1998. Primary and secondary sex ratio manipulation by Zebra Finches. Anim Behav, 56: 155–164
Kölliker M, Heeb P, Werner I, Mateman AC, Lessells CM, Richner H. 1999. Offspring sex ratio is related to male body size in the great tit (Parus major). Behav Ecol, 10: 68–72
Komdeur J, Daan S, Tinbergen JM, Mateman AC. 1997. Extreme adaptive modification in sex ratio of the Seychelles warbler's eggs. Nature, 385: 522–526
Komdeur J, Magrath MJL, Krackow S. 2002. Pre-ovulation control of hatchling sex ratio in the Seychelles Warbler. Proc R Soc B, 269: 1067–1072
Korpimäki E, May CA, Parkin DT, Wetton JH, Wiehn J. 2000. Environmental- and parental condition-related variation in sex ratio of kestrel broods. J Avian Biol, 31: 128–134
Korsten P, Lessells CM, Mateman AC, van der Velde M, Komdeur J. 2006. Primary sex ratio adjustment to experimentally reduced male UV attractiveness in blue tits. Behav Ecol, 17: 539–546
Krackow S. 1999. Avian sex ratio distortions: the myth of maternal control. Proc Int Ornithol Congr, 22: 425–433
Krijgsveld KL, Daan S, Dijkstra C, Visser GH. 1998. Energy requirements for growth in relation to sexual size dimorphism in Marsh Harrier (Circus aeruginosus) nestlings. Physiol Biochem Zool, 71: 693–702
Laaksonen T, Fargallo JA, Korpimäki E, Lyytinen S, Valkama J, Pöyri V. 2004. Year- and sex-dependent effects of experimental brood sex ratio manipulation on fledging condition of Eurasian kestrels. J Anim Ecol, 73: 342–352
Ležalová R, Tkadlec E, Obornák M, Šimek J, Honza M. 2005. Should males come first? The relationship between offspring hatching order and sex in the Black-headed Gull (Larus ridibundus). J Avian Biol, 36: 478–483
Legge S, Heinsohn R, Double MC, Griffiths R, Cockburn A. 2001. Complex sex allocation in the Laughing Kookaburra. Behav Ecol, 12: 524–533
Martínez-Padilla J, Fargallo JA. 2007. Food supply during prelaying period modifies the sex-dependent investment in eggs of Eurasian Kestrels. Behav Ecol Sociobiol, 61: 1735–1742
Martins TLF. 2004. Sex-specific growth rates in zebra finch nestlings: a possible mechanism for sex ratio adjustment. Behav Ecol, 15: 174–180
McDonald PG, Olsen PD, Cockburn A. 2005. Sex allocation and nestling survival in a dimorphic raptor: does size matter? Behav Ecol, 16: 922–930
Nager RG, Monagham P, Griffiths R, Houston DC, Dawson R. 1999. Experimental demonstration that offspring sex ratio varies with maternal condition. Proc Natl Acad Sci USA, 96: 570–573
Newton I, Marquiss M. 1979. Sex ratio among nestlings of the European sparrowhawk. Am Nat, 113: 309–315
Oddie KR, Reim C. 2002. Egg sex ratio and paternal traits: using within-individual comparisons. Behav Ecol, 13: 503–510
Olsent PD, Cockburn A. 1991. Female-biased sex allocation in Peregrine Falcons and other raptors. Behav Ecol Sociobiol, 28: 417–428
Pike TW, Petrie M. 2003. Potential mechanisms of avian sex manipulation. Biol Rev, 78: 553–574
Pike TW, Petrie M. 2005. Maternal body condition and plasma hormones affect offspring sex ratio in peafowl. Anim Behav, 70: 745–751
Pike TW, Petrie M. 2006. Experimental evidence that corticosterone affects offspring sex ratios in quail. Proc R Soc B, 273: 1093–1098
Riedstra B, Dijkstra C, Daan S. 1998. Daily energy expenditure of male and female marsh harrier nestlings. Auk, 115: 635–641
Rutkowska J, Badyaev AV. 2008. Meiotic drive and sex determination: molecular and cytological mechanisms of sex ratio adjustment in birds. Philos T R Soc B, 363: 1675–1686
Rutkowska J, Cichoń M. 2006. Maternal testosterone affects the primary sex ratio and offspring survival in Zebra Finches. Anim Behav, 71: 1283–1288
Rutstein AN, Slater PJB, Graves JA. 2004. Diet quality and resource allocation in the zebra finch. Proc R Soc Lond B, 271(Suppl. ): 286–289
Sheldon BC. 1998. Recent studies of avian sex ratios. Heredity, 80: 397–402
Slagsvold T, Sandvik J, Rofstad G, Husby M. 1984. On the adaptive value of intraclutch egg-size variation in birds. Auk, 101: 685–697
Tella JL, Donazar JA, Negro JJ, Hiraldo F. 1996. Seasonal and interannual variations in the sex-ratio of Lesser Kestrel (Falco naumanni) broods. Ibis, 138: 342–345
Torres R, Drummond H. 1997. Female-biased mortality in nestlings of a bird with size dimorphism. J Anim Ecol, 66: 859–865
Trivers RL, Willard DE. 1973. Natural selection of parental ability to vary the sex ratio of offspring. Science, 179: 90–92
Village A. 1990. The Kestrel. Poyser, London
Wegge P. 1980. Distorted sex ratio among small broods in a declining Capercaillie population. Ornis Scand, 11: 106–109
West SA, Sheldon BC. 2002. Constraints in the evolution of sex ratio adjustment. Science, 295: 1685–1688
Xiang GQ, Gao W, Feng HL. 1991. The study on breeding ecology of European Magpie (Pica pica). In: Gao W (ed) Chinese Bird Study. Chinese Science Press, Beijing, pp 102–106 (in Chinese)
Zhou T, Wang HT, Liu Y, Lei FM, Gao W. 2009. Patterns of magpie nest utilization by a nesting raptor community in a secondary forest. Prog Nat Sci, 19: 1253–1259
Table
1.
LMM analyses of the effects of clutch size and laying order on the sex of the eggs in Eurasian Kestrels
Source
NDF
DDF
FNDF, DDF
p
Clutch size
3
8
1.570
0.271
Laying order
1
8
0.004
0.951
Egg mass
1
8
1.796
0.217
Clutch size × Laying order
1
8
0.000
0.983
Covariance parameter estimates:
Estimate
SE
Z
p
Laying date
0.262
0.130
2.000
0.046
The model accounts for random variation between laying date. NDF represents the degrees of freedom of the numerator and DDF that of the denominator. FNDF, DDF stands for Fisher's test statistics, p represents the significance level, "estimate" represents the estimated covariance parameters, SE stands for standard error, Z represents Z statistic value of the Wald test.
Table
2.
LMM analyses of the effects of clutch size, laying order and hatching order on the sex of the fledgings in Eurasian Kestrel
Source
NDF
DDF
FNDF, DDF
p
Clutch size
3
11
5.297
0.017
Laying order
1
11
0.851
0.376
Hatching order
1
11
0.128
0.728
Egg mass
1
11
8.099
0.016
Clutch size × Laying order
1
11
6.447
0.028
Clutch size × Hatching order
1
11
5.508
0.039
Covariance parameter estimates:
Estimate
SE
Z
p
Laying date
0.194
0.008
2.345
0.019
The model accounts for random variation between laying date. NDF is the degree of freedom of the numerator and DDF of the denominator, FNDF, DDF is Fisher's test statistic, p represents significance level, "estimate" represents the estimated covariance parameters, SE stands for standard error, and Z represents the Z statistic value of the Wald test.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds