Bing-Run Zhu, Mo A. Verhoeven, Chris J. Hassell, Katherine K-S Leung, Dmitry Dorofeev, Qiang Ma, Krairat Eiamampai, Jonathan T. Coleman, Uchrakhzaya Tserenbat, Gankhuyag Purev-Ochir, David Li, Zhengwang Zhang, Theunis Piersma. 2023: Predicting the non-breeding distributions of the two Asian subspecies of Black-tailed Godwit using morphological information. Avian Research, 14(1): 100069. DOI: 10.1016/j.avrs.2022.100069
Citation:
Bing-Run Zhu, Mo A. Verhoeven, Chris J. Hassell, Katherine K-S Leung, Dmitry Dorofeev, Qiang Ma, Krairat Eiamampai, Jonathan T. Coleman, Uchrakhzaya Tserenbat, Gankhuyag Purev-Ochir, David Li, Zhengwang Zhang, Theunis Piersma. 2023: Predicting the non-breeding distributions of the two Asian subspecies of Black-tailed Godwit using morphological information. Avian Research, 14(1): 100069. DOI: 10.1016/j.avrs.2022.100069
Bing-Run Zhu, Mo A. Verhoeven, Chris J. Hassell, Katherine K-S Leung, Dmitry Dorofeev, Qiang Ma, Krairat Eiamampai, Jonathan T. Coleman, Uchrakhzaya Tserenbat, Gankhuyag Purev-Ochir, David Li, Zhengwang Zhang, Theunis Piersma. 2023: Predicting the non-breeding distributions of the two Asian subspecies of Black-tailed Godwit using morphological information. Avian Research, 14(1): 100069. DOI: 10.1016/j.avrs.2022.100069
Citation:
Bing-Run Zhu, Mo A. Verhoeven, Chris J. Hassell, Katherine K-S Leung, Dmitry Dorofeev, Qiang Ma, Krairat Eiamampai, Jonathan T. Coleman, Uchrakhzaya Tserenbat, Gankhuyag Purev-Ochir, David Li, Zhengwang Zhang, Theunis Piersma. 2023: Predicting the non-breeding distributions of the two Asian subspecies of Black-tailed Godwit using morphological information. Avian Research, 14(1): 100069. DOI: 10.1016/j.avrs.2022.100069
Until recently, Limosa limosa melanuroides was thought to be the only subspecies of Black-tailed Godwit in the East Asian–Australasian Flyway. For this reason, all previous occurrences and counts of Black-tailed Godwits in the flyway have been assigned to melanuroides. However, a larger-bodied subspecies, bohaii, has recently been discovered in the flyway. As a result, the occurrence of Black-tailed Godwits in the flyway needs to be reconsidered such that the specific distribution of each subspecies becomes known. To this end, we developed a simple discriminant function to assign individuals to subspecies based on their bill and wing length. Cross-validation with individuals known to be bohaii or melanuroides, based on molecular analysis, showed the developed function to be 97.7% accurate. When applied to measurements of godwits captured at 22 sites across 9 countries in East–Southeast Asia and Australia, we found that bohaii and melanuroides occurred at most sites and overlapped in their distribution from Kamchatka to Australia. We examined photos from all along the flyway to verify this surprising result, confirming that both subspecies co-occur in most locations. Based on these results, we hypothesise that bohaii and melanuroides from the west of their breeding ranges mostly migrate over Chinese mainland. Birds of both subspecies from the east of their ranges are expected to migrate along the Pacific Ocean. We encourage ringing groups in East–Southeast Asia and Australia to use this simple method to keep adding knowledge about Black-tailed Godwits in the East Asian–Australasian Flyway.
Obtaining information on the distribution of different avian subspecies is often challenging due to subtle differences in morphology, plumage colour or songs (Clayton, 1990; Inouye et al., 2001; Wojczulanis-Jakubas et al., 2011). This can be even more complicated for migratory animals that occur at multiple sites throughout the year. Such knowledge on the distribution of subspecies and populations, however, is essential to understand morphological variation, selective pressures, and phylogenetic relationships (Piersma and Baker, 2000; Phillimore and Owens, 2006). When such information is scarce, it also becomes very hard to assess population trends and develop conservation strategies (Chan et al., 2019a; Smith et al., 2020).
This problem is now relevant in the case of Black-tailed Godwits (Limosa limosa), a long-distance migratory shorebird, in the East Asian–Australasian Flyway (EAAF), where recently a new and large-bodied subspecies named bohaii has been discovered (Zhu et al., 2021a). Bohaii godwits breed in the Russian Far East subarctic region and share the EAAF with melanuroides godwits that breed further south in the steppes of Mongolia and Inner Mongolia, China (Zhu et al., 2022). Previously, melanuroides was thought to be the only godwit subspecies in this flyway (Zhu et al., 2021a, b). For this reason, all previous occurrences and counts of Black-tailed Godwits in the EAAF were attributed to melanuroides alone. As a result, there is a real need to update our knowledge on the distribution of both subspecies in the EAAF to make new population estimates, fill knowledge gaps and design conservation strategies.
In recent years, satellite tracking has become a leading method for obtaining the annual distributions of migratory animals (Zhu et al., 2021b; Kuang et al., 2022). It represents a powerful but expensive method and is often limited to a few individuals from one spot rather than including many birds from multiple locations (Lindberg and Walker, 2007). Our study aims to provide the first information on the non-breeding distributions and potential migratory corridors of both godwit subspecies by using morphological information. Morphological measurements of godwits are available from 22 ringing sites across 9 countries in the EAAF. To analyse this data set, we developed a simple discriminant function that can distinguish between the two subspecies on the basis of the individual's bill and wing length.
Discriminant functions have been widely used in species classification (Genovart et al., 2003; Santiago-Alarcon and Parker, 2007; Izenman, 2013). Gunnarsson et al. (2006) demonstrated that this method can also be used to reliably predict the sex of individual islandica godwits. Since the differences between godwit subspecies are more significant than the previously analysed sexual size differences, we expect that a discriminant function will also be able to predict subspecies reliably (Zhu et al., 2020, 2021a). We validated the accuracy of our discriminant function by checking its performance on individuals known to be bohaii or melanuroides through molecular analysis (see Zhu et al., 2021a). Our discriminant function provides an accessible and affordable tool to keep adding knowledge about godwits in the EAAF that will hopefully help design effective conservation strategies, as this flyway continues to face threats from human-caused habitat destruction (Szabo et al., 2016).
2.
Methods
2.1
Discriminant function analysis
To develop our discriminant function, we used reference measurements from Black-tailed Godwits (hereafter 'godwits'), for which the subspecies was genetically determined using a genetic marker from mitochondrial DNA (see Zhu et al., 2021a). Godwits have reversed sexual size dimorphism (Engelmoer and Roselaar, 1998; Zhu et al., 2020). Fortunately, our reference dataset for both subspecies contained a near-equal number of males and females which minimises the potential for bias as a result of the existing sexual dimorphism (bohaii: 21♀ and 24♂; melanuroides: 85♀ and 87♂). Samples of bohaii were collected in Bohai Bay, China (39.2° N, 118.0° E) and melanuroides samples in Roebuck Bay, Australia (17.9° S, 122.3° E). To limit the potential for measurement errors, the same team performed ringing and measuring at each site (led by B.-R. Zhu in Bohai Bay and C.J. Hassell in NW Australia).
These reference measurements were checked for normality with a Shapiro-Wilk Test (p ≥ 0.11), for homogeneity with variance-covariance matrices (χ2 = 93.979, df = 1980, p = 1). The two linear dimensions with the lowest collinearity and variance inflation factor (VIF = 3.1) were selected using R-packages "covTestR" and "car", namely bill and wing (the flattened wing chord). Next, we normalised the reference data to minimise the effect on the model caused by the scale or unit of the predictors and randomly split the reference data into a training set (80%) and a testing set (20%; Picard and Berk, 1990). We then obtained the discriminant function by fitting a linear model to the bill and wing lengths in the training set with the function 'lda' from the R-package "MASS".
We cross-validated the performance of the resulting discriminant function by applying it to the testing set. This method is proven to be more precise than re-substitution or sample splitting (Dechaume-Moncharmont et al., 2011). Lastly, to estimate the error rate of the function when discriminating subspecies, we examined the discriminant function using 50 samples (each sample: n = 217) that were resampled by bootstrapping from the reference data. The resulting prediction accuracy and 95% confidence interval (CI) are reported (Shinmura, 2015).
2.2
Predicting and mapping the non-breeding distributions
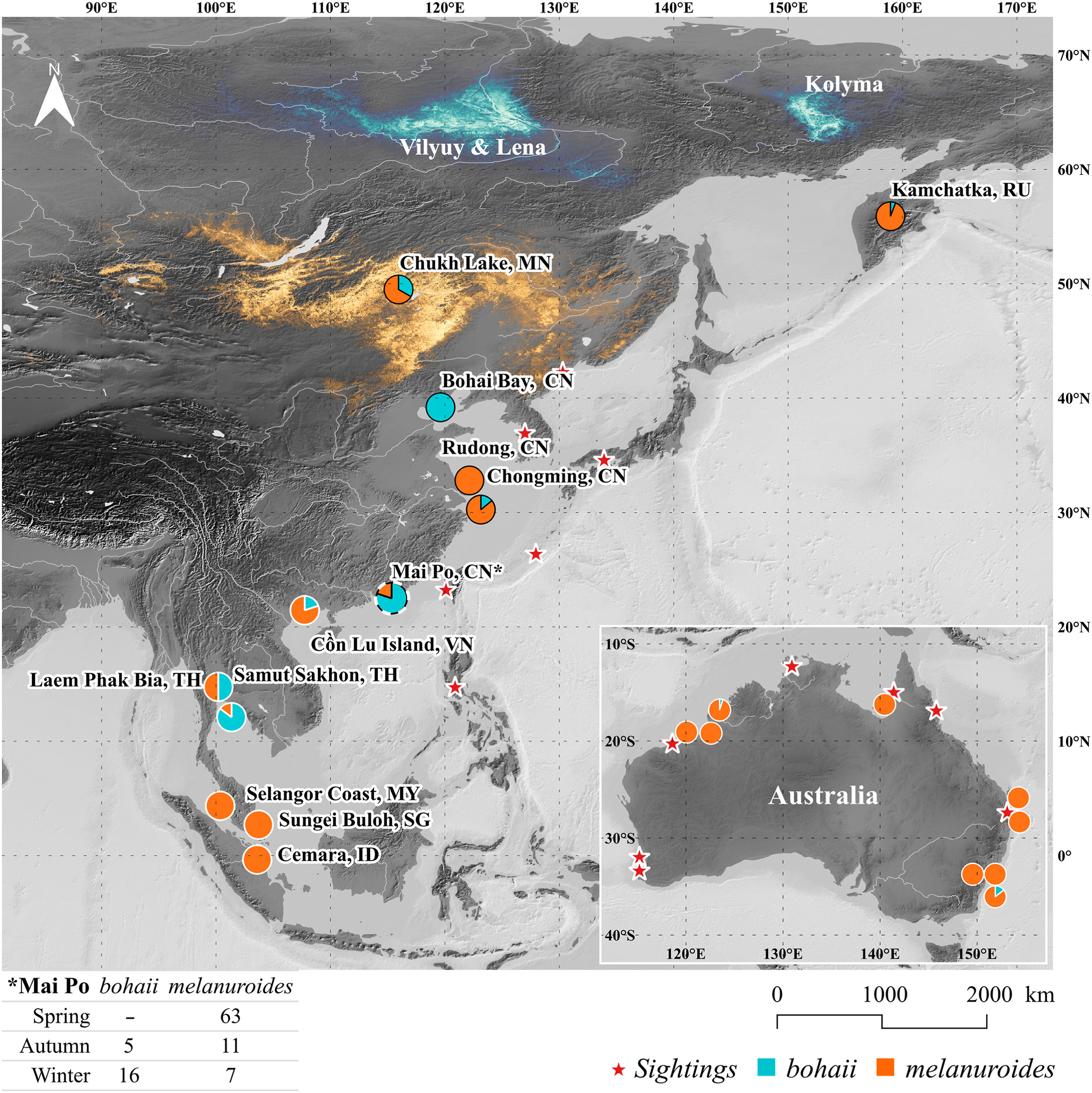
To investigate the non-breeding distribution of both subspecies in East-Southeast Asia and Australia, we applied our discriminant function to the ringing data (e.g., biometric measurements, ringing dates, locations and ring numbers) of 994 adults captured at 22 ringing stations in 9 countries (Fig. 1; Appendix Table S1). We set the western border of our data collection area in Thailand (ca. 95° E) for two reasons: (1) this is the westernmost site where both subspecies have been confirmed to occur through molecular methods (Zhu et al., 2016–2018 unpubl. data, 2021a); (2) at sites further west, e.g., in Bangladesh, India and Sri Lanka, there is anecdotal evidence suggesting the occurrence of L. l. limosa, which is similar in body size to bohaii and would therefore influence our non-breeding range predictions (Zhu et al., 2020). We classified each ringing site as a stopping or non-breeding site by the month in which the ringing work was conducted, i.e., spring (April–June), autumn (July–October) and winter (November–March; Fig. 1).
Figure
1.
Predicted non-breeding distributions of bohaii and melanuroides. Sightings of bohaii type godwits confirmed by photos are shown with stars. Stopping sites are marked with black outlines, and non-breeding sites are white. Mai Po Nature Reserve (black and white outline) was the northernmost non-breeding site for both subspecies and also a stopping site for melanuroides. The known breeding range of bohaii (ca. 61–66° N) is shown in light blue and that of melanuroides (ca. 42–52° N) in orange (adapted from Zhu et al., 2022). Ring stations and countries are in bold.
If a bird was expected to belong to the bohaii subspecies, but the ringing site was not previously known as a bohaii-site from satellite tracking, i.e., Russia or Australia (see Zhu et al., 2021b). We performed additional checks for the possibility of their occurrence by examining photos of godwits taken in those regions. Godwit photos from Kamchatka, Russia, were collected from the local ringing group, and photos of non-breeding godwits in Australia were collected from eBird (https://ebird.org/home) and individual researchers. The godwit photos were assessed by B.-R. Zhu and K.K.-S. Leung who have the most experience with identifying bohaii in the field. We first shuffled the photos and concealed their locations to reduce potential bias. Then, both examiners verified the photos independently and their results were cross-checked and turned out to be identical. As a result, 75 photos were examined (Fig. 1).
We collected additional information about the presence of Black-tailed Godwits in regions beyond our sampling sites, i.e., the Korean Peninsula, Japan, Taiwan of China, the Philippines and Australia by interviewing bird watchers, photographers and shorebird researchers along the flyway. When the presence of godwits was reported, we also collected and examined photos from these sites. We present the predicted proportion of each subspecies in the ringing stations on a map using Qgis 3.1.6, sightings of godwits from the interviews were also marked out.
3.
Results
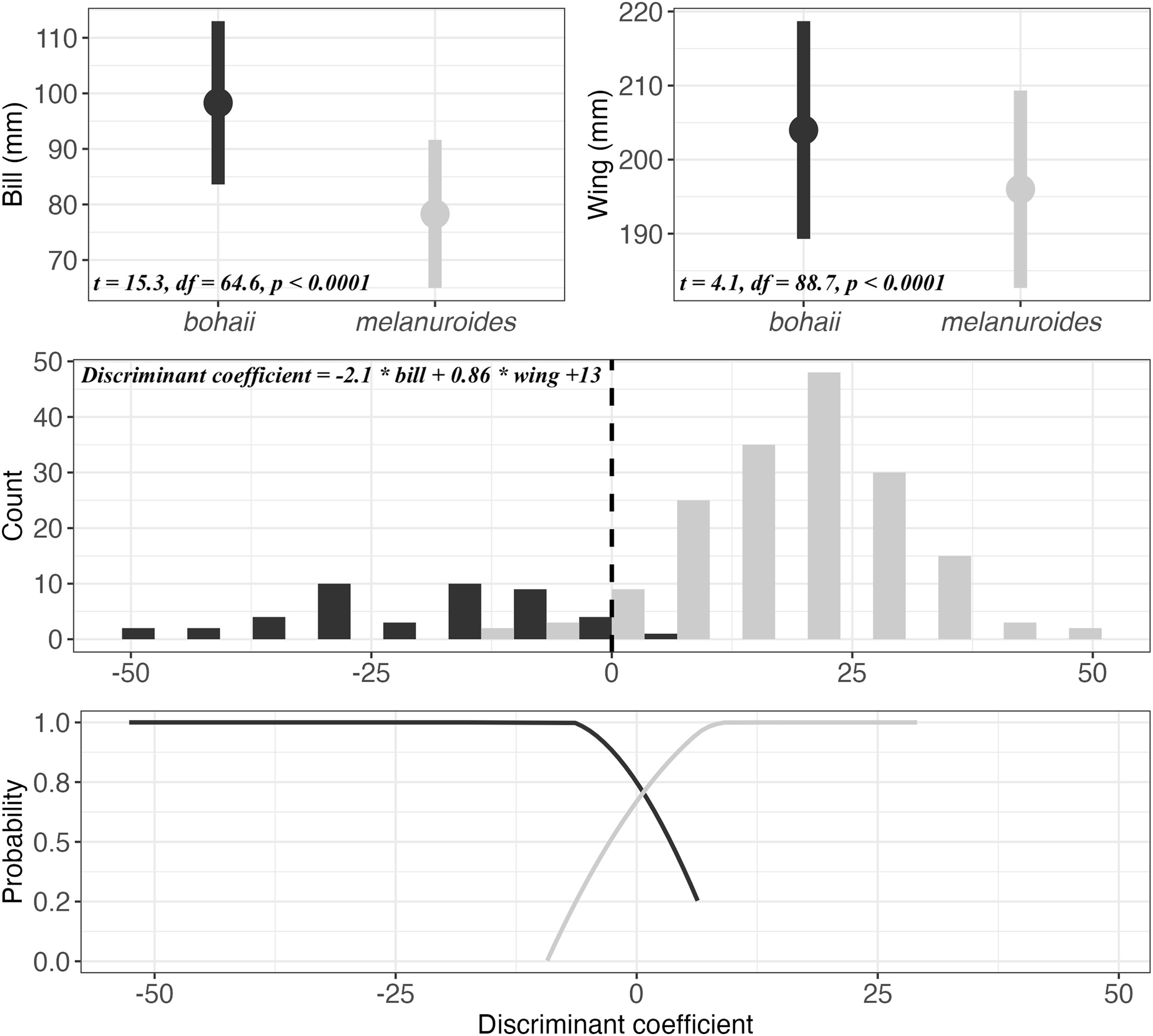
The discriminant analysis yielded the following function with a cut-off value of 13 (also see Fig. 2):
Figure
2.
(A) average bill and wing length (95% CI) of bohaii and melanuroides. The results of the Student t-tests are displayed in the bottom left. (B) subspecies assignment of all individuals from our reference dataset based on the discriminant function (top left; black = melanuroides, grey = bohaii). The dashed line indicates the cut-off value (13). (C) the probability of correct subspecies prediction at different discriminant coefficients. The sample size for all figures: bohaii: 45, melanuroides: 172.
An individual is predicted to be bohaii when the discriminant coefficient is less than 0 and melanuroides when it is greater than 0. The cross-validation of this function on our testing set (n = 44) showed it to be 97.7% accurate in predicting subspecies. The prediction accuracy of the discriminant function on the bootstrapped samples was 97.0% (95% CI: 95.7–98.3%, n = 2357) for bohaii and 95.5% (95% CI: 95–96.1%, n = 8493) for melanuroides.
Out of the 22 ringing stations that provided biometric data on godwits, Mai Po Nature Reserve in Hong Kong, China (ca. 22° N) was the northernmost non-breeding site of both subspecies (Fig. 1; Appendix Table S1). The morphological data showed that bohaii was captured at nine ringing stations in six countries and that melanuroides was captured at all ringing stations (Fig. 1; Appendix Table S1). Moreover, the distribution of the two subspecies overlapped in six countries, i.e., Russia, Mongolia, China, Vietnam, Thailand and Australia (Fig. 1; Appendix Table S1).
The discriminant function identified bohaii occurred at sites not previously known from satellite tracking: Kamchatka, Russia, Nudgee Beach, Australia and Roebuck Bay, Australia (Fig. 1; Appendix Table S1; Zhu et al., 2021b). In Kamchatka, 6% of the measured birds (four in total) were identified as bohaii. This was 14% in Nudgee Beach (three birds) and 5% in Roebuck Bay (28 birds). These proportions are higher than the model's average error rate (3%) in discriminating bohaii and thus support the presence of bohaii at these new locations. To verify this finding, we examined photos of one of the four godwits predicted to be bohaii in Kamchatka and both examiners agreed that the individual was bohaii. Similarly, we also identified numerous bohaii type individuals in photos from near Nudgee Beach and Roebuck Bay (see Appendix Table S2 for details).
Additionally, amongst photos from bird watchers, photographers and researchers, we found bohaii type godwits together with melanuroides type godwits in Rason (North Korea, N. Moores, pers. comm.), Chungcheongnam-do (South Korea, H–K. Nam, pers. comm.), Okinawa and Hiroshima (Japan, T. Kawasumi pers. comm.), Tainan (China, B. Chen, pers. comm.), Bulacan (the Philippines, I. Dy, pers. comm.) and the Gulf of Carpentaria, Australia (R. Jaensch, pers. comm. Fig. 1).
4.
Discussion
We developed a discriminant function based on the bill and wing length, which was > 95% accurate in assigning godwits captured throughout the EAAF to either the bohaii or melanuroides subspecies. It is worth noting that the difference in body size between the analysed godwit subspecies is greater than the sexual dimorphism within the subspecies (Zhu et al., 2020, 2021a). Therefore, even without considering an individual's sex, the discriminant function was able to assign it to subspecies with high accuracy. We applied this function to godwits captured at 22 sites across 9 countries in East-Southeast Asia and Australia. We found that bohaii and melanuroides co-occurred in most sampled sites except for the stopping site in Rudong, China and the non-breeding sites in Malaysia, Singapore, and Indonesia, where only melanuroides godwits were captured (Fig. 1). The overlap in the distribution of bohaii and melanuroides extends as far south as Australia. Previously, only the latter was thought to occur here (Fig. 1; van Gils et al., 2020).
The discriminant function revealed that bohaii has stopping sites in Chinese mainland and non-breeding sites in Thailand. This is consistent with previous satellite tracking results (Zhu et al., 2021b) and supports the subspecies assignment based on our function. Surprisingly, the discriminant function also uncovered some godwits captured in Kamchatka, Russia and Australia to be bohaii. These sites are outside of bohaii's previously known distribution. Still, the number of godwits assigned as bohaii in these sites was higher than expected on the basis of the function's error rate. In addition, our examination of photos from these regions also confirmed the occurrence of bohaii here. Therefore, we conclude that the actual distribution of bohaii is larger than previously identified via tracking (Zhu et al., 2021b). This is further supported by photos from the Korean peninsula, Taiwan of China and the Philippines, which also include bohaii in addition to melanuroides. Noteworthy is that photos from the southeastern Gulf of Carpentaria, Australia even indicated that bohaii is the dominant non-breeding population there (R. Jaensch, pers. comm.).
Based on these results, we hypothesise that bohaii from the west of their breeding range (e.g., the Lena and Vilyuy River basins, see Fig. 1 and Zhu et al., 2022) predominantly migrate over Chinese mainland. In contrast, bohaii from the east of their range (e.g., the Kolyma River basin) are expected to predominantly migrate along the Pacific Ocean via the Kamchatka Peninsula, Japan, Korean Peninsula, Taiwan of China and the Philippines to Australia (Fig. 1). We believe that the same is true for melanuroides, i.e., an inland corridor is mainly used to reach their western range and an oceanic corridor is used to reach their eastern range (see Fig. 1 and Zhu et al., 2022). This idea is supported by two satellite tagged adult melanuroides godwits from the west of their range in Chukh Lake, Mongolia. They migrated from inland China, over the Himalayas, and overwintered on the Indian subcontinent (Bangladesh and Sri Lanka; Tserenbat and Purev-Ochir, 2019–2020, unpubl. data). There is no published information about the eastern breeding locations of melanuroides that use the Kamchatka Peninsula as a stopping site. However, they are likely to breed in Chukotka or on islands in the Bering Strait (Lappo et al., 2012). Furthermore, molecularly determined melanuroides from Northwest Australia, were sighted in Taiwan, Lianyungang and Kamchatka (Chan et al., 2019b; Hassell et al., 1998–2019 unpubl. data), suggesting an oceanic migratory corridor for the eastern breeding population of this subspecies.
While the geographical and ecological characteristics of the breeding grounds of these two Asian godwits subspecies are considerably different (Zhu et al., 2022), their non-breeding distributions overlap to a large extent. This pattern is also seen in islandica and limosa in the East-Atlantic Flyway (Gill et al., 2007). Outside their core breeding grounds in Iceland and the Netherlands, the two subspecies co-occur from the UK to Morocco during the non-breeding season (Lopes et al., 2013; Verhoeven et al., 2021). That said, islandica is primarily found in intertidal habitats while limosa mostly uses rice fields (Alves et al., 2010). The same might be the case for bohaii and melanuroides in Southeast Asia. For example, in Thailand, tracked bohaii rarely used saline habitats. In contrast, large groups of melanuroides type godwits were recorded on the Samut Prakan coast near Bangkok (Zhu et al., 2016–2018 unpubl. data, 2021b).
We encourage ringing groups in East-Southeast Asia and Australia to use this accessible and effective method to examine their data and contribute to our collective knowledge of Black-tailed Godwits, such as their non-breeding range. However, we caution against using this method when subspecies of similar body sizes co-occur, e.g., in Myanmar, Bangladesh, India and Sri Lanka, where limosa and bohaii may co-exist, we recommend using the genetic markers already developed and applied in four godwit subspecies to assess population structure (Lopes et al., 2013; Zhu et al., 2021b).
Authors' contribution
ZBR designed this study, analysed the data, wrote and reviewed the manuscript; MAV analysed the data and reviewed the manuscript; CJH, KKSL, DD, QM, KE, JTC, UT, GPO, DL collected the data in the field and reviewed the manuscript. ZZ, TP reviewed the manuscript. All authors read and approved the final manuscript.
Ethics statement
The local forestry or nature reserve authorities approved the ringing work in Chinese mainland. The Agriculture, Fisheries and Conservation Department of the Government of the Hong Kong SAR approved the ringing work at Mai Po Nature Reserve, China. The Federal Service for Supervision of Natural Resources (Rosprirodnadzor) approved the ringing work in Kamchatka, Russia. The Ministry of Environment and Tourism of Mongolia approved the work in Mongolia. The National Parks Board, Singapore, authorised the ringing work in Singapore. The Department of National Park, Wildlife and Plant Conservation, Ministry of Natural Resource and Environment, approved the ringing work in Thailand. The Australian Federal Government, the Australian Bird and Bat Banding Scheme (ABBBS) approved the ringing work in Australia.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We are grateful to the colleagues along the East Asian-Australasian Flyway, for providing sightings and ringing information on Black-tailed Godwits. These include Qingquan Bai and Qing Chang from China, Preeyaporn Kala, Jirut Khamaye and Philip D. Round from Thailand, Iwan Londo from Indonesia, Bisharga Das and Sayam Chowdhury from Bangladesh, David Wells from Malaysia, and Leo Joseph from the Australian National Wildlife Collection, CSIRO (ror.org/059mabc80; grid.510155.5). We acknowledge the Australasian Wader Studies Group for providing ringing data along the EAAF. We acknowledge the Queensland Wader Study Group for providing the ringing data for Queensland, Australia. We acknowledge the Yawuru People via the offices of Nyamba Buru Yawuru Limited for permission to catch birds on the shores of Roebuck Bay, traditional lands of the Yawuru people. We also acknowledge the Karajarri and Nyangumarta people for permission to catch birds to be marked for this project on the shores of 80 Mile Beach, the traditional lands of the Karajarri and Nyangumarta people. We appreciate the many anonymous photographers for their photos of Black-tailed Godwits in Japan and Australia. CJH would like to thank GFN's major funders for the NWA work, Spinoza Premium of Netherlands Organisation Prize for Scientific Research to Theunis Piersma (2014–2017), WWF Netherlands (2010–2017 and 2019–2021) and MAVA, Foundation pour la nature (2018). This study was funded by the National Natural Science Foundation of China [31830089, 31801985, 32270518].
Alves, J.A., Lourenco, P.M., Piersma, T., Sutherland, W.J., Gill, J.A., 2010. Population overlap and habitat segregation in wintering Black-tailed Godwits Limosa limosa. Bird Study 57, 381-391.
Chan, Y.C., Tibbitts, T.L., Lok, T., Hassell, C.J., Peng, H.B., Ma, Z., et al., 2019a. Filling knowledge gaps in a threatened shorebird flyway through satellite tracking. J. Appl. Ecol. 56, 2305-2315.
Chan, Y.C., Peng, H.B., Han, Y.X., Chung, S.S., Li, J., Zhang, L., et al., 2019b. Conserving unprotected important coastal habitats in the Yellow Sea: shorebird occurrence, distribution and food resources at Lianyungang. Global Ecol. Conserv. 20, e00724.
Clayton, N.S., 1990. Subspecies recognition and song learning in zebra finches. Anim. Behav. 40, 1009-1017.
Dechaume-Moncharmont, F.X., Monceau, K., Cezilly, F., 2011. Sexing birds using discriminant function analysis: a critical appraisal. Auk 128, 78-86.
Engelmoer, M., Roselaar, C.S., 1998. Geographical Variation in Waders. Springer Science and Business Media, Dordrecht.
Genovart, M., McMinn, M., Bowler, D., 2003. A discriminant function for predicting sex in the Balearic Shearwater. Waterbirds 26, 72-76.
Gill, J.A., Langston, R.H., Alves, J.A., Atkinson, P.W., Bocher, P., Vieira, N.C., et al., 2007. Contrasting trends in two Black-tailed Godwit populations: a review of causes and recommendations. Wader Study Group Bull. 114, 43-50.
Gunnarsson, T.G., Gill, J.A., Goodacre, S.L., Gélinaud, G., Atkinson, P.W., Hewitt, G.M., et al., 2006. Sexing of Black-tailed Godwits Limosa limosa islandica: a comparison of behavioural, molecular, biometric and field-based techniques. Bird Study 53, 193-198.
Inouye, C.Y., Hill, G.E., Stradi, R.D., Montgomerie, R., Bosque, C., 2001. Carotenoid pigments in male house finch plumage in relation to age, subspecies, and ornamental coloration. Auk 118, 900-915.
Izenman, A.J., 2013. Linear Discriminant Analysis. Modern Multivariate Statistical Techniques. Springer, New York, NY.
Kuang, F., Wu, W., Li, D., Hassell, C.J., Maglio, G., Leung, K. K-S., et al., 2022. Detecting the non-breeding region and migration route of Whimbrels (Numenius phaeopus rogachevae) in the East Asian-Australasian Flyway. Avian Res. 13, 100011.
Lappo, E., Tomkovich, P., Syroechkovskiy, E., Zharikov, Y.V., 2012. Atlas of Breeding Waders in the Russian Arctic. UF Ofsetnaya Pecha, Moscow.
Lindberg, M.S., Walker, J., 2007. Satellite telemetry in avian research and management: sample size considerations. J. Wildlife Manage. 71, 1002-1009.
Lopes, R.J., Alves, J.A., Gill, J.A., Gunnarsson, T.G., Hooijmeijer, J.C., Lourenço, P.M., et al., 2013. Do different subspecies of Black-tailed Godwit Limosa limosa overlap in Iberian wintering and staging areas? Validation with genetic markers. J. Ornithol. 154, 35-40.
Phillimore, A.B., Owens, I.P., 2006. Are subspecies useful in evolutionary and conservation biology? Proc. R. Soc. B. Biol. Sci. 273, 1049-1053.
Piersma, T., Baker, A.J., 2000. Life history characteristics and the conservation of migratory shorebirds. In: Gosling, L.M., Sutherland, W.J. (Eds.), Behaviour and Conservation. Cambridge University Press, Cambridge, pp. 105–124.
Santiago-Alarcon, D., Parker, P.G., 2007. Sexual size dimorphism and morphological evidence supporting the recognition of two subspecies in the Galápagos Dove. Condor 109, 132-141.
Shinmura, S., 2015. The 95% confidence intervals of error rates and discriminant coefficients. Stat. Optim. Inform. Comp. 3, 66-78.
Smith, P.A., McKinnon, L., Meltofte, H., Lanctot, R.B., Fox, A.D., Leafloor, J.O., et al., 2020. Status and trends of tundra birds across the circumpolar Arctic. Ambio 49, 732-748.
Szabo, J.K., Battley, P.F., Buchanan, K.L., Rogers, D.I., 2016. What does the future hold for shorebirds in the East Asian-Australasian Flyway? Emu 116, 95-99.
van Gils, J.A., Wiersma, J.P., Christie, D.A., Garcia, E.F., Boesman, P.F., 2020. Black-tailed godwit (Limosa limosa), version 1.0. In: del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E. (Eds.), Birds of the World. Cornell Lab of Ornithology, Ithaca, NY.
Verhoeven, M.A., Smart, J., Kitchin, C., Schmitt, S., Whiffin, M., Burgess, M., et al., 2021. Diagnosing the recent population decline of Black-tailed Godwits in the United Kingdom. Wader Study 128, 65-76.
Wojczulanis-Jakubas, K., Jakubas, D., Welcker, J., Harding, A., Karnovsky, N.J., Kidawa, D., et al., 2011. Body size variation of a high-Arctic seabird: the dovekie (Alle alle). Polar Biol. 34, 847-854.
Zhu, B.R., Hassell, C.J., Verkuil, Y.I., Gunnarson, T.G., Hooijmeijer, J.C., Zhang, Z., et al., 2020. Size, shape and sex differences in three subspecies of Black-tailed Godwits Limosa limosa. Bird Study 67, 45-52.
Zhu, B.R., Verkuil, Y.I., Conklin, J.R., Yang, A., Lei, W., Alves, J.A., et al., 2021a. Discovery of a morphologically and genetically distinct population of Black-tailed Godwits in the East Asian-Australasian Flyway. Ibis 163, 448-462.
Zhu, B.R., Verhoeven, M.A., Loonstra, A.J., Sanchez-Aguilar, L., Hassell, C.J., Leung, K. K-S., et al., 2021b. Identification of breeding grounds and annual routines of the newly discovered bohaii subspecies of Black-tailed Godwits. Emu 121, 292-302.
Zhu, B.R., Verhoeven, M.A., Velasco, N., Sanchez-Aguilar, L., Zhang, Z., Piersma, T., 2022. Current breeding distributions and predicted range shifts under climate change in two subspecies of Black-tailed Godwits in Asia. Global Change Biol. 28, 5416-5426.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds