Giampiero Sammuri, Guido Alari Esposito, Giuseppe Anselmi, Francesco Pezzo, Vincenzo Rizzo Pinna, Andrea Sforzi, Flavio Monti. 2025: Moving away from home: Early life movements and space use in juvenile Montagu's Harriers (Circus pygargus) from central Italy. Avian Research, 16(1): 100239. DOI: 10.1016/j.avrs.2025.100239
Citation:
Giampiero Sammuri, Guido Alari Esposito, Giuseppe Anselmi, Francesco Pezzo, Vincenzo Rizzo Pinna, Andrea Sforzi, Flavio Monti. 2025: Moving away from home: Early life movements and space use in juvenile Montagu's Harriers (Circus pygargus) from central Italy. Avian Research, 16(1): 100239. DOI: 10.1016/j.avrs.2025.100239
Giampiero Sammuri, Guido Alari Esposito, Giuseppe Anselmi, Francesco Pezzo, Vincenzo Rizzo Pinna, Andrea Sforzi, Flavio Monti. 2025: Moving away from home: Early life movements and space use in juvenile Montagu's Harriers (Circus pygargus) from central Italy. Avian Research, 16(1): 100239. DOI: 10.1016/j.avrs.2025.100239
Citation:
Giampiero Sammuri, Guido Alari Esposito, Giuseppe Anselmi, Francesco Pezzo, Vincenzo Rizzo Pinna, Andrea Sforzi, Flavio Monti. 2025: Moving away from home: Early life movements and space use in juvenile Montagu's Harriers (Circus pygargus) from central Italy. Avian Research, 16(1): 100239. DOI: 10.1016/j.avrs.2025.100239
Assessing individual differences and variability in animal movement patterns is essential to improve our understanding of the evolution and ontogeny of migratory strategies. In long-distance migratory species, fledged juveniles often rely on an extremely restricted time span to learn the essential skills for survival and to prepare for migration, possibly the most risky phase of their lives. Collecting detailed information on the dynamics of the movements during the crucial pre-migratory phase is hence essential to understand the solutions developed by migratory species in different environmental contexts. Here, we used high-resolution GPS/GSM transmitters to collect information on the movement ecology of seven juvenile Montagu's Harriers (Circus pygargus) born in central Italy, investigating their early life stages, namely the post-fledging dependence period (PFDP) and the pre-migratory phase (PMP), until autumn migration. After fledging, individuals showed high variability (both in space and time) in home range size, daily distances covered (6.88 ± 11.44 km/day), distance from the nest (1.45 ± 2.8 km) and PFDP length (23.3 ± 5.3 days). Residence time at the natal site significantly decreased, while time interval between revists in the natal area significantly increased, as the PFDP progressed. During the PMP, explored areas and distance from the nest (max value up to 320.8 km) varied among individuals, despite daily distances covered (27 ± 40 km/day) and time allocation between traveling (60.7%) and foraging (39.3%) were similar across individuals. The PMP lasted 38 ± 14 days. Land cover composition of foraging locations was mostly represented by agricultural lands (~78.2%), though habitat use differed among individuals. More than 76% of such locations were outside protected areas. This individual-based tracking study represents a novel approach that improves previous knowledge based on field studies on the early life stages of the Montagu's Harrier. High inter-individual variability in movement patterns, broad-range exploratory movements and foraging locations outside the protected area network make the application of standard conservation measures difficult, raising concerns about the long-term preservation of this vulnerable migratory species in Italy.
Among migratory birds, long-distance migratory species are particularly constrained by annual migration programs, which in turn can influence other stages of the annual cycle (Newton, 2010). While adults can count on previous experience in shaping their movement strategies (i.e., by defining departure dates for migration, the duration of the breeding and pre-migratory phase; e.g., Sergio et al., 2014; Vansteelant et al., 2015), juveniles are naive and inexperienced (e.g., Harel et al., 2016; Monti et al., 2018). Immediately after fledging, they enter a risky phase during which they must develop essential flight skills (Harel et al., 2016; Ruaux et al., 2020) and quickly become independent to survive and proficiently prepare themselves for the autumn migration (e.g., Kitowski, 2009; Cavalcante et al., 2019). In migratory birds this phase can be relatively short (e.g., it can last from a few weeks to months, depending on the species; e.g., Newton, 2010) and it is usually made by two successive transitory stages that precede autumn migration: i) a period of dependence on parents, which spans from the first flight until bird's departure from the natal territory, referred to as the post-fledging dependence period (hereafter, PFDP) (e.g., Bustamante and Hiraldo, 1989; Arroyo et al., 2002a; Hemery et al., 2023); and ii) an independence phase during which juveniles are no more tied to the nest site and their parents and learn how to hunt and feed autonomously, until migration (e.g., Arroyo et al., 2002a). During this so called pre-migratory phase (hereafter, PMP), ranging movements are usually undertaken (Todd et al., 2007; Strandberg et al., 2008) and energy reserves stored in preparation for migration (Berthold, 1996). Both these phases can highly vary, in space and time, and differ between species, sexes (Newton, 2011; Krupiński et al., 2021), or even between individuals of the same species (e.g., Amar et al., 2000; Arroyo et al., 2002a; Monti et al., 2012). Hemery et al. (2023) stressed that it would be important to investigate such variability at the inter-individual level, and especially among individuals belonging to the same population, as it can reveal key ecological and behavioral adaptations to local environmental conditions. This variability can be expressed at multiple levels, affecting different behavioral parameters such as: time spent at the nest, number of revisits to the natal area, daily distances traveled, home range size, as well as the extent and duration of exploratory movements before migration (e.g., Züst et al., 2023). Furthermore, this stage could be highly risky, as inexperienced individuals wandering across unknown localities are more prone to encounter multiple threats on their way, ultimately increasing their probability of mortality (Amar et al., 2000; Oltra et al., 2024). Investigating how individuals move and which areas they visit during these stages is crucial not only for a better understanding of individual choices and behaviour, but also to detect potential threats and plan corrective measures aiming at species conservation (e.g., Allen and Singh, 2016; Jahn et al., 2017).
In this respect migratory raptor populations are especially vulnerable to high mortality rates they experience during their complex annual cycle, and particularly sensitive to human-related threats (e.g., Bildstein, 2006; Klaassen et al., 2014; Serratosa et al., 2024). Historically, large declines in migratory raptor populations have been documented in many parts of the world, including the Western Palearctic (Newton, 2004; De Pascalis et al., 2020). For this reason they still represent nowadays a conservation priority, with some populations deserving specific management and research efforts (e.g., McClure et al., 2018).
We focussed on the Montagu's Harrier (Circus pygargus), a long-distance migratory raptor, breeding in Europe, North Africa (mostly in Morocco; Garrido et al., 2021) and Asia and with wintering grounds located in sub-saharan Africa (mostly in the Sahel belt), as well as in the Indian subcontinent and Sri Lanka (Limiñana et al., 2008; Schlaich and Klaassen, 2021; Ganesh et al., 2024). The species is declining across its range, mainly because of habitat loss and fragmentation (Arroyo and Bretagnolle, 2000; Butet and Leroux, 2001; Arroyo et al., 2002b) and other human-related threats (De Pascalis et al., 2020; Serratosa et al., 2024). Breeding areas in western Europe are mostly associated with agricultural landscapes (e.g., cereal crops; Arroyo et al., 2002a), with nests and clutches subjected to a high risk of destruction during crop harvesting procedures and other agricultural practices, commonly taking place in conjunction with the reproductive season and making the species highly dependent on nest protection measures (Pandolfi, 1996; Arroyo et al., 2002a, 2002b; Gameiro et al., 2024). Some breeding nuclei can also be found in natural habitats (e.g., shrublands; Limiñana et al., 2006), at various altitudes, spanning from coastal zones to hilly and mountain areas (Cramp and Simmons, 1980). In Italy, the Montagu's Harrier has a fragmented distribution (Brichetti and Fracasso, 2018; Ravasini, 2022), with a breeding population estimation of about 260–380 breeding pairs (Ravasini, 2022). The species has historically suffered important declines (Pandolfi and Tanferna, 2009) and it is currently ranked as vulnerable (VU) both at regional (Mediterranean; Westrip et al., 2022) and national level, according to the latest IUCN RedList (Rondinini et al., 2022), though being ranked as Least Concern at global scale. Information available for the species at country level mostly relies on distribution atlas data and behavioral studies dating back to the 90s (e.g., Pandolfi et al., 1990; Pandolfi, 1996; Ravasini, 2022), while no studies on Italian populations focussed on movement ecology yet.
In other countries, such as in Spain and France, previous studies focussed on the PFDP by investigating juveniles' behavioral patterns and relationships with parents at nest (Amar et al., 2000; Arroyo et al., 2002a; Millon et al., 2008). In particular, Arroyo et al. (2002a) found that departure dates and length of the PFDP were mostly driven by food abundance and provisioning rate, as well as affected by parent-offspring conflicts. The PFDP was longer in years with scarce food abundance and for individuals in larger broods (Arroyo et al., 2002a). Similarly, Amar et al. (2000) found that birds in poorer condition had longer periods of dependence. This suggests that movement patterns and phases may largely vary according to ecological factors and even considerably differ at the individual level (e.g., Arroyo et al., 2002a). Furthermore, some studies (Limiñana et al., 2008; Krupiński et al., 2021) stressed that the objective identification of each stage during this period (e.g., the transition between the PFDP and the PMP and/or migration) is not always easy to define, requiring the use of standardized methods. GPS-tracking is in this context a powerful tool to investigate, with a high accuracy, the spatio-temporal patterns of movements, providing integrative information on this complex stage and, ultimately, providing evidences for a better distinction of the different phases (e.g., Berger-Geiger et al., 2022; Arroyo et al., 2024).
In 2024, we deployed high-resolution GPS/GSM transmitters to seven juvenile Montagu's Harriers breeding in central Italy to collect information on their movement ecology throughout the annual cycle. This represents the first tracking study on the species in Italy. The current paper reports the results collected during the period encompassing the PFDP and the PMP, until the onset of autumn migration. We were interested in investigating the spatio-temporal variation in movement patterns during different stages, and in particular to: i) define individual departure dates for each period; ii) describe and quantify home range size and movement parameters during the PFDP; and iii) model the potential effects of the time elapsed since the first flight on the residence time and on the time between successive revisits at the natal area, during the PFDP. As juveniles are assumed to improve their flight and hunting abilities with time (e.g., Arroyo et al., 2002a), we expected a gradual increase in both distance covered and home range estimates, over time. Accordingly, we hypothesize a gradual decrease of the time spent at the natal area (residence time), and an increase in the time interval between successive revisits, as the PFDP progressed. For the PMP, we aimed at: iv) investigating distances covered and key staging areas attended for foraging, as well as the duration of these wandering and relative stays. As reported by Limiñana et al. (2008) for adults, we expected wider movements to distant staging areas, possibly located at higher latitudes than the nest sites as food peaks are occurring there later on during the season. Since these foraging locations are of key importance for preparing juveniles for autumn migration, we also investigated their land cover composition and, for conservation purposes, evaluated their protection status (i.e., inside or outside the protected areas network). Finally, departure dates for autumn migration were recorded, for comparisons with other European populations. In particular, we expected the autumn migration timing to be similar to that of other tagged individuals in Europe that nest at comparable latitudes, such as those in Spain (e.g., Arroyo et al., 2024).
2.
Materials and methods
2.1
Study area
The study area is located in the central-eastern part of the province of Grosseto (southern Tuscany), in the Amiata Mountain area. Altitude ranges from ~500 m above sea level in hilly areas to 1193 m, peaking with the Monte Labbro. Average annual rainfall is between 1128.4 and 1264.4 mm, while the average annual temperature fluctuates between 11.1 and 12.9 ℃ (Giovacchini, 2003). The climate is mesothermic and goes from sub-arid, with moderate winter water surplus, to sub-humid or humid, with moderate summer dryness (Giovacchini, 2003). The area is characterized by xerophytic to semi-mesophytic grasslands in the hilly and low mountain plains and mixed forests in the gorge or valley bottom environments. Part of the study area has been recognized as a site of community importance (SIC/ZPS IT51A0018 "Monte Labbro e Alta Valle dell'Albegna") and several protected areas have been established in the immediate surroundings such as the Monte Labbro Nature Reserves and the Faunistic Park of Monte Amiata. There, cultivated and natural areas are used by Montagu's Harriers for breeding and hunting. Nests were located on the ground in the thick fern and/or bramble vegetation and were found often adjacent to areas with agricultural fields.
2.2
Capture and tracking
In July 2024, we trapped 11 Montagu's Harrier chicks from five nests, during the last phases of the nestling period. Nests were closely located (mean distance between nests: 0.96 ± 0.9 km; min-max: 0.034–1.89 km). All chicks were measured at an age of 25–30 days and marked with a metal ring. Capture, handling, and tagging procedures were carried out under formal authorization issued by the Tuscany Regional Administration, after the positive opinion expressed by the Italian Institute for Environmental Protection and Research (ISPRA), in accordance with Law157/1992 [Art.4 (1) and Art 7 (5)], which regulates research on wild bird and mammal species. Blood and feather samples for genetic analyses to determine the sex of the birds were collected from chicks during ringing activities at nests. Bird handling (from capture to release) lasted in total 30–40 min.
Seven out of eleven individuals were equipped with 10.8 g solar powered GPS/GSM transmitter (model Ornitrack-10, Ornitela, Lithuania). We tagged a chick per nest in three nests, while for each of the other two nests two siblings were equipped. Loggers were mounted through a 4.7 mm wide Teflon ribbon backpack harness. The mass of the tracking device including harness were around 3% and never exceeded 5% of the bird's body mass (percentage of body mass range: 2.3–3.9%, mean body mass 342.14 ± 64.3 g; n = 7), in line with recommendations from Kenward et al. (2001) and later studies (e.g., Geen et al., 2019), and similar to previous studies on the species (e.g., Krupiński et al., 2021; Berger-Geiger et al., 2022). It should be noted that heavier loggers have also been used for this species in the past (e.g., 14–17 g; Krupiński et al., 2021). Devices were programmed to collect daytime GPS positions every hour and provided data on instantaneous flight speed (km/h) and altitude (m above ground). One of the birds – T69555 – died 12 days after fledging, thus providing complete data for the first week of the PFDP only. The body was found under a medium voltage pylon in the surroundings of the nest site and the cause of death was ascertained as head trauma from impact, as per the necropsy carried out at the "CRUMA" rehabilitation center.
2.3
Tracking data processing and parameters
GPS positions were imported into QGIS (v. 3.6.1) and projected to the Universal Transverse Mercator (UTM) coordinate system for all spatial analyses. To characterize and spatio-temporally define the PFDP, we followed Egea-Casas et al. (2023), calculating the following parameters: (1) the onset of the PFDP, (2) the end of the PFDP and (3) the length of the PFDP, as the difference between the previous dates.
The onset of the PFDF was set as the date of the first flight, defined by means of the "recurse" R package (Bracis et al., 2018). This package allows to look for returns to a previously visited area, i.e., revisitations, and computes the number of visits and time spent inside and outside a circular area around a defined central point (i.e., nest site). Accordingly, for each bird, we set the coordinates of its nest as the central point around which we built a 50 m radius circular buffer area (hereafter: "natal area"). Following Egea-Casas et al. (2023), the first flight > 50 m away from the natal area for more than 1 h (corresponding to the sampling interval of our data; Bracis et al., 2018) followed by regular revisits was considered as the onset of the PFDF. The threshold chosen (50 m) represents twice the nominal error of the GPS (e.g., Egea-Casas et al., 2023). Moreover, before tracking birds we tested all devices by putting them outdoors on a spot where they collected locations for two consecutive days. Overall, the average GPS error was of 6.81 ± 6.56 m (n = 555 fix from 7 devices), while the maximum error distance recorded was of 47.12 m, thus supporting the choice of the selected threshold.
As pointed out by Limiñana et al. (2008), the transition between the PFDP and the PMP is indicated by a peak in the variability of the distance covered between consecutive locations, regardless of the differences among individuals in their movement patterns. Thus, following Limiñana et al. (2008) and Egea-Casas et al. (2023), the end of the PFDP was defined as coinciding with the onset of the PMP. To exactly define this moment, both spatial and temporal thresholds were set. Spatially, we calculated the net squared displacement (NSD), as the squared distance between the nest site and subsequent locations (Bunnefeld et al., 2011; Cagnacci et al., 2016; Egea-Casas et al., 2023). Thus, we distinguished the end of the PFDP as an abrupt change in the NSD not followed by any return within an area of 5 km radius from the nest. This radius corresponds to the average linear distance to the nest (or activity center) of breeding adults during the reproductive period (e.g., 3.5–3.7 km in Poland; Krupiński et al., 2021; 4.4–4.9 km in Spain; Berger-Geiger et al., 2022), thus representing parents' territories. From a temporal point of view we took into account birds not returning in that area for at least 7 days, which is a reasonable time to consider individuals fully capable of hunting alone (Cramp and Simmons, 1980). Since adults were not equipped with tracking devices, considering that they could abandon the breeding area and start migration before juveniles (e.g., Arroyo et al., 2002a), both the above mentioned thresholds helped us to disentangle the end of the juveniles' dependency from the parents and/or their territory. The duration of the PFDF (expressed in days) was calculated as the difference between the date of the onset of the pre-migratory phase and that of the first flight.
For the PFDP, we also gathered using the "recurse" R package: (4) the residence time, as time spent in the natal area, (5) the number of revisits to the natal area and (6) the interval time between successive revisits. Total residence time was calculated as the sum of the hours spent within the natal area and divided by 24 to obtain the total amount in days. In addition to that, individual home ranges and core areas were estimated for each bird as the 95% (hereafter "HR") and 50% (hereafter "CA") Utilization Distribution (UD) respectively, using the fixed kernel density estimation (KDE) with reference bandwidth (href) in the R package adehabitatHR (Calenge, 2006). As an alternative metric for literature comparison, we also computed the Minimum Convex Polygon (MCP) encompassing all GPS locations obtained for each bird (Worton, 1989). At this scope, the QGIS tool "Minimum Bounding Geometry" was used, selecting the option "convex hull". Mean and maximum daily distances covered and mean and maximum distance from nest were also computed for this period. To investigate the spatio-temporal progression of the space use by harriers during the PFPD, we also reported similar estimates on a weekly basis (Appendix Table S1).
The same approach used above applies to the transition between the PMP and the start of the autumn migration (Limiñana et al., 2008). In this case, we defined the end of the PMP as coinciding with the onset of the autumn migration, which in turn was defined as an abrupt change in the NSD indicated by a southward movement of > 100 km/day without a return flight towards the North (e.g., Berger-Geiger et al., 2022). For the PMP, we reported: i) mean and maximum daily distances covered (km/d); ii) maximum distance from the nest (km); iii) duration (in days). The function 'as.ltraj' from the 'adehabitatLT' R package (Calenge, 2006) was adopted to distinguish between: i) traveling (i.e., tortuosity index < 1.5 – as a proxy of transit movements between areas); and ii) foraging (tortuosity index > 1.5 – as a proxy for prospecting and foraging movements) tracks' segments (e.g., Roshier et al., 2008). From that, we calculated the time spent (in days) for both movement categories during the entire PMP.
For foraging locations we investigated habitats assessed with the CORINE Land Cover (CLC, 2020), using the QGIS software. Following McKinlay et al. (2024), we simplified the CLC level-3 land nomenclature into six classes: (1) urbanized, (2) agricultural, (3) woodland, (4) grassland, (5) water bodies and (6) other habitats, including natural spaces without or with poor vegetation and/or dunes. The proportion of foraging locations in each habitat category was then computed at individual level. Furthermore, for conservation purposes, we evaluated whether foraging grounds were located inside or outside protected areas, using the World Database on Protected Areas, the most comprehensive global database on terrestrial and marine protected areas (WDPA; https://www.protectedplanet.net/; UNEP-WCMC, IUCN, 2024). The proportion of foraging locations within/outside protected areas was then evaluated, over the entire PMP, at the individual level and across all individuals. Finally, we defined as key staging areas (hereafter "KSA") that zones where birds likely prepared for an energetic challenge (sensu Warnock, 2010), with a concentration of foraging locations whose distance did not overcome 10 km apart and that were used for more than 3 consecutive days (i.e., thus not considering occasional foraging locations). For each individual we counted them, and we calculated the number of days spent in the last visited KSA before autumn migration.
2.4
Statistical analyses
All spatial and statistical analyses were carried out using R software (v4.3.1; R Core Team, 2022). Due to the limited sample size and the non-normal distribution of data non-parametric tests were used. Differences in total residence time, number of revisits, home ranges and movement patterns between successive weeks during the PFDP were tested with the non-parametric Friedman test, while the two-tailed Wilcoxon signed rank test was used to test differences between the first two weeks after fledging, for which more data were available. Differences between sexes were not analysed, given the low sample size and individuals' age class. Data are reported as mean ± standard deviations (SD).
To investigate if the time elapsed since the first flight (in days) affected residence time and time interval between successive revisits at the natal area during the PFDP, we used generalized linear mixed models (GLMMs; Zuur and Ieno, 2016). For this specific analysis these two response variables were treated in two separate models and both modeled with tweedie errors, as it is customary with continuous, 0 + variables. The time elapsed from first flight (continuous variable) was set as fixed predictor. In all models, we included 'individual' as a random effect. The model selection used Akaike's Information Criterion corrected for small sample sizes (AICc). Models were selected if they had ΔAICc ≤ 2, and if their AICc value was lower than that of any simpler alternative (Harrison et al., 2018). Therefore, we obtained either a set of top-ranked models or a single best model for each response variable. Inference about the effect of the predictor was made using the best model: we estimated coefficients and 95% confidence intervals (CIs) of the predictor, assessing whether 95% CIs overlapped "0" to identify informative predictor. GLMMs and model selections were performed using the function glmmTMBglmmTMB (Brooks et al., 2017) for fitting models and dredgeMuMIn (Bartoń, 2020) for model selection. Best models were validated through visual inspection of residuals through the 'DHARMa' package (Hartig, 2022). Plots for the top-ranked models were obtained using package "effects" version 4.2–0 (Fox, 2003; Fox and Weisberg, 2019).
3.
Results
Overall, the first flight of tagged birds occurred in the second half of July (Table 1). After fledging, birds showed high variability both in movement patterns and duration of the different stages (PFDP and PMP) (Fig. 1). All tagged birds, except T69555 who died prematurely (cfr. Methods), completed the PFDP and wandered for about one month across different Italian regions until autumn migration, which started in the second half of September.
Table
1.
Key movement metrics during the post-fleding dependence period (PFDP) of seven juvenile Montagu's Harriers tracked in central Italy in 2024.
ID bird
Sex of the bird
Start tracking
Onset PFDP
End PFDP
Duration PFDP (days)
Abrupt departure (yes/no)
HR (km2)
CA (km2)
MCP (km2)
Mean daily distance (km)
Max daily distance (km)
Mean distance from the nest (km)
Max distance from the nest (km)
No. of revisits
Total residence time (days)
Residence time/PFDP (%)
T69553
F
06/07/2024
11/07/2024
02/08/2024
22
yes
0.326
0.034
3.600
1.936 ± 2.2
7.913
0.105 ± 0.2
1.964
60
15.911
72.323
T69554
F
15/07/2024
16/07/2024
03/08/2024
18
yes
0.229
0.032
0.430
1.417 ± 1.2
5.113
0.148 ± 0.1
0.861
18
7.534
41.853
T69555*
F
20/07/2024
26/07/2024
07/08/2024*
12*
na
0.042
0.005
0.080
0.308 ± 0.4
1.823
0.061 ± 0.1
0.465
7
9.455
78.794*
T69556
F
20/07/2024
26/07/2024
26/08/2024
31
yes
1495.540
111.759
3533.955
17.234 ± 38.8
132.132
4.176 ± 11.8
96.722
14
9.197
29.668
T69558
F
20/07/2024
30/07/2024
27/08/2024
28
no
141.259
20.151
423.905
16.703 ± 20.5
82.844
4.025 ± 4.5
39.568
45
6.294
22.479
T69559
F
20/07/2024
26/07/2024
13/08/2024
18
no
1.045
0.118
11.348
2.842 ± 3.8
14.454
0.204 ± 0.2
4.999
29
5.763
32.017
T69563
M
20/07/2024
23/07/2024
15/08/2024
23
yes
0.147
0.014
2.264
1.146 ± 1.9
6.847
0.075 ± 0.1
2.890
42
18.273
79.449
ID of the bird (ring code), start tracking date, onset, end and duration of the PFDP (expressed in days), indication on abrupt departure from natal site (yes or no), size of home range (HR, 95% kernel density estimate (KDE)), core areas (CA, 50% KDE) and maximum convex polygon (MCP) estimates expressed in km2 are provided. We also provided estimates of mean and maximum daily distances covered, mean and maximum distance from the nest, number of revisits and total residence time at the natal area, and percentage of residence time over the entire PFDP length. Mean values are reported with their mean ± standard deviation. The asterisk indicates the individual that died prematurely 12 days after fledging and for which the estimates refer only to this period. "na" means data not available.
Table
2.
Parameters estimated from top-ranked GLMMs predicting residence time (A) and time since last visit at the natal area (B) during the post-fleding dependence period (PFDP) of six juvenile Montagu's Harriers tracked in central Italy in 2024 in relation to the time elapsed since the first flight (elapsed days).
Model
Response variable
Predictor
β coefficient
95% CI
A
Residence time at nest (n = 212)
Intercept
3.298
2.673; 3.924
σ2 Individual = 0.5561 (best model)
Elapsed days
−0.155
−0.182; −0.127***
B
Time since last visit (n = 205)
Intercept
0.653
−0.026; 1.332
σ2 Individual = 0.5892 (best model)
Elapsed days
0.093
0.064; 0.123***
Variance of random intercepts (σ2), predictors' coefficient estimates (β), and their 95% confidence intervals (CIs). An asterisk marks coefficients whose CIs do not include "0".
Figure
1.
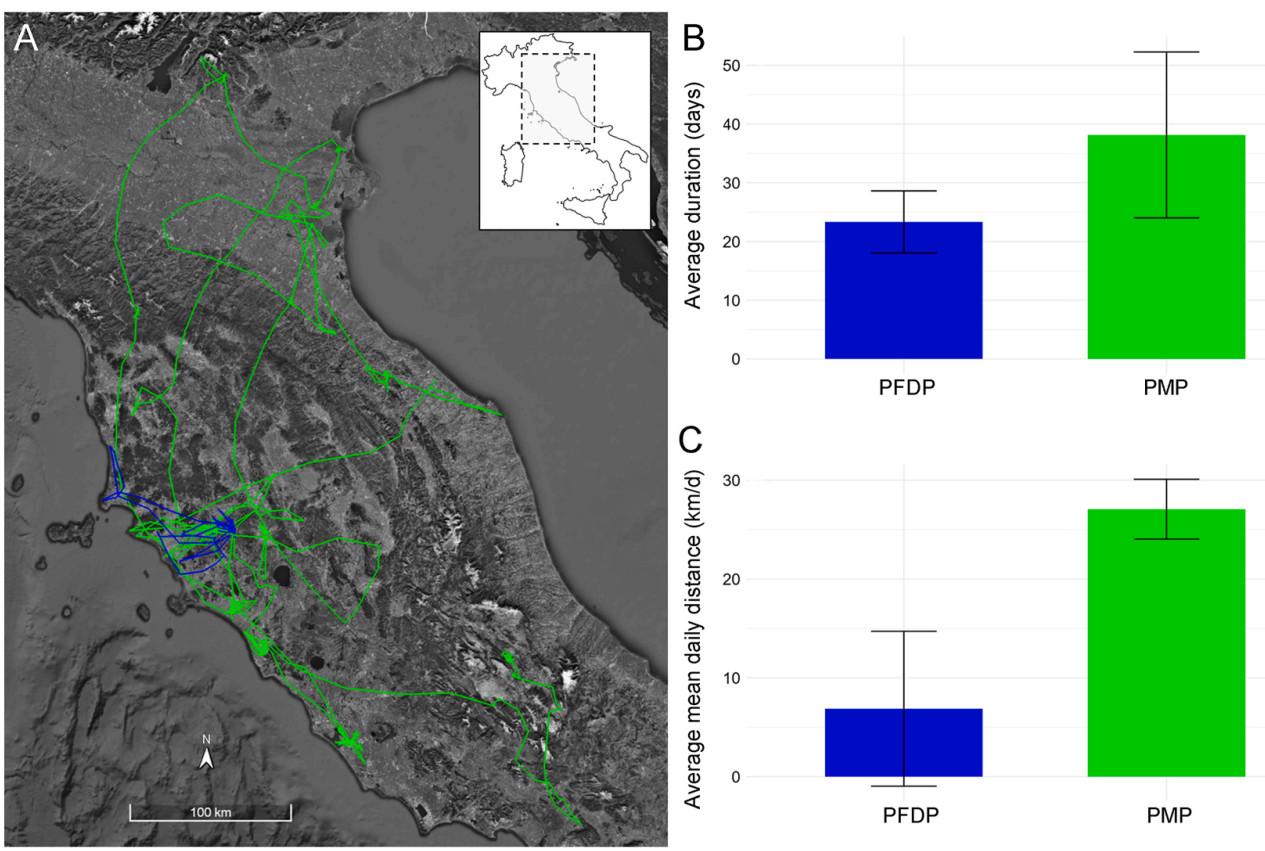
Early life movements and their spatio-temporal metrics of juvenile Montagu's Harriers (Circus pygargus) tracked in central Italy in 2024. (A) Tracks of movements carried out during the PFDP (in blue) and PMP (in green), for all individuals pooled. The inlet shows a map of Italy with a light grey rectangle highlighting the zoomed-in area where the movements occurred. (B) Average duration (in days) and (C) average mean daily distance (in km/day) during both periods, using the same color scheme as in panel A. Bars indicate standard deviations.
The PFDP lasted on average 23.3 ± 5.3 days and varied among individuals (min-max: 18–31 days, n = 6). During this period, birds stayed in the surrounding of the nest (mean distance from the nest: 1.45 ± 2.8 km), with an average daily distance covered of 6.88 ± 11.44 km/day. Among all six birds, the maximum distance from the nest and the maximum daily distance recorded were 96.72 km and 132.13 km, respectively (Table 1). Home range size varied among individuals (Table 1; Fig. 2). On average, home ranges and core areas were 273.09 ± 601.5 km2 and 22.02 ± 44.7 km2, respectively (n = 6). The mean MCP was 662.58 ± 1416.65 km2 (n = 6). Partial estimates for individual T69555 which died during the PFDP are provided in Table 1.
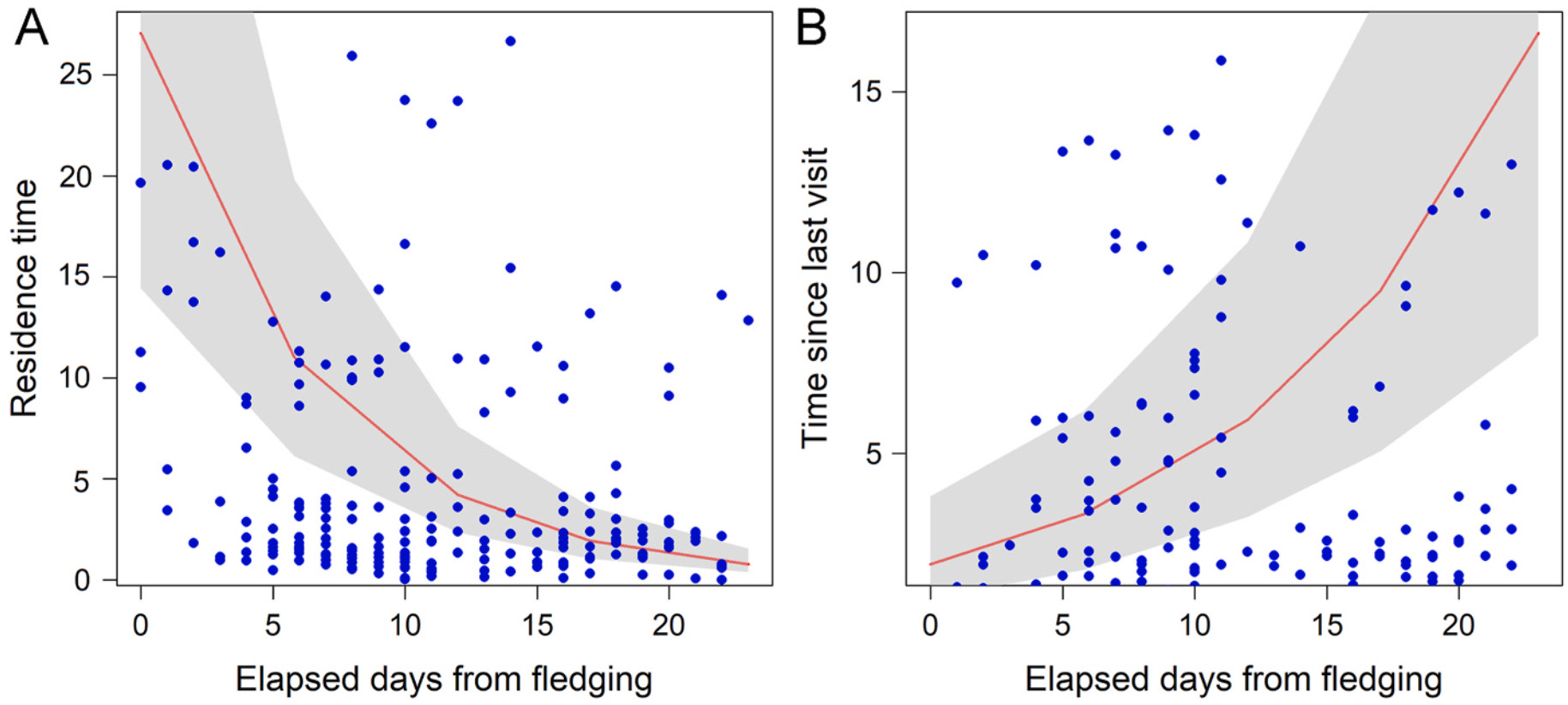
The mean number of revisits at the natal area during the PFDP was 36 ± 19.9, varying between individuals (min-max: 14–68 revisits, Table 1). The mean individual residence time at the nest was 10.17 ± 4.9 days, representing on average 46.23 ± 23.8% of the total PFDP duration. This percentage ranged between 22.5% and 79.44% among individuals. While residence time at the natal area significantly decreased over time (Table 1; Fig. 3A), time interval between revisits significantly increased as the PFDP progressed (Table 1; Fig. 3B).
Figure
3.
Plots of the top-ranked GLMMs (see Table 2) predicting the effect of time elapsed from first flight on residence time at the natal area (A) and the effect of elapsed days from fledging on time since last visits at the natal area (B) during the post-fleding dependence period (PFDP) of six juveniles Montagu's Harriers tracked in central Italy in 2024. Plots for the top-ranked models were obtained using package "effects" version 4.2–0 (Fox, 2003; Fox and Weisberg, 2019).
More specifically across weeks, the Friedman test revealed a significant difference in all parameters (i.e., an increase in home range, core area, cumulative distance, distance from the nest, number of revisits and a decrease in total residence time at the natal area) during the first three weeks of the PFDP (p < 0.001 in all cases; n = 17; Appendix Table S1). Similarly, the Wilcoxon signed rank test confirmed a significant difference in all parameters (same direction as mentioned above) between the first and the second week of the PFDP (p < 0.05 in all cases; n = 13). Four birds out of the six that completed this stage departed abruptly from the natal area (after ca. 18–23 days), without any return. In contrast, the two remaining birds visited closer areas and returned several times to the natal area before starting their PMP (Table 1).
3.2
Variation in PMP parameters
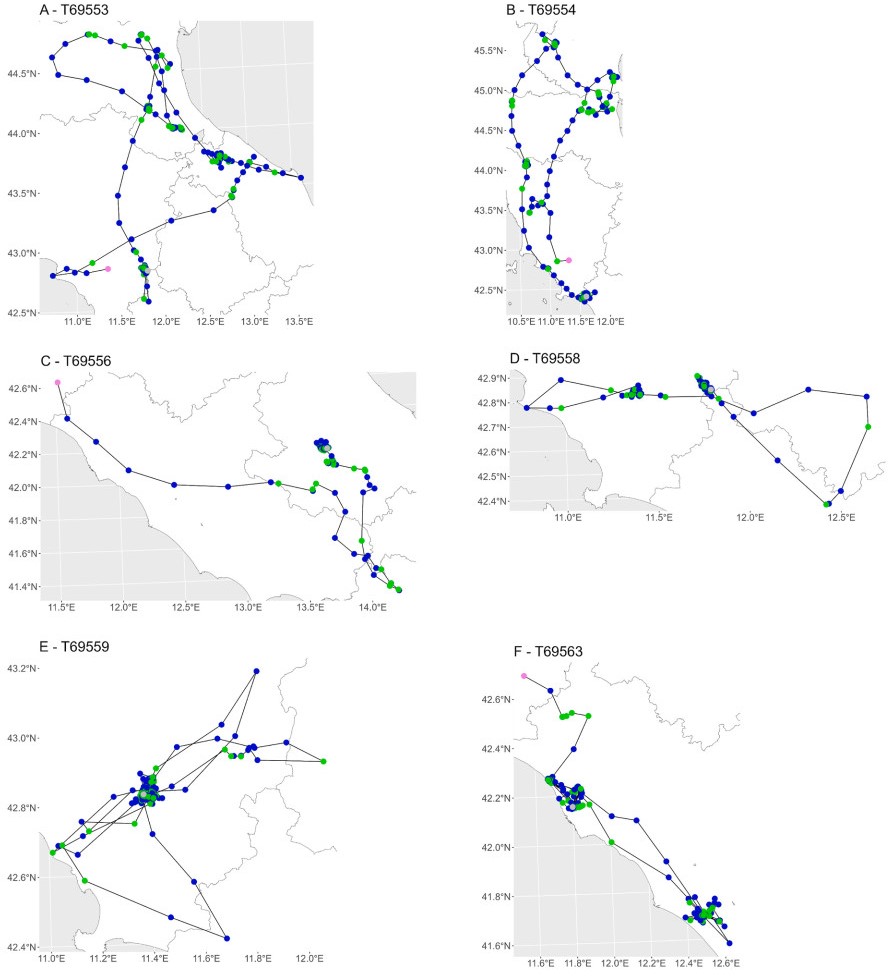
Explored areas and distances from the nest differed among individuals during the PMP. The great majority of juvenile harriers (five out of six) showed marked ranging behaviour, visiting several Italian regions (Fig. 1 and Fig. 4). Maximum distance from the nest was on average 188.6 ± 105.8 km, with a maximum of 320.8 km recorded by individual T69554 (Table 3). However, mean and maximum daily distances covered were 27.06 ± 40.4 km/day and 178.7 ± 56.2 km (n = 6) respectively, were similar among individuals. The foraging locations were sparsely distributed across central-northern Italy (Fig. 4). In only one case, individual T69559 stayed for 46 days in the same area located at ca. 15 km from its natal site, before starting its autumn migration from there (Fig. 4E). Harriers spent on average 60.7% of the PMP time traveling between foraging locations, and 39.3% in foraging movements (Appendix Fig. S1).
Figure
4.
Movements of juvenile Montagu's Harriers during the PMP, tagged in central Italy in 2024. For each individual, foraging locations are reported in green and traveling locations in blue, while first and last position are indicated with violet and grey, respectively.
Table
3.
Key movement metrics during the pre-migratory phase (PMP) of six juvenile Montagu's Harriers tracked in central Italy in 2024.
ID bird
Onset PMP
End PMP
Duration PMP (days)
Mean daily distance (km/day)
Max daily distance (km/day)
Max distance from the nest (km)
T69553
02/08/2024
28/09/2024
57
28.598 ± 53.9
254.274
221.631
T69554
03/08/2024
20/09/2024
48
31.943 ± 44.1
170.447
320.814
T69556
26/08/2024
18/09/2024
23
28.013 ± 51.3
231.293
280.804
T69558
27/08/2024
20/09/2024
24
23.963 ± 38.9
163.041
93.675
T69559
13/08/2024
28/09/2024
46
24.645 ± 31.2
154.854
49.113
T69563
15/08/2024
15/09/2024
31
25.252 ± 23.2
98.421
165.916
ID of the bird (ring code), start and end of the PMP and its duration are provided. Mean daily distances and maximum daily distance covered are reported, together with the maximum reached distance from the nest for each individual. Mean values are reported with their ± standard deviation.
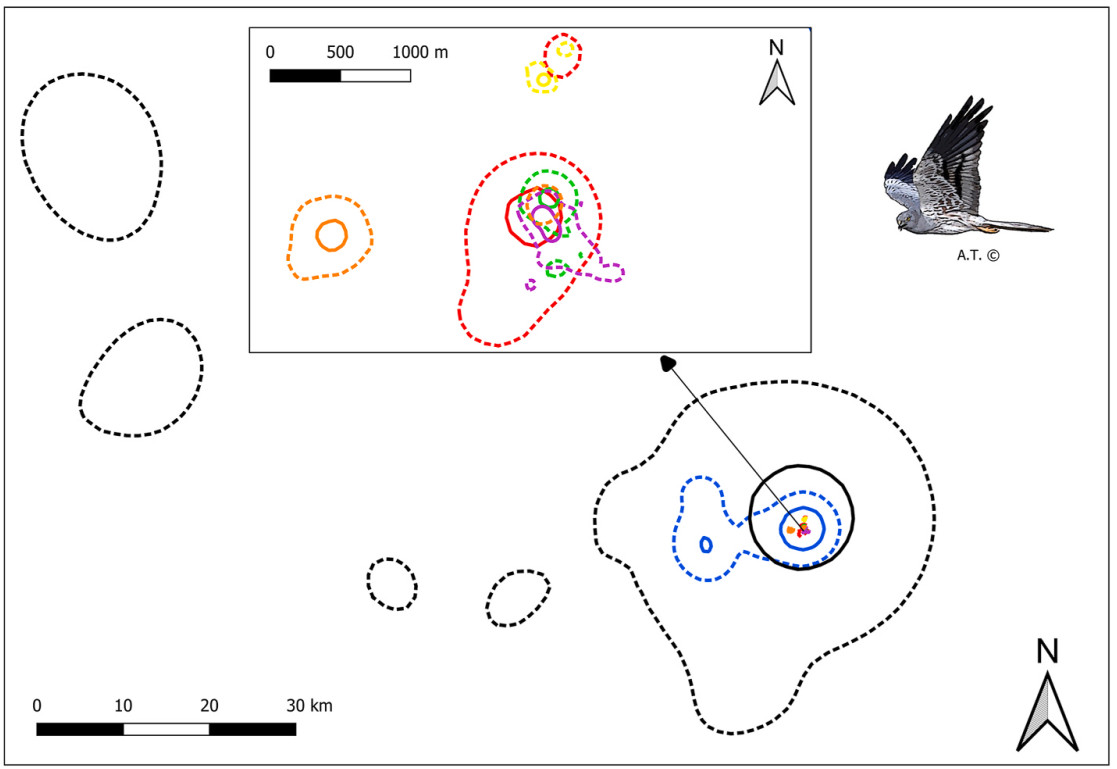
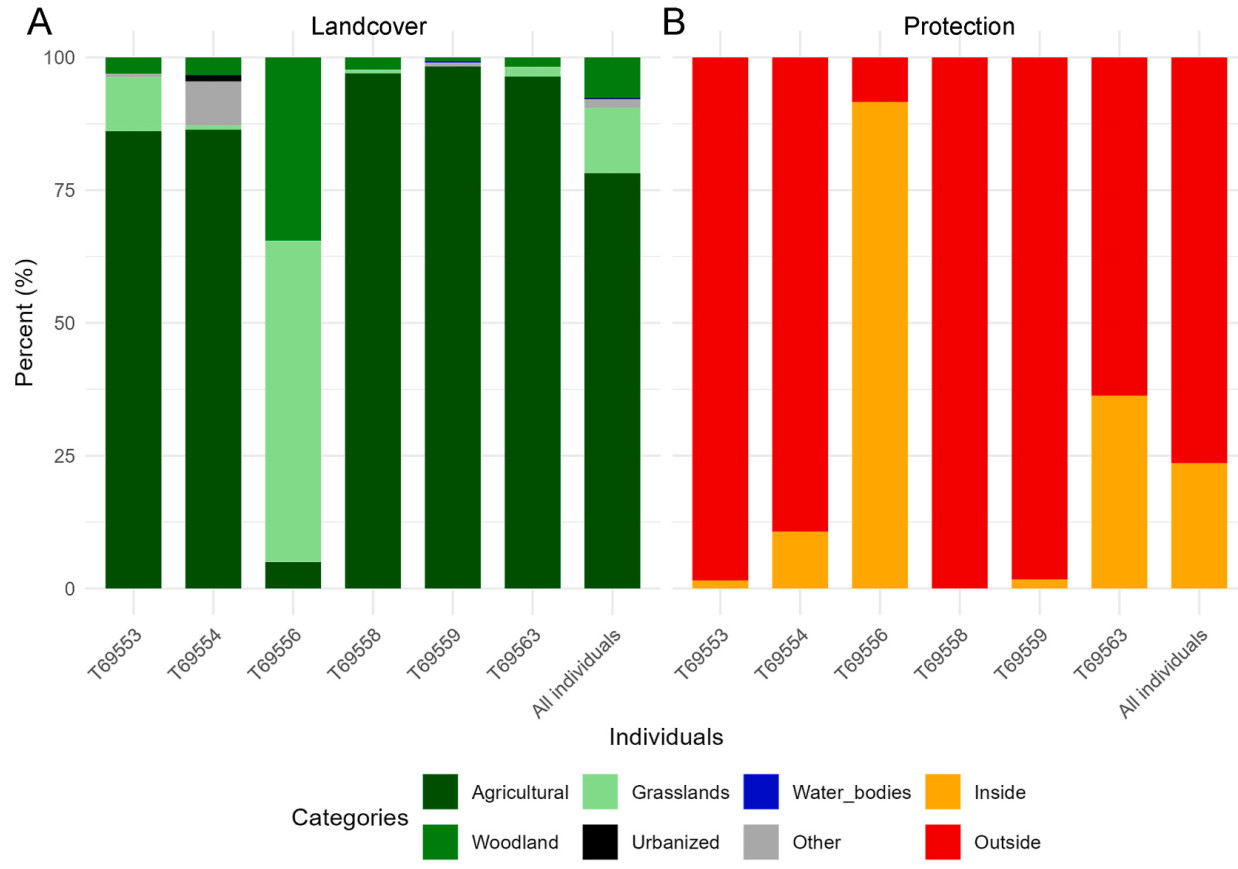
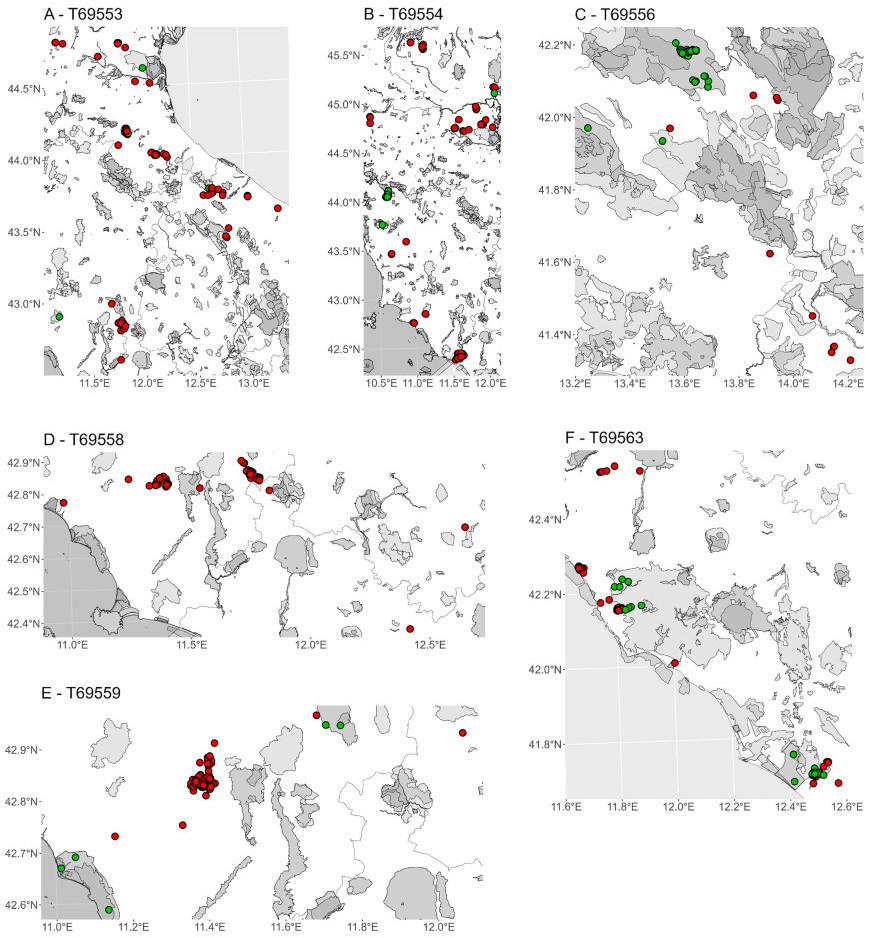
On average, agricultural land was the most frequently used habitat (mean across individuals: 78.2%), followed by grasslands (12.3%), woodlands (7.6%), other habitats (1.6%), and by urbanized (0.2%) and water bodies (0.1%) that were scarcely used (Fig. 5A). Land cover composition of foraging locations was similar among individuals, except for T69556 which it mostly foraged in grassland habitat (Fig. 5A). Foraging locations were mostly located outside protected areas (mean across individuals: 76.4%), with differences between individuals (Fig. 5, Fig. 6). The exception was individual T69556 which settled in the Sirente-Velino Regional Park in the Apennines, where it spent 90% of its foraging time inside the protected area (Fig. 5, Fig. 6C).
Figure
5.
Percentage composition of land cover (A) and protected areas use (B) for foraging locations of six juvenile Montagu's Harriers tracked in central Italy in 2024 during their pre-migratory phase (PMP). Used habitats were assessed using the CORINE Land Cover (CLC, 2018) layer and land nomenclature, simplified into six classes: (1) urbanized, (2) agricultural, (3) woodland, (4) grassland, (5) water bodies and (6) other habitats including natural spaces without or with poor vegetation and/or dunes. Protected areas use was evaluated using the World Database on Protected Areas (WDPA; https://www.protectedplanet.net/; UNEP-WCMC, IUCN, 2024). Each bar represents one individual, with mean percentages for all individuals pooled together reported in the rightmost bars.
Figure
6.
Protected areas utilization during foraging by six juvenile Montagu's Harriers tracked in central Italy in 2024 during the pre-migratory phase (PMP). In red foraging locations outside protected areas and in green those inside protected areas. Protected areas are represented by light grey polygons with transparency. Intersections between foraging locations and protected areas were performed using the World Database on Protected Areas (WDPA; https://www.protectedplanet.net/; UNEP-WCMC, IUCN, 2024).
All harriers but one – T69559 – had more than one KSA during the PMP (Appendix Table S2). All harriers, including those with a greater number of KSAs, stopped wandering and remained in a single KSA for on average 21.8 ± 14 days (range: 3–46 days), before initiating autumn migration (Appendix Table S2). On average, the PMP lasted 38 ± 14 days (n = 6; Table 3), with all birds departing for autumn migration in the second half of September (range: 15/09–28/09; Appendix Table S2).
4.
Discussion
Our study assessed the space use of juvenile Montagu's Harriers in Italy during the post-fledging dependence period and pre-migratory phase. The PFDP is considered as one of the most critical stages in the life cycle of raptors (e.g., López-López et al., 2014; Hemery et al., 2023) as flight and foraging skills are developed. However the way in which such development occurs as well as the decision to emigrate from the natal territory may vary among individuals (Delgado et al., 2009). In line with that, we found substantial individual spatio-temporal variation in movement patterns across the different phases, from fledging till the onset of migration.
As expected, after fledging a progressive decrease in the residence time in the natal area and, conversely, an expansion of the time elapsed between subsequent revisits to the same area were recorded, suggesting that juveniles gradually improved their flight skills and started exploring more widely the surroundings. This was also evident from the comparisons of the various parameters on a weekly basis (Appendix Table S1). The occurrence and frequency of first explorations generally increased with time, even though juveniles continued to attend the natal area, as described for other raptor species (e.g., Black Kite Milvus migrans, Bustamante and Hiraldo, 1989; Red Kite Milvusmilvus, Bustamante, 1993; Osprey Pandion haliaetus, Bustamante, 1995; Monti et al., 2012; Eagle Owl Bubobubo, Delgado et al., 2009; Bearded Vulture Gypaetus barbatus, López-López et al., 2014; Golden Eagle Aquila chrysaetos, Hemery et al., 2023). However, although this general pattern occurred for all individuals, they differedin the use of the natal area, both in space and time. The PFDP movement metrics varied consistently among individuals, with some of them showing larger home ranges and distances covered than others. Arroyo et al. (2002a) showed that the duration of the PFDP in Montagu's Harriers may be adjusted according to fledging date (shorter for birds that fledge later in the season) and/or food condition in the natal area (longer in poorer food conditions for equivalent fledging date). Our sample size was too limited to statistically support these hypotheses, though such variability was recorded even for individuals with similar fledging date (cfr. Methods). For example, the fleding dates of the two siblings T6558 and T6559, with almost the same weight at ringing (< 5 g difference), differed by three days, but the length of their PFDP differed by 10 days. Parental aggression toward fledglings or reduction in food provisioning are known to be other factors potentially affecting PFDP duration (Alonso et al., 1987; Bustamante, 1993), though they were not investigated in this study.
Overall, the average length of the PFDP across all six GPS-tracked harriers (ca. 23 days) showed a figure comparable to that reported for field studies in Spain (25 days; Arroyo et al., 2002a), Poland and Italy (e.g., 24 days; Kitowski, 2005; Pandolfi, 1996). In this respect, our study provides further support to previous findings gathered without the use of current technological devices. Interestingly, we recorded that the two individuals with longer PFDP (28 and 31 days, respectively) also had larger home ranges, higher mean daily distances covered and mean and maximum distance from the nest. They also spent less time (residence time < 30%) in the natal area in proportion to the total PFDP length, though no clear patterns were observed in the number of revisits at the nest. This could suggest that individuals that took longer to become independent, showed larger movements during the PFDP, could acquire a better knowledge of the natal area and its surroundings compared to individuals with shorter PFDP that moved relatively less during the PFDP and showed higher rates of residence time in the natal area (up to 79%). In the future, following more GPS-tracked juveniles and adults from the same area will help to extend the investigation on the PFDP and to verify these preliminary findings/hypotheses.
The way in which the PMP was initiated also varied considerably at the individual level. Both long-distance exploratory movements and local displacements to nearby foraging areas were recorded. Four out of six individuals abruptly abandoned the natal area, starting traveling across several Italian regions in different directions, up to > 320 km from the nest, and switching between different foraging locations. In contrast, the two remaining birds mostly attended an area in the vicinity of the natal site (ca. 15 km). This variability in movement patterns is even more interesting if we consider that all birds came from and were raised in the same area, within a closely spaced cluster of nests. One could speculate that, trophic resources' accessibility and availability being the same for all pairs, inter-individual variability in juveniles' movement patterns could be dependent on intrinsic factors and/or differences in parental investment/feeding rate, successively affecting chicks' growth and development, and ultimately the duration of the PMP. Nevertheless, beyond this variability some common patterns have been recorded. For example all the harriers, including those that had explored various areas in different directions during the PMP, successively stabilized in a single KSA, where they remained for more than 20 days on average before departing for autumn migration. It is therefore possible that individuals have chosen to maximize their foraging efforts in a single area, rather than continuing traveling around. Another advantage of this strategy might be that the birds are safer in an area they know well, thus increasing their likelihood of survival.
As recently pointed out by Barbraud et al. (2024) for another long-distance migratory land bird that mostly feeds on the ground on invertebrates, the White Stork (Ciconiaciconia), quantifying spatial and temporal habitat use is fundamental for addressing ecological issues related to habitat and species conservation strategies. In our case all harriers, except the one who settled in the Apennines and foraged on grassland habitats, mostly exploited agricultural lands, both inland and close to the coast, in some cases adjacent to peri-urban areas. Our analyses also showed that the great majority of these foraging sites fall outside protected areas. This means that these areas, so important for juvenile harriers preparing for their migration, do not benefit from any protection. This translates into a high potential risk for the species and conflicts with human activities, namely those linked to agriculture, as questioned for the Eurasian Stone-curlew (Burhinus oedicnemus) (Gaget et al., 2019). These areas not only offer shelter from human disturbance but also safeguard essential habitats from degradation due to agricultural or industrial expansion, while maintaining ecological connectivity for migratory species and supporting the ecological processes necessary for long-distance migration (e.g., Cooke et al., 2024). In another raptor species, the Osprey, Montillo et al. (2022) demonstrated how that the availability of suitable habitats within protected areas can significantly improve survival rates for juvenile birds. The absence of harrier foraging sites within these protected areas further underscores the urgency of expanding conservation efforts to encompass a broader range of habitats, including those utilized by migratory species in their preparatory phases (e.g., Gameiro et al., 2024).
On the other hand, the observed inter-individual differences in movement patterns, while exposing juvenile birds to greater risks and vulnerabilities, also indicate a level of flexibility and potential adaptability to changing environmental conditions. This flexibility may enhance the species' ability to navigate novel or changing habitats, suggesting potential adaptive strategies that could help mitigate the impacts of habitat loss. Such adaptability may prove to be an essential factor in the species' resilience to shifting ecological circumstances, especially in the context of climate change and increasing human influence on the landscape.
Finally, with regards to the onset of autumn migration, ranges in departure dates spanned less than two weeks across all individuals, starting from the second half of September. Historical ringing recoveries and data from migration surveys at watchpoints/migratory bottlenecks (García and Arroyo, 1998) evidenced that passage frequency is higher in the autumn than the spring migration in the Gibraltar Strait, but the opposite occurs in the central Mediterranean (e.g., Sicily Channel). In Italy, Panuccio et al. (2005) found that few Montagu's Harriers cross the central Mediterranean during post-breeding movements. The cited survey, carried out both at Panarea island and at the Straits of Messina bottleneck, was carried out until the 10th of September, thus possibly missing juveniles/immature individuals departing later in the season, as in the case of our tagged birds. However, being a facultative soaring migrant, the species can attain long flight bouts over the open sea, resulting in broad front migration (Schlaich and Klaassen, 2021), de facto not concentrating at migration bottlenecks and watchpoints (e.g., De Pascalis et al., 2020), as also evidenced by Meyer et al. (2003) for the southern Mediterranean coasts of Spain. This means that a large part of the population is actually missing from being counted, at least in autumn (e.g., Schlaich and Klaassen, 2021). Studies using tracking technology in Montagu's Harriers showed that the departure date of autumn migration occurred from the second half of August, as recorded in Spain (Limiñana et al., 2012a, Limiñana et al., 2012b) and in the Netherlands (Bengtsson and Jonzén, 2001; Trierweiler et al., 2007). However, these values nearly exclusively refer to adult individuals that are known to leave earlier from breeding grounds than juveniles (Arroyo et al., 2002a). Further tracked-individuals from other regions could help depicting a better timing of autumn migration departures date and routes followed for juveniles Montagu's Harriers of the Italian population.
5.
Conclusions
By means of GPS-tracking technology, this work sheds light on movement ecology during the early life stages of the vulnerable Montagu's Harrier in Italy. From a conservation point of view, this novel study provides key elements for identifying gaps in the protection measures currently adopted for this species at national level. Actually, the protected areas network does not fulfill the species' requirements as the majority of key areas used by juvenile harriers during the PMP are located outside protected areas, raising concerns for its long-term protection. Since the Montagu's Harrier is closely linked to agricultural environments, habitat protection becomes of fundamental importance for its long-term preservation (e.g., Gameiro et al., 2024). However, agricultural landscapes often represent environments that are "off-the-radar" of conservation or protected area designation, either because they are used for productive/economic purposes or because they are considered less important than other habitats. The current standard approach of designating protected areas to conserve species seems to be inefficient to ensure the protection of wide-ranging species (e.g., Montillo et al., 2022), as in this case for the Montagu's Harrier. Furthermore, the high inter-individual variability in movement patterns and wide-ranging exploratory behaviour further complicates the implementation of fully satisfactory management measures. These environments deserve greater attention, tailor-made management measures and a more sustainable use of their resources, aiming at safeguarding farmland bird populations and the rich biodiversity that populate them.
All the activities of capturing and handling birds were carried out under formal authorization issued by the Tuscany Regional Administration (N: 10543, 16/052024), after the positive opinion expressed by the Italian Institute for Environmental Protection and Research (ISPRA) (Rif. Int. 18502/2024), in accordance with Law157/1992 [Art.4 (1) and Art 7 (5)], which regulates research on wild bird and mammal species.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We want to thank the Veterinarian Renato Ceccherelli and its staff at the Centro Recupero Uccelli Marini e Acquatici (CRUMA-LIPU), for the important assistance and support provided during ringing activities at the nest. All volunteers from the Gruppo Ornitologico Maremmano (GOM) who took part in field surveys and helped in detecting Montagu's Harriers activity areas are also deeply acknowledged. FM was funded by EU – Next Generation EU Mission 4 "Education and Research" – Component 2: "From research to business" – Investment 3.1: "Fund for the realization of an integrated system of research and innovation infrastructures" – Project IR0000032 – ITINERIS – Italian Integrated Environmental Research Infrastructures System – CUP B53C22002150006. The author acknowledges the Research Infrastructures participating in the ITINERIS project with their Italian nodes: ACTRIS, ANAEE, ATLaS, CeTRA, DANUBIUS, DISSCO, e-LTER, ECORD, EMPHASIS, EMSO, EUFAR, Euro-Argo, EuroFleets, Geoscience, IBISBA, ICOS, JERICO, LIFEWATCH, LNS, N/R Laura Bassi, SIOS, SMINO. The authors are grateful to A.E. Schlaich and other three anonymous reviewers for their valuable suggestions on a previous version of the manuscript.
Allen, A.M., Singh, N.J., 2016. Linking movement ecology with wildlife management and conservation. Front. Ecol. Evol. 3, 155.
Alonso, J.C., Gonzalez, L.M., Heredia, B., Gonzalez, J.L., 1987. Parental care and the transition to independence of Spanish imperial eagles Aquila heliaca in doñana national Park, southwest Spain. Ibis 129, 212-224.
Amar, A., Arroyo, B.E., Bretagnolle, V., 2000. Post-fledging dependency and dispersal in released and wild Montagu's harriers Circus pygargus. Ibis 142, 21-28.
Arroyo, B.E., Bretagnolle, V., 2000. Evaluating the long-term effectiveness of conservation practices in Montagu's Harrier Circus pygargus. In: Chancellor, R.D., Meyburg, B.-U. (Eds.), Raptors at Risk. Pica Press, Bodmin, Cornwall, pp. 403-408.
Arroyo, B.E., De Cornulier, T.H., Bretagnolle, V., 2002a. Parental investment and parent-offspring conflicts during the postfledging period in Montagu's harriers. Anim. Behav. 63, 235-244.
Arroyo, B., García, J.T., Bretagnolle, V., 2002b. Conservation of the Montagu's harrier (Circus pygargus) in agricultural areas. Anim. Conserv. 5, 283-290.. .
Arroyo, B., Fernández-Tizón, M., Puche, S., 2024. Comportamiento espacial de la población española de aguilucho cenizo. Monografía n.°10 del programa Migra. SEO/BirdLife. Madrid. .
Barbraud, C., Kato, A., Trehou, G., Dugué, H., Musseau, R., 2024. Factors affecting time activity budgets of breeding White Storks Ciconia ciconia assessed using bio-logging. ARDEOLA 71, 321-335.
Bartoń, K., 2020. MuMIn: multi-model inference. R package version.
Bengtsson, D., Jonzén, N., 2001. Autumn migration of Montagu's harrier Circus pygargus and marsh harrier C. aeruginosus at the southern tip of öland. Ornis Svec. 11, 1-6.. .
Berger-Geiger, B., Heine, G., Kumaraswamy, A., Galizia, C.G., 2022. Changing places: spatial ecology and social interactions of female and male Montagu's Harrier (Circus pygargus) in the Spanish Extremadura. J. Ornithol. 163, 165-179.. .
Berthold, P., 1996. Control of Bird Migration. Chapman and Hall, London, United Kingdom.
Bildstein, K.L., 2006. Migrating Raptors of the World: Their Ecology and Conservation. Cornell University Press, Ithaca, NY, U.S.A.
Bracis, C., Bildstein, K.L., Mueller, T., 2018. Revisitation analysis uncovers spatio-temporal patterns in animal movement data. Ecography 41, 1801-1811.
Brichetti, P., Fracasso, G., 2018. The Birds of Italy: Anatidae-Alcidae; Edizioni Belvedere, Milano, Italy.
Brooks, M.E., Kristensen, K., van Benthem, K.J., Magnusson, A., Berg, C.W., Nielsen, A., et al., 2017. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J 9, 378-400.
Bunnefeld, N., Börger, L., van Moorter, B., Rolandsen, C.M., Dettki, H., Solberg, E.J., et al., 2011. A model‐driven approach to quantify migration patterns: individual, regional and yearly differences. J. Anim. Ecol. 80, 466-476.
Bustamante, J., Hiraldo, F., 1989. Postfledging dependence period and maturation of flight skills in the black kite Milvus migrans. Bird Study 36, 199-204.
Bustamante, J., 1993. Post-fledging dependence period and development of flight and hunting behaviour in the red kite Milvus milvus. Bird Study 40, 181-188.
Bustamante, J., 1995. The duration of the postfledging dependence period of ospreys Pandion haliaetus at Loch Garten, Scotland. Bird Study 42, 31-36.
Butet, A., Leroux, A.B., 2001. Effects of agriculture development on vole dynamics and conservation of Montagu's harrier in western French wetlands. Biol. Conserv. 100, 289-295.
Calenge, C., 2006. The package adehabitat for the R software: a tool for the analysis of space and habitat use by animals. Ecol. Model. 197, 516-519.
Cagnacci, F., Focardi, S., Ghisla, A., Van Moorter, B., Merrill, E.H., Gurarie, E., et al., 2016. How many routes lead to migration? Comparison of methods to assess and characterize migratory movements. J. Anim. Ecol. 85, 54-68.
Cavalcante, T., Tuyama, C.A., Mourthe, I., 2019. Insights into the development of a juvenile harpy eagle's hunting skills. Acta Amazonica 49, 114-117.
Cramp, S., Simmons, K.E.L., 1980. Handbook of the Birds of Europe, the Middle East and North Africa. The Birds of the Western Palearctic, Vol. 2. Oxford University Press, Oxford.
CLC (Copernicus Land Monitoring Service), 2020. CORINE Land Cover 2018 (vector/raster 100 m), Europe, 6-yearly. European Environment Agency, Version V2020_20u1. .
Cooke, S.J., Piczak, M.L., Singh, N.J., Åkesson, S., Ford, A.T., Chowdhury, S., et al., 2024. Animal migration in the Anthropocene: threats and mitigation options. Biol. Rev. 99, 1242-1260.
Delgado, M.M., Penteriani, V., Nams, V.O., 2009. How fledglings explore surroundings from fledging to dispersal. A case study with eagle owls (Bubo bubo). Ardea 97, 7-15.
De Pascalis, F., Panuccio, M., Bacaro, G., Monti, F., 2020. Shift in proximate causes of mortality for six large migratory raptors over a century. Biol. Conserv. 251, 108793.
Egea-Casas, O., López-López, P., Álvarez, E., Cortone, G., Galán, M., Iglesias-Lebrija, J.J., et al., 2023. Similar post-fledging behavior observed in wild and reintroduced juvenile Bonelli's Eagle in two Mediterranean islands. Ornithol. Appl. 125, duad037.
Fox, J., 2003. Effect displays in R for generalised linear models. J. Stat. Software 8, 1-27. .
Fox, J., Weisberg, S., 2019. An R Companion to Applied Regression, third ed. Sage, Thousand Oaks, CA.
Gaget, E., Fay, R., Augiron, S., Villers, A., Bretagnolle, V., 2019. Long‐term decline despite conservation efforts questions Eurasian Stone‐curlew population viability in intensive farmlands. Ibis 161, 359-371.
Gameiro, J., Marques, A.T., Venâncio, L., Valerio, F., Pacheco, C., Guedes, A., et al., 2024. Evidence of a twofold ecological trap driven by agricultural change causing a priority farmland bird population crash. Conserv. Sci. Pract. 6, e13168.
Ganesh, T., Kannan, A., Prashanth, M.B., Samrat, A., 2024. Stopover by migrant Montagu's Harriers in the Thar Desert is determined by vegetation greenness and grasshopper abundance but not locust outbreaks. J. Arid Environ. 225, 105253.
García, J.T., Arroyo, B.E., 1998. Migratory movements of western European Montagu's Harrier Circus pygargus: a review. Bird Study 45, 188-194.
Garrido, J.R., Numa, C., Barrios, V., Qninba, A., Riad, A., Haitham, O., et al., 2021. The Conservation Status and Distribution of the Breeding Birds of Prey of North Africa. IUCN, Gland, Switzerland. xvi + 102pp.
Geen, G.R., Robinson, R.A., Baillie, S.R., 2019. Effects of tracking devices on individual birds–a review of the evidence. J. Avian Biol. 50, e01823. .
Giovacchini, P., 2003. L'avifauna del Sito di Importanza Comunitaria "Monte Labbro e Alta Valle dell'Albegna" (Grosseto, Toscana meridionale). Atti Soc. Tosc. Sci. Nat. Res. B 110, 71-82.
Harel, R., Horvitz, N., Nathan, R., 2016. Adult vultures outperform juveniles in challenging thermal soaring conditions. Sci. Rep. 6, 27865.
Harrison, X.A., Donaldson, L., Correa-Cano, M.E., Evans, J., Fisher, D.N., Goodwin, C.E., et al., 2018. A brief introduction to mixed effects modelling and multi-model inference in ecology. PeerJ 6, e4794.
Hartig, F., 2022. DHARMa: residual diagnostics for hierarchical (multilevel/mixed) regression models. . (Accessed 14 October 2024).
Hemery, A., Mugnier‐Lavorel, L., Itty, C., Duriez, O., Besnard, A., 2023. Timing of departure from natal areas by golden eagles is not constrained by acquisition of flight skills. J. Avian Biol. 2023, e03111.
Jahn, A.E., Bejarano, V., Cueto, V.R., Di Giacomo, A.S., Fontana, C.S., 2017. Movement ecology research to advance conservation of South America's grassland migratory birds. Perspect. Ecol. Conserv. 15, 209-215.
Janowski, S., Pürckhauer, C., Krüger, R., Tietze, D.T., Wink, M., 2018. Recruitment rates, natal and breeding dispersal of Montagu's harriers (Circus pygargus) by means of microsatellite analysis. Open Ornithol. J. 11, 39-55.
Kenward, R.E., Pfeffer, R.H., Al-Bowardi, M.A., Fox, N.C., Riddle, K.E., Bragin, E.A., et al., 2001. Setting harness sizes and other marking techniques for a falcon with strong sexual dimorphism. J. Field Ornithol. 72, 244-257.
Kitowski, I., 2005. Play behaviour and active training of Montagu's harrier (Circus pygargus) offspring in the post-fledging period. J. Ethol. 23, 3-8.
Kitowski, I., 2009. Social learning of hunting skills in juvenile marsh harriers Circus aeruginosus. J. Ethol. 27, 327-332.
Klaassen, R.H., Hake, M., Strandberg, R., Koks, B.J., Trierweiler, C., Exo, K.M., et al., 2014. When and where does mortality occur in migratory birds? Direct evidence from long‐term satellite tracking of raptors. J. Anim. Ecol. 83, 176-184.
Klaassen, R.H.G., Schlaich, A.E., Both, C., Bouten, W., Koks, B., 2019. Individual variation in home range size reflects different space use strategies in a central place foraging raptor bird. In: Schlaich, A.E. (Ed.), Migrants in Double Jeopardy. Doctoral Thesis. University of Groningen, The Netherlands.
Krupiński, D., Kotowska, D., Recio, M.R., Żmihorski, M., Obłoza, P., Mirski, P., 2021. Ranging behaviour and habitat use in Montagu's Harrier Circus pygargus in extensive farmland of Eastern Poland. J. Ornithol. 162, 325-337.
Limiñana, R., Soutullo, A., Urios, V., Surroca, M., 2006. Vegetation height selection in Montagu's Harriers Circus pygargus breeding in a natural habitat. Ardea 94, 280-284.
Limiñana, R., Soutullo, A., Urios, V., 2007. Autumn migration of Montagu's harriers Circus pygargus tracked by satellite telemetry. J. Ornithol. 148, 517-523.
Limiñana, R., Soutullo, A., López-López, P., Urios, V., 2008. Pre-migratory movements of adult Montagu's Harriers Circus pygargus. Ardea 96, 81-90.
Limiñana, R., García, J.T., González, J.M., Guerrero, Á., Lavedán, J., Moreno, J.D., et al., 2012a. Philopatry and natal dispersal of Montagu's harriers (Circus pygargus) breeding in Spain: a review of existing data. Eur. J. Wildl. Res. 58, 549-555.
Limiñana, R., Soutullo, A., Urios, V., Reig-Ferrer, A., 2012b. Migration and wintering areas of adult Montagu's Harriers (Circus pygargus) breeding in Spain. J. Ornithol. 153, 85-93.
López-López, P., Gil, J.A., Alcántara, M., 2014. Post-fledging dependence period and onset of natal dispersal in bearded vultures (Gypaetus barbatus): new insights from GPS satellite telemetry. J. Raptor Res. 48, 173-181.
Meyer, K.S., Spaar, R., Bruderer, B., 2003. Sea crossing behaviour of falcons and harriers at the southern Mediterranean coast of Spain. Avian Sci. 3, 153-162.
McClure, C.J., Westrip, J.R., Johnson, J.A., Schulwitz, S.E., Virani, M.Z., et al., 2018. State of the world's raptors: distributions, threats, and conservation recommendations. Biol. Conserv. 227, 390-402.
McKinlay, S.E., Morganti, M., Mazzoleni, A., Labate, A., Sorrenti, M., Rubolini, D., 2024. Non-breeding ranging behaviour, habitat use, and pre-breeding migratory movements of Fieldfares (Turdus pilaris) wintering in southern Europe. J. Ornithol. 165, 337-346.
Millon, A., Arroyo, B.E., Bretagnolle, V., 2008. Variable but predictable prey availability affects predator breeding success: natural versus experimental evidence. J. Zool. 275, 349-358.
Monti, F., Sforzi, A., Dominici, J.M., 2012. Post-fledging dependence period of ospreys Pandion haliaetus released in central Italy: home ranges, space use and aggregation. ARDEOLA 59, 17-30.
Monti, F., Grémillet, D., Sforzi, A., Dominici, J.M., Bagur, R.T., Navarro, A.M., et al., 2018. Migration distance affects stopover use but not travel speed: contrasting patterns between long‐and short‐distance migrating ospreys. J. Avian Biol. 49, e01839.
Montillo, F., Sforzi, A., Sammuri, G., Duriez, O., Monti, F., 2022. Effectiveness of protected areas for osprey survival at intercontinental scale. Biodivers. Conserv. 31, 1379-1405.
Newton, I., 2004. Population limitation in migrants. Ibis 146, 197-226.
Newton, I., 2010. Bird Migration. Collins, London.
Newton, I., 2011. Migration within the annual cycle: species, sex and age differences. J. Ornithol. 152, 169-185.
Oltra, J., García, J., Carbonell, I., Jambas, J., Álvarez, E., Iglesias‐Lebrija, J.J., et al., 2024. Early life movements and mortality of Egyptian vultures: implications for transcontinental conservation. Ecol. Evol. 14, e70291.
Pandolfi, M., Rossana, P., D'Astore, P., 1990. Analysis of breeding behaviour in Montagu's Harrier Circus pygargus in a site of CentraI Italy. Avocetta 14, 97-102.
Pandolfi, M., 1996. Play activity in young Montagu's Harriers (Circus pygargus). Auk 113, 935-938.
Pandolfi, M., Tanferna, A., 2009. Long-term change in population size and reproductive parameters of Montagu's harriers (Circus pygargus) in Italy. J. Raptor Res. 43, 155-159.
Panuccio, M., Agostini, N., Mellone, U., 2005. Autumn migration strategies of honey buzzards, black kites, marsh and Montagu's harriers over land and over water in the central Mediterranean. Avocetta 29, 27-32.
R Core Team, 2022. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. .
Ravasini, M., 2022. Albanella Minore Cyrcus pygargus. In: Lardelli, R., Bogliani, G., Brichetti, P., Caprio, E., Celada, C., Conca, G., et al. (Eds.), Atlante Degli Uccelli Nidificanti in Italia. Edizioni Belvedere, Latina, pp. 286-287.
Rondinini, C., Battistoni, A., Teofili, C., 2022. Lista Rossa IUCN dei vertebrati italiani 2022. Comitato Italiano IUCN e Ministero dell'Ambiente e della Sicurezza Energetica, Roma, Italy.
Roshier, D.A., Doerr, V.A.J., Doerr, E.D., 2008. Animal movement in dynamic landscapes: interaction between behavioural strategies and resource distributions. Oecologia 156, 465-477.
Ruaux, G., Lumineau, S., de Margerie, E., 2020. The development of flight behaviours in birds. Proc. R. Soc. B 287, 20200668.
Schlaich, A., 2019. Migrants in Double Jeopardy: Ecology of Montagu’s Harriers on Breeding and Wintering Grounds. University of Groningen, NL. Doctoral thesis.
Schlaich, A.E., Klaassen, R.H., 2021. Montagu’s harrier Circus pygargus. In: Panuccio, M., Mellone, U., Agostini, N. (Eds.), Migration Strategies of Birds of Prey in Western Palearctic. CRC Press, pp. 152–159.
Schlaich, A.E., Bretagnolle, V., Both, C., Koks, B.J., Klaassen, R.H., 2023. On the wintering ecology of Montagu's Harriers in West Africa: itinerancy in relation to varying annual environmental conditions. Ardea 111, 321-342.
Sergio, F., Tanferna, A., De Stephanis, R., Jiménez, L.L., Blas, J., Tavecchia, G., et al., 2014. Individual improvements and selective mortality shape lifelong migratory performance. Nature 515, 410-413.
Serratosa, J., Oppel, S., Rotics, S., Santangeli, A., Butchart, S.H., Cano-Alonso, L.S., et al., 2024. Tracking data highlight the importance of human-induced mortality for large migratory birds at a flyway scale. Biol. Conserv. 293, 110525.
Strandberg, R., Klaassen, R.H., Hake, M., Olofsson, P., Thorup, K., Alerstam, T., 2008. Complex timing of Marsh Harrier Circus aeruginosus migration due to pre-and post-migratory movements. Ardea 96, 159-171.
Todd, L.D., Poulin, R.G., Brigham, R.M., Bayne, E.M., Wellicome, T.I., 2007. Pre-migratory movements by juvenile Burrowing Owls in a patchy landscape. Avian Conserv. Ecol. 2, 4.
Trierweiler, C., 2010. Travels to Feed and Food to Breed: the Annual Cycle of a Migratory Raptor, Montagu’s Harrier, in a Modern World. Doctoral Thesis. University of Groningen. University of Groningen, the Netherlands.
Trierweiler, C., Koks, B.J., Drent, R.H., Exo, K.M., Komdeur, J., Dijkstra, C., et al., 2007. Satellite tracking of two Montagu's Harriers (Circus pygargus): dual pathways during autumn migration. J. Ornithol. 148, 513-516.
Trierweiler, C., Klaassen, R.H., Drent, R.H., Exo, K.M., Komdeur, J., Bairlein, F., et al., 2014. Migratory connectivity and population-specific migration routes in a long-distance migratory bird. Proc. R. Soc. B 281, 20132897.
UNEP-WCMC, IUCN, 2024. Protected planet: the world database on protected areas (WDPA) and world database on other effective area-based conservation measures (WD-OECM), September 2024, Cambridge, UK. .
Vansteelant, W.M.G., Bouten, W., Klaassen, R.H.G., Koks, B.J., Schlaich, A.E., van Diermen, J., et al., 2015. Regional and seasonal flight speeds of soaring migrants and the role of weather conditions at hourly and daily scales. J. Avian Biol. 46, 25-39.
Warnock, N., 2010. Stopping vs. staging: the difference between a hop and a jump. J. Avian Biol. 41, 621-626.
Westrip, J.R.S., Burfield, I.J., Allen, D.J., Numa, C., 2022. The Conservation Status of Breeding Raptors in the Mediterranean. IUCN, Málaga, Spain.
Worton, B.J., 1989. Kernel methods for estimating the utility distribution in home-range studies. Ecology 70, 164-168.
Zuur, A.F., Ieno, E.N., 2016. A protocol for conducting and presenting results of regression-type analyses. Methods Ecol. Evol. 7, 636-645.
Züst, Z., Mukhin, A., Taylor, P.D., Schmaljohann, H., 2023. Pre-migratory flights in migrant songbirds: the ecological and evolutionary importance of understudied exploratory movements. Movement Ecol. 11, 78.
Table
1.
Key movement metrics during the post-fleding dependence period (PFDP) of seven juvenile Montagu's Harriers tracked in central Italy in 2024.
ID bird
Sex of the bird
Start tracking
Onset PFDP
End PFDP
Duration PFDP (days)
Abrupt departure (yes/no)
HR (km2)
CA (km2)
MCP (km2)
Mean daily distance (km)
Max daily distance (km)
Mean distance from the nest (km)
Max distance from the nest (km)
No. of revisits
Total residence time (days)
Residence time/PFDP (%)
T69553
F
06/07/2024
11/07/2024
02/08/2024
22
yes
0.326
0.034
3.600
1.936 ± 2.2
7.913
0.105 ± 0.2
1.964
60
15.911
72.323
T69554
F
15/07/2024
16/07/2024
03/08/2024
18
yes
0.229
0.032
0.430
1.417 ± 1.2
5.113
0.148 ± 0.1
0.861
18
7.534
41.853
T69555*
F
20/07/2024
26/07/2024
07/08/2024*
12*
na
0.042
0.005
0.080
0.308 ± 0.4
1.823
0.061 ± 0.1
0.465
7
9.455
78.794*
T69556
F
20/07/2024
26/07/2024
26/08/2024
31
yes
1495.540
111.759
3533.955
17.234 ± 38.8
132.132
4.176 ± 11.8
96.722
14
9.197
29.668
T69558
F
20/07/2024
30/07/2024
27/08/2024
28
no
141.259
20.151
423.905
16.703 ± 20.5
82.844
4.025 ± 4.5
39.568
45
6.294
22.479
T69559
F
20/07/2024
26/07/2024
13/08/2024
18
no
1.045
0.118
11.348
2.842 ± 3.8
14.454
0.204 ± 0.2
4.999
29
5.763
32.017
T69563
M
20/07/2024
23/07/2024
15/08/2024
23
yes
0.147
0.014
2.264
1.146 ± 1.9
6.847
0.075 ± 0.1
2.890
42
18.273
79.449
ID of the bird (ring code), start tracking date, onset, end and duration of the PFDP (expressed in days), indication on abrupt departure from natal site (yes or no), size of home range (HR, 95% kernel density estimate (KDE)), core areas (CA, 50% KDE) and maximum convex polygon (MCP) estimates expressed in km2 are provided. We also provided estimates of mean and maximum daily distances covered, mean and maximum distance from the nest, number of revisits and total residence time at the natal area, and percentage of residence time over the entire PFDP length. Mean values are reported with their mean ± standard deviation. The asterisk indicates the individual that died prematurely 12 days after fledging and for which the estimates refer only to this period. "na" means data not available.
Table
2.
Parameters estimated from top-ranked GLMMs predicting residence time (A) and time since last visit at the natal area (B) during the post-fleding dependence period (PFDP) of six juvenile Montagu's Harriers tracked in central Italy in 2024 in relation to the time elapsed since the first flight (elapsed days).
Model
Response variable
Predictor
β coefficient
95% CI
A
Residence time at nest (n = 212)
Intercept
3.298
2.673; 3.924
σ2 Individual = 0.5561 (best model)
Elapsed days
−0.155
−0.182; −0.127***
B
Time since last visit (n = 205)
Intercept
0.653
−0.026; 1.332
σ2 Individual = 0.5892 (best model)
Elapsed days
0.093
0.064; 0.123***
Variance of random intercepts (σ2), predictors' coefficient estimates (β), and their 95% confidence intervals (CIs). An asterisk marks coefficients whose CIs do not include "0".
Table
3.
Key movement metrics during the pre-migratory phase (PMP) of six juvenile Montagu's Harriers tracked in central Italy in 2024.
ID bird
Onset PMP
End PMP
Duration PMP (days)
Mean daily distance (km/day)
Max daily distance (km/day)
Max distance from the nest (km)
T69553
02/08/2024
28/09/2024
57
28.598 ± 53.9
254.274
221.631
T69554
03/08/2024
20/09/2024
48
31.943 ± 44.1
170.447
320.814
T69556
26/08/2024
18/09/2024
23
28.013 ± 51.3
231.293
280.804
T69558
27/08/2024
20/09/2024
24
23.963 ± 38.9
163.041
93.675
T69559
13/08/2024
28/09/2024
46
24.645 ± 31.2
154.854
49.113
T69563
15/08/2024
15/09/2024
31
25.252 ± 23.2
98.421
165.916
ID of the bird (ring code), start and end of the PMP and its duration are provided. Mean daily distances and maximum daily distance covered are reported, together with the maximum reached distance from the nest for each individual. Mean values are reported with their ± standard deviation.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds