Harsh environmental conditions often impose strong selection on the phenotype of natural populations through impacts on their fitness. For overwintering birds, winter is an important period for survival because the weather conditions in winter is usually severer than in other seasons and birds often suffer more thermoregulation costs while food is in short supply. Thus, the selective effect of weather conditions on phenotype in winter is expected to be strong. In this study, we examined the relationship of overwinter survival of Silver-throated Tits (Aegithalos glaucogularis) with their morphological traits under different levels of winter conditions (winters with and without snowstorms) to explore the differential selective effect of winter weather on their morphology. We found that regardless of whether the winter experienced a snowstorm, the female Silver-throated Tits with a smaller bill surface area or smaller bill depth and a smaller surface area:volume ratio were more likely to survive during the winter, which supported the hypothesis of selection for heat retention. Furthermore, the females with a smaller body length survived better than the larger females, indicating that the lesser food requirements for smaller body sizes may confer advantages during the winter when food availability was reduced. In addition, in agreement with the finding in many short-lived birds that survival rate increases with the increase of age, older (≥2-year-old) female Silver-throated Tits had higher overwinter survival than 1-year-old females. However, we did not find any correlation of morphological traits and age with survival in male Silver-throated Tits. These results illustrate differential selective effects of winter weather on female and male Silver-throated Tits and contribute to the knowledge of sex-specific selection on the phenotype of natural populations.
Natural selection is widely accepted as the main force shaping phenotypic variation within populations (Rieseberg et al., 2002; Funk et al., 2006). Numerous studies have shown that harsh environmental conditions can lead to strong selection on animal phenotype through impacts on their fitness (Wood and Brodie III, 2016; Grant et al., 2017). The most classic example is probably the directional selection of bill size and shape during an extreme drought in Darwin's Medium Ground Finches (Geospiza fortis) in the Galapagos islands: as the smaller and softer seeds were eaten first when dry caused a food shortage, the finches who could not eat the larger and harder seeds were starved to death, whereas those with larger bills survived better (Boag and Grant, 1981). A similar but a more recent example is from a study of a Lizard (Anolis scriptus): after being battered by two hurricanes, the individuals that had survived were those with strong clinging capacity which enabled them to hold tight during the hurricanes (Donihue et al., 2018). These examples illustrate that individuals with different phenotypes have different fitness under certain environmental conditions. Since morphological traits are often heritable (Roff, 1997; Wood and Brodie, 2016), those surviving individuals will then have the opportunity to pass on the traits that help them survive to the next generation, leading to changes in the population's phenotype after selection.
For many bird species, weather conditions in winter may impose strong selective pressure because the impact of climate on their survival is often considered to be severe in winter when thermoregulatory cost is high and food is in short supply (Perdeck et al., 2000; Sæther et al., 2000). To this end, Bergmann's rule about the link between body size and geographic distribution says that the animals living in colder climates are relatively larger to reduce thermoregulatory costs (Bergmann, 1847) and as a supplementary, Allen's rule suggests that the animals living in colder environments should have relatively smaller appendages to reduce heat loss (Allen, 1877). Both theorems have received supports from various studies of birds (Murphy, 1985; Nudds and Oswald, 2007; Symonds and Tattersall, 2010; Friedman et al., 2017; Probst et al., 2021). On the other hand, winter weather often increases birds' foraging difficulty, and this is particularly true for insectivorous birds who often change their diet in winter when invertebrate foods are not available. For instance, the Great Tit (Parus major), as an insectivorous bird, has been observed to eat beech mast as a supply of emergency food whenever the weather condition in winter is bad (Van Balen, 1980). The foraging difficulty in winter is also likely to increase due to extremely climatic events such as snowstorms that may cover food with snow and ice (Robinson et al., 2007). Some studies have shown that the survival rates and abundances of wintering birds decline in years with higher snowfall, and the survivorship has a negative relationship with the extent of snow cover (Doherty and Grubb, 2002; Jones et al., 2003; Keyser et al., 2022). It can thus be expected that in the extremely adverse environments, the individuals with some advantageous phenotypes can survive better. As an example, it has been reported in Cliff Swallows (Petrochelidon pyrrhonota) that smaller-brained individuals are more likely to die during an extreme late-spring cold snap, presumably because their brain size can influence foraging success and that bigger brain size may allow advantages when conditions exceed the normal environment (Wagnon and Brown, 2020).
Furthermore, the selective effect of weather conditions on phenotype in years with severe weather conditions can differ from that in normal years. For example, Møller (2011) found that the behavioral and life history responses of Barn Swallows (Hirundo rustica) to years with extreme weather (including extremely cold and extremely warm) and normal years were different, suggesting that the strength and direction of selection may be different under different environmental conditions. Also, there may be competing selection pressures imposing on the same phenotypic trait of an individual. In this regard, although a larger body size may confer the thermal advantage of retaining heat and allow more fat to be stored (Kendeigh, 1972; Leyequién et al., 2006), limited food resources under harsh environmental conditions may offset the benefits of being large due to larger individual’ greater amount of food intake requirement (Lehikoinen, 1986). For example, a study on the correlation between morphology and winter climatic conditions of House Sparrows (Passer domesticus) found that selection favored small body size for females as winter conditions got severer (Fleischer and Johnston, 1984).
Moreover, the phenotype of males and females may suffer different selection pressure of the environments. In addition to the example of House Sparrows mentioned above (Fleischer and Johnston, 1984), an analysis of the selective effect of winter mortality on the size of Great Tits found a significant tendency of directional selection for large size in colder winters in adult males, but not in females (Lehikoinen, 1986) and similarly, a study of Barn Swallows showed the wing shape of females but not males covaried with conditions in winter (Saino et al., 2017). Therefore, different mechanisms of selection may exist for the two sexes and subtle behavioral or ecological differences between the two sexes may lead to different selection outcomes.
In this study, we examined the relationship of overwinter survival of Silver-throated Tits (Aegithalos glaucogularis) with their morphological traits under different levels of winter selection pressure (winters with and without snowstorms) to explore the differential selective effect of winter weather on their morphology. The Silver-throated Tit is an insectivorous bird that is endemic to China. The nature of the small body size (6.0–9.3 g) of this species (Li et al., 2010) makes it particularly sensitive to fluctuations in weather conditions (Leech and Crick, 2007), so it is an ideal species for studying the impact of harsh climatic conditions on its phenotype. Given that small birds have high surface area to volume ratio and limited fat storage capacity, they are expected to minimize heat loss while maximizing energy intake and thermogenic capacity during winter (Andreasson et al., 2020). We hypothesized that winter conditions would impose strong selection pressures on their morphological traits through the effects on their survival. Based on the findings in other birds, we specifically tested the following predictions. First, as birds with larger body sizes may have energetic advantages during winter, individuals with larger body sizes may survive better in winter; alternatively, larger birds may have a worse ability to survive for their greater requirement of food. Second, those individuals with smaller appendages (e.g., bill and tarsus) may have a higher survival rate for their advantage in heat retention. Third, the birds with enhanced flight abilities (i.e., large wings) would have fitness advantages during winter because they can forage over a larger area (Saino et al., 2017). Also, we expected that the above selective effect of winter weather on Silver-throated Tits’ morphological traits might differ between females and males, given that males are larger than females (Li et al., 2010), which may lead to their differences in fat storage and heat retention. Besides, we expected that the selective effect of winter weather on their morphological traits might depend on winter conditions (i.e., with and without snow storms). For example, although we expected that the individuals with a smaller bill might be more advantageous in reducing heat dissipation, the individuals with a larger bill might have superiority in obtaining food covered under snow.
2.
Methods
2.1
Study population and data collection
The Silver-throated Tit population in this study is located in the Zhanglou Village and its surrounding area (ca. 4.5 km2) of the Dongzhai National Nature Reserve (31.95° N, 114.25° E; altitude 100–840 m) in central China and is in the transitional region between temperate and subtropical zones. The Silver-throated Tits are resident at the study area and are small passerine birds with sexually similar plumage, despite that males are lightly larger than females in morphometric measurements (Li et al., 2010). We started to monitor this Silver-throated Tit population in the spring of 2011 and thereafter, annual fieldwork was generally carried out from each year's December to May of the following year. In winters (December to January), we mist-netted adults and banded each of them with a unique combination of metal and color rings. We also measured their morphological traits and collected a small blood sample from each bird for sex identification using genetic method (for detailed information, see Li et al., 2010; Wang et al., 2010). During the breeding seasons (February to May), we searched for nests and monitored the nests regularly (usually every two days). When any adult of a nest was found unbanded, we mist-netted and banded it in the same way as in winter. Besides, we banded the nestlings from the nests when they were around 10 days old.
A total of seven morphological traits were measured for each bird. Bill length, bill depth, bill-head length and tarsus length were measured using a digital caliper (±0.01 mm), and wing length, tail length and total body length were measured using a ruler (±0.5 mm) (for detailed information about morphometric measurements, see Li et al., 2010). We also weighed each bird with an electronic balance (±0.01 g). In addition, we used a bird's bill head length minus bill length to represent its head length (Carvalho et al., 2021), and total body length minus tail length to represent its body length. The measurement of most individuals in this study was conducted by the same person. To provide consistency and eliminate interobserver error, only the measurement data of the same person were used in the analyses. Where a bird was measured more than once in the same winter, we only used the measurement when it was measured for the first time.
The age of each individual when they were captured was determined based on our annual banding records. As the Silver-throated Tit population in the study area has been well monitored since 2011, we treated the age of the adult birds that were banded during the first year of work in the new area as unknown, and in the following years, any newly captured unbanded adults were assumed as the birds that were born outside the study area in the previous spring and dispersed into the study area (i.e., their age was one year old). This is reasonable because the adults of Silver-throated Tits normally do not disperse after their first year's breeding, and through extensive fieldwork, most breeding adults (>90%) and fledglings (>90%) born in the study area had been banded every year. The birds whose actual age had not been determined were excluded from the analyses.
Since no banding work was performed in the winters of 2011, 2012 and 2013, and for the winter of 2016 we only have the banding data after a snowstorm, this study only uses the winter banding data of 2014–2015 and 2017–2019, among which 2015 and 2018 were the years that experienced snowstorm, while 2014, 2017 and 2019 were the years that did not experience snowstorm. For the distinguishment of the years with and without the snowstorm, we followed the definition of a snowstorm in meteorology, i.e., the 24-h precipitation is not less than 10.0 mm during a snowfall event (Zang, 2009). The daily climate data were from the Xinyang meteorological station (~32 km from the center of our study area) and were further confirmed for the occurrence of snowstorms based on the historical weather of Xinyang on the “Weather Post Report Website” (http://www.tianqihoubao.com/lishi/xinyang.html). This resulted in the identification of the winters of 2015 and 2018 as having snowstorms (Appendix Table S1). The study on Cliff Swallows (20–28 g, about 2–3 times larger than Silver-throated Tits) has shown that if the cold lasted for 4 days, it would cause a large number of individual deaths (Brown and Brown, 1998). In the snowstorm events of 2015 and 2018, the temperature all dropped by at least 4 ℃ within a day and the mean temperature was all below 0 ℃ for at least 6 days thereafter (Appendix Table S1), so under the combinations of low temperature and snow cover, we believe that the weather was severe enough to produce strong impacts on the survival of Silver-throated Tits.
2.2
Statistical analyses
The final sample sizes for this study are shown in Table 1. We performed all statistical analyses in R 4.1.1 (R Development Core Team, 2021). To analyze the effect of different morphological traits on the overwinter survival of Silver-throated Tit in winter with or without snowstorm, we built generalized linear mixed models (GLMM) with a binomial distribution and a logit link using the “glmer” function in the package “lme4” (Bates et al., 2020). In the GLMM, whether the winter-banded individuals survived from winter to the breeding season (1 and 0 represent surviving and not surviving to the breeding season, respectively) was treated as the response variable. Since we were able to monitor the vast majority of individuals that appeared in the breeding season, for individuals banded in winter, if it was recorded again in the breeding season, it was regarded to have survived the winter; otherwise, it was regarded to have died in winter. Although there is a possibility that individuals presumed dead may have dispersed outside our study area, this will not affect our results unless there is also a morphological difference between the birds that dispersed and those did not disperse. For the explanatory variables, we included each morphological trait (including body mass, bill length, bill depth, head length, tarsus length, wing length, tail length and body length), whether the winter experienced a snowstorm, the relative capture date of the bird (calculated as the number of days from the date when the individual was captured to December 1st), the age of the bird (classified as 1-year-old and ≥ 2-year-old), and the interactions of whether the winter experienced a snowstorm with every morphological trait. In addition, the year and bird ID were included as the random factors to account for the nonindependence of data from the same year and from the same individual that occurred across years. Females and males were analyzed separately because the selection pressure may be different between the sexes. In addition, because the bill surface area is related to the heat loss in cold environments (Tattersall et al., 2017), which may affect the thermoregulatory ability in winter and thus affect the overwinter survival, we also replaced the bill length and bill depth with bill surface area to repeat the same analyses as above. Each bird's bill surface area was estimated using the following formula (Greenberg et al., 2012a; Greenberg and Danner, 2012):
bill depth 2× bill length
(1)
Table
1.
The sample size for generalized linear mixed model analyses.
Sex
Not survive into the breeding season
Survive into the breeding season
Suffer snowstorm
Not suffer
Suffer snowstorm
Not suffer
Male (N = 163)
30
37
31
65
Female (N = 200)
45
61
32
62
Note: the sample size refers to the number of observations; a total of 25 birds have repeated observations, including 23 birds produced records across two years and the remaining two birds had records across three years.
Assuming the bill of a Silver-throated Tit is a cone, the above formula is adapted from the formula for the lateral surface area of a cone A = πrL, where L is the slant height of the cone (~ bill length) and r is the radius of the base circle (~1/2 bill depth).
Besides, we also followed Dubiner and Meiri (2022) and explored the effect of the surface area:volume ratio on Silver-throated Tit survival. Because of the long tails (the tail length is about half of the total body length) of our study species, we calculated the surface area:volume ratio in two ways: the total body length2/body mass and body length2/body mass (the “total body length” was measured from the tip of the bill to the end of the tail, whereas the “body length” was obtained by subtracting tail length from total body length). We then replaced body length and body mass with the above two “surface area:volume ratio” respectively in the explanatory variables of the generalized linear mixed models for analysis.
Before the model analyses, we used the “vif” function in the package “car” (John et al., 2019) to assess collinearity among the predictor variables by calculating the variance inflation factor (VIF). All VIF values were well below a threshold of 2, which indicated no evidence of collinearity among variables (Zuur et al., 2010). To simplify the models, we compared all possible combinations of global model terms by using the “dredge” function in the package “MuMIn” (Bartoń, 2019). Akaike's information criterion corrected for small sample size (AICc) was used to assess model fit (Burnham and Anderson, 2002), we considered models with ΔAICc ≤2 as equivalent alternative models. If there were more than one model with ΔAICc ≤2, we used the “model.avg” function in the package “MuMIn” to output the results and considered an effect to be significant when P < 0.05.
3.
Results
More than one best-supported models were obtained after the model selection procedure (see Appendix Tables S2–S7). For the analyses while using body length and body mass (instead of surface area:volume ratio) and including either bill length and bill depth or bill surface area as the model terms, the final models for males (obtained after averaging the best-supported models in Appendix Tables S2 and S3) did not detect any significant effects of the explanatory variables on male's overwinter survival (Table 2). This suggests that the morphological traits and whether the winter experienced a snowstorm did not affect the overwinter survival of the males. A notable point is that a bird's age had a marginally significant effect on its survival, with the older (≥2-year-old) males tending to survive better than the younger (1-year-old) males (Table 2).
Table
2.
Model average results from the best model set for the factors (including body length and body mass) that affect the overwinter survival rates of male Silver-throated Tits.
Fixed effects
Analysis using bill length and bill depth
Analysis using bill surface area
Estimate
SE
Z
P
Estimate
SE
z
P
Intercept
3.555
7.053
0.502
0.616
1.870
5.956
0.313
0.755
Age 2a
0.643
0.348
1.833
0.067
0.667
0.346
1.910
0.056
Suffer 1b
2.637
8.296
0.317
0.751
3.771
9.452
0.398
0.691
Bill surface area
−0.414
0.268
1.538
0.124
Bill length
−1.000
0.663
1.497
0.134
Bill depth
−1.228
1.614
0.756
0.450
Head length
−0.225
0.505
0.443
0.658
−0.156
0.511
0.304
0.761
Head length × suffer 1b
−1.311
0.837
1.553
0.120
−1.311
0.837
1.553
0.120
Wing length
0.086
0.110
0.776
0.438
0.096
0.110
0.867
0.386
Tarsus length
0.222
0.393
0.559
0.576
0.262
0.398
0.655
0.513
Tail length
0.046
0.085
0.535
0.593
Body mass
0.012
0.590
0.020
0.984
0.044
0.573
0.076
0.939
Body mass × suffer 1b
−1.373
0.892
1.527
0.127
−1.373
0.892
1.527
0.127
a Age 1 (1-year-old) is considered as the reference level. Age 2 represents all individuals older than one year. b Years without snowstorm are the reference category. 2015 and 2018 are the years that experienced snowstorm, and 2014, 2017 and 2019 are the years that without experienced snowstorm.
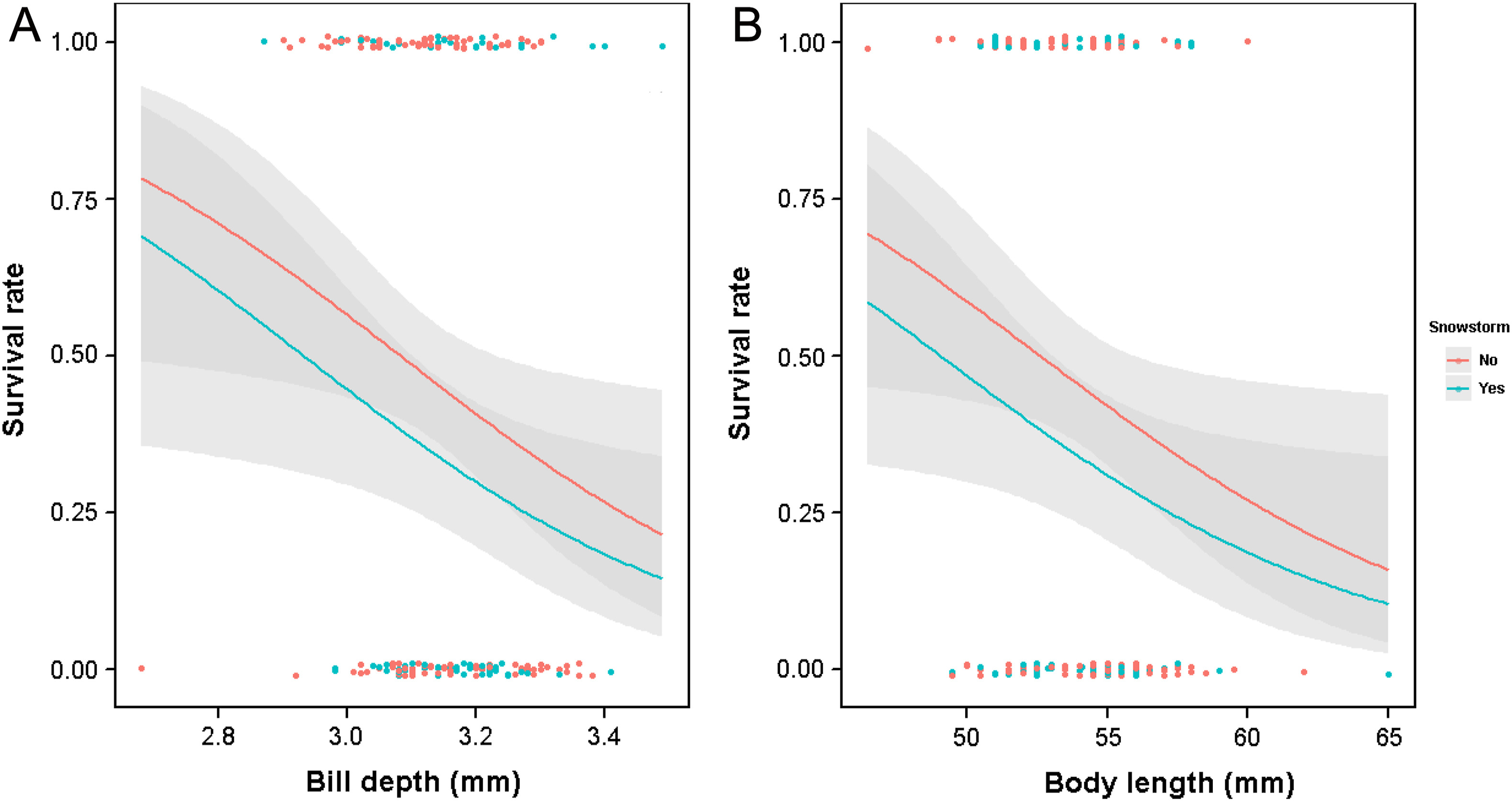
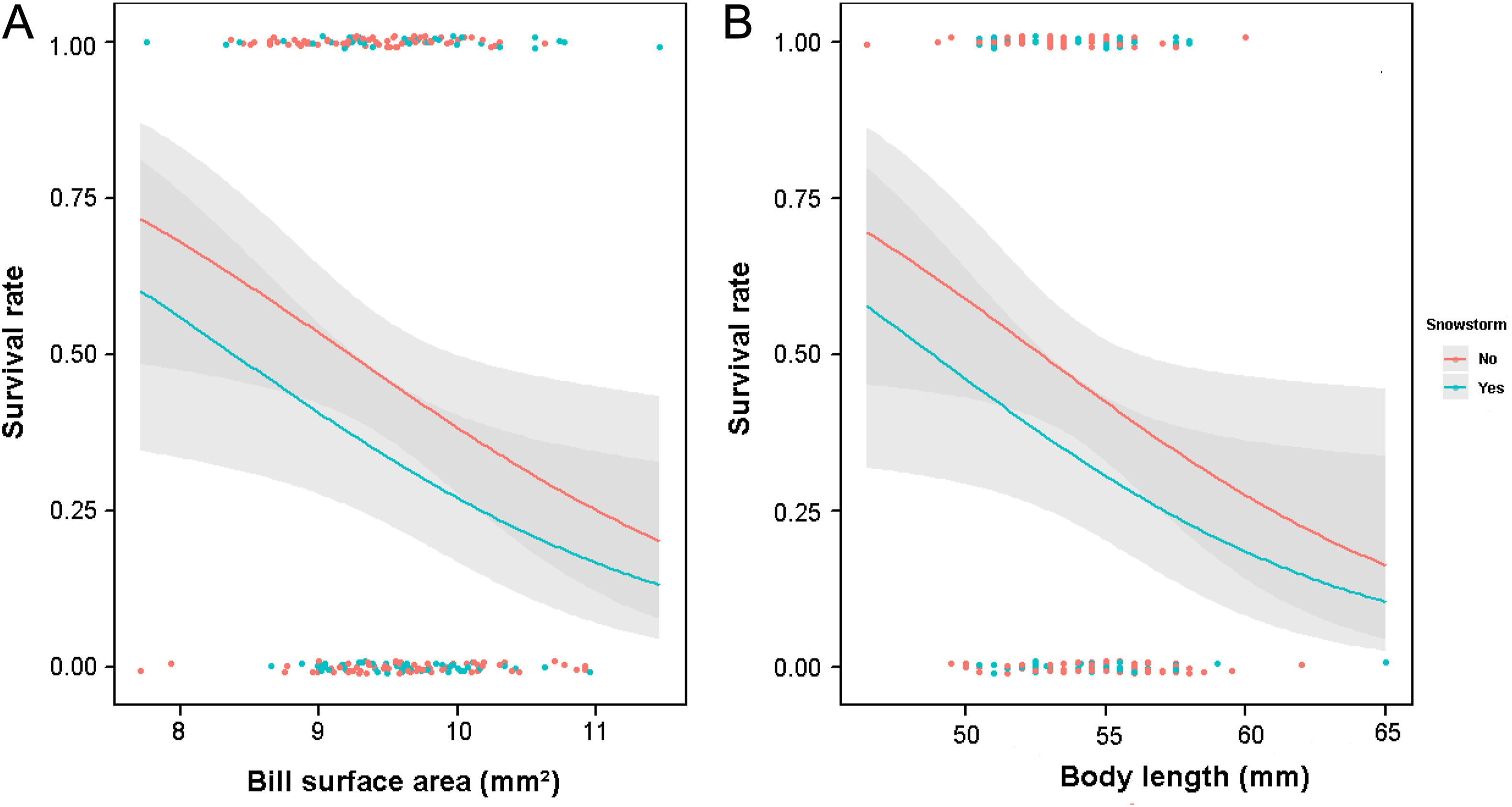
For the females, the average model from the best-supported models (Appendix Table S2) suggests that regardless of whether the winter experienced a snowstorm, both bill depth and body length had significantly negative effects on female overwinter survival, with the females having smaller bill depth and shorter body length surviving better (Table 3; Fig. 1). Also, female age had a significant effect on their overwinter survival and the older (≥2-year-old) females survived better than the younger (1-year-old) females (Table 3). When replacing bill length and bill depth with bill surface area in the analysis, the average model obtained from the best-supported models (Appendix Table S3) additionally included tail length in the model, except that bill length and bill depth were replaced by bill surface area (Table 3). It was found that the females with smaller bill surface area and shorter body length survived significantly better in winter (Table 3; Fig. 2), and again, the older females (≥2-year-old) had a significantly better survivorship than the younger females (1-year-old).
Table
3.
Model average results from the best model set for the factors (including body length and body mass) that affect the overwinter survival rates of female Silver-throated Tits.
Fixed effects
Analysis using bill length and bill depth
Analysis using bill surface area
Estimate
SE
z
P
Estimate
SE
z
P
Intercept
18.237
7.231
2.509
0.012
12.398
5.755
2.143
0.032
Age 2a
1.242
0.400
3.086
0.002
1.235
0.401
3.061
0.002
Suffer 1b
−6.787
8.860
0.763
0.445
−4.184
6.344
0.657
0.511
Date
−0.005
0.011
0.436
0.663
−0.007
0.011
0.646
0.518
Bill surface area
−0.663
0.289
2.278
0.023
Bill surface area × suffer 1b
0.554
0.527
1.044
0.296
Bill length
−0.673
0.612
1.094
0.274
Bill depth
−3.515
1.667
2.097
0.036
Bill depth × suffer1b
3.793
2.910
1.295
0.195
Body length
−0.153
0.075
2.031
0.042
−0.155
0.077
2.012
0.044
Body length × suffer 1b
0.160
0.131
1.217
0.223
0.159
0.131
1.211
0.226
Body mass
0.495
0.427
1.152
0.249
0.537
0.424
1.257
0.209
Wing length
0.108
0.119
0.900
0.368
0.114
0.119
0.956
0.339
Tarsus length
0.151
0.351
0.428
0.669
0.285
0.370
0.764
0.445
Tail length
0.039
0.070
0.552
0.581
a Age 1 (1-year-old) is considered as the reference level. Age 2 represents all individuals older than one year. b Years without snowstorm are the reference category. 2015 and 2018 are the years that experienced snowstorm, and 2014, 2017 and 2019 are the years that without experienced snowstorm.
Figure
1.
Predicted overwinter survival rates (with 95% CI) of female Silver-throated Tits in relation to the bill depth (A) and body length (B) using the best model in Appendix Table S2 (analysis using bill length and bill depth).
Figure
2.
Predicted overwinter survival rates (with 95% CI) of female Silver-throated Tits in relation to the bill surface area (A) and body length (B) using the best model in Appendix Table S3 (analysis using bill surface area).
When repeating the above analyses by replacing body length and body mass with surface area:volume ratio to explore the effect of surface area:volume ratio on Silver-throated Tit survival (Appendix Tables S4–S7), the surface area:volume ratio calculated by body length2/body mass had no significant effect on male survival (Table 4) but had a significantly negative effect on female survival (Table 5). When using the surface area:volume ratio calculated by total body length2/body mass for the analysis, no significant effect on the survival of either sex was detected (Appendix Tables S8 and S9). In addition, the effects of all other variables (i.e., age, bill depth and bill surface area) in these analyses were qualitatively similar to those above (Table 2 and 3).
Table
4.
Model average results from the best model set for the factors (including surface area:volume ratio calculated with body length) that affect the overwinter survival rates of male Silver-throated Tits.
Fixed effects
Analysis using bill length and bill depth
Analysis using bill surface area
Estimate
SE
z
P
Estimate
SE
z
P
Intercept
3.640
6.952
0.521
0.602
1.937
5.860
0.329
0.742
Age 2a
0.643
0.348
1.836
0.066
0.669
0.346
1.922
0.055
Suffer 1b
1.914
8.097
0.236
0.814
2.908
9.474
0.306
0.760
Surface area:volume ratio
0.002
0.005
0.465
0.642
0.002
0.005
0.422
0.673
Bill surface area
−0.415
0.268
1.538
0.124
Bill length
−0.998
0.663
1.494
0.135
Bill depth
−1.228
1.614
0.756
0.450
Head length
−0.225
0.505
0.443
0.658
−0.156
0.511
0.304
0.761
Head length × Suffer 1b
−1.311
0.837
1.553
0.120
−1.311
0.837
1.553
0.120
Wing length
0.086
0.109
0.776
0.438
0.096
0.110
0.867
0.386
Tarsus length
0.222
0.393
0.559
0.576
0.262
0.398
0.655
0.513
Tail length
0.046
0.085
0.535
0.593
a Age 1 (1-year-old) is considered as the reference level. Age 2 represents all individuals older than one year. b Years without snowstorm are the reference category. 2015 and 2018 are the years that experienced snowstorm, and 2014, 2017 and 2019 are the years that without experienced snowstorm.
Table
5.
Model average results from the best model set for the factors (including surface area: volume ratio calculated with body length) that affect the overwinter survival rates of female Silver-throated Tits.
Fixed effects
Analysis using bill length and bill depth
Analysis using bill surface area
Estimate
SE
Z
P
Estimate
SE
z
P
Intercept
16.370
6.108
2.666
0.008
10.122
4.385
2.295
0.022
Age 2a
1.196
0.396
3.005
0.003
1.189
0.397
2.977
0.003
Suffer 1b
−6.275
8.644
0.724
0.469
−1.860
3.198
0.579
0.562
Date
−0.005
0.011
0.468
0.640
−0.008
0.011
0.719
0.472
Surface area:volume ratio
−0.010
0.005
2.122
0.034
−0.010
0.004
2.175
0.030
Surface area:volume ratio × Suffer 1b
0.009
0.008
1.123
0.262
0.008
0.008
1.045
0.296
Bill surface area
−0.690
0.284
2.416
0.016
Bill surface area × Suffer 1b
0.513
0.538
0.948
0.343
Bill length
−0.733
0.612
1.189
0.234
Bill depth
−3.650
1.667
2.177
0.029
Bill depth × Suffer 1b
3.642
2.934
1.234
0.217
Wing length
0.046
0.116
0.396
0.692
0.058
0.117
0.489
0.625
Tail length
0.034
0.069
0.488
0.626
Head length
−0.163
0.378
0.430
0.667
a Age 1 (1-year-old) is considered as the reference level. Age 2 represents all individuals older than one year. b Years without snowstorm are the reference category. 2015 and 2018 are the years that experienced snowstorm, and 2014, 2017, 2019 are the years that without experienced snowstorm.
By comparing the characteristics of the Silver-throated Tits that survived the winter and those did not, our study show that some morphological traits, such as bill and body size, might be important for their survival and this phenomenon was sex-specific. However, despite that we expect the selective effect on the morphological traits of Silver-throated Tits to be different between winters with and without snowstorms, we fail to find differing relationships between individual survival and morphological traits with respect to the presence or absence of snowstorms.
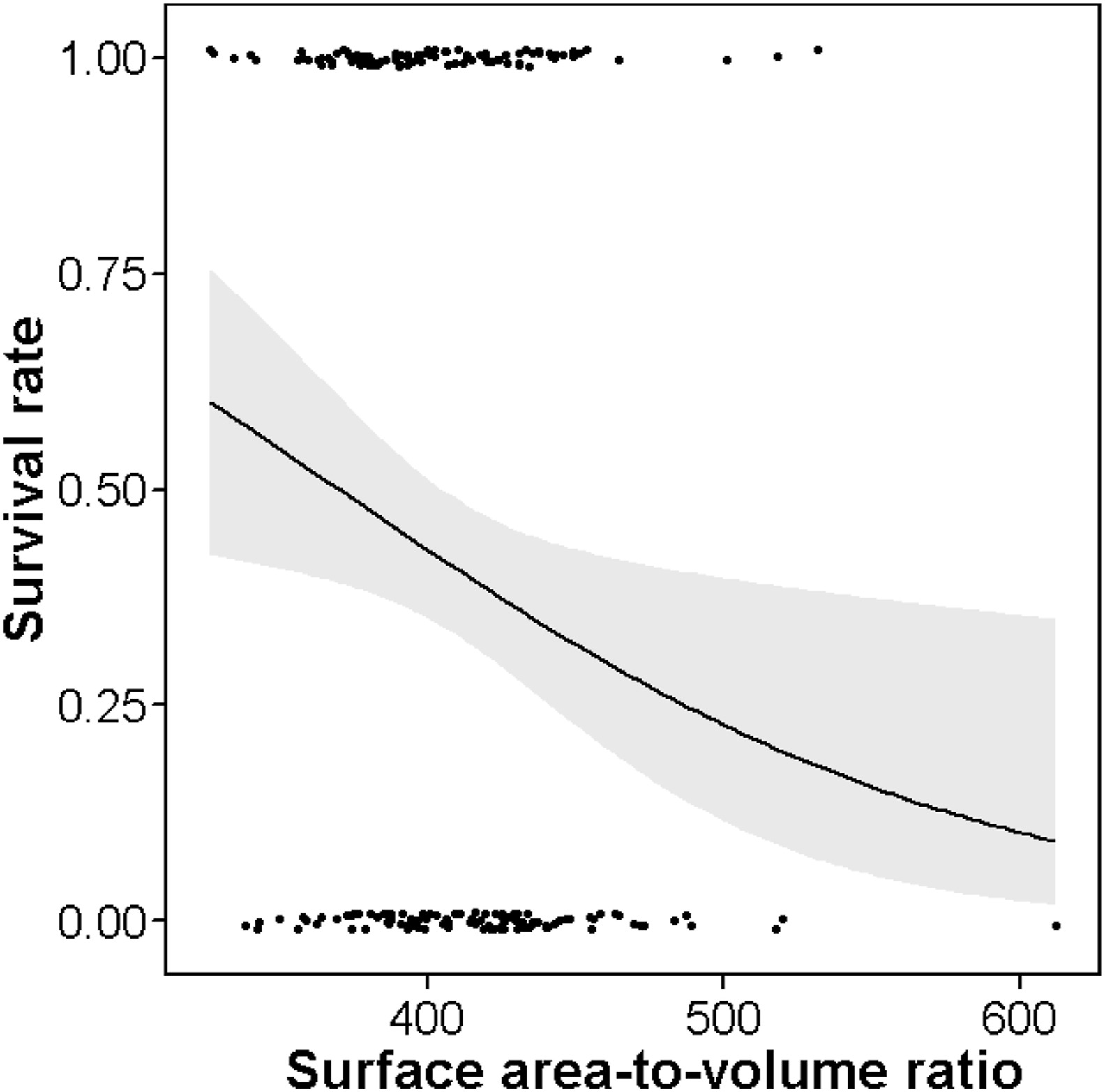
Recent findings suggest that the bill of birds is an effective heat-exchange organ and plays an important role in thermoregulation in birds (Tattersall et al., 2009, 2017; Greenberg et al., 2012a; Economo et al., 2017). This is also true for even small-billed passerine birds. As an instance, although Song Sparrow's (Melospiza melodia) bill represents only 2% of the body surface area, when the ambient temperatures down to 15 ℃, the total body heat loss through its surface can be as high as 10% of basal heat production (Greenberg et al., 2012b). Hence, the size of the bird's bill may be selected by the need for thermoregulation. In this regard, it has been found that colder winters were associated with the smaller bill surface area of the song sparrows in eastern North America (Danner and Greenberg, 2015). Similar findings have also been reported in interspecific comparisons: Friedman et al. (2017) found that low winter temperature was correlated with low bill surface area, length and width of 158 species of the Australasian honeyeaters and allies (Meliphagoidea) and Symonds and Tattersall (2010) found that species in colder climates had significantly shorter bills in a phylogenetic comparative analyses of 214 bird species. All of these suggests that selection on bill size can be explained by the bill's role in thermoregulation. For female Silver-throated Tits, we found that the individuals with smaller bill surface area or smaller bill depth were more likely to survive during winter. Supporting the hypothesis of selection for heat retention (Danner and Greenberg, 2015), we think the result may relate to the thermoregulatory function of their bills. Because of their small body size, Silver-throated Tits may have a high thermoregulatory burden and a smaller bill relative to body size may help to reduce the loss of body heat to the environment when experiencing colder climates during winter. In line with these results, we also found that the females with smaller surface area:volume ratios had better survivorship in the winter (Fig. 3), as the individuals with lower surface area:volume ratios can retain heat better (Bergmann, 1847). However, we did not find an association between tarsus length and survival in the present study. This result is in line with the findings of Symonds and Tattersall (2010) and Onley et al. (2020). They suggested that this may be due to physiological differences in how the tarsus controls heat loss: the tarsus has a countercurrent heat exchange mechanism, and the central artery is surrounded by veins, through which heat from arterial blood can circulate without losing too much heat to the environment (Steen and Steen, 1965; Symonds and Tattersall, 2010; Onley et al., 2020).
Figure
3.
Predicted overwinter survival rates (with 95% CI) of female Silver-throated Tits in relation to the surface area:volume ratio calculated by body length2/body mass using the best model in Appendix Table S4.
Contrary to the findings for the Great Tits mentioned earlier (Lehikoinen, 1986), our research shows a significantly negative correlation between body length and overwinter survival in females but not in male Silver-throated Tits, i.e., the females with smaller body lengths were more likely to survive. During the period of cold weather and food scarcity, a larger body size may confer both the thermal advantage of retaining heat and the benefit of allowing more fat to be stored (Kendeigh, 1972; Leyequién et al., 2006), so larger birds may have advantages over small birds in winter. However, this may not always be true. Because of larger individuals’ greater amount of food intake, limited food resources may offset the benefits of their large size under harsh environmental conditions (Lehikoinen, 1986). A study about the correlations between the morphology of House Sparrows and winter climatic conditions found that selection led to small body size for females as winter conditions got severer (Fleischer and Johnston, 1984). Therefore, it is possible that the female Silver-throated Tits with shorter body lengths in our study may survive better than larger females for their relatively lesser food requirement during the winter when food availability was reduced.
Furthermore, no morphological traits of male Silver-throated Tits were found to have a significant effect on their survival, indicating that winter weather had a sex-specific selective effect on Silver-throated Tits’ morphological traits. We think this may be due to the physiological or behavioral differences between males and females that lead to different selection outcomes. For example, male Silver-throated Tits are larger than females (Li et al., 2010), which means that they may have more fat reserves and relatively lower heat loss. Also, male birds may be dominant over females when foraging (e.g., Great Tits, Barluenga et al., 2000; Common Ravens Corvus corax, Boucherie et al., 2022) and can gain more foods than females. All these may result in relatively weaker selection pressure on males than on females. However, these possibilities need to be further investigated in future studies.
We also found an effect of individual age on over-winter survival: the older female Silver-throated Tits had higher overwinter survival than those 1-year-old females and a similar trend (though non-significant) was also observed for males (Table 3). In our study population, the longevity of most Silver-throated Tits is 1–3 years, so our result agrees with the finding that the survival rate increases with the increase of age in many short-lived birds (Sæther and Bakke, 2000). In addition, a study of Blue Tits (Cyanistes caeruleus) showed that young Blue Tits maintain a higher body temperature than older individuals when at predation risk under cold ambient temperatures at night, which indicates that young birds may need to generate more heat than older birds (Andreasson et al., 2019, 2020). For the Silver-throated Tits, it is likely that the young birds need to generate more heat to maintain a higher body temperature but have a worse ability of foraging than the older birds, thereby experience a higher mortality rate in winter.
However, we did not detect that the winters with snowstorms had a significantly different influence on bird's survival than the winters without snowstorms in either sex. Previous research suggested that the negative effect of high snowfall on birds' survival is primarily through its impacts on food availability (Shipley et al., 2020; Keyser et al., 2022). One possible reason for our result is that the impact of heavy snow at our study site was not strong enough to affect the food availability of Silver-throated Tits. Unlike the species that forage on the ground, Silver-throated Tits mainly forage in the canopy or tree trunks, and still have insect eggs that lie on the surface or inside the branches and plant buds to eat when snow covers. A study on the feeding habits of congeneric Long-tailed Tit (A. caudatus) in Northeast China showed that the animal and plant foods accounted for about 50% of their diet respectively in winter (Song, 1981), which indicated that these species have flexible foraging strategies under the situation of food shortage. Since snowstorms provide no additional influence on Silver-throated Tit survival, our result also implies that cold temperature may be more important in determining winter survival rate of Silver-throated Tits at our study site.
5.
Conclusions
In conclusion, through a comparation of Silver-throated Tits that survived and did not survive in winters, we found that whether the winter experienced snowstorms did not affect the survival of Silver-throated Tits and the selection pressure on morphological traits of males and females in winter is sex-specific. The result adds to the knowledge of the sex-specific effects of natural selection pressure on morphological traits in birds. Since the direction and strength of selection pressure in nature often change, making its relationship with phenotypes complicated (Siepielski et al., 2009), we suggest that more such studies on wild populations to be conducted in future to fully understand the role of different selective pressures on animal morphological traits. It should be noted that our study has assumed that dispersal would not affect our result as long as no morphological difference exists between the birds that dispersed and those did not disperse. Given that morphological traits may correlate with dispersal ability in some species (Dawideit et al., 2009; Sheard et al., 2020), the assumption needs to be verified in future studies of Silver-throated Tits.
Authors’ contributions
JL and JX conceived the study; YW, QH and JL conducted fieldwork; YW and QH conducted analyses; YW and JL wrote the manuscript. All authors read and approved the final manuscript.
Ethics statement
Our study was carried out in agreement with the Law of the People's Republic of China on the Protection of Wildlife and was approved by the Administrative Bureau of the Dongzhai National Nature Reserve.
Declaration of competing interest
The authors declare that they have no competing interests.
Acknowledgements
This work was made possible thanks to the Dongzhai National Nature Reserve. Special thanks are given to Prof. Zhengwang Zhang at Beijing Normal University for his earlier advice and ongoing support of this project. We are also grateful to all students, volunteers and field assistants, especially Peng Zhang, who helped with data collection in the field. This work was supported by the National Natural Science Foundation of China (31970421, 31472011, 31101644).
Allen, J.A., 1877. The influence of physical conditions in the gensis of species. Radical Rev. 1, 108-140.
Andreasson, F., Nord, A., Nilsson, J.Å., 2019. Age-dependent effects of predation risk on night-time hypothermia in two wintering passerine species. Oecologia 189, 329-337.
Andreasson, F., Nord, A., Nilsson, J.Å., 2020. Age differences in night-time metabolic rate and body temperature in a small passerine. J. Comp. Physiol. B 190, 349-359.
Van Balen, J.H., 1980. Population fluctuations of the great tit and feeding conditions in winter. Ardea 68, 143-164.
Barluenga, M., Barbosa, A., Moreno, E., 2000. Social relationships due to sex, age and morphology in great tits Parus major wintering in a mountainous habitat of Central Spain. Ardeola 47, 19-27.
Bartoń, K., 2019. Package MuMIn.
Bates, D., Maechler, M., Bolker, B., Walker, S., Bojesen, R.H., Singmann, H., et al., 2020.
Package Lme4: Linear Mixed-Effects Models Using Eigen and S4.
Bergmann, C., 1847. Über die Verhältnisse der warmeokonomiedie Thiere zu ihrer grösse. Göttinger Studien 3, 359-708.
Boag, P.T., Grant, P.R., 1981. Intense natural selection in a population of Darwin’s finches (Geospizinae) in the Galápagos. Science 214, 82-85.
Boucherie, P.H., Gallego-Abenza, M., Massen, J.J.M., Bugnyar, T., 2022. Dominance in a socially dynamic setting: hierarchical structure and conflict dynamics in ravens’ foraging groups. Philos. T. Roy. Soc. B 377, 20200446.
Brown, C.R., Brown, M.B., 1998. Intense natural selection on body size and wing and tail asymmetry in cliff swallows during severe weather. Evolution 52, 1461-1475.
Burnham, K.P., Anderson, D.R., 2002. Model Selection and Multimodel Inferernce: A
Practical Information-Theoretic Approach. Springer Science and Business Media,
New York.
Carvalho, C.F., Batalha, H.R., Cardoso, G.C., 2021. Personality is independent of morphological differences in common waxbills. Anim. Behav. 175, 175-179.
Danner, R.M., Greenberg, R., 2015. A critical season approach to Allen’s rule: bill size declines with winter temperature in a cold temperate environment. J. Biogeogr. 42, 114-120.
Dawideit, B.A., Phillimore, A.B., Laube, I., Leisler, B., Böhning-Gaese, K., 2009. Ecomorphological predictors of natal dispersal distances in birds. J. Anim. Ecol. 78, 388-395.
Doherty, P.F., Grubb, T.C., 2002. Survivorship of permanent-resident birds in a fragmented forest landscape. Ecology 83, 844-857.
Donihue, C.M., Herrel, A., Fabre, A.C., Kamath, A., Geneva, A.J., Schoener, T.W., et al., 2018. Hurricane-induced selection on the morphology of an island lizard. Nature 560, 88-91.
Dubiner, S., Meiri, S., 2022. Widespread recent changes in morphology of old world birds, global warming the immediate suspect. Glob. Ecol. Biogeogr. 31, 791-801.
Fleischer, R.C., Johnston, R.F., 1984. The relationships between winter climate and selection on body size of house sparrows. Can. J. Zool. 62, 405-410.
Funk, D.J., Nosil, P., Etges, W.J., 2006. Ecological divergence exhibits consistently positive associations with reproductive isolation across disparate taxa. P. Natl. Acad. Sci. USA 103, 3209-3213.
Grant, P.R., Rosemary Grant, B., Huey, R.B., Johnson, M.T.J., Knoll, A.H., Schmitt, J., 2017. Evolution caused by extreme events. Philos. T. Roy. Soc. B 372, 20160146.
Greenberg, R., Danner, R., Olsen, B., Luther, D., 2012a. High summer temperature explains bill size variation in salt marsh sparrows. Ecography 35, 146-152.
Greenberg, R., Cadena, V., Danner, R.M., Tattersall, G., 2012b. Heat loss may explain bill size differences between birds occupying different habitats. PLoS One 7, e40933.
Greenberg, R., Danner, R.M., 2012. The influence of the california marine layer on bill size in a generalist songbird. Evolution 66, 3825-3835.
John, A., Weisberg, S., Price, B., Adler, D., Baud-bovy, G., et al., 2019. Package Car. .
Kendeigh, S.C., 1972. Energy control of size limits in birds. Am. Nat. 106, 79-88.
Keyser, S.R., Fink, D., Gudex-Cross, D., Radeloff, V.C., Pauli, J.N., Zuckerberg, B., 2022. Snow cover dynamics: an overlooked yet important feature of winter bird occurrence and abundance across the United States. Ecography 2022, e06378.
Leech, D.I., Crick, H.Q.P., 2007. Influence of climate change on the abundance, distribution and phenology of woodland bird species in temperate regions. Ibis 149, 128-145.
Lehikoinen, E., 1986. Dependence of winter survival on size in the great tit Parus major. Ornis Fenn. 63, 10-16.
Leyequien, E., De Boer, W.F., Cleef, A., 2006. Influence of body size on coexistence of bird species. Ecol. Res. 22, 735-741.
Li, J., Wang, N., Wang, Y., Lin, S., Li, Q., Liu, Y.Y., et al., 2010. Sexual size dimorphism and sex identification using morphological traits of two Aegithalidae species. Zool. Sci. 27, 946-951.
Moeller, A.P., 2011. Behavioral and life history responses to extreme climatic conditions: Studies on a migratory songbird. Curr. Zool. 57, 351-362.
Murphy, E.C., 1985. Bergmann’s Rule, seasonality, and geographic variation in body size of house sparrows. Evolution 39, 1327-1334.
Nudds, R.L., Oswald, S.A., 2007. An interspecific test of Allen’s rule: Evolutionary implications for endothermic species. Evolution 61, 2839-2848.
Onley, I.R., Gardner, J.L., Symonds, M.R.E., 2020. Spatial and temporal variation in morphology in Australian whistlers and shrike-thrushes: Is climate change causing larger appendages? Biol. J. Linn. Soc. 130, 101-113.
Perdeck, A.C., Visser, M.E., Balen, J.H.V., 2000. Great tit Parus major survival, and the beech-crop cycle. Ardea 88, 99-106.
Probst, C.M., Ralston, J., Bentley, I., 2021. Effects of climate on bill morphology within and across toxostoma thrashers. J. Avian Biol. 2022, e02871.
R Development Core Team, 2021. R: A Language and Environment for Statistical
Computing Version 4.1.1. R Foundation for Statistical Computing. .
Rieseberg, L.H., Widmer, A., Arntz, A.M., Burke, J.M., 2002. Directional selection is the primary cause of phenotypic diversification. P. Natl. Acad. Sci. USA 99, 12242-12245.
Robinson, R.A., Baillie, S.R., Crick, H.Q.P., 2007. Weather-dependent survival: implications of climate change for passerine population processes. Ibis 149, 357-364.
Roff, D.A., 1997. Evolutionary Quantitative Genetics. Springer, New York.
Saether, B.E., Bakke, Ø., 2000. Avian life history variation and contribution of demographic traits to the population growth rate. Ecology 81, 642-653.
Saether, B.E., Tufto, J., Engen, S., Jerstad, K., Røstad, O.W., Skatan, J.E., 2000. Population dynamical consequences of climate change for a small temperate songbird. Science 287, 854-856.
Saino, N., Ambrosini, R., Caprioli, M., Liechti, F., Romano, A., Rubolini, D., et al., 2017. Wing morphology, winter ecology, and fecundity selection: evidence for sex-dependence in barn swallows (Hirundo rustica). Oecologia 184, 799-812.
Sheard, C., Neate-Clegg, M.H.C., Alioravainen, N., Jones, S.E.I., Vincent, C., MacGregor, H.E.A., et al., 2020. Ecological drivers of global gradients in avian dispersal inferred from wing morphology. Nat. Commun. 11, 2463.
Shipley, A.A., Cruz, J., Zuckerberg, B., 2020. Personality differences in the selection of dynamic refugia have demographic consequences for a winter-adapted bird. Proc. R. Soc. B 287, 20200609.
Siepielski, A.M., Dibattista, J.D., Carlson, S.M., 2009. It’s about time: The temporal dynamics of phenotypic selection in the wild. Ecol. Lett. 12, 1261-1276.
Song, Y., 1981. Studies on the breeding behaviour and feeding habits of long-tailed tits. Zool. Res. 2, 235-242.
Steen, I., Steen, J.B., 1965. The importance of the legs in the thermoregulation of birds. Acta Physiol. Scand. 63, 285-291.
Symonds, M.R.E., Tattersall, G.J., 2010. Geographical variation in bill size across bird species provides evidence for Allen’s rule. Am. Nat. 176, 188-197.
Tattersall, G.J., Andrade, D.V., Abe, A.S., 2009. Heat exchange from the toucan bill reveals a controllable vascular thermal radiator. Science 325, 468-470.
Tattersall, G.J., Arnaout, B., Symonds, M.R.E., 2017. The evolution of the avian bill as a thermoregulatory organ. Biol. Rev. 92, 1630-1656.
Wagnon, G.S., Brown, C.R., 2020. Smaller brained cliff swallows are more likely to die during harsh weather. Biol. Lett. 16, 20200264.
Wang, N., Li, J-Q., Liu, Y-Y., Zhang, Z-W., 2010. Improvement on molecular sex identification primers for Passeriform bird species. Chinese Birds 1, 65-69.
Wood, C.W., Brodie III, E.D., 2016. Evolutionary response when selection and genetic variation covary across environments. Ecol. Lett. 19, 1189-1200.
Zang, H., 2009. Temporal and spatial distribution characters of the snowfall with various intensities during 1954-2005 in China. J. Anhui Agr. Sci. 37, 6064-6066.
Zuur, A.F., Ieno, E.N., Elphick, C.S., 2010. A protocol for data exploration to avoid common statistical problems. Method. Ecol. Evol. 1, 3-14.
Table
1.
The sample size for generalized linear mixed model analyses.
Sex
Not survive into the breeding season
Survive into the breeding season
Suffer snowstorm
Not suffer
Suffer snowstorm
Not suffer
Male (N = 163)
30
37
31
65
Female (N = 200)
45
61
32
62
Note: the sample size refers to the number of observations; a total of 25 birds have repeated observations, including 23 birds produced records across two years and the remaining two birds had records across three years.
Table
2.
Model average results from the best model set for the factors (including body length and body mass) that affect the overwinter survival rates of male Silver-throated Tits.
Fixed effects
Analysis using bill length and bill depth
Analysis using bill surface area
Estimate
SE
Z
P
Estimate
SE
z
P
Intercept
3.555
7.053
0.502
0.616
1.870
5.956
0.313
0.755
Age 2a
0.643
0.348
1.833
0.067
0.667
0.346
1.910
0.056
Suffer 1b
2.637
8.296
0.317
0.751
3.771
9.452
0.398
0.691
Bill surface area
−0.414
0.268
1.538
0.124
Bill length
−1.000
0.663
1.497
0.134
Bill depth
−1.228
1.614
0.756
0.450
Head length
−0.225
0.505
0.443
0.658
−0.156
0.511
0.304
0.761
Head length × suffer 1b
−1.311
0.837
1.553
0.120
−1.311
0.837
1.553
0.120
Wing length
0.086
0.110
0.776
0.438
0.096
0.110
0.867
0.386
Tarsus length
0.222
0.393
0.559
0.576
0.262
0.398
0.655
0.513
Tail length
0.046
0.085
0.535
0.593
Body mass
0.012
0.590
0.020
0.984
0.044
0.573
0.076
0.939
Body mass × suffer 1b
−1.373
0.892
1.527
0.127
−1.373
0.892
1.527
0.127
a Age 1 (1-year-old) is considered as the reference level. Age 2 represents all individuals older than one year. b Years without snowstorm are the reference category. 2015 and 2018 are the years that experienced snowstorm, and 2014, 2017 and 2019 are the years that without experienced snowstorm.
Table
3.
Model average results from the best model set for the factors (including body length and body mass) that affect the overwinter survival rates of female Silver-throated Tits.
Fixed effects
Analysis using bill length and bill depth
Analysis using bill surface area
Estimate
SE
z
P
Estimate
SE
z
P
Intercept
18.237
7.231
2.509
0.012
12.398
5.755
2.143
0.032
Age 2a
1.242
0.400
3.086
0.002
1.235
0.401
3.061
0.002
Suffer 1b
−6.787
8.860
0.763
0.445
−4.184
6.344
0.657
0.511
Date
−0.005
0.011
0.436
0.663
−0.007
0.011
0.646
0.518
Bill surface area
−0.663
0.289
2.278
0.023
Bill surface area × suffer 1b
0.554
0.527
1.044
0.296
Bill length
−0.673
0.612
1.094
0.274
Bill depth
−3.515
1.667
2.097
0.036
Bill depth × suffer1b
3.793
2.910
1.295
0.195
Body length
−0.153
0.075
2.031
0.042
−0.155
0.077
2.012
0.044
Body length × suffer 1b
0.160
0.131
1.217
0.223
0.159
0.131
1.211
0.226
Body mass
0.495
0.427
1.152
0.249
0.537
0.424
1.257
0.209
Wing length
0.108
0.119
0.900
0.368
0.114
0.119
0.956
0.339
Tarsus length
0.151
0.351
0.428
0.669
0.285
0.370
0.764
0.445
Tail length
0.039
0.070
0.552
0.581
a Age 1 (1-year-old) is considered as the reference level. Age 2 represents all individuals older than one year. b Years without snowstorm are the reference category. 2015 and 2018 are the years that experienced snowstorm, and 2014, 2017 and 2019 are the years that without experienced snowstorm.
Table
4.
Model average results from the best model set for the factors (including surface area:volume ratio calculated with body length) that affect the overwinter survival rates of male Silver-throated Tits.
Fixed effects
Analysis using bill length and bill depth
Analysis using bill surface area
Estimate
SE
z
P
Estimate
SE
z
P
Intercept
3.640
6.952
0.521
0.602
1.937
5.860
0.329
0.742
Age 2a
0.643
0.348
1.836
0.066
0.669
0.346
1.922
0.055
Suffer 1b
1.914
8.097
0.236
0.814
2.908
9.474
0.306
0.760
Surface area:volume ratio
0.002
0.005
0.465
0.642
0.002
0.005
0.422
0.673
Bill surface area
−0.415
0.268
1.538
0.124
Bill length
−0.998
0.663
1.494
0.135
Bill depth
−1.228
1.614
0.756
0.450
Head length
−0.225
0.505
0.443
0.658
−0.156
0.511
0.304
0.761
Head length × Suffer 1b
−1.311
0.837
1.553
0.120
−1.311
0.837
1.553
0.120
Wing length
0.086
0.109
0.776
0.438
0.096
0.110
0.867
0.386
Tarsus length
0.222
0.393
0.559
0.576
0.262
0.398
0.655
0.513
Tail length
0.046
0.085
0.535
0.593
a Age 1 (1-year-old) is considered as the reference level. Age 2 represents all individuals older than one year. b Years without snowstorm are the reference category. 2015 and 2018 are the years that experienced snowstorm, and 2014, 2017 and 2019 are the years that without experienced snowstorm.
Table
5.
Model average results from the best model set for the factors (including surface area: volume ratio calculated with body length) that affect the overwinter survival rates of female Silver-throated Tits.
Fixed effects
Analysis using bill length and bill depth
Analysis using bill surface area
Estimate
SE
Z
P
Estimate
SE
z
P
Intercept
16.370
6.108
2.666
0.008
10.122
4.385
2.295
0.022
Age 2a
1.196
0.396
3.005
0.003
1.189
0.397
2.977
0.003
Suffer 1b
−6.275
8.644
0.724
0.469
−1.860
3.198
0.579
0.562
Date
−0.005
0.011
0.468
0.640
−0.008
0.011
0.719
0.472
Surface area:volume ratio
−0.010
0.005
2.122
0.034
−0.010
0.004
2.175
0.030
Surface area:volume ratio × Suffer 1b
0.009
0.008
1.123
0.262
0.008
0.008
1.045
0.296
Bill surface area
−0.690
0.284
2.416
0.016
Bill surface area × Suffer 1b
0.513
0.538
0.948
0.343
Bill length
−0.733
0.612
1.189
0.234
Bill depth
−3.650
1.667
2.177
0.029
Bill depth × Suffer 1b
3.642
2.934
1.234
0.217
Wing length
0.046
0.116
0.396
0.692
0.058
0.117
0.489
0.625
Tail length
0.034
0.069
0.488
0.626
Head length
−0.163
0.378
0.430
0.667
a Age 1 (1-year-old) is considered as the reference level. Age 2 represents all individuals older than one year. b Years without snowstorm are the reference category. 2015 and 2018 are the years that experienced snowstorm, and 2014, 2017, 2019 are the years that without experienced snowstorm.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds