The Korea Institute of Ornithology and Department of Biology, Graduate School, Kyung Hee University, 26 Kyungheedae-ro, Dongdaemun-gu, Seoul, 02447, Republic of Korea
b.
National Institute of Biological Resources, 42 Hwangyeong-ro, Seo-gu, Incheon, 22689, Republic of Korea
c.
Endangered Species Research Center, National Institute of Ecology, Yeongyang, 36351, Republic of Korea
d.
Wildlife Sciences and Conservation Center of Mongolia, Union Building, B-701, UNESCO str., Ulaanbaatar, 14210, Mongolia
Funds: This work was supported by a grant the National Institute of Biological Resources, funded by the Ministry of Environment, Republic of Korea (grant numbers NIBR202216101 and NIBR202223101). The funder had no role in the study design; in the collection, analysis or interpretation of data; in the writing of the report; or in the decision to submit the article for publication
Long-distance migratory birds travel more rapidly in spring than in autumn, as they face temporal breeding constraints. However, several species travel slower in spring owing to environmental influences, such as food availability and wind conditions. GPS trackers were attached to 17 Whooper Swans (Cygnus cygnus) inhabiting northeastern Mongolia, to determine their migration routes and stopover sites in spring and autumn. Differences between spring and autumn migrations, migration-influencing parameters, and the effect of spring stopover site temperatures were analyzed. Six swans completed perfect tours between their wintering and breeding sites, and these data were used for analysis. Spring migration lasted 57 days, with 49.2 days spent at 3.7 stopover sites. Autumn migration lasted 21.5 days, with 17.5 days spent at 1.0 stopover sites. Thus, the swans traveled more rapidly in autumn than in spring. Migration distance, number of stopovers, migration speed, and straightness were important migration determinants in both spring and autumn. Migration distance, stopover duration, number of stopovers, daily travel speed, travel duration, and migration speed differed significantly between spring and autumn. During spring migration, the temperature at the current stopover sites and that at the future stopover sites displayed significant variations (t = 1585.8, df = 631.6, p < 0.001). These findings are critical for the conservation and management of Whooper Swans and their key habitats in East Asian regions, and the data are anticipated to make a particularly significant contribution toward developing detailed management plans for the conservation of their key habitats.
Long-distance migratory birds are generally under greater pressure from temporal constraints during their spring migration than during their autumn migration, as they must ensure an advantageous position in the competition for breeding territories. These birds are known to migrate more rapidly in spring than in autumn (Fransson, 1995; Kokko, 1999; Alerstam, 2006; Yohannes et al., 2009; Klaassen et al., 2012; Nilsson et al., 2013). Birds arriving at the breeding site early (having ensured an advantageous position in the competition) can also start breeding early (Moore et al., 2005), which consequently increases reproductive capacity (Van Noordwijk et al., 1995). Another advantage is that these birds can secure a greater amount of time for breeding, as well as for long-distance migration to their wintering sites after breeding (McNamara et al., 1998). This leads to the general expectation that autumn migration is slower than spring migration because the migratory birds are not driven by such intense time pressure in autumn (Nilsson et al., 2013; Kölzsch et al., 2016).
Nevertheless, certain species of birds display contrasting patterns of spring and autumn migration (Shamoun-Baranes et al., 2003; Mosbech et al., 2006; Nolet, 2006; Klaassen et al., 2011; Limiñana et al., 2012; Bustnes et al., 2013; Mellone et al., 2015; Deng et al., 2019). Most birds with contrasting migratory patterns share a characteristic large body (Kölzsch et al., 2016). Compared to small birds, large birds face a higher time-related burden, as their breeding and moulting demand a greater amount of time and energy (Rahn and Ar, 1974). In addition, it is not possible to save the amount of energy required to migrate the entire distance to their final destination; the maximum distance that can be traveled without refueling at a stopover site is limited (Alerstam et al., 2003). For certain species of large, long-distance migratory birds, a relatively slower spring migration than autumn migration can mainly be attributed to the following four causes (Ao et al., 2020): (1) the cost of reaching the breeding site early is high (Bêty et al., 2004); (2) food availability fluctuates seasonally; (3) wind conditions vary according to season; and (4) ice formation occurs at the stopover site (Shamoun-Baranes et al., 2003).
Whooper Swans (Cygnus cygnus) are a well-known large migratory waterbird species whose spring migration is slower than their autumn migration (Ao et al., 2020). Whooper Swans are categorized into five groups based on their wintering and breeding sites: Iceland, North-Western Europe, the Black and East Caribbean Seas, the Caspian Sea and West-Siberia, and East Asia (Rees et al., 2019; Wetlands International, 2022). Recently, the East Asian population has been estimated to be approximately 57,690 birds (Wetlands International, 2022), and the number of birds that spend their winter in South Korea was estimated to be 9152 in January 2021 (National Institute of Biological Resources, 2021), accounting for approximately 16% of the East Asian population. The number of Whooper Swans wintering in South Korea has continuously increased each year (Choi et al., 2018). Thus, the geographical significance of South Korea as a wintering destination for Whooper Swans is expected to show a consistent increase.
Previous studies on the migration routes of ducks and geese included studies on the characteristics of the spring and autumn migrations of White-Fronted Geese (Anser albifrons), which showed that the variation in migration speed was determined by the use of stopover sites (Kölzsch et al., 2016; Deng et al., 2019). For the swan species Bewick's Swan (Cygnus columbianus bewickii), ice formation in the habitat (stopovers and breeding sites) slowed spring migration (Nuijten et al., 2014); however, to the best of our knowledge, no such reports on Whooper Swans exist. Moreover, only a few studies have investigated the detailed migration characteristics of Whooper Swans in South Korea during winter (Ao et al., 2020). Thus, it is necessary to understand the connection between the breeding and wintering sites of Whooper Swans and to identify the underlying mechanisms.
This study aimed to: (1) identify the specific migration routes and key stopover sites of Whooper Swans wintering in South Korea; (2) examine the variation in migration patterns between spring and autumn and identify the influencing factors; and (3) determine the effect of ice formation in stopover habitats during their spring migration. Based on these findings, the ecological significance of the migration characteristics of Whooper Swans wintering in South Korea is discussed.
2.
Methods
2.1
Whooper Swan capture and GPS tracker attachment
Between July 31, 2018, and July 22, 2019, a total of 17 Whooper Swans (6 in 2018 and 11 in 2019) were captured at Buir Lake (47.801° N, 117.696° E) located in the northeastern part of Mongolia. A neck band for identification and a GPS tracker (WT-300, KoEco, Republic of Korea) were attached to each bird (Table 1). Whilst moulting, the birds display no flight for approximately 5–6 weeks (Boiko et al., 2014). Lake inhabitants that showed moulting characteristics were steered by the boat and directly captured using a dip-net. The GPS tracker was attached to the bird in the form of a backpack using a Teflon ribbon. The weight of the tracker was 65 g, which is approximately 0.73% of the average weight of a Whooper Swan (8903 g; range: 8000–10,300 g). The GPS tracker collected location data through a GPS satellite and transmitted the data via the mobile network. The data were set to be received six times per day at 4 h intervals and were downloaded from the website (http://www.wi-tracker.com). Among the 17 Whooper Swans with trackers attached, six made successful journeys between their wintering and breeding sites, while no data were received from three birds after the day of tracker attachment, likely owing to device malfunction or the death of the bird. Four Whooper Swans traveled to their wintering sites after tracker deployment, but the data collection stopped at the wintering sites. Four other Whooper Swans began to move toward their wintering sites, but data download stopped before they arrived. Data from the six Whooper Swans that completed the entire round-trip between their wintering and breeding sites were used in the final analysis.
Table
1.
General information for Whooper Swans with trackers attached.
2.2
Whooper Swan migration parameters: definitions and estimation
To analyze migration strategies, the annual cycle of Whooper Swans was divided into four periods: breeding and moulting, autumn migration, wintering, and spring migration. The breeding site was defined as the area furthest north on the spring migration route, where the bird stayed for a minimum of 30 days (the incubation period for Whooper Swans is approximately 31–42 days; Kear, 2005). The wintering site was defined as the area furthest south on the autumn migration route, where the bird stayed for a minimum of seven days. A stopover site was defined as an area on the spring or autumn migration routes at which the bird stayed for at least 72 h (3 days) continuously (Li et al., 2018). For each bird, the spring migration was defined as starting at the last location of the wintering site with GPS signal and ending at the first location of the breeding site with GPS signal; the autumn migration was defined as starting at the last location of the breeding or moulting site with GPS signal and ending at the first location of the wintering site with GPS signal.
To analyze the migration characteristics of Whooper Swans, eight migration parameters (migration duration, migration distance, stopover duration, stopover number, migration speed, travel duration, daily travel speed, and straightness index) were determined (Benhamou, 2004; Nilsson et al., 2013). Each parameter was defined as shown in Table 2.
Table
2.
Define of migration parameters of Whooper Swans.
Variable
Descript
Unit
Migration duration
Total length of the spring or autumn migration including the time spent at stopovers
days
Migration distance
Total distance traveled between the wintering and breeding sites, excluding distances within the wintering, breeding, and stopover sites—it was calculated as the sum of the distances between locations along a continuous path and not as a linear distance connecting the start and end points
km
Stopover duration
The sum of the days spent at each stopover site during the spring and autumn migrations
days
Number of stopovers
The number of stopover sites where the bird spent at least three days during their spring or autumn migrations
–
Daily travel speed
The value calculated by dividing migration distance by travel duration
km/day
Travel duration
The value calculated difference between migration duration and stopover duration
days
Migration speed
The value calculated by dividing migration distance by migration duration
km/day
Straightness index
The value calculated by dividing the linear distance between the start and end points by the migration distance (Benhamou, 2004)
Given that Whooper Swans predominantly feed on the roots of water plants at shallow depths (Shimada et al., 2014), ice formation can have a considerable impact on the availability of this food source for energy refueling during migration (Shamoun-Baranes et al., 2003). To determine the likelihood of ice formation in the stopover habitats, temperature data for the stopover sites used by Whooper Swans during their spring migrations were analyzed. Based on the arrival date of the Whooper Swans at each stopover site used during spring migration, the daily mean, maximum, and minimum temperatures were obtained for the two weeks prior to arrival at the current stopover site and future stopover sites. Temperature data for each stopover site were obtained from the Ventusky website (www.ventusky.com).
2.4
Statistical analyses
A generalized linear mixed-effects model (GLMM) was used to determine the factors influencing the variation between the spring and autumn migrations of the six tracked Whooper Swans. Prior to applying the GLMM, the multicollinearity of the variables was checked by evaluating the variance inflation factor (VIF) and performing a correlation analysis for the variables with high VIFs. Variables with a VIF ≥ 10 were regarded as exhibiting high multicollinearity and were thus excluded from the model (Kutner et al., 2004). Akaike's information criterion (AIC) was used in model selection (Burnham and Anderson, 2002). All possible models were tested and only those with AIC values below that of the null model were applied. All eight migration parameters showed VIFs ≥ 10. Thus, the migration duration was used as the representative parameter, and a wintering site parameter was added in the final model. Using AIC, the Akaike weight (wi; the weight of evidence in favor of model i) of each model and the model order were estimated. Additionally, the relative importance of each parameter included in the model was calculated as the sum of wi (Burnham and Anderson, 2002). The models exhibiting differences in the AIC of < 2 were determined to be the best models.
To examine the variation between the spring and autumn migrations, one-tailed Wilcoxon's signed-rank tests were performed on the variables with confirmed importance in the models and the variables that were strongly correlated with them. In addition, two-tailed t-tests were performed on the daily mean, maximum, and minimum temperature data to comparatively analyze the mean values for the current and future stopover sites for two weeks prior to the arrival date at each stopover site in the spring migration. For all statistical analyses, R version 3.6.1 was used (R Core Team, 2019). The Car package was used for the VIF (Fox and Weisberg, 2011), the Performance Analytics package for the correlation analysis (Peterson and Carl, 2014), the lme4 package for the GLMM (Bates et al., 2014), the MuMIn package for model selection (Barton and Barton, 2015), and the PairedData package for the one-tailed Wilcoxon's signed-ranks test (Champely, 2018). For all the statistical tests, p < 0.05 was considered statistically significant and statistical results were presented as mean ± standard error.
3.
Results
3.1
Autumn migration routes
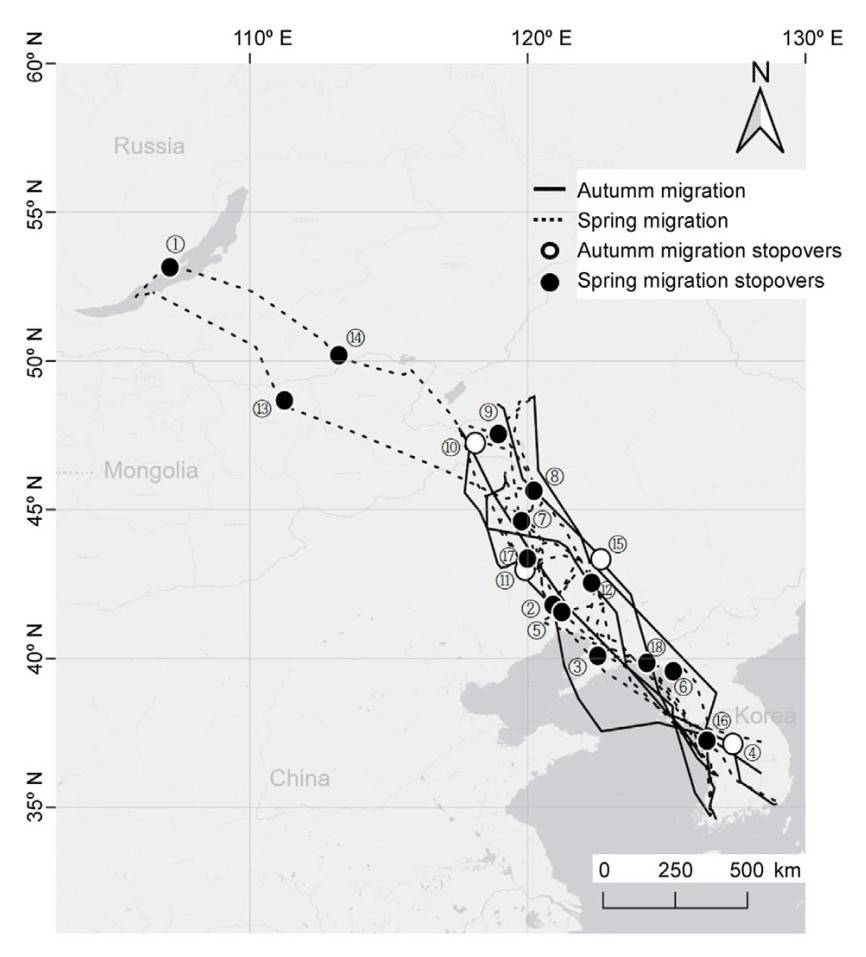
The GPS tracked Whooper Swans stayed at Buir Lake in Mongolia until early October, starting their autumn migration between October 13 and 26. Four stopover sites were used for autumn migration (Table 3). Among the Whooper Swans, WS_1 migrated approximately 1430 km to its wintering site in Taean, South Korea, in one trip without resting. WS_3 traveled from Buir Lake in Mongolia straight to Cheongmi-cheon, South Korea (1418 km linear distance) for its first stopover, stayed at the site for approximately 16 days, and then traveled to its wintering site in Busan, South Korea. WS_6, WS_10, and WS_17 traveled from Buir Lake in Mongolia to their first stopover at the Horqin Left Rear Banner reservoir in Inner Mongolia, China (622 km linear distance), then migrated straight to their wintering sites in Yeongam and Seocheon in South Korea. WS_11 traveled from Buir Lake in Mongolia to its first stopover at Khalkhgol Lake, Mongolia, approximately 62 km south of Buir Lake. Its second stopover was on the Laoha River, Inner Mongolia, China, and from there, it traveled to its wintering site in Gumi, South Korea. The autumn migrations of the tracked Whooper Swans were completed between October 29 and November 18, and all of these birds wintered at sites in South Korea (Fig. 1).
Table
3.
Key stopovers during the spring and autumn migrations of Whooper Swans.
Stopover sites
Latitude
Longitude
Bird ID
Arrival date
Departure date
Season
Lake Baikal, Russia
107.116
53.145
WS_1
04/19/2019
04/23/2019
Spring
Baishi Reservoir, China
120.915
41.794
WS_10
03/03/2020
03/18/2020
Spring
WS_11
02/28/2020
03/23/2020
Spring
WS_17
03/08/2020
03/13/2020
Spring
Biliu River, China
122.523
40.090
WS_1
03/04/2019
03/16/2019
Spring
Cheongmi Stream, Republic of Korea
127.388
37.127
WS_3
10/29/2018
11/14/2018
Autumn
Daling River, China
121.229
41.556
WS_1
03/17/2019
03/25/2019
Spring
Farmland in Jungju-gun, North Korea
125.241
39.571
WS_3
04/07/2019
04/22/2019
Spring
Haiha'er River, Inner Mongolia, China
119.779
44.615
WS_10
03/25/2020
03/28/2020
Spring
WS_6
04/28/2019
05/21/2019
Spring
Huolin River, Inner Mongolia, China
120.223
45.636
WS_6
05/22/2019
05/27/2019
Spring
Khalkh River, Mongolia
118.943
47.543
WS_3
05/12/2019
06/02/2019
Spring
Khalkhgol Lake, Mongolia
118.113
47.242
WS_11
10/13/2019
10/22/2019
Autumn
Laoha River, Inner Mongolia, China
119.903
42.980
WS_11
10/25/2019
11/14/2019
Autumn
WS_10
03/18/2020
03/24/2020
Spring
WS_11
03/23/2020
04/03/2020
Spring
Liuhe River, China
122.303
42.540
WS_6
03/18/2019
04/12/2019
Spring
Onon River, Mongolia
111.240
48.674
WS_11
04/04/2020
04/06/2020
Spring
Onon River, Russia
113.198
50.196
WS_1
04/13/2019
04/18/2019
Spring
Reservoir of the Horqin Left Rear Banner, Inner Mongolia, China
Figure
1.
Migration routes and key stopovers used by individual Whooper Swans, linked by lines. (1) Lake Baikal, Russia; (2) Baishi Reservoir, China; (3) Biliu River, China; (4) Cheongmi Stream, Republic of Korea; (5) Daling River, China; (6) Farmland of Jungju-gun, North Korea; (7) Haiha'er River, Inner Mongolia, China; (8) Huolin River, Inner Mongolia, China; (9) Khalkh River, Mongolia; (10) Khalkhgol Lake, Mongolia; (11) Laoha River, Inner Mongolia, China; (12) Liuhe River, China; (13) Onon River, Mongolia; (14) Onon River, Russia; (15) Reservoir of the Horqin Left Rear Banner, Inner Mongolia, China; (16) Youngheungdo Wetland, Republic of Korea; (17) Xar Moron River, Inner Mongolia, China; (18) Yalu River Estuary, North Korea.
The spring migrations of the Whooper Swans after wintering started between February 19 and March 14. Fifteen stopover sites were used for spring migration (Table 3). WS_1 started its spring migration at Taean, South Korea, flew 536 km to its first stopover site at the Biliu River in China, and then made a series of stopovers at the following sites (in order): the Daling River in China, Xar Moron River in Inner Mongolia, Reka Onon in Russia, and an area west of Lake Baikal in Russia. Finally, the bird arrived at a breeding site in the southeastern area of Lake Baikal in Russia. WS_3 started its spring migration in Busan, South Korea, flew 337 km to its first stopover site at Yeongheung-do, South Korea, where it spent approximately 31 days, and then resumed travel via a series of stopover sites, as follows: farmland in Jungju-gun, North Korea; the Yalu River Estuary in North Korea; and the Khalkh River in Mongolia. The bird settled at a breeding site at Buir Lake in Mongolia. WS_6 started its spring migration at Yeongam, South Korea, flew 974 km to its first stopover site on the Liuhe River in China, and then made a series of stopovers at the following sites: the Xar Moron River, Inner Mongolia Autonomous Region, China; the Haiha'er River, Inner Mongolia, China; and the Huolin River, Inner Mongolia, China. The bird settled at a breeding site in Hesi Genaori Lake, Inner Mongolia, China. WS_10 started its spring migration at Seocheon, South Korea. The bird spent approximately 15 days at its first stopover site at the Baishi Reservoir in China, and then resumed travel via 2 stopover sites in Inner Mongolia, China, the Laoha River and the Haiha'er River. The bird settled at a breeding site in a wetland near the Buridu Nuori River in Inner Mongolia, China. WS_11 started its spring migration at Yeongwol, South Korea, flew 509 km to its first stopover site in the Yalu River Estuary, North Korea, and then traveled to the following stopover sites: Baishi Reservoir, China; Laoha River, Inner Mongolia, China; and Onon River, Mongolia. The bird settled at a breeding site in the southeastern area of Lake Baikal in Russia. WS_17 started its spring migration in Yeongam, South Korea. It made its first stopover at the Baishi Reservoir in China, its second stopover at the Xar Moron River in Inner Mongolia, China, and then arrived at its breeding site in a wetland near the Buridu Nuori River in Inner Mongolia, China. The spring migrations of the tracked Whooper Swans were completed between March 29 and June 2 (Fig. 1).
3.3
Correlation analysis of migration parameters
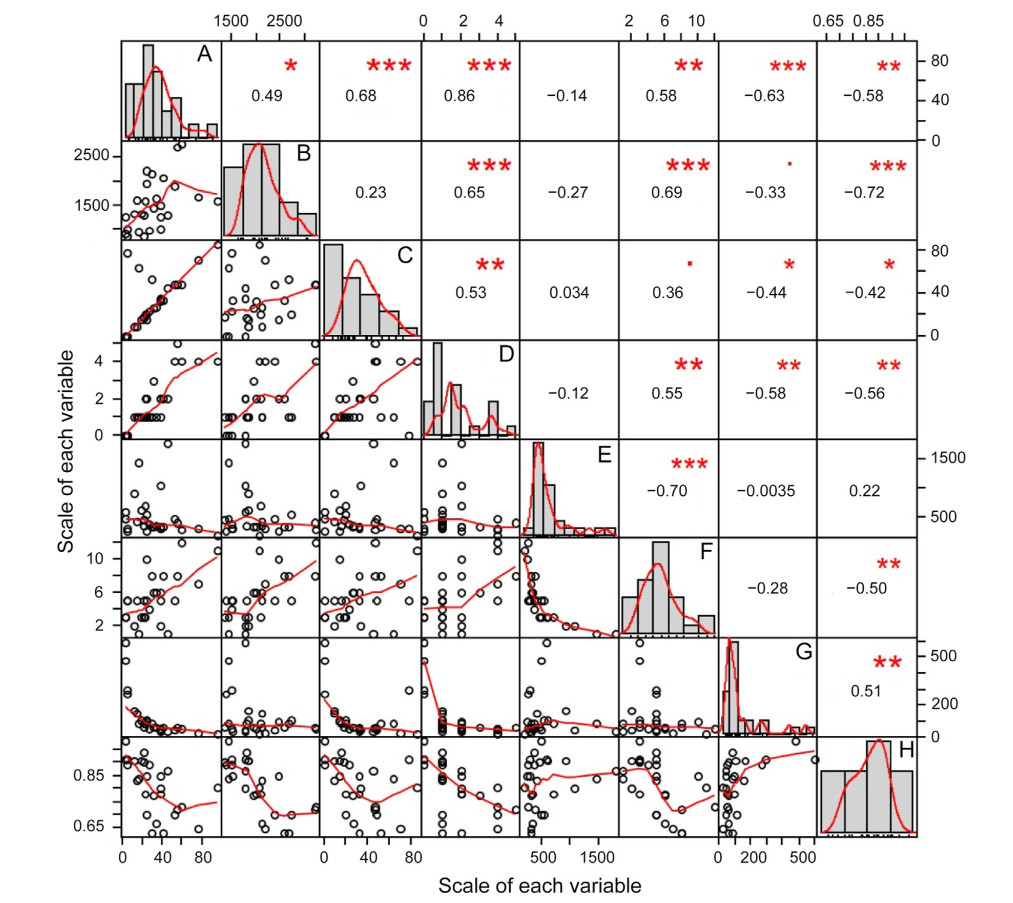
Most of the eight migration parameters exhibited strong correlations (Fig. 2). Notably, the parameters strongly correlated with migration duration were stopover duration (r = 0.68, p < 0.001), stopover number (r = 0.86, p < 0.001), travel duration (r = 0.58, p > 0.01), and migration speed (r = −0.63, p > 0.001), whereas those strongly correlated with migration distance were stopover number (r = 0.65, p > 0.001), travel duration (r = 0.69, p > 0.001), and straightness index (r = −0.72, p > 0.001). Travel duration was strongly correlated with daily travel speed (r = −0.70, p > 0.001). The strong correlations found between the migration duration and migration speed, the migration distance and straightness index, and the daily travel speed and travel duration were negative, whereas all the other strong correlations were positive.
Figure
2.
Correlation matrix. The distribution of each variable is shown on the diagonal. Below the diagonal the bivariate scatter plots are presented with a fitted line (red line). Above the diagonal the correlation coefficient is presented; the size of the text represents the strength of the correlation and the significance level is indicated by asterisks: ***, **, and * indicate p-values of 0.001, 0.01, and 0.05, respectively. (A) migration duration (days), (B) migration distance (km), (C) stopover duration (days), (D) number of stopovers, (E) daily travel speed (km/day), (F) travel duration (day), (G) migration speed (km/day), and (H) straightness index. (For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
3.4
Factors influencing the spring and autumn migrations
The model exhibiting the lowest AIC was that incorporating migration distance and stopover number, and a total of five models showed a difference in AIC < 2 (Table 4). The cumulative AIC weight of the five models was 79%. In these five models, migration distance, stopover number, migration speed, and the straightness index were the determinants with the greatest importance in both the spring and autumn migrations. Notably, in all five models, the number of stopovers was verified, whereas the migration speed was verified in three models, and the migration distance in two models. The capture region and wintering site parameters did not affect the spring or autumn migrations (Table 4).
Table
4.
Candidate model, number of parameters (K), Akaike's information criterion (AIC), difference from lowest AIC value (ΔAIC), and Akaike model weight used to determine factors influencing migration season (spring and autumn) of Whooper Swans.
Parameter
K
logLik
AIC
ΔAIC
Weight
Migration distance + number of stopovers
5
0.35
12.15
0.00
0.22
Number of stopovers
4
−1.46
12.75
0.60
0.16
Migration distance + number of stopovers + migration speed
6
1.67
12.85
0.70
0.15
Number of stopovers + migration speed
5
−0.10
13.06
0.91
0.14
Number of stopovers + migration speed + straightness index
6
1.39
13.42
1.26
0.12
K: number of parameters; AIC: Akaike's information criterion; ΔAIC: difference from lowest Akaike's information criterion value.
Eight migration parameters (migration duration, migration distance, stopover duration, stopover number, daily travel speed, travel duration, migration speed, and straightness index) were determined through correlation analysis and model selection, and all were verified as crucial determinants of the spring and autumn migrations. Among the eight parameters, the migration distance, stopover duration, stopover number, daily travel speed, travel duration, and migration speed exhibited statistical significance in the one-tailed Wilcoxon's signed-rank test comparing the spring and autumn migrations (migration distance, V = 6, p < 0.05; stopover duration, V = 1, p < 0.05; number of stopovers, V = 0, p < 0.05; daily travel speed, V = 0, p < 0.05; travel duration, V = 1, p < 0.05; and migration speed, V = 21, p < 0.05). The migration distance, stopover duration, number of stopovers, and travel duration gave higher values during the spring migration, whereas the daily travel speed and migration speed gave higher values during the autumn migration (Table 5).
Table
5.
Migration parameters of tracked Whooper Swan.
3.5
Relationship between temperature and spring migration
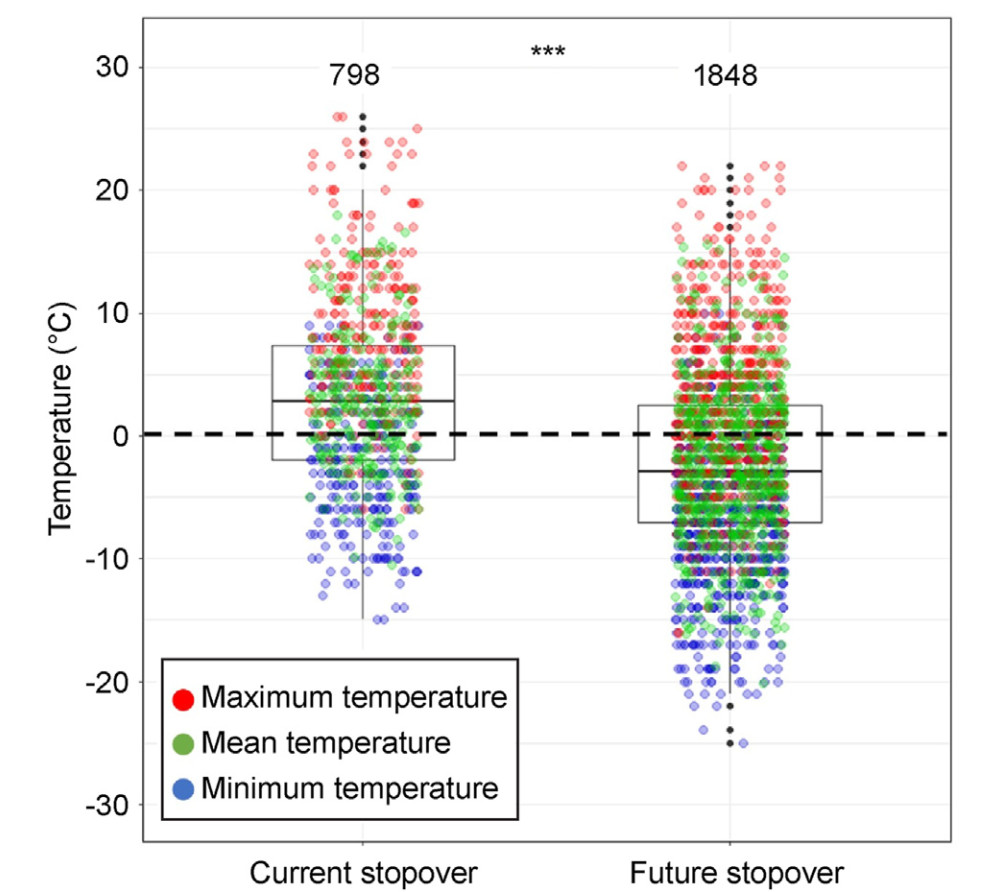
A significant variation was found between the current stopover sites and the future stopover sites, with respect to the daily mean, maximum, and minimum temperatures for the two weeks prior to the arrival date at each stopover site during the spring migration (Fig. 3; t = 1585.8, df = 631.6, p < 0.001). The average maximum and minimum temperatures at the current and future stopover sites were 3.07 ± 0.26 ℃ and −2.21 ± 0.18 ℃, respectively, indicating that the temperature was above 0 ℃ at the current stopover sites and below 0 ℃ at the future stopover sites.
Figure
3.
Comparison of maximum, mean, and minimum temperatures between current and future stopover sites during spring migration. Boxes represent the interquartile temperature ranges, and the line within each box denotes the median value; the whiskers encompass the majority of the points. The sample sizes for each stopover are presented above the box plots. The dotted black line indicates a temperature of 0 ℃. A t-test was used to assess significance; *** indicates p-values < 0.001.
This study identified the specific migration routes of Whooper Swans wintering in South Korea. In addition, analysis of the migration parameters of Whooper Swans showed variations between the spring and winter migrations. Some of the migration parameters identified for the spring and autumn migration patterns of Whooper Swans exhibited distinct variations.
Whooper Swans wintering in South Korea were found to travel through the lower reaches of the Amnokgang River in South Korea, and the Xinkai River near Tongliao, Inner Mongolia, China, to reach breeding sites near Hulun in Mongolia, at Buir Lake in Mongolia, or further north in the southern area of Lake Baikal in Russia. These migration routes differ from those previously reported for East Asia. The population of Whooper Swans that winter in Hokkaido, Japan, are known to migrate from eastern Hokkaido, via the lower reaches of the Amur River and the northern shores of the Okhotsk Sea, to breeding sites in the lower reaches of the Indigirka or Kolyma Rivers in northern Russia (Shimada et al., 2014). The population of Whooper Swans in China is known to winter at the Longyangxia Reservoir, middle reaches of the Yellow River in Shanxi and Henan Provinces, Jiuquan Wetland or along the Rongcheng coast, whereas they breed at the Shaazgai, Uvs, or Airag Lakes in western Mongolia (Ao et al., 2020). The results of this study suggest that the Whooper Swans inhabiting East Asia use varying migration routes depending on their wintering and breeding regions (Mathiasson, 2013; Ao et al., 2020), and that the population in South Korea is clearly distinct from other populations, owing to their use of unique migration routes.
Our results confirmed that the migration duration and stopover duration in spring migration were longer than in autumn for Whooper Swans (Table 5). Similar migration characteristics have been previously observed in other swan species that use different migration routes (Nolet, 2006; Nuijten et al., 2014; Ao et al., 2020). The Tundra Swan (Cygnus columbianus), for example, was shown to migrate more slowly in spring than in autumn in the Western Nearctic, Eastern Nearctic, Western Palearctic, and Eastern Palearctic flyways (Nowak et al., 1990; Higuchi et al., 1991; Ely et al., 1997; Beekman et al., 2002; Kamiya and Ozaki, 2002; Petrie and Wilcox, 2003). The slower spring migration than autumn migration in Whooper Swans is presumed to be caused by a combination of factors, such as breeding strategies and environmental conditions (Nuijten et al., 2014). Most swans, including Whooper Swans, are well-known capital breeders; meaning that the energy required for migration and the energy required for breeding must be strictly balanced (Bromley and Jarvis, 1993; Bruderer and Salewski, 2009; Ely and Meixell, 2016). Hence, energy refueling at stopover sites during spring migration is a prerequisite for the breeding success of these birds (Nolet, 2006; Nuijten et al., 2014). However, as ice formation exerts a significant impact on food sources for energy refueling during migration (Whooper Swans mainly feed on the roots of water plants at shallow depths; Shimada et al., 2014), it is inevitable that food availability is limited (i.e., the ice prevents the use of food or the food is scarce even after the ice melts) (Petrie and Wilcox, 2003; Nuijten et al., 2014). Thus, premature migration to a stopover or breeding site poses a challenge to energy acquisition with a considerable impact on not only the breeding success but also the survival of Whooper Swans (Bauer et al., 2006; Nuijten et al., 2014; Li et al., 2020). As such, owing to the substantial decrease in food availability caused by ice formation, migrating Whooper Swans presumably take more precautions in spring than in autumn, to strike a balance between achieving sufficient energy acquisition and competing for breeding territories (Bauer et al., 2006; Nuijten et al., 2014; Ao et al., 2020).
The results of this study shed some light on the detailed mechanisms behind the slower spring migration of Whooper Swans; the birds appear to increase their migration distance to decrease their migration speed, while increasing both the stopover duration and number. In general, migratory birds are known to optimize stopover duration and number based on seasonal food availability (Alerstam and Lindström, 1990; Sanz et al., 2003; Baker et al., 2004; Becker et al., 2008; Roques et al., 2022). These characteristics also underlie the strategy of using numerous variable stopover sites (Loria and Moore, 1990; Martin and Karr, 1990; Moore, 1992; Mabey et al., 1993). Whooper Swans are presumed to increase not only the time spent at stopover sites, but also the number of stopover sites used, to maximize their energy refueling capacity. In terms of food acquisition, it might be more stable and effective to remain at a single site to accumulate energy before migrating to the breeding site; however, in terms of competition for breeding territories, it is advantageous for the birds to arrive at the breeding site as early as possible through continuous migration (northward, in the case of Whooper Swans) (Ao et al., 2020). Thus, Whooper Swans are presumed to determine a suitable stopover duration and number of stops depending on the stopover sites that are accessible during continuous northward migration.
Although we should be prudent about inferring and concluding too much from just six individuals, this study is significant in identifying new migration routes for Whooper Swans that winter in South Korea, using GPS trackers. Furthermore, distinct variations were found between the spring and autumn migrations. The findings are anticipated to provide valuable data for conservation and management policies for Whooper Swans wintering in South Korea, while offering critical evidence for the development of detailed management plans for the conservation of crucial habitats along the flyways of these long-distance migratory birds (wintering sites, stopover sites, and breeding sites in this study). Moreover, regarding the persisting problem of avian influenza, this data will be essential in predicting potential routes of transmission based on the migration routes of Whooper Swans, which may be vectors of avian influenza in South Korea. Future tracking of more individuals is required to identify the important habitat characteristics, selection, and utilization of this species.
Authors' contributions
The study was conceptualized by JY Lee, HK Nam, WH Hur and JC Yoo. The fieldwork was done by JY Park, SG Kang, N Batbayar, O Tsend, T Natsagdorj, J Nergui and DW Kim. The data and statistical analyses were conducted by JY Lee, HK Nam, JW Hwang. A first draft was written by JY Lee and HK Nam, and commented by all authors. All authors read and approved the final manuscript.
Ethics statement
Research protocols were approved by the Mongolian Ministry of Environment and Tourism (license number: 06/2862).
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper
Acknowledgements
We would like to thank all those who have helpfully cooperated with us in this study, especially Nyambayar Batbayar's entire Mongolian field team for fieldwork.
Alerstam, T., 2006. Strategies for the transition to breeding in time-selected migration. Ardea 94, 347–357.
Alerstam, T., Hedenström, A., Åkesson, S., 2003. Long-distance migration: evolution and determinants. Oikos 103, 247–260. .
Alerstam, T., Lindström, Å., 1990. In: Gwinner, E. (Ed.), Optimal bird migration: the relative importance of time, energy, and safety, Bird Migration. Springer, Berlin, pp. 331–351.
Ao, P., Wang, X., Meng, F., Batbayar, N., Moriguchi, S., Shimada, T., et al., 2020. Migration routes and conservation status of the Whooper Swan Cygnus cygnus in East Asia. Wildfowl 6, 43–72.
Baker, A.J., González, P.M., Piersma, T., Niles, L.J., do Nascimento Ide, L., Atkinson, P.W., et al., 2004. Rapid population decline in red knots: fitness consequences of decreased refuelling rates and late arrival in Delaware Bay. Proc. Biol. Sci. 271, 875–882. .
Barton, K., Barton, M.K., 2015. Package “Mumin”. R package version 1.18. .
Bates, D., Maechler, M., Bolker, B., Walker, S., 2014. lme4: linear mixed-effects models using Eigen and S4_. R package. version 1.1-7. .
Bauer, S., Madsen, J., Klaassen, M., 2006. Intake rates, stochasticity, or onset of spring: what aspects of food availability affect spring migration patterns in Pink-footed Geese Anser brachyrhynchus? Ardea 94, 555–566. .
Becker, P.H., Dittmann, T., Ludwigs, J.D., Limmer, B., Ludwig, S.C., Bauch, C., et al., 2008. Timing of initial arrival at the breeding site predicts age at first reproduction in a long-lived migratory bird. Proc. Natl. Acad. Sci. USA 105, 12349–12352. .
Beekman, J.H., Nolet, B.A., Marcel, K., 2002. Skipping swans: fuelling rates and wind conditions determine differential use of migratory stopover sites of Bewick's Swans Cygnus bewickii. Ardea 90, 437–460.
Benhamou, S., 2004. How to reliably estimate the tortuosity of an animal's path: straightness? J. Theor. Biol. 229, 209–220. .
Boiko, D., Kamp-Persson, H., Morkŭnas, J., 2014. Breeding Whooper Swans Cygnus cygnus in the baltic states, 1973–2013: result of a 207 recolonization. Wildfowl 64, 207–216.
Bromley, R.G., Jarvis, R.L., 1993. The energetics of migration and reproduction of Dusky Canada Geese. Condor 95, 193–210. .
Bruderer, B., Salewski, V., 2009. Lower annual fecundity in long-distance migrants than in less migratory birds of temperate Europe. J. Ornithol. 150, 281–286. .
Bêty, J., Giroux, J.F., Gauthier, G., 2004. Individual variation in timing of migration: causes and reproductive consequences in greater snow geese (Anser caerulescens atlanticus). Behav. Ecol. Sociobiol. 57, 1–8. .
Burnham, K.P., Anderson, D.R., 2002. Model Selection and Multimodel Inference: a Practical Information-Theoretic Approach, second ed. Springer-Verlag, New York.
Bustnes, J.O., Moe, B., Helberg, M., Phillips, R.A., 2013. Rapid long-distance migration in Norwegian Lesser Black-backed Gulls Larus fuscus fuscus along their eastern flyway. Ibis 155, 402–406. .
Champely, S., 2018. PairedData: paired data analysis. R package version 1.1.1. .
Choi, J., Kim, J.Y., Do, Y., Joo, G.J., 2018. Population trends of wintering whooper swan (Cygnus cygnus) in South Korea: data from the winter water-bird census program. Korean J. Ecol. Environ. 51, 365–372. .
Deng, X., Zhao, Q., Fang, L., Xu, Z., Wang, X., He, H., et al., 2019. Spring migration duration exceeds that of autumn migration in far East Asian greater White-fronted geese (Anser albifrons). Avian Res. 10, 19.
Ely, C.R., Douglas, D.C., Fowler, A.C., Babcock, C.A., Derksen, D.V., Takekawa, J.Y., 1997. Migration behavior of Tundra Swan form the Yukon-Kuskokwim Delta, Alaska. Wilson Bull. 109, 679–692.
Ely, C.R., Meixell, B.W., 2016. Demographic outcomes of diverse migration strategies assessed in a metapopulation of tundra swans. Mov. Ecol. 4, 10. .
Fox, J., Weisberg, S., 2011. An R Companion to Applied Regression. Sage, Thousand Oaks, CA.
Fransson, T., 1995. Timing and speed of migration in North and West European populations of Sylvia warblers. J. Avian Biol. 26, 39–48. .
Higuchi, H., Sato, F., Matsui, S., Soma, M., Kanmuri, N., 1991. Satellite tracking of the migration routes of Whistling Swans Cygnus columbianus. J. Yamashina Inst. Ornithol. 23, 6–12. .
Kamiya, K., Ozaki, K., 2002. Satellite tracking of Bewick's Swan migration from Lake Nakaumi, Japan. Waterbirds 25, 128–131.
Kear, J., 2005. Ducks, Geese and Swans: Species Accounts (Cairina to Mergus), vol. 2. Oxford University Press, Oxford.
Klaassen, R.H., Alerstam, T., Carlsson, P., Fox, J.W., Lindström, A., 2011. Great flights by great snipes: long and fast non-stop migration over benign habitats. Biol. Lett. 7, 833–835. .
Klaassen, R.H.G., Ens, B.J., Shamoun-Baranes, J., Exo, K.M., Bairlein, F., 2012. Migration strategy of a flight generalist, the Lesser black-backed Gull Larus fuscus. Behav. Ecol. 23, 58–68. .
Kokko, H., 1999. Competition for early arrival in migratory birds. J. Anim. Ecol. 68, 940–950. .
Kölzsch, A., Müskens, G.J.D.M., Kruckenberg, H., Glazov, P., Weinzierl, R., Nolet, B.A., et al., 2016. Towards a new understanding of migration timing: slower spring than autumn migration in geese reflects different decision rules for stopover use and departure. Oikos 125, 1496–1507. .
Kutner, M.H., Nachtsheim, C.J., Neter, J., Li, W., 2004. Applied Linear Statistical Models, fifth ed. McGraw-Hill, Chicago. Irwin.
Li, H., Fang, L., Wang, X., Yi, K., Cao, L., Fox, A.D., 2020. Does snowmelt constrain spring migration progression in sympatric wintering Arctic-nesting geese? Results from a Far East Asia telemetry study. Ibis 162, 548–555. .
Li, S., Meng, W., Liu, D., Yang, Q., Chen, L., Dai, Q., et al., 2018. Migratory Whooper Swans Cygnus cygnus transmit H5N1 virus between China and Mongolia: combination evidence from satellite tracking and Phylogenetics analysis. Sci. Rep. 8, 7049. .
Limiñana, R., Soutullo, A., Urios, V., Reig-Ferrer, A., 2012. Migration and wintering areas of adult Montagu's Harriers (Circus pygargus) breeding in Spain. J. Ornithol. 153, 85–93. .
Loria, D.E., Moore, F.R., 1990. Energy demands of migration on red-eyed vireos, Vireo olivaceus. Behav. Ecol. 1, 24–35. .
Mabey, S.E., McCann, J., Niles, L.J., Bartlett, C., Kerlinger, P., 1993. The Migratory Songbird Coastal Corridor Final Report. Virginia Department of Environmental Quality Coastal Resources Management Program, Richmond, VA. Report No. NA90AA-HCZ839.
Martin, T.E., Karr, J.R., 1990. Behavioral plasticity of foraging maneuvers of migratory warblers: multiple selection periods for niches. Stud. Avian Biol. 13, 353–359.
Mathiasson, S., 2013. Eurasian Whooper swan Cygnus cygnus migration with particular reference to birds wintering in southern Sweden. Wildfowl 1, 201–208.
McNamara, J.M., Welham, R.K., Houston, A.I., 1998. The timing of migration within the context of an annual routine. J. Avian Biol. 29, 416–423. .
Mellone, U., De La Puente, J., López-López, P., Limiñana, R., Bermejo, A., Urios, V., 2015. Seasonal differences in migration patterns of a soaring bird in relation to environmental conditions: a multi-scale approach. Behav. Ecol. Sociobiol. 69, 75–82. .
Moore, F.R., 1992. Ecophysiological and behavioral response to energy demand during migration. Acta Congr. Int. Onthil. 20, 753–760.
Moore, F.R., Smith, R.J., Sandberg, R., 2005. In: Greenberg, R., Marra, P. (Eds.), Stopover ecology of intercontinental migrants: en route problems and consequences for reproductive performance, Birds of Two Worlds: the Ecology and Evolution of Migration. Johns Hopkins Press, Baltimore, pp. 251–261.
Mosbech, A., Gilchrist, G., Merkel, F., Sonne, C., Flagstad, A., Nyegaard, H., 2006. Year-round movements of northern Common Eiders Somateria mollissima borealis breeding in Arctic Canada and West Greenland followed by satellite telemetry. Ardea 94, 651–665.
National Institute of Biological Resources, 2021. 2020–2021 Winter Water-Bird Census of Korea. National Institute of Biological Resources, Republic of Korea (in Korean).
Nilsson, C., Klaassen, R.H., Alerstam, T., 2013. Differences in speed and duration of bird migration between spring and autumn. Am. Nat. 181, 837–845. .
Nolet, B.A., 2006. Speed of spring migration of Tundra Swans Cygnus columbianus in accordance with income or capital breeding strategy? Ardea 94, 579–591.
Nowak, E., Berthold, P., Querner, U., 1990. Satellite tracking of migrating Bewick's swans. A European pilot study. Naturwissenschaften 77, 549–550. .
Nuijten, R.J.M., Kölzsch, A., Van Gils, J.A., Hoye, B.J., Oosterbeek, K., De Vries, P.P., et al., 2014. The exception to the rule: retreating ice front makes Bewick's swans Cygnus columbianus bewickii migrate slower in spring than in autumn. J. Avian Biol. 45, 113–122. .
Peterson, B.G., Carl, P., 2014. PerformanceAnalytics: econometric tools for performance and risk analysis. .
Petrie, S.A., Wilcox, K.L., 2003. Migration chronology of eastern-population tundra swans. Can. J. Zool. 81, 861–870. .
R Core Team, 2019. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing.
Rahn, H., Ar, A., 1974. The avian egg: incubation time and water loss. Condor 76, 147–152. .
Rees, E.C., Cao, L., Clausen, P., Coleman, J.T., Cornely, J., Einarsson, O., et al., 2019. Conservation status of the world's swan populations, Cygnus sp. and Coscoroba sp. : a review of current trends and gaps in knowledge. Wildfowl 5, 35–72.
Roques, S., Henry, P.-Y., Guyot, G., Bargain, B., Cam, E., Pradel, R., 2022. When to depart from a stopover site? Time since arrival matters more than current weather conditions. Ornithology 139. ukab057.
Sanz, J.J., Potti, J., Moreno, J., Merino, S., Frias, O., 2003. Climate change and fitness components of a migratory bird breeding in the Mediterranean region. Global Change Biol. 9, 461–472. .
Shamoun-Baranes, J., Baharad, A., Alpert, P., Berthold, P., Yom-Tov, Y., Dvir, Y., et al., 2003. The effect of wind, season and latitude on the migration speed of white storks Ciconia ciconia, along the eastern migration route. J. Avian Biol. 34, 97–104. .
Shimada, T., Yamaguchi, N.M., Hijikata, N., Hiraoka, E., Hupp, J.W., Flint, P.L., et al., 2014. Satellite tracking of migrating Whooper Swans Cygnus cygnus wintering in Japan. Ornithol. Sci. 13, 67–75. .
Van Noordwijk, A.J.V., McCleery, R.H., Perrins, C.M., 1995. Selection for the timing of great tit breeding in relation to caterpillar growth and temperature. J. Anim. Ecol. 64, 451–458. .
Yohannes, E., Biebach, H., Nikolaus, G., Pearson, D.J., 2009. Migration speeds among eleven species of long-distance migrating passerines across Europe, the desert and eastern Africa. J. Avian Biol. 40, 126–134. .
Table
2.
Define of migration parameters of Whooper Swans.
Variable
Descript
Unit
Migration duration
Total length of the spring or autumn migration including the time spent at stopovers
days
Migration distance
Total distance traveled between the wintering and breeding sites, excluding distances within the wintering, breeding, and stopover sites—it was calculated as the sum of the distances between locations along a continuous path and not as a linear distance connecting the start and end points
km
Stopover duration
The sum of the days spent at each stopover site during the spring and autumn migrations
days
Number of stopovers
The number of stopover sites where the bird spent at least three days during their spring or autumn migrations
–
Daily travel speed
The value calculated by dividing migration distance by travel duration
km/day
Travel duration
The value calculated difference between migration duration and stopover duration
days
Migration speed
The value calculated by dividing migration distance by migration duration
km/day
Straightness index
The value calculated by dividing the linear distance between the start and end points by the migration distance (Benhamou, 2004)
Table
4.
Candidate model, number of parameters (K), Akaike's information criterion (AIC), difference from lowest AIC value (ΔAIC), and Akaike model weight used to determine factors influencing migration season (spring and autumn) of Whooper Swans.
Parameter
K
logLik
AIC
ΔAIC
Weight
Migration distance + number of stopovers
5
0.35
12.15
0.00
0.22
Number of stopovers
4
−1.46
12.75
0.60
0.16
Migration distance + number of stopovers + migration speed
6
1.67
12.85
0.70
0.15
Number of stopovers + migration speed
5
−0.10
13.06
0.91
0.14
Number of stopovers + migration speed + straightness index
6
1.39
13.42
1.26
0.12
K: number of parameters; AIC: Akaike's information criterion; ΔAIC: difference from lowest Akaike's information criterion value.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds