Siwei An, Fenliang Kuang, Wei Wu, Chris J. Hassell, Jonathan T. Coleman, Zijing Gao, Xuena Sun, Yue Yuan, Grace Maglio, Kar-Sin K. Leung, Xuesong Feng, Zhijun Ma. 2024: Individual consistency in spatiotemporal characteristics of migratory Whimbrels in the East Asian–Australasian Flyway. Avian Research, 15(1): 100191. DOI: 10.1016/j.avrs.2024.100191
Citation:
Siwei An, Fenliang Kuang, Wei Wu, Chris J. Hassell, Jonathan T. Coleman, Zijing Gao, Xuena Sun, Yue Yuan, Grace Maglio, Kar-Sin K. Leung, Xuesong Feng, Zhijun Ma. 2024: Individual consistency in spatiotemporal characteristics of migratory Whimbrels in the East Asian–Australasian Flyway. Avian Research, 15(1): 100191. DOI: 10.1016/j.avrs.2024.100191
Siwei An, Fenliang Kuang, Wei Wu, Chris J. Hassell, Jonathan T. Coleman, Zijing Gao, Xuena Sun, Yue Yuan, Grace Maglio, Kar-Sin K. Leung, Xuesong Feng, Zhijun Ma. 2024: Individual consistency in spatiotemporal characteristics of migratory Whimbrels in the East Asian–Australasian Flyway. Avian Research, 15(1): 100191. DOI: 10.1016/j.avrs.2024.100191
Citation:
Siwei An, Fenliang Kuang, Wei Wu, Chris J. Hassell, Jonathan T. Coleman, Zijing Gao, Xuena Sun, Yue Yuan, Grace Maglio, Kar-Sin K. Leung, Xuesong Feng, Zhijun Ma. 2024: Individual consistency in spatiotemporal characteristics of migratory Whimbrels in the East Asian–Australasian Flyway. Avian Research, 15(1): 100191. DOI: 10.1016/j.avrs.2024.100191
Ministry of Education Key Laboratory for Biodiversity Science and Ecological Engineering, Coastal Ecosystems Research Station of the Yangtze River Estuary, Institute of Biodiversity Science, School of Life Sciences, Fudan University, Shanghai, 200433, China
b.
School of Chemistry and Environment, Yunnan Minzu University, Kunming, 650504, China
c.
Shanghai Chongming Dongtan Nature Reserve Administration Center, Chongming, Shanghai, 202183, China
d.
Global Flyway Network, PO Box 3089, Broome, WA 6725, Australia
e.
Australasian Wader Studies Group, Broome, WA, 6725, Australia
f.
Queensland Wader Study Group, Shailer Park, QLD, 4128, Australia
g.
Hong Kong Waterbirds Ringing Group, Hong Kong, 999077, China
Funds:
the National Key Research and Development Program of China2023YFF1304504
the National Natural Science Foundation of China31830089
the National Natural Science Foundation of China31772467
the Science and Technology Department of Shanghai21DZ1201902
the World Wide Fund for Nature Beijing Office10003881
Many migratory birds exhibit interannual consistency in migration schedules, routes and stopover sites. Detecting the interannual consistency in spatiotemporal characteristics helps understand the maintenance of migration and enables the implementation of targeted conservation measures. We tracked the migration of Whimbrel (Numenius phaeopus) in the East Asian–Australasian Flyway and collected spatiotemporal data from individuals that were tracked for at least two years. Wilcoxon non-parametric tests were used to compare the interannual variations in the dates of departure from and arrival at breeding/nonbreeding sites, and the interannual variation in the longitudes when the same individual across the same latitudes. Whimbrels exhibited a high degree of consistency in the use of breeding, nonbreeding, and stopover sites between years. The variation of arrival dates at nonbreeding sites was significantly larger than that of the departure dates from nonbreeding and breeding sites. Repeatedly used stopover sites by the same individuals in multiple years were concentrated in the Yellow Sea coast during northward migration, but were more widespread during southward migration. The stopover duration at repeatedly used sites was significantly longer than that at sites used only once. When flying across the Yellow Sea, Whimbrels breeding in Sakha (Yakutia) exhibited the highest consistency in migration routes in both autumn and spring. Moreover, the consistency in migration routes of Yakutia breeding birds was generally higher than that of birds breeding in Chukotka. Our results suggest that the northward migration schedule of the Whimbrels is mainly controlled by endogenous factors, while the southward migration schedule is less affected by endogenous factors. The repeated use of stopover sites in the Yellow Sea coast suggests this region is important for the migration of Whimbrel, and thus has high conservation value.
Migratory birds regularly move between breeding and nonbreeding sites every year (Newton, 2008). Migration is endogenously regulated to match periodical changes of environmental conditions. Consequently, the spatiotemporal characteristics of migration activities is predictable (Akesson et al., 2017; Akesson and Helm, 2020). Many studies have shown that the migration activities of the same individual usually have a fixed schedule and route between years and show high temporal and spatial consistency (Potti, 1998; Conklin et al., 2013; Vardanis et al., 2016; Kurten et al., 2022). Through long-term evolution, migratory birds follow schedules and routes that have been proved to be suitable, which help birds efficiently use familiar habitats and food resources along the migration route, save time and energy in exploring new environments, and reduce the risk of predation (Lank et al., 2003; Duijns et al., 2019). Migration consistency therefore plays an important role in ensuring successful migration and improving fitness of migratory birds.
The interannual spatiotemporal consistency of migration helps birds adapt to periodical environmental conditions. Meanwhile, birds can also fine tune the spatiotemporal rhythm of migration to a certain extent in response to altered environmental conditions (Weller et al., 2022). For example, wind conditions largely affect the migration decision of birds at stopover sites (Ma et al., 2011; Schmaljohann et al., 2022); when facing food decline at traditional stopover sites, some individuals can shift to use different stopover sites (Ke et al., 2019), or even change migration routes (Verkuil et al., 2012). Under the current context of global climate change, the ability to adjust migration schedules and routes should benefit birds in adapting to altered environmental conditions. Therefore, clarifying the spatiotemporal consistency of migration activities helps understand their migration ecology and contributes to the protection of migratory birds and their key habitats.
Most studies on the spatiotemporal consistency of migration have focused on the start and end date of migration, while the results varied among studies. Some studies have found that the consistency of northward migration schedules is generally higher than that of southward migration schedules, such as Bar-tailed Godwit (Limosa lapponica) in the East Asian–Australasian Flyway (Conklin et al., 2013), Purple Martin (Progne subis) in the American Flyway (Fraser et al., 2019), and Pallas's Gull (Larus ichthyaetus) breeding in the Tibetan Plateau (Liu et al., 2018). This may be due to that the northward migration schedule is mainly controlled by endogenous regulation while the southward migration schedule may also be affected by reproductive success in the adults as well as by social and environmental conditions (Lourenco et al., 2011; Stanley et al., 2012; Pedersen et al., 2018; Akesson and Helm, 2020; Loonstra et al., 2023). Moreover, many birds do not have territorial behavior in nonbreeding sites, and the southward migration schedule is relatively flexible without reproductive pressure (Carneiro et al., 2019; Fraser et al., 2019). However, some species, such as the Hoopoes (Upupa epops) breeding in Switzerland have higher consistency in southward migration schedules than northward migration schedules (van Wijk et al., 2016).
In addition, the consistency of the start dates of migration is usually higher than that of the arrival dates, especially in northward migration, such as for the Red-backed Shrike (Lanius collurio) (Pedersen et al., 2018) and Western Marsh Harrier (Circus aeruginosus) (Vardanis et al., 2011). This may be due to the departure dates being regulated by endogenous factors more (Lourenco et al., 2011; Pedersen et al., 2018; Watson and Keren, 2019). However, the consistency of arrival dates at breeding sites of Snow Goose (Anser caerulescens) in North America (Bety et al., 2004) and Woodcock (Scolopax rusticola) in Europe (Tedeschi et al., 2020) is higher than that of their departure dates from nonbreeding sites. This may be caused by the time pressure of arrival at breeding sites and the high consistency of migration routes.
There is a hypothesis that for long-distance migratory birds, environmental conditions at breeding sites are difficult to predict when birds are at distant nonbreeding sites. Thus, the date of departure from nonbreeding sites might rely mainly on endogenous regulation and exhibit high consistency in departure dates between years (Ruthrauff et al., 2019). Moreover, many migratory birds use the same breeding and nonbreeding sites in successive years, while there are relatively few studies on the spatial consistency of stopover sites and migration routes. Some studies have found that Great Reed Warbler (Acrocephalus arundinaceus) (Hasselquist et al., 2017), Osprey (Pandion haliaetus) (Vardanis et al., 2016), and Oriental Honey-buzzard (Pernis ptilorhynchus) (Sugasawa and Higuchi, 2019), use the same stopover sites or similar migration routes in different years. These are likely due to their reliance on specific habitats or food sources that are distributed at specific spatial regions. The consistency of migration routes can also be affected by topographic features. For example, Common Swift (Apus apus) breeding in Europe exhibit high consistency in migration routes crossing the Strait of Gibraltar (Wellbrock et al., 2017); Oriental Honey-buzzard breeding in Japan exhibits less consistency in migration routes when crossing oceans than flying over land (Sugasawa and Higuchi, 2019).
Many shorebird species fly tens of thousands of kilometers during one-way migration, across multiple landscapes including land and ocean. Some of them highly rely on tidal flats during stopover. Therefore, shorebirds can be good objects to study on the spatiotemporal consistency in annual cycles. Moreover, many shorebirds have suffered rapid population decline over the past two decades, mainly due to habitat loss and degradation at key stopover sites (Baker et al., 2004; Studds et al., 2017). Detecting the consistency during annual movement helps identify the regions of priority conservation along migration routes. Here we analyzed the spatiotemporal consistency of migration activities for Whimbrels in the East Asian–Australasian Flyway (EAAF). The Whimbrels are long-distance migratory birds with a worldwide distribution. In the EAAF, their breeding sites are scattered across Siberia, and the nonbreeding sites are mainly located in Southeast Asia and Australia (Kuang et al., 2022). During the migration period, they cross East Asia and mainly stopover on the tidal flats of the Yellow Sea region (Kuang et al., 2020).
We tracked the migration of Whimbrels and analyzed: a) the consistency of the timing of arrival at and departure from the breeding/nonbreeding sites; b) the spatial consistency of the breeding, nonbreeding, and stopover sites; and c) the spatial consistency of migration routes at different latitudes, seasons and regions. We hypothesized that Whimbrels exhibit high consistency in migration schedule between years, and the date of departure from nonbreeding sites has the highest consistency. In terms of spatial consistency, since Whimbrels mainly stopover in the Yellow Sea region (Kuang et al., 2020), we hypothesized that stopovers and migration routes have high consistency, and the consistency is higher in the Yellow Sea than in other regions. Due to the wide distribution of the breeding sites of Whimbrels in the EAAF, different individuals either use migration routes along the coast, over the land, or across the Pacific (Kuang et al., 2022). We hypothesized that the route consistency of individuals migrating across the Pacific would be lower than those over land due to variable wind conditions and the lack of stopovers and landmarks.
2.
Materials and methods
2.1
Bird tracking
The tracking data in this study came from Whimbrels captured at Shanghai Chongming Dongtan, China, a stopover site in the southern Yellow Sea, in 2018–2019, and at the nonbreeding sites in Australia at Moreton Bay, Queensland and Roebuck Bay, western Australia. Whimbrels captured in Moreton Bay in 2017–2018 were fitted with 5 g solar platform terminal transmitters (PTTs, Microwave Telemetry, Inc., Columbia, Maryland, USA). The Whimbrels captured in Roebuck Bay were fitted with 5 g solar PTTs or 7 g solar Global Positioning System–Global System for Mobile Communication transmitters (GPS–GSMs, Hunan Global Messenger Technology Co., Ltd., Hunan, China). All Whimbrels captured at Shanghai Chongming Dongtan were deployed with 7 g GPS–GSM transmitters.
The PTTs and GPS-GSM transmitters were attached to the back of Whimbrels using the leg-loop harness. All the tagged birds in this study were identified as adults during time of deployment based on plumage characteristics (Prater et al., 1977). The weight of each transmitter was less than 3% of the body weight of each Whimbrel, to minimise impact on the birds (Kuang et al., 2020). The PPTs working cycle was set to be transmitting for 10 h and then shut down for 48 h. The GPS–GSM transmitters were set to record one geographical coordination every 2–6 h. See Kuang et al. (2020, 2022) for more detailed information.
2.2
Data processing
We used data from individuals that were tracked for more than one year. Only the data with high positioning accuracy (error less than 500 m for PTTs and less than 100 m for GPS–GSM transmitters) were included for analysis. The spatiotemporal information of migration was extracted using the method in Kuang et al. (2020). Briefly, during northward migration, we assumed that an individual has departed from the nonbreeding or stopover site when it flew northward for more than 50 km from the site and did not return; an individual has arrived at the breeding site when it no longer moved northward and the two successive location fixes were less than 50 km apart. If an individual stayed at multiple sites at breeding region during breeding season, we considered the site at which the individual stayed for the longest duration to be the breeding site. Similarly, during southward migration, we assumed an individual had departed from the breeding or stopover site when it flew southward for more than 50 km from the site and did not return; an individual had arrived at the nonbreeding site when it no longer moved southward and the two successive location fixes were less than 50 km apart. During migration, the sites where Whimbrels stayed for more than 48 h and where two successive location fixes were less than 50 km apart were considered to be stopover sites. Considering that individual fixes were extremely concentrated at each site, the geographical locations of nonbreeding, stopover, and breeding sites were measured by using the arithmetic mean of the geographical coordinates of all fixes at the sites for each individual (Giunchi et al., 2019).
The dates of departure from and arrival at the nonbreeding, stopover, and breeding sites were estimated for each individual. If two consecutive location fixes before/after departure from or arrival at a site were recorded on the same day, the dates were used as the departure or arrival date. If two consecutive location fixes before/after departure from or arrival a site were recorded on different dates and if an individual was flying at the latter location fix, we measured the distance between the location fixes and used a flight speed of 50 km/h to estimate the flight duration for the individual to cover the distance (Alves et al., 2016). For the individuals fitted with PTTs, we estimated the status of birds (flying or not flying) according to the time and locations of consecutive fixes before/after departure from or arrival at a site. The departure date was estimated according to the time recorded at the last location fix before departure and the flight duration after departure. Similarly, if two consecutive location fixes before and after arrival at a site fall on different dates and the individual was flying at the earlier fix, the distance between the location fixes and the flight speed of 50 km/h was used to estimate the flight duration and then the arrival date was calculated. The duration of stopover at a site was calculated according to the arrival and departure date at that site.
To analyze the consistency of migration routes between years, we identified the longitudes on which Whimbrels crossed the latitudes of 5° N, 15° N, 25° N, 35° N, 45° N, 55° N, and 65° N (marked latitudes, Table 1) for the first time in a migration period (Sugasawa and Higuchi, 2019). We estimated the longitudes according to the longitudes of successive location fixes before and after the individual crossing the latitudes, assuming straight flight paths.
Table
1.
The geographic regions where Whimbrels from two breeding regions flew across the marked latitudes.
Migration routes of Whimbrels in the EAAF largely differed between breeding sites. Whimbrels breeding in Sakha (Yakutia) mainly migrated along the coast and across inland, while Whimbrels breeding in Chukotka migrated mainly across the ocean (Kuang et al., 2020). Thus, we grouped the Whimbrels by their breeding areas, and compared the consistency of migration routes between the two groups.
2.3
Data analysis
To analyze the consistency of migration date of four migration events (departure from/arrival at the nonbreeding and the breeding sites), we converted the dates into count data (January 1 = 1). Then we calculated the date range and standard deviation (SD) between the first year and the second year tracked of each event for each individual. For individual tracked for more than two years, we calculated the range and standard deviation between the first year and each subsequent year, and then calculated the average of the range and the standard deviation. The lower the range and SD, the higher the consistency. Due to the abnormal distribution of the data, Wilcoxon tests with “exactRankTests” package in R were used to compare the date consistency of the four events (Hothorn and Hornik, 2022).
The “geosphere” package in R was used to calculate the distance of breeding sites and nonbreeding sites between years for each individual (Hijmans, 2022). We calculated the stopover duration of individuals at each stopover site. If the distance between the locations of stopover sites (arithmetic means of the geographical coordinates) of the same individuals in different years was less than 25 km, we assumed that the individual repeatedly used the same stopover site. Wilcoxon tests were used to compare the stopover duration at repeatedly used stopover sites and stopover sites used only once during northward and southward migration, respectively.
We calculated the SDs of the longitudes when birds crossed the marked latitudes within the same season in different years. The lower the SD, the higher the consistency of migration route. Wilcoxon tests were used to compare the SDs of longitudes of individuals from the same breeding sites within the same season. We further compared the SDs of longitudes of individuals from different breeding sites within the same season. One-way ANOVA and Bonferroni's multiple comparisons were used after normalizing the data with Box-Cox transformation to test whether SDs of longitudes differed across marked latitudes within the same season.
All statistical analyses were performed with R Studio version 4.3.2 for Windows (R Core Team, 2021). Significant differences were deemed with P < 0.05 and the values were exhibited as median (interquartile range).
3.
Results
The tracking data from a total of 15 Whimbrels were used in this study, including nine individuals with data for more than two years, and two individuals with data for more than three years (Appendix Table S1). Four individuals had incomplete data in their second tracking year. These tracked Whimbrels included five captured at Roebuck Bay, three captured at Moreton Bay, and seven captured at Shanghai Chongming Dongtan. Eleven of these birds bred in Yakutia and four bred in Chukotka.
3.1
Consistency in migration timings
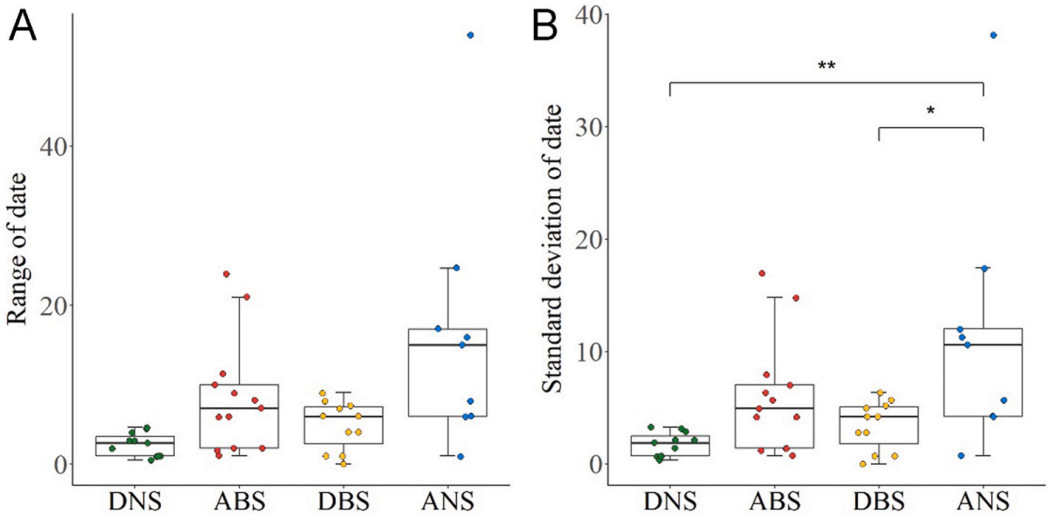
We used departure dates from nonbreeding sites in 11 birds, arrival dates at breeding sites in 13 birds, departure dates from breeding sites in 11 birds and arrival dates at nonbreeding sites in 9 birds (Appendix Table S2). Among the timings of the four migration events, the interannual variation in the date of departure from the nonbreeding sites (range = 2.5 (2.7) days, SD = 1.89 (1.77) days) was the lowest while the interannual variation in the date of arrival at the nonbreeding sites (range = 15 (11) days, SD = 10.61 (7.78) days) was the highest (Fig. 1; Appendix Table S3). The SD of the date of arrival at the nonbreeding sites was significantly larger than that of the date of departure from the nonbreeding sites and the breeding sites (Wilcoxon test, P < 0.01 for departure from the nonbreeding sites, P = 0.02 for departure from the breeding sites; Fig. 1B). There was no significant difference among the SD of the date of the other events.
Figure
1.
The range (A) and standard deviation (B) of interannual difference for each individual in departure dates from nonbreeding sites (DNS) (n=11), arrival dates at breeding site (ABS) (n=13), departure dates from breeding sites (DBS) (n=11) and arrival dates at nonbreeding sites (ANS) (n=9) of the tracked Whimbrels.
Both breeding and nonbreeding sites of the tracked Whimbrels were highly consistent. All the 15 tracked individuals used the same nonbreeding sites in different years. Twelve individuals used the same breeding sites as the previous year, one individual used a breeding site near its location from the previous year (262 km away), and the other two did not return to the breeding region in Siberia in the second year but stayed on the eastern coast of China during the breeding season.
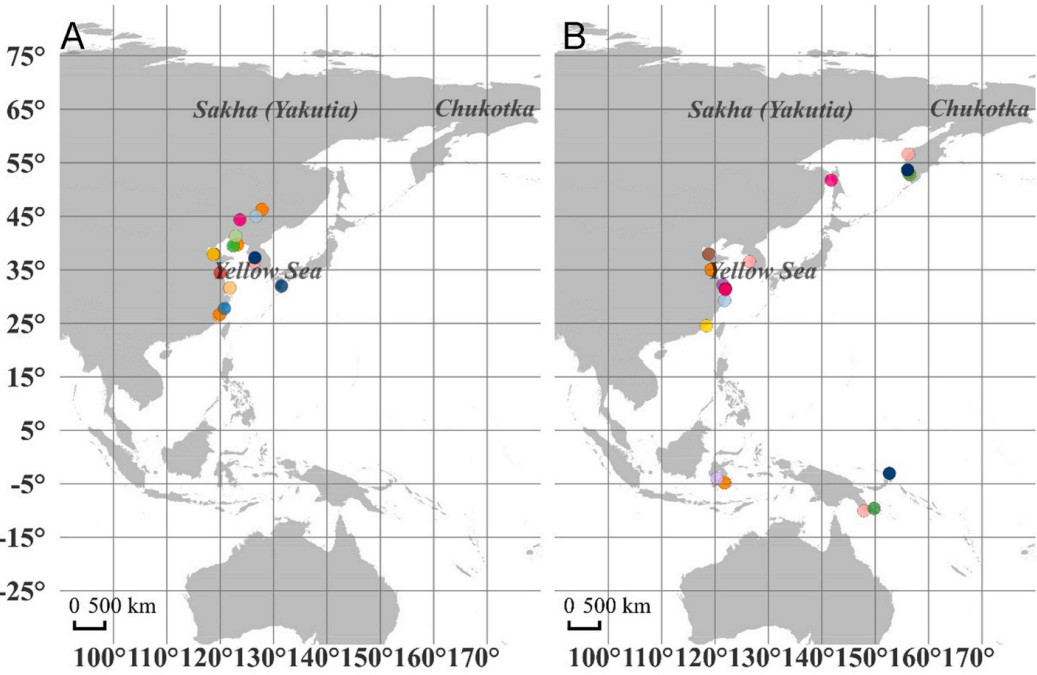
During northward migration, 13 of the 15 birds used at least one identical stopover site in different years, including one bird that used two and one bird that used three identical stopover sites. All together the tracked birds stopped 95 times at 73 stopover sites, of which 16 sites were repeatedly used a total of 37 times. The latitudes of the repeatedly used stopover sites ranged from 26° N to 47° N. Of the 16 repeatedly used stopover sites, nine were located on the Yellow Sea coast, four on the inland area of Northeast China, two along the East China Sea coast, and one on Kyushu Island, Japan (Fig. 2A).
Figure
2.
The repeatedly used stopover sites for the same tagged individuals during northward (A) and southward migration (B). The same colors in (A) and (B) indicate the same individuals. (For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)
Twelve Whimbrels had tracking data during southward migration for more than one year. One bird used three identical stopover sites, six birds used two identical stopover sites, four birds used one identical stopover site, and one bird did not use identical stopover sites between years. All together these 12 Whimbrels stopped 83 times at 54 stopover sites, of which 19 sites were repeatedly used a total of 44 times. During southward migration, the repeatedly used stopover sites were more scattered than those during northward migration. They were located in the Yellow Sea coast (seven sites), the coast of Okhotsk Sea (five sites), the Malay Archipelago (four sites), and the Bismark archipelago (one site), and the East China Sea coast (two sites) (Fig. 2B). All the Whimbrels that stopped at the Malay Archipelago and the coast of Okhotsk Sea were from the breeding sites in Chukotka.
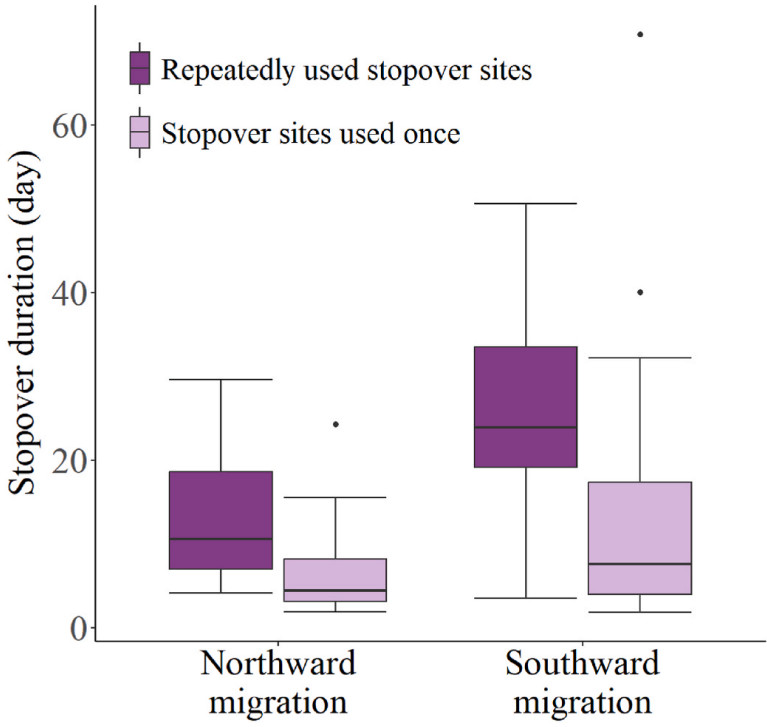
The stopover duration at the repeatedly used stopover sites (10.6 (11.65) days, n = 31 during northward migration; 23.9 (14.4) days, n = 33 during southward migration) were significantly longer than that at stopover sites used once (4.4 (5.05) days, n = 47 during northward migration; 7.6 (13.4) days, n = 33 during southward migration) (P < 0.001 for both; Fig. 3).
Figure
3.
The stopover duration at repeatedly used stopover sites and stopover sites used only once.
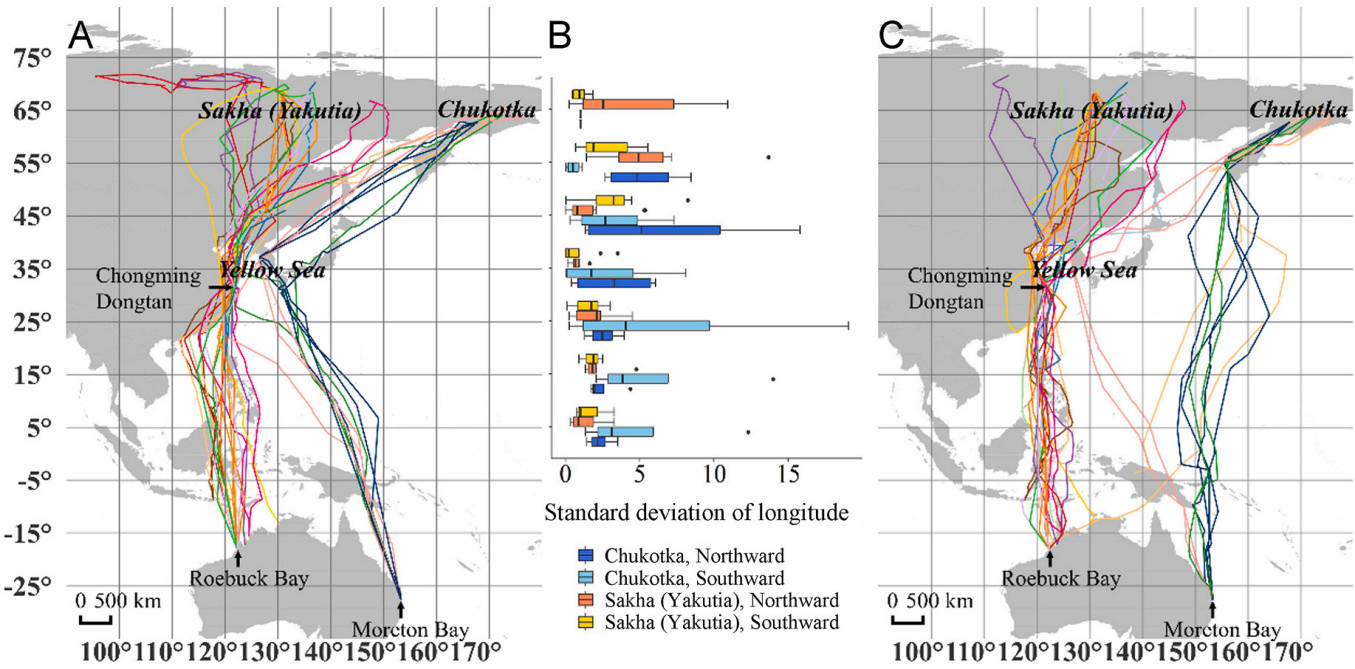
For Whimbrel individuals breeding in Yakutia, there were significant differences in the longitude variation among different latitudes during northward (P < 0.001), and no significant differences during southward (P = 0.05) migration. The SD of longitudes were the lowest at 35° N, indicating that the consistency of migration routes was the highest at that latitude (Fig. 4; Appendix Table S4). There was no significant difference in the SD of longitudes at each marked latitude between seasons (P > 0.05 for all).
Figure
4.
Migration routes of the tracked Whimbrels during northward (A) and southward migration (C), and the standard deviation of longitudes at marked latitudes for individuals breeding at Sakha and Chukotka during northward and southward migration (B). In (A) and (C), different colors indicate different individuals, the same color in (A) and (C) indicate the same individual. (For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)
All four individuals breeding in Chukotka stopped near 35° N during northward migration. During southward migration, three of the four birds flew directly from the Kamchatka Peninsula across the Western Pacific to the stopover sites near 5° S every year. Another individual first flew southwestward to the stopover site near 35° N every year and then flew southeastward to the stopover site in New Guinea. Within both seasons, there was no significant difference in the SD of longitudes among marked latitudes (P = 0.5 for northward and 0.3 for southward migration). Between seasons, there was no significant difference in the SD of longitudes at each marked latitude, except at 55° N where the SD of longitudes was significantly lower during southward than that during northward migration (P = 0.03).
During both migratory seasons, the SD of longitudes for individuals breeding in Yakutia was generally lower than that for individuals breeding in Chukotka at each marked latitude, except at 55° N during southward migration, where the SD of longitudes of individuals breeding in Chukotka is lower than those breeding in Yakutia (Fig. 4; Appendix Table S4). This suggests that the migration routes of Whimbrels breeding in Yakutia are more consistent than that of birds breeding in Chukotka.
4.
Discussion
Using multiple years of tracking data from the same individuals, we analyzed the spatiotemporal consistency between years of migrating Whimbrels in the East Asian–Australasian Flyway. The results indicate that birds exhibited high consistency in the departure date from the nonbreeding sites and the use of breeding and nonbreeding sites. We found that the interannual consistency of the migration routes differed among latitudes, and generally the migration routes of birds breeding in Yakutia were more consistent than that of birds breeding in Chukotka. We also found that birds exhibited high consistency in the use of stopover sites in the Yellow Sea region (near 35° N) between years; stopover durations were significantly longer at repeatedly used stopover sites in multiple years compared to stopover sites being used once.
Migration decisions of birds are affected by many factors, such as endogenous regulation, external environment conditions, and the birds’ body conditions (e.g., the amount of fuel storage). The consistency of migration departure dates reflects the balance among these factors (Akesson and Hedenstrom, 2000; Gronroos et al., 2012; Schmaljohann and Eikenaar, 2017). Consistent with our hypothesis, the time consistency of the Whimbrels was the highest when departing from nonbreeding sites among the four migration events, which is similar to the Whimbrels breeding in Iceland (Carneiro et al., 2019). This may be due to the start date of long-distance migration being mainly endogenously regulated (Helm et al., 2005; Akesson and Helm, 2020). Since nonbreeding sites are far away from breeding sites and the migration distance is very long, environmental conditions at nonbreeding sites cannot be used to speculate the environmental conditions along migration routes and at the breeding site. Therefore, long-distance migratory birds mainly regulate the start date of northward migration by endogenous factors (Ruthrauff et al., 2019). During southward migration, birds face less time pressure at migration destinations and thus migration timing can be more flexible (Carneiro et al., 2019). The date of departure from the breeding grounds can be largely affected by the breeding success. For example, for the altricial birds and in some nidifugous birds such as Whimbrel that parents still guard chicks, the adults that successfully reproduce will take care of the offspring for some time at the breeding site, while adults that have failed to reproduce can start southward migration earlier if they do not try to reproduce again within that breeding season (Carneiro et al., 2019). For the adults that attempt to reproduce the replacement clutch, they will stay at the breeding site for an extended period and thus delay their southward migration. This results in less consistency in the onset of southward migration between years. Due to the weaker time pressure of arrival at nonbreeding sites, birds can adjust their migration schedule based on environmental conditions and their body conditions during southward migration. This results in a more flexible migration timing and thus less interannual consistency in arrival time at nonbreeding sites.
Whimbrels exhibited high consistency in their choice of breeding and nonbreeding sites, and they generally returned to the same sites every year. This is consistent to Whimbrels breeding in Iceland (Carneiro et al., 2019). Our study also found that individuals repeatedly used one or more stopover sites along migration routes in different years. This may be related to the geographical distribution and habitat quality of these sites. Similar to many other shorebirds (Piersma et al., 2004, 2021; Hua et al., 2013; Ma et al., 2013), Whimbrels mainly use tidal habitat with abundant benthic food for fuel deposition during migration stopovers (Kuang et al., 2019). However, these habitats are limited to specific areas, prompting birds to repeatedly use these food-rich stopover sites in different years. Studies on the population dynamics and community composition of shorebirds at stopover sites in the Yellow Sea found that even when facing habitat loss and food decline, most shorebirds still use the same sites, thus maintaining relatively stable populations (Zhang et al., 2018) and communities (Wang et al., 2022) at different periods. We also found that the stopover duration at repeatedly used stopover sites by the same individuals was significantly longer than that at stopover sites being used only once, suggesting those repeatedly used stopover sites are critical habitats for migratory Whimbrels.
During northward migration, these repeatedly used stopover sites were mainly located in the Yellow Sea region. This is consistent with our hypothesis and illustrates the importance of the Yellow Sea region for migrating Whimbrels. During southward migration, besides the Yellow Sea region, the stopover sites repeatedly used by Whimbrels were also distributed on the Kamchatka Peninsula and the Malay Archipelago. The stopover sites on the Kamchatka Peninsula were used by the Whimbrels breeding in Chukotka. After departure from the Kamchatka Peninsula, some individuals flew to the Yellow Sea region and some crossed the Western Pacific and flew to the stopover sites south of the equator. Long-distance cross-sea flights require large amount of fuel store and can benefit from suitable weather conditions (Piersma et al., 2022). This suggests that the Kamchatka Peninsula is critical for the successful southward migration of Whimbrels. The Malay Archipelago is the first available stopover sites after flying thousands of kilometers across the sea, thus allowing birds to restore body condition and replenish fuel.
Analysis on the consistency of migration routes indicates that migration routes exhibited high interannual consistency at the marked latitude of 35° N, which located in the Yellow Sea region, especially for Whimbrels that breed in Yakutia. This is consistent with the fact that many individuals repeatedly used the same stopover sites in the Yellow Sea coast in different years. Although tracked individuals used the same breeding sites in different years, the consistency of migration routes for individuals breeding in Yakutia was low when close to their inland breeding sites at high latitude (55° N). This may be due to that the landmark for the inland migratory flights are less distinctive comparing to the obvious coastline at lower latitudes. Or perhaps it is due to the exploratory behavior of birds near the breeding grounds. Overall, in both seasons, the migration routes of Whimbrels breeding in Yakutia exhibited higher consistency than that of birds breeding in Chukotka. This is consistent with our hypothesis and may be due to that the latter flew across the ocean, where there is a lack of geographical reference for navigation. Since wind conditions are variable over the Pacific Ocean, birds may have to alter the flying direction to maximize wind assistance or avoid headwinds (Gill et al., 2014; Sugasawa and Higuchi, 2019). Meanwhile for individuals flying along the coast, they can choose to land at the stopover sites on the route when facing headwinds (Ma et al., 2011). During southward migration, Whimbrels breeding in Chukotka exhibited high consistency in migration routes at 55° N. This is related to the use of the final stopover sites before crossing the ocean. This once again demonstrates the importance of Kamchatka Peninsula for the Whimbrel migration.
All the four individuals breeding in Chukotka staged in the Yellow Sea region during northward migration. During southward migration, three birds flew from Kamchatka Peninsula across the Western Pacific to the stopover sites in the southern hemisphere (5° S) every year; while one individual staged in the Yellow Sea region in two years. It is still unclear why the birds breeding in the same sites used different but consistent migration routes for several years. In this study, the sample size was small for Whimbrels breeding in Chukotka. In the future, long-term migration tracking of more individuals breeding in Chukotka, combined with surveys on birds and environmental conditions at the stopover sites, can help clarify the reasons for different migration routes.
Ethics statement
Catching, banding and deployment of transmitters of birds were carried out under the license issued by the National Bird Banding Center of China and the Australian Bird and Bat Banding Scheme (ABBBS). Catching birds on the shores of Roebuck Bay was conducted with the permission of the Yawuru People. At Moreton Bay Whimbrels were tagged under licence CVL1337 issued to JTC by the Department of National Parks, Sport and Racing, Queensland Government and Department of Primary Industries Animal Ethics licence CA 2015-03-845. At Roebuck Bay Whimbrels were tagged under Regulation 17 Licence 08-00741-3 authorised to CJH from the Department of Biodiversity, Conservation and Attractions.
CRediT authorship contribution statement
Siwei An: Writing – original draft, Visualization, Methodology, Investigation, Formal analysis, Data curation. Fenliang Kuang: Writing – review & editing, Supervision, Methodology, Investigation, Data curation. Wei Wu: Investigation. Chris J. Hassell: Investigation. Jonathan T. Coleman: Writing – review & editing, Investigation. Zijing Gao: Investigation. Xuena Sun: Investigation. Yue Yuan: Investigation. Grace Maglio: Writing – review & editing, Investigation. Kar-Sin K. Leung: Writing – review & editing, Investigation. Xuesong Feng: Investigation. Zhijun Ma: Writing – original draft, Supervision, Resources, Project administration, Methodology, Investigation, Funding acquisition, Conceptualization.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
We thank the Shanghai Chongming Dongtan National Nature Reserve, Australasian Wader Studies Group, Queensland Wader Study Group, and the Northwest Australia Expedition 2018 team for their support for fieldwork. We appreciate Saijun Yuan, Hongxi Zang, Weipin Chen, Weiguo Jin, Guochang Ni, Roz Jessop, Michael Dawkins, Prue Wright, Robert Bush, Brad Woodworth, Chi-Yeung Choi, Bingrun Zhu and many volunteers for their assistance in the fieldwork. AWSG acknowledges the Yawuru People via the offices of Nyamba Buru Yawuru Limited for permission to catch birds on the shores of Roebuck Bay, traditional lands of the Yawuru people. CJH thanks his funders, WWF Netherlands, Spinoza Premium of Netherlands Organisation Prize for Scientific Research to Theunis Piersma and MAVA (Foundation Pour La Nature). The authors also acknowledge the support of the Australian Bird and Bat Banding Scheme (ABBBS) for the provision of Banding licenses and bands to the A-Class banders involved in this research.
Akesson, S., Hedenstrom, A., 2000. Wind selectivity of migratory flight departures in birds. Behav. Ecol. Sociobiol. 47, 140–144.
Akesson, S., Helm, B., 2020. Endogenous programs and flexibility in bird migration. Front. Ecol. Evol. 8, 78.
Akesson, S., Ilieva, M., Karagicheva, J., Rakhimberdiev, E., Tomotani, B., Helm, B., 2017. Timing avian long-distance migration: from internal clock mechanisms to global flights. Philos. T. Roy. Soc. B 372, 1734.
Alves, J.A., Dias, M.P., Mendez, V., Katrinardottir, B., Gunnarsson, T.G., 2016. Very rapid long-distance sea crossing by a migratory bird. Sci. Rep. 6, 38154.
Baker, A.J., Gonzalez, P.M., Piersma, T., Niles, L.J., de Lima Serrano do Nascimento, I.de L., Atkinson, P.W., et al., 2004. Rapid population decline in red knots: fitness consequences of decreased refuelling rates and late arrival in Delaware Bay. Proc. R. Soc. B 271, 875–882.
Bety, J., Giroux, J.F., Gauthier, G., 2004. Individual variation in timing of migration: Causes and reproductive consequences in greater snow geese (Anser caerulescens atlanticus). Behav. Ecol. Sociobiol. 57, 1–8.
Carneiro, C., Gunnarsson, T.G., Alves, J.A., 2019. Why are whimbrels not advancing their arrival dates into Iceland? Exploring seasonal and sex-specific variation in consistency of individual timing during the annual cycle. Front. Ecol. Evol. 7, 248.
Conklin, J.R., Battley, P.F., Potter, M.A., 2013. Absolute consistency: individual versus population variation in annual-cycle schedules of a long-distance migrant bird. PLoS One 8, 9.
Duijns, S., Anderson, A.M., Aubry, Y., Dey, A., Flemming, S.A., Francis, C.M., et al., 2019. Long-distance migratory shorebirds travel faster towards their breeding grounds, but fly faster post-breeding. Sci. Rep. 9, 9420.
Fraser, K.C., Shave, A., de Greef, E., Siegrist, J., Garroway, C.J., 2019. Individual variability in migration timing can explain long-term, population-level advances in a songbird. Front. Ecol. Evol. 7, 7.
Giunchi, D., Baldaccini, N.E., Lenzoni, A., Luschi, P., Sorrenti, M., Cerritelli, G., et al., 2019. Spring migratory routes and stopover duration of satellite-tracked Eurasian teals Anas crecca wintering in Italy. Ibis 161, 117–130.
Gronroos, J., Green, M., Alerstam, T., 2012. To fly or not to fly depending on winds: shorebird migration in different seasonal wind regimes. Anim. Behav. 83, 1449–1457.
Hasselquist, D., Montras-Janer, T., Tarka, M., Hansson, B., 2017. Individual consistency of long-distance migration in a songbird: significant repeatability of autumn route, stopovers and wintering sites but not in timing of migration. J. Avian Biol. 48, 91–102.
Helm, B., Gwinner, E., Trost, L., 2005. Flexible seasonal timing and migratory behavior: results from stonechat breeding programs. Ann. N. Y. Acad. Sci. 1046, 216–227.
Hijmans, R., 2022. Geosphere: Spherical Trigonometry Version 1, pp. 5–18. .
Hothorn, T., Hornik, K., 2022. exactRankTests: Exact Distributions for Rank and
Permutation Tests Version 0.8-35. .
Hua, N., Piersma, T., Ma, Z., 2013. Three-phase fuel deposition in a long-distance migrant, the red knot (Calidris canutus piersmai), before the flight to high arctic breeding grounds. PLoS One 8, e62551.
Ke, W.J., He, P., Peng, H.-B., Choi, C.-Y., Zhang, S.D., Melville, D.S., et al., 2019. Migration timing influences the responses of birds to food shortage at their refuelling site. Ibis 161, 908–914.
Kuang, F.L., Coleman, J.T., Hassell, C.J., Leung, K.S.K., Maglio, G., Ke, W.J., et al., 2020. Seasonal and population differences in migration of whimbrels in the East Asian-Australasian Flyway. Avian Res. 11, 24.
Kuang, F.L., Wu, W., Ke, W.J., Ma, Q., Chen, W.P., Feng, X.S., et al., 2019. Habitat use by migrating Whimbrels (Numenius phaeopus) as determined by bio-tracking at a stopover site in the Yellow Sea. J. Ornithol. 160, 1109–1119.
Kuang, F.L., Wu, W., Li, D.V., Hassell, C.J., Maglio, G., Leung, K.S.K., et al., 2022. Detecting the non-breeding region and migration route of whimbrels (Numenius phaeopus rogachevae) in the East Asian-Australasian Flyway. Avian Res. 13, 100011.
Kurten, N., Schmaljohann, H., Bichet, C., Haest, B., Vedder, O., Gonzalez-Solis, J., et al., 2022. High individual repeatability of the migratory behaviour of a long-distance migratory seabird. Movement Ecol. 10, 5.
Lank, D.B., Butler, R.W., Ireland, J., Ydenberg, R.C., 2003. Effects of predation danger on migration strategies of sandpipers. Oikos 103, 303–319.
Liu, D.P., Zhang, G.G., Jiang, H.X., Lu, J., 2018. Detours in long-distance migration across the Qinghai-Tibetan Plateau: individual consistency and habitat associations. PeerJ 6, e4304.
Loonstra, A.J., Verhoeven, M.A., Both, C., Piersma, T., 2023. Translocation of shorebird siblings shows intraspecific variation in migration routines that arise after fledging. Curr. Biol. 33, P2535–P2540.
Lourenco, P.M., Kentie, R., Schroeder, J., Groen, N.M., Hooijmeijer, J., Piersma, T., 2011. Repeatable timing of northward departure, arrival and breeding in black-tailed godwits Limosa l. limosa, but no domino effects. J. Ornithol. 152, 1023–1032.
Ma, Z.J., Hua, N., Peng, H.B., Choi, C., Battley, P.F., Zhou, Q.Y., et al., 2013. Differentiating between stopover and staging sites: Functions of the southern and northern yellow sea for long-distance migratory shorebirds. J. Avian Biol. 44, 504–512.
Ma, Z.J., Hua, N., Zhang, X., Guo, H.Q., Zhao, B., Ma, Q., et al., 2011. Wind conditions affect stopover decisions and fuel stores of shorebirds migrating through the south Yellow Sea. Ibis 153, 755–767.
Newton, I., 2008. The Migration Ecology of Birds. Academic Press, London.
Pedersen, L., Jackson, K., Thorup, K., Tottrup, A.P., 2018. Full-year tracking suggests endogenous control of migration timing in a long-distance migratory songbird. Behav. Ecol. Sociobiol. 72, 139.
Piersma, T., Gill, R.E., Ruthrauff, D.R., Guglielmo, C.G., Conklin, J.R., Handel, C.M., 2022. The Pacific as the world's greatest theater of bird migration: extreme flights spark questions about physiological capabilities, behavior, and the evolution of migratory pathways. Ornithology 139, ukab086.
Piersma, T., Kok, E.M.A., Hassell, C.J., Peng, H.-B., Verkuil, Y.I., Lei, G., et al., 2021. When a typical jumper skips: itineraries and staging habitats used by red knots (Calidris canutus piersmai) migrating between northwest Australia and the New Siberian Islands. Ibis 163, 1235–1251.
Piersma, T., Rogers, D., Gonz´ alez, P., Zwarts, L., Niles, L., Lima, I., et al., 2004. Fuel
storage rates before northward flights in red knots worldwide: facing the severest
ecological constraint in tropical intertidal environments? In: Greenberg, R., Marra, P.
P. (Eds.), Birds of Two Worlds: Ecology and Evolution of Migration. The Johns
Hopkins University Press, Baltimore, pp. 262–273.
Potti, J., 1998. Arrival time from spring migration in male pied flycatchers: individual consistency and familial resemblance. Condor 100, 702–708.
Prater, A.J., Marchant, J.H., Vuorinen, J., 1977. Guide to the Identification and Ageing of Holarctic Waders. BTO, Tring.
R Core Team, 2021. R: A Language and Environment for Statistical Computing. R
Foundation for Statistical Computing, Vienna, Austria. .
Ruthrauff, D.R., Tibbitts, T.L., Gill, R.E., 2019. Flexible timing of annual movements across consistently used sites by marbled godwits breeding in Alaska. Auk 136, uky007.
Schmaljohann, H., Eikenaar, C., 2017. How do energy stores and changes in these affect departure decisions by migratory birds? A critical view on stopover ecology studies and some future perspectives. J. Comp. Physiol. 203, 411–429.
Schmaljohann, H., Eikenaar, C., Sapir, N., 2022. Understanding the ecological and evolutionary function of stopover in migrating birds. Biol. Rev. 97, 1231–1252.
Stanley, C.Q., MacPherson, M., Fraser, K.C., McKinnon, E.A., Stutchbury, B.J.M., 2012. Repeat tracking of individual songbirds reveals consistent migration timing but flexibility in route. PLoS One 7, e40688.
Studds, C.E., Kendall, B.E., Murray, N.J., Wilson, H.B., Rogers, D.I., Clemens, R.S., et al., 2017. Rapid population decline in migratory shorebirds relying on Yellow Sea tidal mudflats as stopover sites. Nat. Commun. 81, 14895.
Sugasawa, S., Higuchi, H., 2019. Seasonal contrasts in individual consistency of oriental honey buzzards' migration. Biol. Lett. 15, 20190131.
Tedeschi, A., Sorrenti, M., Bottazzo, M., Spagnesi, M., Telletxea, I., Ibanez, R., et al., 2020. Interindividual variation and consistency of migratory behavior in the Eurasian woodcock. Curr. Zool. 66, 155–163.
van Wijk, R.E., Bauer, S., Schaub, M., 2016. Repeatability of individual migration routes, wintering sites, and timing in a long-distance migrant bird. Ecol. Evol. 6, 8679–8685.
Vardanis, Y., Klaassen, R.H.G., Strandberg, R., Alerstam, T., 2011. Individuality in bird migration: routes and timing. Biol. Lett. 7, 502–505.
Vardanis, Y., Nilsson, J.A., Klaassen, R.H.G., Strandberg, R., Alerstam, T., 2016. Consistency in long-distance bird migration: contrasting patterns in time and space for two raptors. Anim. Behav. 113, 177–187.
Verkuil, Y.I., Karlionova, N., Rakhimberdiev, E.N., Jukema, J., Wijmenga, J.J., Hooijmeijer, J.C.E.W., et al., 2012. Losing a staging area: eastward redistribution of Afro-Eurasian ruffs is associated with deteriorating fuelling conditions along the western flyway. Biol. Conserv. 149, 51–59.
Wang, X.D., Chen, Y., Melville, D.S., Choi, C.Y., Tan, K., Liu, J.J., et al., 2022. Impacts of habitat loss on migratory shorebird populations and communities at stopover sites in the Yellow Sea. Biol. Conserv. 269, 109547.
Watson, J.W., Keren, I.N., 2019. Repeatability in migration of ferruginous hawks (Buteo regalis) and implications for nomadism. Wilson J. Ornithol. 131, 561–570.
Wellbrock, A.H.J., Bauch, C., Rozman, J., Witte, K., 2017. Same procedure as last year? Repeatedly tracked swifts show individual consistency in migration pattern in successive years. J. Avian Biol. 48, 897–903.
Weller, F.G., Beatty, W.S., Webb, E.B., Kesler, D.C., Krementz, D.G., Asante, K., et al., 2022. Environmental drivers of autumn migration departure decisions in midcontinental mallards. Movement Ecol. 10, 1.
Zhang, S.D., Ma, Z.J., Choi, C.Y., Peng, H.B., Bai, Q.Q., Liu, W.L., et al., 2018. Persistent use of a shorebird staging site in the yellow sea despite severe declines in food resources implies a lack of alternatives. Bird Conserv. Int. 28, 534–548.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds