Cheng WEN, Dong HAN. 2013: Raptor migration monitoring in the spring of 2009 at Baiwangshan, Beijing. Avian Research, 4(4): 319-327. DOI: 10.5122/cbirds.2013.0031
Citation:
Cheng WEN, Dong HAN. 2013: Raptor migration monitoring in the spring of 2009 at Baiwangshan, Beijing. Avian Research, 4(4): 319-327. DOI: 10.5122/cbirds.2013.0031
Cheng WEN, Dong HAN. 2013: Raptor migration monitoring in the spring of 2009 at Baiwangshan, Beijing. Avian Research, 4(4): 319-327. DOI: 10.5122/cbirds.2013.0031
Citation:
Cheng WEN, Dong HAN. 2013: Raptor migration monitoring in the spring of 2009 at Baiwangshan, Beijing. Avian Research, 4(4): 319-327. DOI: 10.5122/cbirds.2013.0031
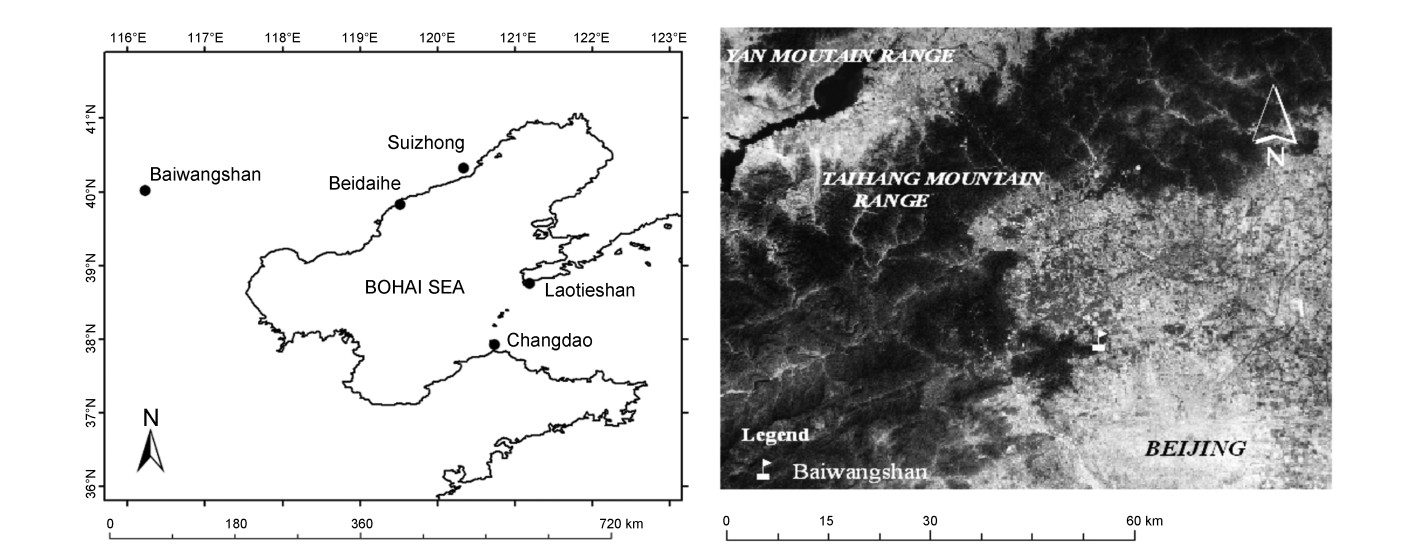
Following a series of surveys since 2003,a full-seasonal raptor count was undertaken at Baiwangshan,a mountain in the vicinity of Beijing,from 6 March to 22 May 2009, when 3802 individual birds of 23 species were recorded. Four species,i.e.,Oriental Honey Buzzard (Pernis ptilorhynchus),Common Buzzard (Buteo buteo),Eurasian Sparrow Hawk (Accipiter nisus) and Gray-faced Buzzard (Butastur indicus) together accounted for up 90% of the total and P. ptilorhynchus alone for 70%. Raptor passage of such magnitude and diversity at an inland site in China was unheard of up to then. As well,113 other migrants were recorded at the site during this period,including 24 water birds,74 passerines and 15 others (cuckoos,owls and other migrants).
Global biodiversity is facing increasing threats due to climate change and anthropogenic impacts (Myers et al., 2000; Grenyer et al., 2006). Identification of important species, under risk of extinction, would be of great help to biologists in a more efficient allocation of limited funds to improve and conserve biological diversity by focusing on these unique species (Grenyer et al., 2006). A common practice to identify conservation values of species is to evaluate the rarity of the population of a species, its distribution ranges (He, 2012) and changes in habitat conditions as is practised in the IUCN Red List (The World Conservation Union, 2010). In recent years, the importance of evolutionary history has become recognized (Cadotte and Davies, 2010; Martyn et al., 2012) and many phylogenetic diversity (PD) indices for quantifying evolutionary heritage of species have been proposed in a number of studies (Faith, 1992, 2002; Faith et al., 2004; Tucker et al., 2012).
The avifauna of China is composed of around 1314 species of which 52 are said to be endemic (only found within the political boundaries of China, i.e., the mainland and Taiwan) (Lei and Lu, 2006). Conservation priorities of these endemic and other bird species in China have been widely established in previous studies (Lei et al., 2002a, 2007; Chen, 2007, 2008; Chang et al., 2013). However, the importance of conservation of endemic birds of China from a phylogenetic perspective has never been evaluated up till now. As such, the present report presents a way to fill such a knowledge gap by proposing conservation priority of endemic birds of China using a series of phylogenetic diversity metrics.
Table
1.
Conservation priorities of species based on alternative phylogenetic diversity metrics. IUCN categories: EN (endangered), VU (vulnerable), NT (near threatened), LC (least concerned), DD (data deficient). "-" denotes the IUCN information is not available. Codes for phylogenetic diversity metrics: ES (equal split), FP (fair proportions), ED (evolutionary distinctiveness), TD (taxonomic distinctiveness), PL (pendant edge's length) and Node-based I and W indices.
The phylogenetic relationships between the 52 endemic birds of China's mainland are extracted from the BirdTree.org database (http://www.birdtree.org), which is derived from a full phylogeny of the global bird species in a previous study (Jetz et al., 2012). However one species, Ficedula beijingnic, was omitted from the tree files of the database. It was therefore decided to exclude this species for further analyses, while the remaining 51 species were used for analysis since these are all included in the retrieved phylogenetic trees. From the 3000 trees tested for possible phylogenetic affinities, the 51 endemic birds were retrieved and the resultant consensus tree, with average branch lengths, was obtained using the DendroPy python library (Sukumaran and Holder, 2010). Molecular dating of the tree was carried out using a penalized likelihood method (Sanderson, 2002). The result, a dated tree (Fig. 1), is used for all subsequent analyses.
Figure
1.
Consensus phylogenetic tree for 51 endemic birds of mainland China
As a comparison, the category of the IUCN Red list for each species was collected from BirdLife International (http://www.birdlife.org/) and included in Table 1. The abbreviations for each category are as follows: EN (endangered), VU (vulnerable), NT (near threatened), LC (least concerned) and DD (data deficient). When comparing the rankings of IUCN and PD indices, the IUCN categories are transformed into discrete integers, i.e., EN (1), VU (2), NT (3), LC (4) and DD (5). The Wilcox signed rank test and Pearson's correlation test were implemented to compare both rankings. A significant Wilcox signed rank or a non-significant Pearson's correlation coefficient implies that both rankings are fundamentally different.
Results
As shown in Table 1, the top five endemic birds based on the combined rankings of seven PD indices in order of priority, are Arborophila ardens, A. gingica, A. rufipectus, Lophophorus lhuysii and Alectoris magna. Their corresponding IUCN classes are VU, NT, EN, VU and LC, respectively. As seen, the PD-based ranking accurately identified the conservation importance of Arborophilarufipectus, an endangered species that should have a high priority for conservation.
Arborophila rufipectus has a very limited range of distribution in the southern part of Sichuan Province. The size of its adult population is estimated to be around 1000 (BirdLife International, http://www.birdlife.org/). Due to continuous hunting and habitat loss, it was classified as Critically Endangered in previous IUCN reports(1994, 1998). It is now still listed in the category of Endangered species. Based on a phylogenetic perspective this species, along with two other Arborophila species, has a very unique and long evolutionary history, when compared to other endemic birds (Fig. 1).
The PD-ranking for Arborophila ardens and Lophophorus lhuysii are also fairly accurate (top-1 and 4), both of which are listed as VU in the IUCN Red List (The World Conservation Union, 2010). This implies that both should deserve high conservation priority from an ecological perspective (based on IUCN ranking) as well as from an evolutionary (based on PD ranking) perspective.
In contrast, there are some inconsistencies between PD and IUCN rankings. For example, Arborophilagingica and Alectoris magna are ranked 2nd and 5th in the PD ranking for their unique evolutionary histories. However, in the IUCN ranking, these species are merely grouped into the categories of NT and LC.
Overall, the difference of rankings based on the IUCN Red List and the combined PD ranking is statistically significant (Wilcox signed rank test: V = 159, p < 0.001; Pearson's correlation coefficient: r = 0.217, p = 0.129). These tests show a significant difference between IUCN and PD rankings for endemic birds of mainland China.
Discussion
Conservation importance of endemic or rare species has been widely recognized (Linder, 1995; Lamoreux et al., 2006; Gaston, 2012). In the present study, I quantified the conservation importance of avian species, endemic to China, by utilizing a variety of phylogenetic diversity metrics. The results show a statistically significant difference between the priority rankings based on PD metrics and that derived from the IUCN Red list. Therefore, a PD-based conservation emphasis on endemic birds of China might offer some new views when establishing relevant conservation strategies by considering evolutionary heritage and genetic resources of these species.
The present study carried out analyses on 51 endemic birds. However, it might be a bit ambiguous given the number of endemic birds in China (Zhang, 2004), when one considers migration during the breeding season (Lei et al., 2002b). In a previous study, the number of endemic birds in China was believed to be around 100 (Lei et al., 2002a), while in a more recent publication, this number is said to be 105 (Lei and Lu, 2006). If the definition of endemic birds were extended to include, for example, Taiwan and Hong Kong, the number of endemic birds should be at least 70 (Zhang, 2004; Lei et al., 2002b). When considering only mainland China, the number of endemic birds should be at least 50, based on the information from the World Bird Database (http://avibase.bsc-eoc.org/). Therefore, the present study should be relatively accurate in suggesting conservation priorities for endemic birds in mainland China based on the phylogenetic diversity framework. Any re-analyses with the addition of a few more possible endemic birds should not greatly affect the quantitative results presented in the present study.
In this study, I have only used pure phylogeny-based diversity indices without considering other weighted phylogenetic diversity indices. Weighted phylogenetic diversity metrics such as the abundance-weighted PD index (Cadotte et al., 2010), the biogeography-weighted PD index (Tucker et al., 2012), the endemism-weighted PD index (Rosauer et al., 2009) and possibly other indices might provide more insights into conservation priorities because these can explicitly incorporate the role of some biological factors when studying the evolutionary history of species. For future implications, integration of other weighted phylogenetic diversity indices into the systematic conservation planning of birds would offer new insights and thus should be considered in the next levels of research.
Acknowledgements
This work was supported by the University of British Columbia and now supported by China Scholarship Council.
Agostini N, Mellone U. 2007. Migration strategies of Oriental Honey-buzzards Pernis ptilorhyncus breeding in Japan. Forktail, 23: 182–183.
ARRCN. 2008. Results of cooperative study of aisan raptor migration in Japan. The Hazy Project Report.
Ash JS. 1993. Raptor migration on Bali, Indonesia. Forktail, 9: 3–11.
Besten JWD. 2004. Migration of Steppe Eagles Aquila nipalensis and other raptors along the Himalayas past Dharamsala, India, in autumn 2001 and spring 2002. Forktail, 20: 9–13.
China Ornithological Society. 2004–2007. China Bird Report 2004–2007. China Ornithological Society. Beijing, China.
DeCandido R, Allen D, Bildstein KL. 2006. Spring Migration of Oriental Honey-buzzards Pernis ptilorhyncus and other raptors at Tanjung Tuan, Malaysia, 2000-01. Forktail, 22: 152–160.
DeCandido R, Allen D, Yosef R. 2005. Comparison of spring migration phenology of Bee-eaters and Oriental Honey-buzzards Pernis ptilorhyncus at Tanjung Tuan, Malaysia, 2000-01. Ardea, 92(2): 169–174.
DeCandido R, Nualsri C, Allen D, Bildstein KL. 2004. Autumn 2003 raptor migration at Chumphon, Thailand: a globally significant raptor migration watch site. Forktail, 20: 49–54.
Ellis DH, Kepler AK, Kepler CB. 1990. Evidence for a fall raptor migration pathway across the South China Sea. J Raptor Res, 24(1): 12–18.
Fan P, Zhong HB, Zhao F, Fan QJ. 2006. Research on raptor banding at Changshan Archipelago. Shandong Forest Sci Technol, 2006(3): 43–45. (in Chinese)
Fan QD, Sun WL, Yuan YT, Zhao F. 1992. Research on raptor banding at Changshan Archipelago, Shangdong. Sichuan J Zool, 11(1): 16–19. (in Chinese)
Ferguson-Lees J, Christie DA. 2001. Raptors of the World. Christopher Helm, London.
Fuller M R, Mosher J A. Methods of detecting and counting raptors: a review. Stud Avian Biol, 1981, 6: 235–246.
Germi F, Waluyo D. 2006. Additional information on the autumn migration of raptors in east Bali, Indonesia. Forktail, 22: 71–76.
Germi F. 2005. Raptor migration in east Bali, Indonesia: observations from a bottleneck watch site. Forktail, 21: 93–98.
Higuchi H, Shiu HJ, Nakamura H, Uematsu A, Kuno K, Saeki M, Hotta M, Tokita K, Moriya E, Morishita E, Tamura M. 2005. Migration of Honey-buzzards Pernis ptilorhyncus based on satellite tracking. Ornithol Sci, 4: 109–115.
Lang CQ, Zhang XQ, Shi RY, Xue HZ. 2003. Raptor census in Shanxi. Sichuan J Zool, 22(3): 151–152. (in Chinese)
Li XJ. 2005. Status and biodiversity conservation on raptors in the mountainous area around Beijing. Dissertation. Beijing Forestry University, Beijing. (in Chinese)
Lin WH, Severinghaus LL. 1998. Raptor migration and conservation in Taiwan. In: Chancellor RD, Meyburg BU, Ferrero JJ (eds) Holarctic Birds of Prey. ADENEX and World Working Group on Birds of Prey, Merida Spain, pp 631–639.
Mackinnon JR, Phillipps K, He FQ. 2000. A Field Guide to the Birds of China. Hunan Education Press, Changsha.
Nijman V, Germi F, van Balen B. 2006. Relative status of two species of migrant sparrowhawks on Java and Bali, Indonesia. Emu, 106(2): 157–162.
Nijman V. 2001a. Autumn migration of raptors on Java, Indonesia: composition, direction and behaviour. Ibis, 143: 99–106.
Nijman V. 2001b. Spatial and temporal variation in migration raptors on Java, Indonesia. Emu, 101: 259–263.
Nijman V. 2004. Magnitude and timing of migrant raptors in central Java, Indonesia. Ardea, 92(2): 161–168.
Niu HX, Lu JQ, Lu JQ, Pu YZ, Zhang XF, Zhang KY, Zhu JG. 2002. Survey on Carnivorous birds in Henan Province. Chin J Zool, 37(1): 36–38.
Nuraeni S, Suparman U. 2000. Raptor migration at Pumcak and Cibulao, West Java, Indonesia: October and November 1999. Asian Rap Bull, 1: 7–9.
Ran JC. 1998. A preliminary study on ann-dynamic of bird of prey in Maolan region. Chin Ornithol Res, 253–257. (in Chinese)
Shiu HJ, Tokita K, Morishita E, Hiraoka E, Wu Y, Nakamura H, Higuchi H. 2006. Route and site fidelity of two migratory raptors: Grey-faced Buzzards Butastur indicus and Honey-buzzards Pernis apivorus. Ornithol Sci, 5: 151–156.
Sukmantoro W, Suparman U, Wen Sin L. 2005. A study on migratory raptors in Sumatra, Kalimantan, Java, Bali and Musa Tenggara, Indonesia: 2001–2004. Report presented at the Fourth Symposium on Asian Raptors. ARRCN and Malaysian Nature Society, Taiping, Malaysia.
Wan DM, Gao W, Zhao J, Wang HT, Liu MY. 2002. Research on the migration law of raptor in Liaoning Province. J Northeast Normal Univ, 34(2): 78–83. (in Chinese)
Yue F, Luo ZK, Wu FQ, Shu S, Wu SB, Wang TH. 2008. Migrant time new records of three harriers in Hubei Province. Forest Invent Plan, 33(5): 31–32. (in Chinese)
Table
1.
Conservation priorities of species based on alternative phylogenetic diversity metrics. IUCN categories: EN (endangered), VU (vulnerable), NT (near threatened), LC (least concerned), DD (data deficient). "-" denotes the IUCN information is not available. Codes for phylogenetic diversity metrics: ES (equal split), FP (fair proportions), ED (evolutionary distinctiveness), TD (taxonomic distinctiveness), PL (pendant edge's length) and Node-based I and W indices.

 DownLoad:
DownLoad:






 Email Alerts
Email Alerts RSS Feeds
RSS Feeds