Nóra Ágh, Henriett Anna Dalvári, Krisztián Szabó, Ivett Pipoly, András Liker. 2024: Hard life for sons in the nest? Sex-dependent offspring mortality in Great Tits in urban and forest areas. Avian Research, 15(1): 100169. DOI: 10.1016/j.avrs.2024.100169
Citation:
Nóra Ágh, Henriett Anna Dalvári, Krisztián Szabó, Ivett Pipoly, András Liker. 2024: Hard life for sons in the nest? Sex-dependent offspring mortality in Great Tits in urban and forest areas. Avian Research, 15(1): 100169. DOI: 10.1016/j.avrs.2024.100169
Nóra Ágh, Henriett Anna Dalvári, Krisztián Szabó, Ivett Pipoly, András Liker. 2024: Hard life for sons in the nest? Sex-dependent offspring mortality in Great Tits in urban and forest areas. Avian Research, 15(1): 100169. DOI: 10.1016/j.avrs.2024.100169
Citation:
Nóra Ágh, Henriett Anna Dalvári, Krisztián Szabó, Ivett Pipoly, András Liker. 2024: Hard life for sons in the nest? Sex-dependent offspring mortality in Great Tits in urban and forest areas. Avian Research, 15(1): 100169. DOI: 10.1016/j.avrs.2024.100169
HUN-REN-PE Evolutionary Ecology Research Group, University of Pannonia, H-8200, Veszprém, Pf. 1158., Hungary. E-mail address: czikkelyne.agh.nora@gmail.com (N. Ágh)
1 Nóra Ágh and Henriett Anna Dalvári are joint first authors.
Sex-biased mortality can occur in birds during development, for example due to sexual differences in energy requirement and/or environmental sensitivity, or the effects of sex hormones or sex differences in the expression of mutations linked to sex chromosomes. The extent of sex-bias in mortality may also be related to environmental conditions that influence offspring development and survival. Urban areas often provide poorer conditions for nestling development resulting in higher offspring mortality compared to natural areas, which may accelerate sex differences in offspring mortality in cities. To test this hypothesis, we examined the sex ratio of dead offspring in Great Tits (Parus major), using 427 samples of unhatched eggs and dead nestlings collected in two urban and two forest sites between 2013 and 2019. The ratio of males in the whole sample of dead offspring (56.9%) was significantly higher than expected by an 1:1 ratio, and the strongest sex biases were detected in urban areas (57.6% males) and in young nestlings (< 14 days old, 59.0% males). However, the sex ratios of dead offspring did not differ significantly among study sites and between offspring developmental stages. 29.3% of unhatched eggs contained a visible embryo, and the proportion of embryo-containing unhatched eggs did not differ significantly between urban and forest study sites. These results suggest male-biased offspring mortality in Great Tits, and highlight the need of large datasets to detect subtle differences between habitats and developmental stages.
Offspring sex ratios may differ from 1:1 in birds, and the key factors affecting the magnitude and direction of the deviation are often habitat-, species- and population-specific (West and Sheldon, 2002). Sex-specific offspring mortality, the differential mortality rate between male and female embryos and nestlings during development, is one of the crucial elements that could lead to skewed offspring sex ratios. Survival differences between the sexes during growth may also have significant impacts on the adult sex ratio, which can have knock-on effects on population demography and viability (reviewed in Donald, 2007; Székely et al., 2014; Schacht et al., 2022).
Several theories have been proposed to explain sex-specific offspring mortality. First, when sexual size dimorphism occurs, the larger sex may have a faster rate of development and a higher energetic need (e.g. due to higher daily energy expenditure; Santoro et al., 2015). Therefore, both the rearing energy cost and environmental sensitivity of the larger offspring may be higher (costly sex hypothesis; Myers, 1978). Thus, during unfavourable conditions (e.g. food shortage, low-quality parents), the larger sex can have higher mortality (Clutton-Brock et al., 1985; Kalmbach et al., 2005; Benito and González-Solís, 2007). Alternatively, the larger offspring can have better competitive ability for food than its smaller brood-mates, which may results in increased mortality in the smaller sex, especially during food shortage (Råberg et al., 2005; Nicolaus et al., 2009). Second, due to the immunosuppressant effect of the androgenic hormones, males may have less effective immune function compared to females and hence have greater vulnerability to pathogens or parasites (Folstad and Karter, 1992). Consequently, under unfavourable environmental conditions (e.g. high parasitic prevalence), males would have a lower survival rate due to their weaker immune system compared to females (Swenson, 1986; Folstad and Karter, 1992 but see Bize et al., 2005). Third, mutations linked to sex chromosomes may also be a reason for sex-specific mortality. According to the unguarded sex chromosome hypothesis (Haldane, 1922), the heterogametic sex (females in birds) may suffer higher mortality because harmful mutations in genes that are present in a single copy on the two different sex chromosomes can express easier than in the homogametic sex that have two gene copies (Pipoly et al., 2015; Xirocostas et al., 2020).
A comparison between natural and urbanized areas offers an excellent opportunity to test how habitat quality affects sex differences in offspring mortality. Compared to natural/semi natural habitats, cities and suburban areas have a higher presence of humans (Sol et al., 2013), altered predator pressure (Vincze et al., 2017), changed quantity and quality of food (Seress et al., 2018; Sinkovics et al., 2021) and higher degree of pollution (Kekkonen, 2017). The effects of urbanization on the reproductive success of birds are not entirely consistent, but it usually involves an earlier start of breeding, a smaller clutch size, reduced hatching rates, lower nestling weight, and lower offspring survival (e.g. Chamberlain et al., 2009; Bailly et al., 2016; Seress et al., 2018, 2020). In general, urban areas may be a harsher environment for developing offspring than the more natural habitats for many bird populations, which can accelerate the sex differences in offspring mortality.
Despite the large number of studies focusing on offspring mortality and its relationship with anthropogenic effects (e.g. Rejt et al., 2005; Bailly et al., 2016; Seress et al., 2020), to our knowledge only a single study investigated the sex differences in mortality of urban birds, reporting male-biased nestling mortality in Eurasian Kestrels (Falco tinnunculus) breeding in urban areas (Sumasgutner et al., 2014). Experimental studies that compared pre-fledging survival between the sexes in contrasting environments produced contradictory results: either the larger (Nager et al., 2000; Tschirren et al., 2003) or the smaller sex (Bize et al., 2005; Rowland et al., 2007) suffered higher mortality in poor rearing conditions. In addition, the vulnerability of the offspring to the external and internal factors may also vary across developmental stages (see for example Kato et al., 2017; Marques-Santos and Dingemanse, 2020). Thus, our understanding of the effects of unfavourable conditions, particularly in the context of urbanization, on sex-specific offspring mortality is incomplete.
Our aim is to examine whether the sex ratio of dead offspring (SRDO, i.e. proportion of males in all dead offspring) is male- or female-biased in Great Tit (Parus major). This species is frequently used as a model system for long-term, individual-based studies investigating sex allocation, reproductive success, and adaptation to urban environments (e.g. Clutton-Brock and Sheldon, 2010; Bailly et al., 2016; Seress et al., 2018). Although several studies examined how urbanization, metal pollution and other anthropogenic disturbances correlate with fledglings' sex ratio or mortality in Great Tits (e.g. Eeva et al., 2012; Bouvier et al., 2016; Corsini et al., 2021), these studies did not determine primary sex ratio or the sex of the dead offspring to infer sex differences in offspring mortality. Both studies that investigated sex-specific offspring mortality in this species were conducted in natural forest habitats (Oddie, 2000; Cichoń et al., 2005).
Here we study sex-specific offspring mortality in two urban and two forest areas. We have already shown that the urban sites have lower availability of high-quality nestling food (caterpillars) and reduced reproductive success by great tits (Seress et al., 2018; Sinkovics et al., 2021, 2023). Neither the primary sex ratio (49%) nor the fledgling sex ratio (52%) differ significantly from a balanced sex ratio in these populations, or differ between urban and forest sites (Ágh et al., 2020). However, we found remarkably high variance in primary and fledgling sex ratios among broods that may be explained by early sex-specific mortalities. We have three main goals in this study: (1) We investigate if there is sex-specific offspring mortality in our study populations. The hypotheses discussed above could predict either higher male or higher female mortality. (2) We investigate whether the sex ratio of dead offspring differs between study sites and between urban and forest areas, or between different stages of development i.e. embryonic, young (<14 days) and old nestling stages (≥14 days). Because urbanization has a negative impact on offspring development and survival (see above), we predict a higher ratio of the more sensitive sex in dead offspring in urban sites. (3) In addition to testing sex differences in early mortality, we also use our sample of unhatched eggs to estimate their fertilization rate and compare this between urban and forest areas.
2.
Material and methods
2.1
Fieldwork and sample collection
We studied Great Tit populations in urban and forest areas in Hungary. The two forest sites were located in deciduous woodlands: a beech forest near Szentgál (47°06′39.75″ N, 17°41′17.94″ E) and an oak forest in Vilma-puszta (47°05′06.7″ N, 17°51′51.4″ E). The two urban sites were located mostly in public parks and university campuses in the cities of Veszprém (47°05′17.29″ N, 17°54′29.66″ E) and Balatonfüred (46°57′30.82″ N, 17°53′34.47″ E) (Seress et al., 2017, 2018). In each of the four sites, there were 80–100 numbered artificial nest boxes. For the purposes of the present study, we monitored the nest boxes and collected data and samples for seven consecutive years, from 2013 to 2019. In each year, the nest monitoring started in March and continued until the last brood fledged in early July. We monitored nest boxes at least twice a week and recorded the date of laying of the first egg and date of hatching, and the number of eggs and nestlings present. We collected unhatched eggs from the fifth day after the first offspring hatched in the brood to avoid collecting late hatching eggs. When we found dead nestlings in the nests during nestling development, we collected tissue samples from them by clipping a toe or picking a few feathers. We ringed nestlings alive 14–16 days after hatching, and from most them took small blood samples from their brachial vein. We stored the blood samples in either AVL or Queens's lysis solution or in 96% ethanol, and the other tissues and unhatched eggs in 96% ethanol at −20 ℃ until further analysis (Ágh et al., 2020). We revisited nests after the broods fledged, usually 5–7 days (maximum 14 days) after ringing (i.e. 19–30 days after hatching). At this last nest checking, we recorded the number of dead nestlings and identified them by their ring numbers.
According to the above sample collection protocol, we divided the samples into three developmental stages: embryonic (unhatched eggs), nestlings died before ringing (toe and feather samples, "young nestlings" henceforward) and nestlings died after ringing (blood samples, "old nestlings" henceforward). Although the range of ringing age was 14–16 days, for simplicity we refer to the age of young and old nestling as < 14 days and ≥14 days, respectively, because most nestlings were ringed at that age. Altogether, we collected 634 samples of unhatched eggs and dead nestlings from 331 broods (see details in Appendix Table S1).
2.2
Laboratory methods
Sex determination was conducted by using the CHD1-Z and CHD1-W genes of the avian sex chromosomes which differ in their intronic gene sequence length hence are suitable to differentiate between the sexes (Ellegren, 1996). We extracted DNA by using silica membrane isolation kits like GeneAid Genomic DNA Mini Kit (Tissue) and Omega E.Z.N.A. Tissue DNA Kit, following the manufacturer's instructions. We examined the egg contents after cracking them in a Petri-dish with sterilized tools. We further analysed only those samples that had a visible embryo by taking a small tissue sample, while eggs without any sign of a developing embryo were categorized as apparently infertile. We call these eggs "unfertilized" henceforward. We successfully sexed 427 offspring (85 embryos, 236 young and 106 old nestlings). Two blood samples and one tissue sample had insufficient DNA for sex determination, and we did not find embryos in 204 of the 289 unhatched eggs.
During the polymerase chain reaction (PCR), the final volume of the sample mixture was 17 μL, which contained 0.66 μL dNTP Mix (2 mM, Fermentas), 0.66 μL MgCl2 (25 mM), 0.066 μL DreamTaqTM (Fermentas) polymerase enzyme, 1.7 μL 10x DreamTaqTM solution (Fermentas), 10.5 μL MQ water, 2 μL primer mix (5 pmol/μL) and 2 μL solution containing the dissolved DNA. We used CHD-i16 (Suh et al., 2011) and p2p8 primers (Griffiths et al., 1998) for molecular sexing as both primers multiply different sized intronic sequences in the CHD1 gene of the W- and the Z-chromosomes. CHD1-i16 primer was used for analysing blood, toe and feather samples while p2p8 primer was used for egg samples, because this primer pair is suitable for fragmented DNA as well (Jensen et al., 2003). Several samples (in 24 eggs, 12 blood and 14 toe samples randomly chosen from every examined year) were tested with both primers for double-checking, and these repeated sexing provided consistent results in all cases. We used "touchdown" function during the PCR with the CHD1-i16 primer for better specificity (Ágh et al., 2018). For the gel electrophoresis, 2% agarose gel (Lonza SeaKem® LE) was used with 20, 000-fold diluted dye (ECO Safe Nucleic Acid Staining Solution; Pacific Image Electronics, Inc.). The running time was set to be 30 min for the i16 primer and 45 min for the p2p8 primer with a current of 100 V. At the end, the amplified CHD1 fragments were visualized with UV light thus we could determine the sex of the samples by the number of CHD1 band(s) (Ágh et al., 2018).
2.3
Statistical analyses
All statistical analyses were conducted using R version 3.6.1 (R Development Core Team, 2019). We used the "glmer" function in package "lme4" (Bates et al., 2014) to build the models. The level of significance was tested using Type Ⅱ analysis of variance (ANOVA) tables with Wald chi-square tests from package "car" (Fox and Weisberg, 2019).
We built two sets of statistical models to investigate the sex ratio of dead offspring (SRDO, expressed as % of males). First, we tested whether the SRDO differs from a balanced sex ratio (i.e. from the 1:1 male: female ratio) in the full sample (embryos + nestlings) using a generalized linear mixed model (GLMM) with binomial error distribution and 'logit' link function. We used the sex of the dead embryo or nestling as binary response variable (0 = female, 1 = male) with brood ID within pair ID and year as random factors to control for the non-independence of nestlings died in the same brood (Wilson and Hardy, 2002). This model did not contain any fixed effect. It tested whether the odds ratio (OR, the proportional difference of the odds of a dead offspring being male) differed from 1. OR = 1 means an equal probability for a dead offspring being a male or a female, which is equivalent with a 1:1 sex ratio in the sample of dead offspring. For easier interpretation of the results, we calculated the estimated proportion of males from the model's OR and provided this measure as an estimate of SRDO. We used OR = 1 as reference value in the model because in a previous work we showed that the primary sex ratio (at conception) did not differ statistically from the 1:1 sex ratio in all of our study populations during years partially overlapping with this study (using data from completely sexed broods; Ágh et al., 2020). In addition, we repeated the above GLMM analysis using subsamples of (1) the four different study sites, (2) the two types of sites (urban and forest), and (3) the three developmental stages of nestlings. We ran this latter set of analyses to provide separate SRDO estimates for these specific subsamples.
Second, we tested whether the SRDO differed between sites and offspring developmental stages using a GLMM model with binomial error distribution and 'logit' link function. The sex of the offspring as a binary variable (0 = female, 1 = male) was the response, and fixed effects were the study sites (four factor levels), and the offspring developmental stages (three factor levels) with brood ID within pair ID and year as the random factors. Model estimates were back-transformed from the log-scale to provide the odds ratio (i.e. site difference in the odds of a dead nestling being male) with 95% confidence interval (CI).
Finally, to compare the fertilization rate of the unhatched eggs among the sites we used a GLMM model where the presence of embryo in the egg was the binary response variable (0 = absent, 1 = present), study site was a fixed effect and the brood ID within pair ID and year were included as a random factor.
3.
Results
In our study populations the mean ± SD number of the unhatched eggs per nest was 0.60 ± 1.00 in the urban and 0.59 ± 1.1 in forest areas, whereas the mean number of dead nestlings per brood was 2.84 ± 2.12 in urban and 1.69 ± 2.13 in forest sites (during the period from 2013 to 2019).
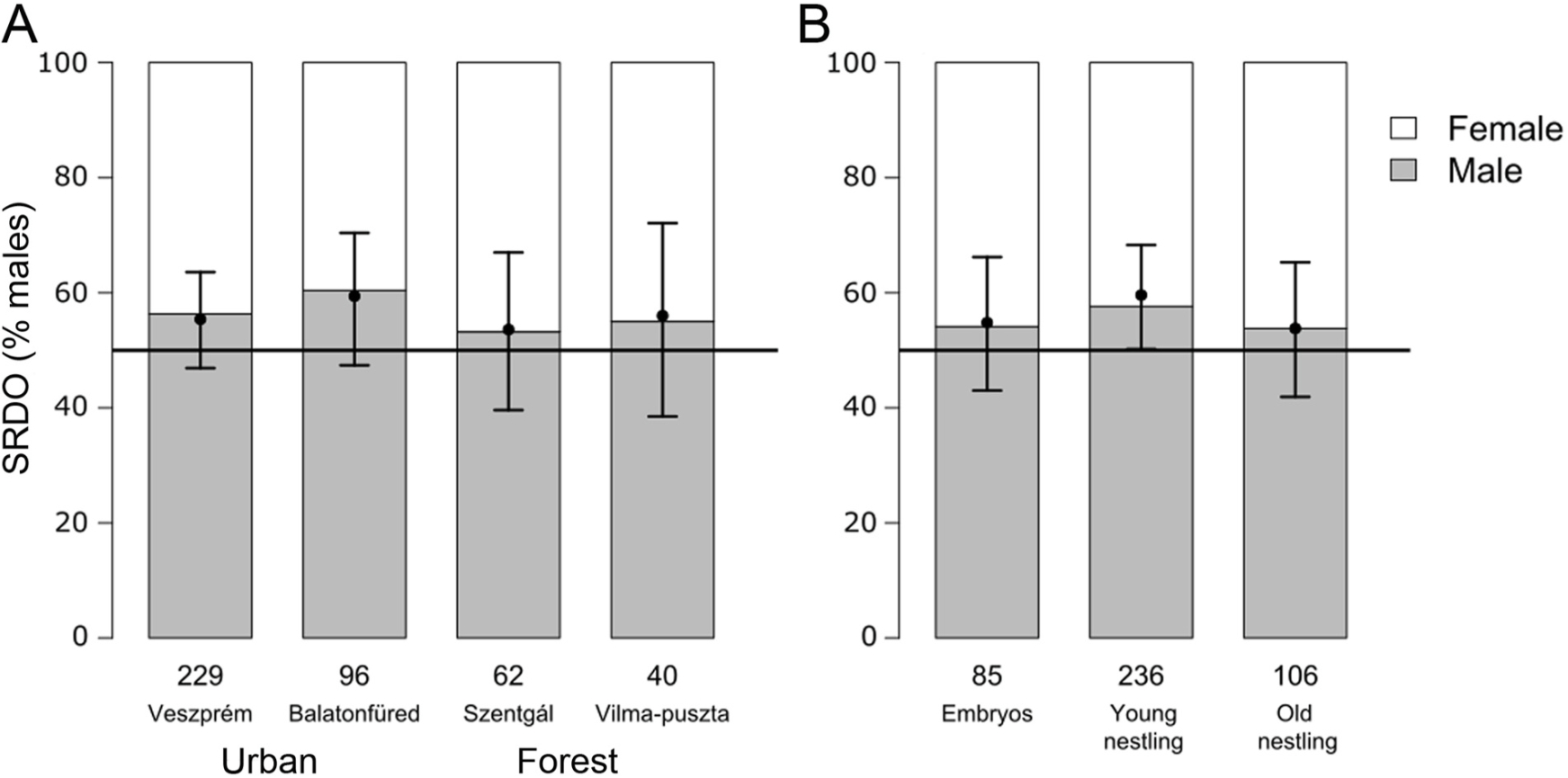
In the full sample, the estimated SRDO was male-biased (56.9% males) and significantly differed from a balanced sex ratio (all sites and offspring age stages combined: OR [95%CI] = 1.31 [1.06–1.63], n = 427; Table 1A). When we tested the sites separately, we observed significant male-bias from the 1:1 sex ratio in the city of Veszprém and a marginally non-significant male-bias in the other urban site Balatonfüred (Table 1B; Fig. 1A). Furthermore, we found a significantly male-biased SRDO in the urban area (Veszprém and Balatonfüred combined), whereas no significant bias was detected in the forests (Table 1B, C). When we analysed developmental stages separately, we found a significantly male-biased SRDO in young nestlings, but the SRDO of embryos in unhatched eggs and old nestlings did not differ statistically from the 1:1 sex ratio (Table 1D; Fig. 1B). However, the model that compared the SRDO between populations and developmental stages (i.e. combined all samples and contained both site and developmental stages as predictors) did not indicate statistically significant differences either among the sites (ANOVA, χ2 = 0.502, df = 3, p = 0.918) or between the offspring age stages (χ2 = 0.969, df = 2, p = 0.616).
Table
1.
Sex ratio of dead offspring (SRDO) estimated by generalized linear mixed models (GLMM) for different sites and developmental stages. N indicates the sample size in each category (dead offspring with known sex), OR is the odds ratio (i.e. the proportional difference of the odds of a dead offspring being male compared to being female in the studied group), back-transformed from log scale. SRDO indicates the sex ratio estimates for dead offspring (expressed as the percentages of males in all dead individuals in the category), calculated from the OR. 95% confidence intervals (CI) for OR were calculated from the statistical models that did not contain any fixed effect (see Methods in the main text for explanation). Bold values highlight estimates where the CIs did not include 1, meaning that the OR is statistically different from 1, with OR > 1 indicating more dead males than dead females.
Groupa
N
OR
95% CI
SRDO
A) Full sample
All samples (embryos + nestlings)
427
1.31
1.06–1.63
56.88%
B) Site groups
Veszprém (urban)
229
1.29
1.01–1.68
56.33%
Balatonfüred (urban)
96
1.55
0.86–2.79
60.83%
Szentgál (forest)
62
1.20
0.63–2.30
54.45%
Vilma-puszta (forest)
40
1.23
0.66–2.28
55.00%
Urban and forest areas
All urban samples
325
1.36
1.08–1.73
57.63%
All forest samples
102
1.21
0.77–1.91
54.81%
D) Offspring age stages
Embryos
85
1.18
0.76–1.84
54.23%
Young nestlings (before 14 days)
236
1.44
1.09–1.91
59.01%
Old nestlings (after 14–16 days)
106
1.17
0.78–1.75
53.83%
All nestlings (young + old)
342
1.35
1.07–1.71
57.46%
a All estimates (percentages, OR, and CI) were obtained from a separate model for each group of samples. Thus, for example, the estimates for the urban area are not the simple average of the values for the two urban sites.
Figure
1.
Sex ratio dead offspring (SRDO) (A) at each study site and (B) in each offspring developmental stage. Shading shows the raw data, whereas dots and whiskers show point estimates and their 95% confidence interval, respectively, from the GLMM model that contained the sites and the age stage categories as explanatory variables. Numbers below the bars refer to the number of offspring in each group. None of the differences between the categories was statistically significant (see the main text for the details of the statistical results).
The proportion of eggs that failed to hatch was similar between the sites (Table S2). 30–40% of the unhatched eggs contained identifiable embryos in each study site (Fig. S1) and this proportion did not differ significantly between the sites (χ2 = 3.992, df = 3, p = 0.262; Appendix Fig. S1, Table S2).
4.
Discussion
4.1
Male-biased offspring mortality
We found an overall male-bias in offspring mortality in Great Tits, with an estimated SRDO of 56.9%. Since dead offspring were more often male, and the primary offspring sex ratio of the population was likely balanced (based on data from 2012 to 2014; Ágh et al., 2020), our data imply a male biased offspring mortality. We consider our finding robust, because our analysis was based on a relatively large sample size, covered seven breeding seasons, and the offspring mortality tended to be male-biased in every subset (i.e. site or developmental stages, see Table 1) of the dataset. Our findings did not support the unguarded sex chromosomes hypothesis, which would predict higher mortality for heterogametic females (Haldane, 1922). Instead, it is consistent with the costly sex hypothesis, which states that larger male offspring may be more sensitive to environmental stressors during development (e.g. Benito and González-Solís, 2007; Rosivall et al., 2010; Bowers et al., 2015). The results are also in line with the immune suppression hypothesis predicting that males may have a lower survival rate compared to females because of the immunosuppressant effect of testosterone (e.g. Folstad and Karter, 1992; Tobler et al., 2010, but see Von Engelhardt et al., 2006).
The costly sex hypothesis could be relevant both before and after hatching, because in some birds sexual size dimorphism emerges already in the embryonic stage, for example eggs containing a male embryo could be larger (e.g. Martyka et al., 2010). The sexual size dimorphism is detectable in the young nestling stage in Great Tits (Oddie, 2000; Stauss et al., 2005; Nicolaus et al., 2009). For example, in our study populations males are heavier by 3.6% and have longer tarsus by 2.5% than female nestlings by the age of ringing (14–16 days after hatching; Ágh et al., 2020). So, the larger size and the presumably greater energy demand during development may make males more sensitive to adverse environmental effects than females (Swenson, 1986; Jones et al., 2009; Rosivall et al., 2010; Bouvier et al., 2016). Unfavourable conditions may be particularly common in urban areas, for example due to the limited availability of high-quality nestling food (e.g. Seress et al., 2018; Sinkovics et al., 2021, 2023), and perhaps the greater competition for food between breeding birds due to the higher population density that characterize some urban bird populations (Møller et al., 2012). On the other hand, some studies did not find an influence of adverse conditions on sex-specific survival in Great Tits, for instance by increased pollution levels (in 15 days old nestlings; Eeva et al., 2012), by increased competition among siblings (Nicolaus et al., 2009) or by handicapping the parents (Kubacka and Cichoń, 2020). These latter results suggest that male-biased offspring mortality may not be fully explained by higher sensitivity of male offspring to adverse environmental conditions after hatching. Both our and Cichoń et al. (2005) observations suggest that offspring mortality could be slightly (although not significantly in our study) male-biased already in the embryo stage. This can also be explained by the higher sensitivity of male embryos to harsh conditions, which may occur during incubation for example due to the starvation of parents (which may reduce the effectiveness of incubation) or unfavourable weather.
The higher incidence of males than females among dead offspring predicts an offspring sex ratio biased towards females. Although we do not know the sex ratio of alive offspring in broods where mortality occurred, we found that the fledging sex ratio did not differ significantly from 1:1 in the same populations investigated during a period partially overlapping with the current study (2012–2014; Ágh et al., 2020) which is in contrast with the prediction of female bias in alive offspring. One explanation for this discrepancy could be that the primary sex ratio (i.e. that of the fertilized eggs) is male-biased and this is compensated by the male-biased offspring mortality. However, in our earlier study, the primary sex ratio (estimated from broods without any egg or nestling mortality) was 48.7% and did not differ significantly from the 1:1 ratio (95%CI = [0.371; 0.602], n = 72 broods; Ágh et al., 2020), thus does not support the above hypothesis. An alternative explanation may be that the frequency of offspring mortality is relatively small to affect brood sex ratios in a statistically detectable way. For example, in our study populations, the mean number of the unhatched eggs per nest is less than one in both the urban and forest areas, whereas the mean number of dead nestlings per brood was nearly 2–3 (see Results). These relatively low offspring mortality (compared to the mean clutch size of 8–11 in urban and 11–12 in forest sites in the same study populations; Seress et al., 2018) combined with its small sex difference may results in an even smaller sex bias in the brood sex ratio that may not be detectable when analysed using a moderate sample size of offspring (e.g. 1152 alive nestlings in 126 broods in our earlier study, see Ágh et al., 2020).
4.2
Differences between sites and offspring developmental stages
Based on our long-term dataset on the breeding success of Great Tits, nestling mortality was higher at the urban than at the forest sites (see above, and also Seress et al., 2018, 2020, 2021), similarly to results reported in other studies (e.g. Bailly et al., 2016). Thus, we expected that the more sensitive male offspring die with higher frequency relative to females in the less favourable urban areas. Although some of our results were in line with this prediction, others did not provide support for it. First, we found slightly more dead males in all study sites (SRDO between 54.45% and 60.83%, Table 1B), but that was significant only in urban areas (i.e. in Veszprém and when data of two urban sites were combined), which may be explained by a more pronounced male-biased mortality in cities. However, we also have larger sample sizes in the analyses for urban sites that may help statistically detect biased mortality. In contrast, we did not detect significant differences in SRDO between urban and forest areas when we compared sites in a single model. The most probable reasons for this discrepancy between the results can include (1) the consistent tendency for a higher male than female mortality in all sites with only small difference between them (1–5% higher ratio of males in urban sites, see Table 1C), and (2) the large confidence intervals of the estimates which limit the power of the statistical inference. Thus, it seems that offspring mortality in our Great Tit populations is male-biased in both urban and forest areas, and further research involving larger sample sizes and/or more explanatory variables or another habitat division (e.g. an urban gradient approach) would be required to confirm the slightly higher relative mortality of male offspring in urban areas suggested by some of the results.
We also investigated whether sex-specific mortality occurs at different age stages during development. We found slightly male-biased SRDOs in both the embryos and nestlings, with the largest bias occurring in the young nestling stage. Some other studies showed that Great Tit nestlings may be more sensitive in the earlier developmental stage than later on (Marques-Santos and Dingemanse, 2020). The most critical time period for growing and for nestling survival is around the 4–8 days, because the thermoregulation start to develop in this phase and the nestlings also have an increasingly high food demands (Marques-Santos and Dingemanse, 2020). This time period overlaps with our young nestling stage category. So, if males are more sensitive than females during development, one would expect the highest male-bias in SRDO in this most sensitive period, and our result appear to support this idea. However, this was also the developmental stage with the largest sample size, which facilitates the detection of a significant difference. In addition, when we compared the developmental stages in a single model there was no significant differences between their SRDOs. Again, this model may fail to detect any difference because the tendency of sex ratio bias is similar in each developmental stages, and the wide confidence intervals associated with the estimates may make small differences undetectable.
Our study showed that most unhatched eggs contain no sign of embryos in all study sites (62.8%–77.6%, Appendix Table S2). Although there were no statistically significant differences between sites, the 10% higher presence of embryos in unhatched eggs in urban sites (Appendix Table S2) may indicate that the development of fertilized eggs may be stopped by internal or external factors more often in cities than in the forest, and thus the embryos fail to develop fully and hatch. In contrast, in forest populations fertilized eggs may have a higher chance of hatching after the start of embryonic development, resulting in a smaller proportion of eggs with embryos that failed to hatch. The possibility that urban environments affect the viability of eggs deserves for further research.
Finally, it is important to consider that our study has some limitations. First, it is quite difficult to collect all dead offspring from the studied broods, especially in the young nestling phase, due to the removal of dead nestlings by parents (e.g. Kirkpatrick et al., 2009; our personal observation). This may potentially introduce a sampling bias, although it seems unlikely that parents preferentially remove either the male or female dead nestlings from the nests. Second, our method based on the presence of visible embryos may underestimate the fertility rate in unhatched eggs. This can explain why we found a higher infertility rate than some other studies in Great Tits using different detection methods (e.g. 13.7% in Hemmings and Evans, 2020).
5.
Conclusion
Overall, our results show that male Great Tit offspring have a higher mortality rate than females, and this is largely independent from the degree of urbanization of the sites or the developmental stages of the offspring. Although the SRDO did not differ significantly between sites or between developmental stages, we found that urban sites and the young nestlings (died before ringing at 14–16 days old) exhibited especially high proportion of dead males, which is in line with some theoretical predictions and may be worth further testing. Although our findings showed a relatively small male bias in offspring mortality, this could have important consequences for reproduction and population demography in Great Tits. For example, sex-biased mortality of offspring may contribute to bias in the adult sex ratio, which in turn could affect sex allocation, mating system, and population persistence (Eberhart-Phillips et al., 2017). In the light of current habitat urbanisation, it is especially important to understand how the special ecological conditions in the cities influence sex-biased offspring mortality, its potential mechanisms, and the impacts of sex-biased mortality on the demography and reproductive adaptations of urban populations. Our study also highlights the need for and potential of sampling and molecular sexing dead offspring that can help understand more clearly the reasons of changing sex ratios from hatching to fledging.
Ethics statement
All applicable international, National and/or Institutional Guidelines for the care and use of animals were followed. Research was permitted by the Middle Transdanubian Inspectorate for Environmental Protection, Natural Protection and Water Management (Permission Number: 31, 559/2011). All procedures were in accordance with the guidelines for animal care outlined by ASAB/ABS and Hungarian laws.
CRediT authorship contribution statement
Nóra Ágh: Writing – review & editing, Writing – original draft, Formal analysis, Data curation, Conceptualization. Henriett Anna Dalvári: Writing – original draft, Methodology, Data curation. Krisztián Szabó: Methodology, Data curation. Ivett Pipoly: Writing – review & editing, Writing – original draft, Data curation, Conceptualization. András Liker: Writing – review & editing, Writing – original draft, Supervision, Data curation, Conceptualization.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We thank all former and current members of the HUN-REN-PE Evolutionary Ecology Research Group who contributed to the fieldwork and collected the samples for this study. We are grateful to the members of Molecular Ecology Research Group of the University of Veterinary Medicine Budapest who helped in the laboratory work.
This work was financially supported by the Hungarian National Research, Development and Innovation Office (NKFIH, grant K132490 to AL and grant PD142106 to IP), by the HUN-REN Hungarian Research Network (grant 16007 to AL), and by the Sustainable Development and Technologies National Programme of the Hungarian Academy of Sciences (NP2022-Ⅱ-6/2022).
Ágh, N., Kovács, S., Nemesházi, E., Szabó, K., 2018. Univerzális, ivarhatározáshoz használt CHD1 markerek alkalmazhatósága különbözö madárrendekben. Magy. Állatorvosok Lapjallatorvosok Lapja 140, 47-59.
Ágh, N., Pipoly, I., Szabó, K., Vincze, E., Bókony, V., Seress, G., et al., 2020. Does offspring sex ratio differ between urban and forest populations of great tits (Parus major)? Biol. Futur. 71, 99-108.
Bailly, J., Scheifler, R., Berthe, S., Clément-Demange, V.A., Leblond, M., Pasteur, B., et al., 2016. From eggs to fledging: negative impact of urban habitat on reproduction in two tit species. J. Ornithol. 157, 377-392.
Bates, D., Maechler, M., Bolker, B., Walker, S., 2014. Fitting linear mixed-effects models using lme4. J. Stat. Softw. arXiv, 1406.5823.
Benito, M.M., González-Solís, J., 2007. Sex ratio, sex-specific chick mortality and sexual size dimorphism in birds. J. Evol. Biol. 20, 1522-1530.
Bize, P., Roulin, A., Tella, J.L., Richner, H., 2005. Female-biased mortality in experimentally parasitized Alpine Swift Apus melba nestlings. Funct. Ecol. 19, 405-413.
Bouvier, J.C., Boivin, T., Charmantier, A., Lambrechts, M., Lavigne, C., 2016. More daughters in a less favourable world: breeding in intensively-managed orchards affects tertiary sex-ratio in the great tit. Basic Appl. Ecol. 17, 638-647.
Bowers, E.K., Thompson, C.F., Sakaluk, S.K., 2015. Persistent sex-by-environment effects on offspring fitness and sex-ratio adjustment in a wild bird population. J. Anim. Ecol. 84, 473-486.
Chamberlain, D., Cannon, A.R., Toms, M.P., Leech, D.I., Hatchwell, B.J., Gaston, K.J., 2009. Avian productivity in urban landscapes: a review and meta-analysis. Ibis 151, 1-18.
Cichoń, M., Sendecka, J., Gustafsson, L., 2005. Male-biased sex ratio among unhatched eggs in great tit Parus major, blue tit P. caeruleus and collard flycatcher Ficedula albicollis. J. Avian Biol. 36, 386-390.
Clutton-Brock, T., Sheldon, B.C., 2010. Individuals and populations: the role of long-term, individual-based studies of animals in ecology and evolutionary biology. Trends Ecol. Evol. 25, 562-573.
Clutton-Brock, T.H., Albon, S.D., Guinness, F.E., 1985. Parental investment and sex differences in juvenile mortality in birds and mammals. Nature 313, 131-133.
Corsini, M., Schöll, E.M., Di Lecce, I., Chatelain, M., Dubiec, A., Szulkin, M., 2021. Growing in the city: urban evolutionary ecology of avian growth rates. Evol. Appl. 14, 69-84.
Donald, P.F., 2007. Adult sex ratios in wild bird populations. Ibis 149, 671-692.
Eberhart-Phillips, L.J., Küpper, C., Miller, T.E., Cruz-López, M., Maher, K.H., dos Remedios, N., et al., 2017. Sex-specific early survival drives adult sex ratio bias in snowy plovers and impacts mating system and population growth. Proc. N. Aca. Sci., 114, E5474-E5481.
Eeva, T., Sillanpää, S., Lehikoinen, E., 2012. Metal pollution does not bias offspring sex ratio in great tit (Parus major). Environ. Sci. Pollut. Res. 19, 2870-2878.
Folstad, I., Karter, A.J., 1992. Parasites, bright males, and the immunocompetence handicap. Am. Nat. 139, 603-622.
Griffiths, R., Double, M.C., Orr, K., Dawson, R.J.G., 1998. A DNA test to sex most birds. Mol. Ecol. 7, 1071-1075.
Haldane, J.B.S., 1922. Sex ratio and unisexual sterility in hybrid animals. J. Genet. 12, 101-109.
Hemmings, N., Evans, S., 2020. Unhatched eggs represent the invisible fraction in two wild bird populations. Biol. Lett., 16, 20190763.
Jensen, H., Sæther, B.E., Ringsby, T.H., Tufto, J., Griffith, S.C., Ellegren, H., 2003. Sexual variation in heritability and genetic correlations of morphological traits in house sparrow (Passer domesticus). J. Evol. Biol. 16, 1296-1307.
Jones, K.S., Nakagawa, S., Sheldon, B.C., 2009. Environmental sensitivity in relation to size and sex in birds: meta-regression analysis. Am. Nat. 174, 122-133.
Kalmbach, E., Furness, R.W., Griffiths, R., 2005. Sex-biased environmental sensitivity: natural and experimental evidence from a bird species with larger females. Behav. Ecol. 16, 442-449.
Kato, T., Matsui, S., Terai, Y., Tanabe, H., Hashima, S., Kasahara, S., et al., 2017. Male-specific mortality biases secondary sex ratio in Eurasian tree sparrows Passer montanus. Ecol. Evol. 7, 10675-10682.
Kekkonen, J., 2017. Pollutants in urbanized areas: direct and indirect effects on bird populations. In: Murgui, E., Hedblom, M. (Eds.), Ecology and Conservation of Birds in Urban Environments. Springer International Publishing, Cham, pp. 227–250.
Kirkpatrick, C., Conway, C.J., Ali, M.H., 2009. Sanitation of entire broods of dead nestlings may bias cause-specific nest failure rates. Ibis. 151, 207-211.
Kubacka, J., Cichoń, M., 2020. An immune challenge of female great tits decreases offspring survival and has sex-specific effects on offspring body size. Acta Ethol. 23, 173-181.
Marques-Santos, F., Dingemanse, N.J., 2020. Weather effects on nestling survival of great tits vary according to the developmental stage. J. Avian Biol. 51 (jav), 02421.
Martyka, R., Rutkowska, J., Dybek-Karpiuk, A., Cichoń, M., Walasz, K., 2010. Sexual dimorphism of egg size in the European Blackbird Turdus merula. J. Ornithol. 151, 827-831.
Møller, A.P., Diaz, M., Flensted-Jensen, E., Grim, T., Ibáñez-Álamo, J.D., Jokimäki, J., et al., 2012. High urban population density of birds reflects their timing of urbanization. Oecologia, 170, 867-875.
Myers, J.H., 1978. Sex ratio adjustment under food stress: maximization of quality or numbers of offspring? Am. Nat. 112, 381-388.
Nager, R.G., Monaghan, P., Houston, D.C., Genovart, M., 2000. Parental condition, brood sex ratio and differential young survival: an experimental study in gulls (Larus fuscus). Behav. Ecol. Sociobiol. 48, 452-457.
Nicolaus, M., Michler, S.P.M., Ubels, R., van der Velde, M., Komdeur, J., Komdeur, J., et al., 2009. Sex-specific effects of altered competition on nestling growth and survival: an experimental manipulation of brood size and sex ratio. J. Anim. Ecol. 78, 414-426.
Oddie, K.R., 2000. Size matters: competition between male and female great tit offspring. J. Anim. Ecol. 69, 903-912.
Pipoly, I., Bókony, V., Kirkpatrick, M., Donald, P.F., Székely, T., Liker, A., 2015. The genetic sex-determination system predicts adult sex ratios in tetrapods. Nature 527, 91-94.
Råberg, L., Stjernman, M., Nilsson, J. Å., 2005. Sex and environmental sensitivity in blue tit nestlings. Oecologia 145, 496-503.
Rejt, L., Gryczynska-Siemaiatkowska, A., Rutkowski, R., Malewska, A., 2005. Does egg sex ratio in urban kestrels (Falco tinnunculus) differ from parity? Pol. J. Ecol. 53, 545-552.
Rosivall, B., Szöllősi, E., Hasselquist, D., Török, J., 2010. Males are sensitive — sex-dependent effect of rearing conditions on nestling growth. Behav. Ecol. Sociobiol. 64, 1555-1562.
Rowland, E., Love, O.P., Verspoor, J.J., Sheldon, L., Williams, T.D., 2007. Manipulating rearing conditions reveals developmental sensitivity in the smaller sex of a passerine bird, the European starling Sturnus vulgaris. J. Avian Biol. 38, 612-618.
Santoro, S., Green, A.J., Speakman, J.R., Figuerola, J., 2015. Facultative and non-facultative sex ratio adjustments in a dimorphic bird species. Oikos 124, 1215-1224.
Schacht, R., Beissinger, S.R., Wedekind, C., Jennions, M.D., Geffroy, B., Liker, A., et al., 2022. Adult sex ratios: causes of variation and implications for animal and human societies. Commun. Biol. 5, 1273.
Seress, G., Vincze, E., Pipoly, I., Hammer, T., Papp, S., Preiszner, B., et al., 2017. Effects of capture and video-recording on the behavior and breeding success of Great Tits in urban and forest habitats. J. Field Ornithol. 88, 299-312.
Seress, G., Hammer, T., Bókony, V., Vincze, E., Preiszner, B., Pipoly, I., et al., 2018. Impact of urbanization on abundance and phenology of caterpillars and consequences for breeding in an insectivorous bird. Ecol. Appl. 28, 1143-1156.
Seress, G., Sándor, K., Evans, K.L., Liker, A., 2020. Food availability limits avian reproduction in the city: an experimental study on great tits Parus major. J. Anim. Ecol. 89, 1570-1580.
Seress, G., Sándor, K., Vincze, E., Pipoly, I., Bukor, B., Ágh, N., et al., 2021. Contrasting effects of the COVID-19 lockdown on urban birds' reproductive success in two cities. Sci. Rep. 11, 17649.
Sinkovics, C., Seress, G., Pipoly, I., Vincze, E., Liker, A., 2021. Great tits feed their nestlings with more but smaller prey items and fewer caterpillars in cities than in forests. Sci. Rep. 11, 24161.
Sinkovics, C., Seress, G., Pipoly, I., Vincze, E., Liker, A., 2023. Comparison of nestling diet between first and second broods of great tits Parus major in urban and forest habitats. Anim. Biodiv. Conserv. 46, 199-212.
Sol, D., Lapiedra, O., González-Lagos, C., 2013. Behavioural adjustments for a life in the city. Anim. Behav. 85, 1101-1112.
Stauss, M., Segelbacher, G., Tomiuk, J., Bachmann, L., 2005. Sex ratio of Parus major and P. caeruleus broods depends on parental condition and habitat quality. Oikos 109, 367-373.
Suh, A., Kriegs, J.O., Brosius, J., Schmitz, J., 2011. Retroposon insertions and the chronology of avian sex chromosome evolution. Mol. Biol. Evol. 28, 2993-2997.
Sumasgutner, P., Nemeth, E., Tebb, G., Krenn, H.W., Gamauf, A., 2014. Hard times in the city – attractive nest sites but insufficient food supply lead to low reproduction rates in a bird of prey. Front. Zool. 11, 48.
Swenson, J.E., 1986. Differential survival by sex in juvenile sage grouse and gray partridge. Ornis Scand. 17, 14.
Székely, T., Liker, A., Freckleton, R.P., Fichtel, C., Kappeler, P.M., 2014. Sex-biased survival predicts adult sex ratio variation in wild birds. Proc. R. Soc. B Biol. Sci. 281, 20140342.
Tobler, M., Hasselquist, D., Smith, H.G., Sandell, M.I., 2010. Short- and long-term consequences of prenatal testosterone for immune function: an experimental study in the zebra finch. Behav. Ecol. Sociobiol. 64, 717-727.
Tschirren, B., Fitze, P.S., Richner, H., 2003. Sexual dimorphism in susceptibility to parasites and cell-mediated immunity in great tit nestlings. J. Anim. Ecol. 72, 839-845.
Vincze, E., Seress, G., Lagisz, M., Nakagawa, S., Dingemanse, N.J., Sprau, P., 2017. Does urbanization affect predation of bird nests? A meta-analysis. Front. Ecol. Evol. 26, e01475.
Von Engelhardt, N., Carere, C., Dijkstra, C., Groothuis, T.G.G., 2006. Sex-specific effects of yolk testosterone on survival, begging and growth of zebra finches. Proc. R. Soc. B Biol. Sci. 273, 65-70.
West, S.A., Sheldon, B.C., 2002. Constraints in the evolution of sex ratio adjustment. Science 295, 1685-1688.
Xirocostas, Z.A., Everingham, S.E., Moles, A.T., 2020. The sex with the reduced sex chromosome dies earlier: a comparison across the tree of life. Biol. Lett. 16, 20190867.
Table
1.
Sex ratio of dead offspring (SRDO) estimated by generalized linear mixed models (GLMM) for different sites and developmental stages. N indicates the sample size in each category (dead offspring with known sex), OR is the odds ratio (i.e. the proportional difference of the odds of a dead offspring being male compared to being female in the studied group), back-transformed from log scale. SRDO indicates the sex ratio estimates for dead offspring (expressed as the percentages of males in all dead individuals in the category), calculated from the OR. 95% confidence intervals (CI) for OR were calculated from the statistical models that did not contain any fixed effect (see Methods in the main text for explanation). Bold values highlight estimates where the CIs did not include 1, meaning that the OR is statistically different from 1, with OR > 1 indicating more dead males than dead females.
Groupa
N
OR
95% CI
SRDO
A) Full sample
All samples (embryos + nestlings)
427
1.31
1.06–1.63
56.88%
B) Site groups
Veszprém (urban)
229
1.29
1.01–1.68
56.33%
Balatonfüred (urban)
96
1.55
0.86–2.79
60.83%
Szentgál (forest)
62
1.20
0.63–2.30
54.45%
Vilma-puszta (forest)
40
1.23
0.66–2.28
55.00%
Urban and forest areas
All urban samples
325
1.36
1.08–1.73
57.63%
All forest samples
102
1.21
0.77–1.91
54.81%
D) Offspring age stages
Embryos
85
1.18
0.76–1.84
54.23%
Young nestlings (before 14 days)
236
1.44
1.09–1.91
59.01%
Old nestlings (after 14–16 days)
106
1.17
0.78–1.75
53.83%
All nestlings (young + old)
342
1.35
1.07–1.71
57.46%
a All estimates (percentages, OR, and CI) were obtained from a separate model for each group of samples. Thus, for example, the estimates for the urban area are not the simple average of the values for the two urban sites.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds