Ministry of Education Key Laboratory of Ecology and Resource Use of the Mongolian Plateau & Inner Mongolia Key Laboratory of Grassland Ecology, School of Ecology and Environment, Inner Mongolia University, Hohhot, 010020, China
b.
Key Laboratory of Marine Biotechnology of Fujian Province, Institute of Oceanology, College of Marine Sciences, Fujian Agriculture and Forestry University, Fuzhou, 350002, China
c.
Key Laboratory of Zoological Systematics and Evolution, Institute of Zoology, Chinese Academy of Sciences, Beijing, 100101, China
d.
Laboratory of Island Biogeography and Conservation Biology, College of Life Sciences, Nanjing Normal University, Nanjing, 210023, China
Funds:
the Natural Science Foundation of Inner Mongolia, China2023JQ01
the National Key R & D Program of China2019YFA0607103
the Central Government Guides Local Science and Technology Development Fund Projects2022ZY0224
the Open Project Program of 'Ministry of Education Key Laboratory of Ecology and Resources Use of the Mongolian PlateauKF2023003
the Major Science and Technology Project of Inner Mongolia Autonomous Region: Monitoring, Assessment and Early Warning Technology Research of Biodiversity in Inner Mongolia2021ZD0011
Biological specimens are fundamental for taxonomy and flora/fauna research. More importantly, they also play crucial roles in recording environmental impacts on morphology and behavior, which is vital for biodiversity research and conservation. However, there are few systematic studies on the patterns and drivers of bird specimen number at regional scales. This study is the first attempt to examine the relationships between bird specimen number and species traits as well as climate niche breadth in China, aiming to answer two questions: 1) how do species' temperature niche breadth and precipitation niche breadth influence specimen number? 2) which trait is most associated with bird specimen number? The associations between bird specimen number and explanatory variables were examined using ordinary least squares, generalized linear models, phylogenetic generalized linear mixed models, and multiple comparisons. The results showed that Muscicapidae was the family with the highest specimen number, and Passeriformes was the order with the highest specimen number. Bird specimen number significantly increased with larger temperature niche breadth and precipitation niche breadth. Specimen number was also positively associated with geographic range size, habitat specificity, hunting vulnerability and clutch size, but negatively associated with body size. These findings suggest that future bird specimen collection should pay more attention to birds with limited ecological niches, large body sizes, and small clutch sizes. This research enhances the use of bird specimen data to study and preserve biodiversity.
In the 21st century, the earth is undergoing the sixth mass extinction, particularly for terrestrial vertebrates (Ceballos et al., 2015; Tilman et al., 2017). Nevertheless, our ability to forecast the effects of these losses and to create practical mitigation plans is constrained by taxonomic and geographic information (Gardner et al., 2014; Miller et al., 2014). Thus, it is essential to record biodiversity as thoroughly as possible (Hromada et al., 2015). Fortunately, specimen data can provide both temporal and spatial information of species occurrence, and they are reliable data that can document past states of biodiversity (Hromada et al., 2015; Page et al., 2015; Schmitt et al., 2019; He et al., 2021). To solve the biodiversity crisis, advanced ecological and population analyses are needed, and more than 60% of the data for these analyses are provided by specimen collections archived in museums (Kress, 2014; Hromada et al., 2015). For instance, a study in southern Finland examined the association between feather color and climate change using skin specimens from wild brown owls and field survey information, and revealed that recent climate changes have impacted natural selection in wild populations (Karell et al., 2011). IUCN Red List criteria requires specific and detailed information about life history and biology, especially for widely distributed species (Hromada et al., 2015). Therefore, without voucher specimens, the extinction risk of many species cannot be properly assessed (Rocha et al., 2014). In summary, specimens and their related data are crucial for biodiversity research and conservation (Hromada et al., 2015; Zizka et al., 2021). They are also very instrumental and indispensable in fields such as taxonomy, biogeography, molecular biology, ecomorphology and others (Peterson et al., 1998).
However, specimen collections themselves are also at risk (Hromada et al., 2015). Thousands of unidentified and unclassified specimens in museums around the world, sometimes unrecognized for a century or longer, are forgotten, misidentified and unlabeled (Winker et al., 1991; Hromada et al., 2015). Several studies have demonstrated that the value of museum collections is declining and deteriorating (Kress, 2014; Minteer et al., 2014; Rocha et al., 2014; Kemp, 2015). In addition, the rate of animal specimens being collected by natural museums has slowed down significantly in recent times (Prather et al., 2004; Kemp, 2015). Aside from financial and personnel constraints, some individuals and institutions think that removing biological specimens from natural populations may negatively influence community structure, population density, or the survivability of rare species, endangering the integrity of natural populations (Patterson, 2002; McLean et al., 2016; Hope et al., 2018). However, studies have also shown that scientific collection has a negligible contribution to mortality compared with other natural or human-caused impacts (Hope et al., 2018). A survey in Bolivia discovered that the influence of bird specimen collection on local bird populations was insignificant (Remsen, 1995). In particular, continued specimen collection is still crucial in identifying species-level categorization in birds; and conservation priorities could not be determined accurately until species limits are established correctly (Barrowclough et al., 2016; Keck, 2020).
With the development of science and technology, observation data appear in large numbers (Johnston et al., 2020), but studies have shown that observation data can not completely replace the specimen data (Krell and Wheeler, 2014; Daru and Rodriguez, 2023). Although alternative forms of documentation, like photographs, are useful in recording distribution, they may be insufficient for other ecological research aspects (Peterson et al., 1998). Besides, most of the collectors of observational data are from different unprofessional teams (Johnston et al., 2020; Daru and Rodriguez, 2023). They may collect data in areas with strong human influence, so the human bias of data collection is large (Daru and Rodriguez, 2023). The specimen record is of great importance because it allows repeated examination for scientific verification and further study (Peterson et al., 1998). The primary goal of an animal collection is to preserve the species record, serving as a vital resource for both current and future research (Peterson et al., 1998). Each specimen is unique and irreplaceable (Winker et al., 1991). Consequently, it is essential to continuously and scientifically collect biological specimens for biodiversity investigation and research.
The morphological, physiological, and behavioral characteristics that a species develops to adapt to its environment are known as species traits (Killen et al., 2013; Wilman et al., 2014; Isaksson, 2018). These characteristics/traits are closely related to species distribution, range change, and extinction risk (Croci et al., 2008; Maclean and Beissinger, 2017). For example, the extinction probability for birds might increase with larger body size and sensitive hunting vulnerability (Price and Gittleman, 2007; Wang et al., 2018). In addition, birds with wide geographic ranges and high trophic levels are more prone to shift their ranges (Davey et al., 2013; Yang et al., 2020). Collectors have tended to focus on more easily detectable groups of species in the field as well as larger, charismatic groups of organisms (Graham et al., 2004; Newbold et al., 2009). Therefore, the species' traits could also affect specimen number. A study in West Africa found that the long-term, patrol-based wildlife monitoring system's observations are biased, and larger, diurnal and gregarious species (e.g., larger ungulates and primates) are more detectable (Burton, 2012). Unfortunately, few studies have linked species traits to the animal specimen number at regional scales. Environmental factors (e.g., climate and elevation, etc.) can also affect the collection of species specimens (Yang et al., 2014; Monsarrat et al., 2019; Vargas et al., 2022). These factors can influence the distribution, abundance, and behavior of organisms, ultimately affecting the availability and accessibility of specimens for collection (McGill et al., 2006; Vargas et al., 2022). Furthermore, certain studies have observed a phenomenon that specimen collectors and researchers tend to focus their collection efforts on high-altitude areas characterized by greater environmental heterogeneity and richer biodiversity (Parnell et al., 2003; Sánchez-Fernández et al., 2008; Engemann et al., 2015; de Araujo and Ramos, 2021). However, this will cause bias in specimen collection. Studies have indicated that species with narrow environmental niches, including endemic species that are limited to specific geographic areas, can be particularly vulnerable to collection bias (Inman et al., 2021; Baker et al., 2022).
As one of the world's richest countries in biodiversity resources, China has currently recorded 1505 avian species (Zheng, 2023) and abundant biological specimens (He et al., 2021). According to statistics, the National Specimen Information Infrastructure (NSII, http://www.nsii.org.cn/) alone holds more than 20 million animal specimens until 2022. However, few studies have been conducted on the factors that influence bird specimen number in China. Therefore, this study offers the first quantitative analysis of the link between species traits, climate niche breadth, and specimen number in Chinese birds to enhance the use of bird specimen data to study and preserve biodiversity. The following three questions are our focus: 1) which order and family have the most specimens? 2) what is the link between bird specimen number and species' temperature niche breadth and precipitation niche breadth? 3) which species trait is most associated with bird specimen number in China?
2.
Materials and methods
2.1
Data collection
Bird specimen data, comprising 22, 185 specimens, was collected from the NSII (http://www.nsii.org.cn/2017/home.php). The data cover 21 orders, 82 families, and 941 species. Mean annual temperature and mean annual precipitation were obtained from the WorldClim database (https://worldclim.org/). The temperature/precipitation niche breadth of each bird species was calculated as the difference between the maximum and minimum mean annual temperature/precipitation in its distribution area.
Parameters of eleven bird traits were compiled from a recent publication (Wang et al., 2018). These traits include geographic range size (km2), habitat specificity (the number of habitats a species is recorded in), hunting vulnerability (rarely/never hunted or killed, occasionally hunted or killed, and often hunted or killed), body size (mm), clutch size (median number of eggs per nest), nest site (cavity, tree, shrub, water and ground), trophic level (omnivores, granivores, frugivores, nectarivores, insectivores, piscivores and carnivores), migrant status (resident, partial resident and full migrant), nest type (exposed or not), flocking tendency (strictly solitary, occasionally social and strictly social), and dispersal ratio (mean wing length divided by the cube root of mean mass).
To conduct the phylogenetic generalized linear mixed models (PGLMM), a phylogeny for the 941 bird species was pruned from a mega bird phylogeny, which was based on a global species pool and composed of 10,000 trees with 9993 Operational Taxonomic Units (Jetz et al., 2014). The Maximum Clade Credibility tree was built using mean node height, and this process involved sampling 5000 pseudo-posterior distributions and utilizing Tree Annotator in the BEAST package (version 1.8.2) (Drummond and Rambaut, 2007).
2.2
Statistical analyses
The correlations between specimen number and climate niche breadth and traits were examined using the ordinary least squares (OLS) models. The independent variable (bird specimen number) was logarithmically transformed to achieve normally distributed residuals.
Since the dependent variable comprised discrete data and the response variable was counting data, the generalized linear model's (GLM) negative binomial model was utilized to test the links between bird traits and specimen number. Phylogenetic generalized linear mixed models (PGLMM) were also used to correct phylogenetic effects, whereby closely related species tend to share similar traits. In addition, we standardized (mean = 0 and SD = 1) species trait data and climate data to eliminate the effect of the different dimensions of the explanatory variable on the regression coefficient.
Multiple variable analyses were also conducted to test the relative importance of different combinations of species traits. We only used the significant variables based on the previous PGLMM single variable analyses, i.e., geographic range size, habitat specificity, hunting vulnerability, body size and clutch size (Table 1).
Table
1.
Results of single variable phylogenetic generalized linear mixed models (PGLMM) and generalized linear models (GLM). The standardized coefficient (Coef) of PGLMM and GLM and Akaike's information criterion (AIC) of PGLMM and GLM were listed. **p < 0.01; *p < 0.05.
Some trait data are not continuous values but discrete grade variables. For instance, the trophic level of birds was represented by numbers, that is, 1 stand for omnivores, 2–granivores, 3–frugivore, 4–nectarivores, 5–insectivores, 6–piscivores and 7–carnivores. Therefore, this study further examined the effects of nest type, nest site, flocking tendency, hunting vulnerability, trophic level and migration status by one-way Analysis of Variance (one-way ANOVA), as OLS and GLM fitting results did not clearly show the relationships among species traits groups.
Standardized coefficients and Akaike's information criterion (AIC) of each PGLMM and GLM model were listed to assess their relative importance. PGLMM was conducted using the function 'pglmm' in the 'phyr' R package, and GLM was conducted using the function 'glm. nb' in the 'MASS' R package. All analyses were conducted in R 4.2.1.
3.
Results
3.1
Specimen number across bird families and orders
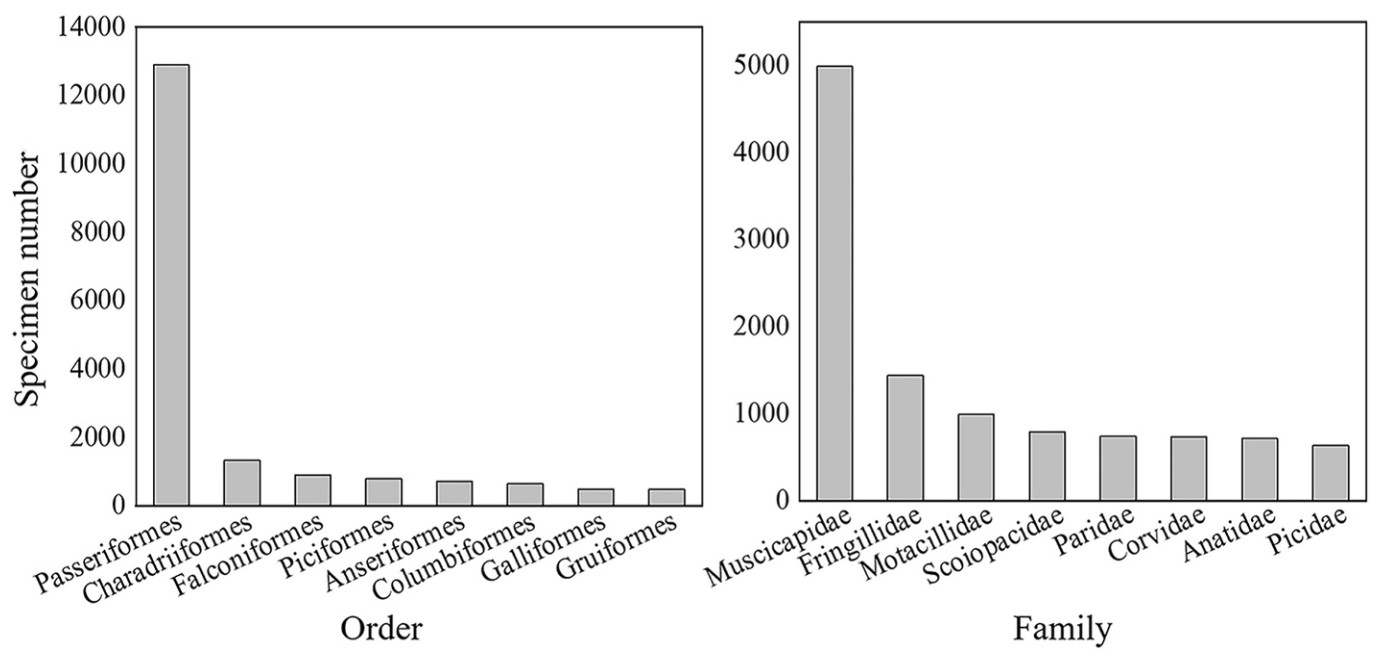
Among the collected bird specimens, Passeriformes accounted for the most specimens, with 12, 904 specimens (63% of all bird specimens). This was followed by Charadriiforme, Falconiformes, Piciformes, Anseriformes, Columbiformes, Galliformes and Gruiformes (Fig. 1), which accounted for more than 17% of the total bird specimen. The family with the most specimens was the Muscicapidae, with 4991 specimens (23% of all bird specimens). This was followed by Fringillidae, Motacillidae, Scoiopacidae, Paridae, Corvidae, Anatidae and Picidae (Fig. 1), which account for about 27% of the total specimen number.
Figure
1.
Specimen number from birds of different orders and families. Only the eight families and orders with the highest values were shown.
3.2
Relationships between specimen number and climate niche breadth and species traits
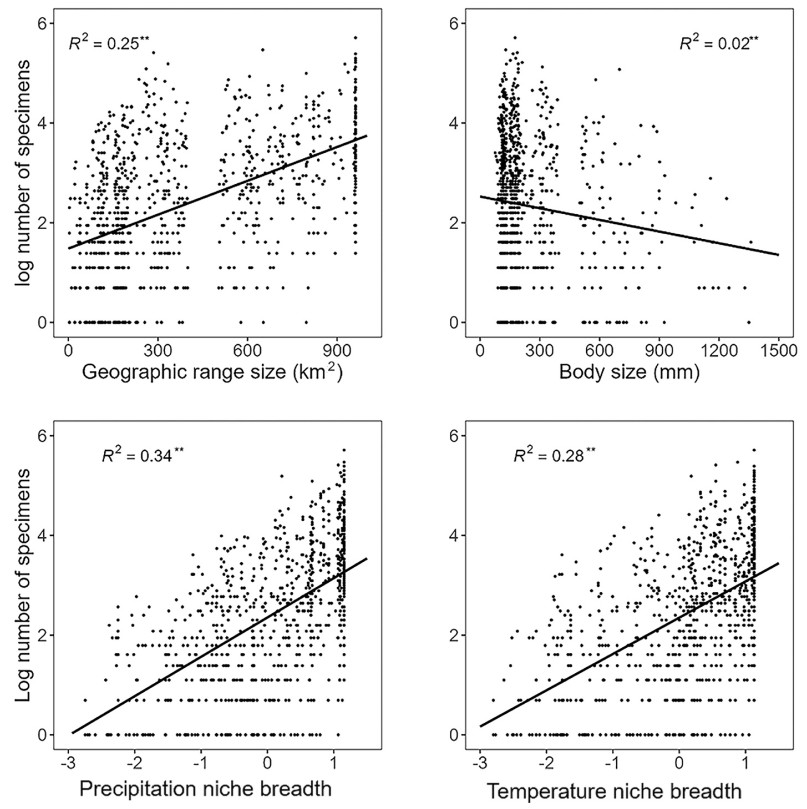
The results of the Ordinary Least Squares (OLS) analysis showed significant positive correlations between bird specimen number and both precipitation niche breadth and temperature niche breadth (Fig. 2). The traits that exhibited the strongest correlations with the bird specimen number were geographic range size (R2 = 0.25**) and body size (R2 = 0.02**) in OLS analysis (Fig. 2). Notably, climate niche breadth had greater influence than species traits, with precipitation niche breadth (R2 = 0.34**) showing a stronger correlation with specimen number compared to temperature niche breadth (R2 = 0.28**, Fig. 2).
Figure
2.
Relationships between specimen number and the climatic niche breadth and species traits (only the two traits with the highest correlation were shown). **p < 0.01; *p < 0.05.
Single variable phylogenetic generalized linear mixed models for discrete data (PGLMM) and generalized linear models (GLM) showed similar patterns regarding the associations between bird species traits and specimen number, i.e., the five traits with the lowest Akaike's Information Criterion (AIC) and highest standardized coefficients were geographic range size, habitat specificity, hunting vulnerability, body size and clutch size (Table 1). However, nest sites and migrant status were only significant in GLM (Table 1). Notably, specimen number showed a positive association with geographic range size, habitat specificity, hunting vulnerability and clutch size, but a negative association with body size.
Multiple variable GLM models revealed that the combination of species traits most strongly associated with specimen number included geographic range size, habitat specificity, hunting vulnerability and body size (Table 2). Notably, the AIC (7456–7554) of GLM for the best seven combinations of variables tended to be lower than the AIC (7590) of the best single variable, i.e., geographic range size (Table 1 and 2).
Table
2.
Results of multiple variables generalized linear models (GLM). The standardized coefficient and Akaike's information criterion (AIC) of GLM were listed. **p < 0.01; *p < 0.05.
GLM
Model description
AIC
Geographic range size (0.58**) + Habitat specificity (0.12**) + Hunting vulnerability (0.35**) + Body size (−0.39**)
7456
Geographic range size (0.58**) + Habitat specificity (0.13**) + Hunting vulnerability (0.36**) + Body size (−0.38**) + Clutch size (−0.04)
7457
Geographic range size (0.61**) + Hunting vulnerability (0.35**) + Body size (−0.40**)
7468
Geographic range size (0.55**) + Habitat specificity (0.12**) + Body size (−0.26**)
7537
Geographic range size (0.55**) + Habitat specificity (0.12**) + Clutch size (0.02) + Body size (−0.26**)
7540
Geographic range size (0.58**) + Body size (−0.26**)
7548
Geographic range size (0.58**) + Hunting vulnerability (0.23**)
7554
Hunting vulnerability (0.29**) + Body size (−0.31**)
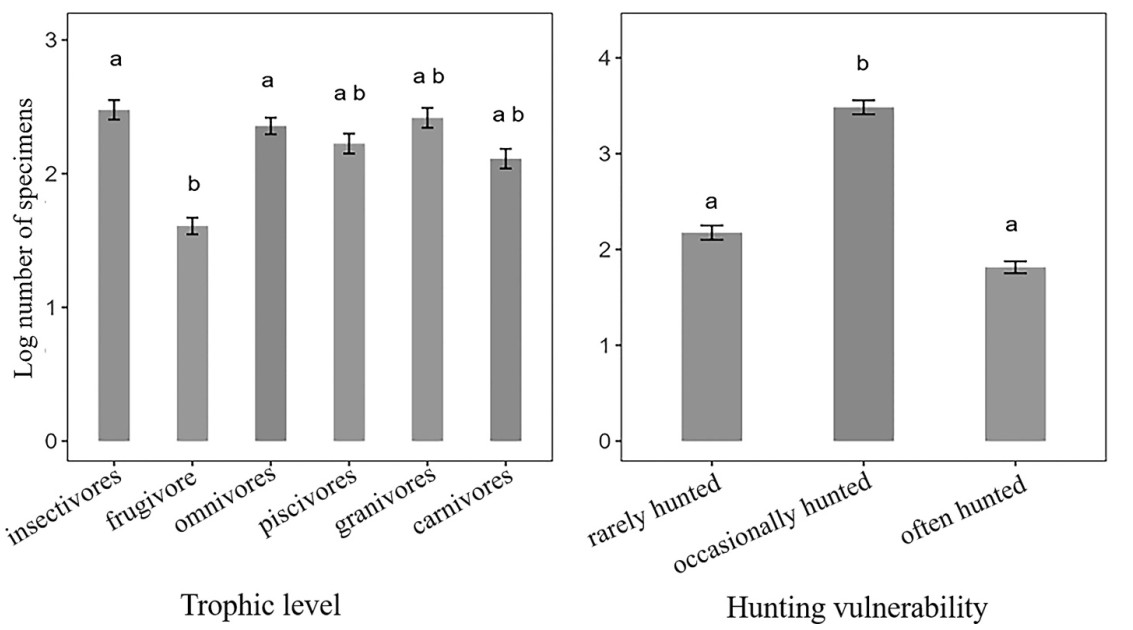
One-way variance analysis showed significant differences in the mean values of nest site, flocking tendency, hunting vulnerability and trophic level. Multiple comparisons revealed significant differences between hunting vulnerability and trophic level groups, while no significant variation was observed among flocking tendency groups. Notably, the specimen number of insectivores and omnivores was significantly higher compared to frugivores (Fig. 3) Specimen number of occasionally hunted was higher than often hunted and rarely hunted (Fig. 3).
Figure
3.
Results of one-way analysis of variance. Only multiple comparison results for trophic level and hunting vulnerability were shown.
This study linked bird specimen number with climatic niche breadth and species traits in China. The results showed that bird specimen number was positively associated with species' temperature niche breadth and precipitation niche breadth. For species traits, bird specimen number was positively associated with geographic range size, habitat specificity, hunting vulnerability and clutch size, but negatively associated with body size.
4.1
Specimen number across bird families and orders
Previous studies indicated that the total number of bird specimens in the world systematic collections surpassesed 9 × 106 (Remsen, 1995; Peterson et al., 1998). If this number was evenly among species, approximately 1000 specimens would exist for each of the 9600 bird species in the world (Peterson et al., 1998). However, despite the record of 1505 bird species in China, the NSII holds only 22, 185 bird specimen records. The bird specimen number recorded in NSII is significantly below the average level, insufficient to support ecological research on geographic variation, species limits, and numerous other life history details (Remsen, 1995). Our analysis indicates that Passeriformes specimens constitute 63% of the total specimen. Passerines are relatively common and well-collected species, indicating that specimens for non-Passeriformes species are considerably less.
4.2
Temperature niche breadth and precipitation niche breadth
Niche breadth typically refers to the range of environmental conditions in which a species can survive and reproduce (Wellborn and Langerhans, 2015), and is generally positively correlated with the distribution range of species (Köckemann et al., 2009; Boucher-Lalonde and Currie, 2016). A study about 3277 bird species and 1659 mammal species in the Americas found a strong positive correlation between species' distribution ranges and ecological niche breadth (Boucher-Lalonde and Currie, 2016). Therefore, larger temperature and precipitation niches can indicate that species may have broader ecological niches, allowing them to occupy a wider range of habitats and exhibit a more extensive geographic distribution (Brown, 1984; Boucher-Lalonde and Currie, 2016). Species that occur in a broader range of climatic conditions may hold greater tolerance to climatic variability, and could therefore face less risk of extinction (Maclean and Beissinger, 2017; Hällfors et al., 2023). In this study, we tried to link species' temperature niche breadth and precipitation niche breadth to specimen number and found that species' temperature niche breadth and precipitation niche breadth were positively correlated with bird specimen number. For example, Alcedo atthis has larger temperature niche breadth (range from −4.26 ℃ to 23.37 ℃) and precipitation niche breadth (range from 41.61 mm to 2064.64 mm), as well as most specimen number (302 specimens). The temperature and precipitation niche breadth of Melanocorypha calandra and Lagopus muta were smaller (4.41–22.64 ℃ and 211.16–229.10 mm, 4.41–29.14 ℃ and 179.04–211.16 mm, respectively), and these two species only had one specimen. In line with these findings, many studies have shown that species with narrow environmental niches (e.g., endemic species) were more vulnerable to collection bias (Inman et al., 2021; Baker et al., 2022).
4.3
Geographic range size
Geographic range size is used as a criterion by the IUCN Red List of Threatened Species in estimating species extinction risk (Gupta et al., 2020). Previous studies have indicated that species with narrow geographic ranges are more prone to extinction due to limited population sizes resulting from local ecological constraints (Cohen and Johnston, 1987; Maurer, 1999). In addition, research has demonstrated that species with large geographic ranges generally exhibit higher local abundances (Brown, 1984; Köckemann et al., 2009; Dallas et al., 2017). Positive associations between species abundance and range size have been observed in many groups of organisms (Gaston, 1996; Kotze et al., 2003; Boucher-Lalonde and Currie, 2016). Birds with a wide geographic range often possess greater tolerance to their environment because of their ecological, behavioral, or physiological flexibility (Bonier et al., 2007). Consequently, birds with wide geographic ranges are more likely to be captured as specimens (Meineke and Daru, 2021). For birds, geographic range size provides an effective prediction of collection frequency (Meineke and Daru, 2021), consistent with our results. For instance, Motacilla alba is mainly distributed in Eurasia and widely distributed also in China, with 302 specimens deposited (the highest specimen number in this study).
4.4
Habitat specificity
Habitat specificity is based on the number of habitats used or the number of habitats in which a species is recorded, and a lower value indicates higher habitat specificity (Martin and Boruta, 2013). In general, a species with a broader ecological niche has a higher ability to tolerate climate change by increasing the likelihood of finding a suitable habitat with sufficient resources, thereby enhancing species' survival in the new environment (Angert et al., 2011; Ding et al., 2013; Estrada et al., 2016). A study using a dataset of Danish terrestrial birds found a positive correlation between habitat breadth and species abundance (Bowler et al., 2018). Consistent with these previous studies, the results of this study showed a positive association between bird specimen number and habitat specificity. This suggests that birds with wider habitat breadth are more likely to be detected and collected by collectors.
4.5
Hunting vulnerability
Previous studies have shown that birds most vulnerable to extinction are those that are heavily hunted and persecuted (Wang et al., 2018; Chen et al., 2019). While our findings showed a positive correlation between hunting vulnerability and specimen number. And the one-way ANOVA revealed that occasionally hunted birds were more likely to be collected as specimens than rarely and often hunted birds. This could be related to the fact that local hunters were once used by researchers to collect bird specimens (Boutan, 1993; Keck, 2020). A study in Spain discovered that hunting is still a severe problem for endangered birds; throughout the study period (1989–2016), for species "Near Threatened", "Vulnerable" and "Endangered", a stable number of individuals are hunted each year, but there has been a decrease in the number of "Least Concern" shot over the years (Balmori, 2019). Hunters generally prefer threatened species, and threatened species are less numerous (Altrichter, 2006; Balmori, 2019; Delany et al., 2003). Therefore, the common species that are occasionally hunted are more likely to be collected as specimens.
4.6
Body size
Body size has traditionally been regarded as the most informative trait in avian species due to its direct correlation with several other traits in bird species, including dispersion capabilities, resource needs, trophic levels, extinction vulnerability, and reproductive strategies (Ding et al., 2013; Estrada et al., 2016). A study conducted in North America revealed a positive relationship between body size and abundance in bird communities with no closely related species (ancient lineages), indicating that larger species tend to be more abundant than smaller ones (Cotgreave, 1994). Conversely, another study using a dataset on the abundance and distribution of breeding birds in the UK showed a negative correlation between abundance and body size across bird species (Gregory, 1995). Furthermore, previous research has consistently demonstrated that smaller bird species exhibit greater adaptability to climate change and lower extinction risk compared to larger birds (Spooner et al., 2018; Wang et al., 2018; Xu et al., 2023). Small birds often possess shorter generation times, higher reproductive rates, and are generally easier to be detected (Coetzee and Chown, 2016). Hence, in theory, smaller birds with lower extinction risk and high reproductive rates are more abundant (Collen et al., 2011; Harnik et al., 2012). In alignment with these previous findings, this research found a negative correlation between specimen number and body size, meaning that smaller birds were more likely to be collected as specimens.
4.7
Clutch size
Clutch size refers to the median number of eggs laid in a single brood by a nesting pair of birds, and it has been shown to have a significant predictive value for species extinction risk (Lyon, 2003; Wang et al., 2018). Smaller clutch sizes can render animals more vulnerable to random events and anthropogenic activities, increasing their sensitivity to habitat change and extinction risk (Lino et al., 2019). On the contrary, a study on capercaillie in European Russia indicated that clutch size reflects the degree of saturation of the entire environment as a whole by birds, and the population reduction was always followed by an increase in clutch size when the habitat is stable (Borchtchevski, 2017). In this study, the clutch size showed strong positive correlations with bird specimen number. Large clutch sizes indicate bird populations with a flush of resources, low competition, and high reproduction rates (Alerstam and Högstedt, 1982; Lundblad and Conway, 2021). This also indirectly supports our conclusion.
4.8
Trophic levels
Higher trophic levels increase species' vulnerability to extinction since they are expected to be less likely to respond to climate changes (Purvis et al., 2000; Buckley and Kingsolver, 2012). The one-way ANOVA findings revealed that omnivorous and insectivorous birds were more likely to be collected as specimens than frugivorous birds. What is noteworthy is that insectivorous birds have higher trophic levels than frugivorous birds. The majority of insectivorous birds are small in body size, which supports our conclusion of smaller birds were more likely to be collected as specimens. Insectivorous birds mainly belong to the order Passeriformes and Muscicapidae (Supahan, 2022), which explains patterns of Chinese bird specimen number, i.e., Passeriformes and Muscicapidae have the most specimens. Omnivorous birds, like sparrows, dwell in urban or rural areas and interact directly with the local community (Isaksson, 2018). They are more prevalent and more detectable to zoologists.
5.
Conclusions
This study analyzed the associations between bird specimen number and species traits as well as climate niche breadth in China. Although there are many other important factors affecting bird specimen number, e.g., social economic and technological development conditions, these factors were not the focus of this study. The current analyses based on available datasets could also provide some useful results. Specifically, the results showed that bird specimen number was significantly correlated with both species' traits and the species' temperature niche breadth and precipitation niche breadth. In the future, modern monitoring technologies (e.g., camera traps and bird watching) should pay more attention to the occurrence data of birds with limited ecological niches, limited geographic range size, low hunting vulnerability, large body size, and small clutch size, which could help to compensate for the bias in specimen collection. This study helps to better utilize bird specimen data for the study and preservation of biodiversity. Future research may pay more attention to the relationships between specimen number and environmental and socio-economic development to amplify specimen data implication in biodiversity research.
Declaration of competing interests
These authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Ethics statement
Ethics approval was not required for this study, though institutional guidelines for the care and use of animals were followed.
Altrichter, M., 2006. Wildlife in the life of local people of the semi-arid Argentine Chaco. Biodivers. Conserv. 15, 2719-2736.
Alerstam, T., Högstedt, G., 1982. Bird migration and reproduction in relation to habitats for survival and breeding. J. Avian Biol. 13, 25-37. .
Angert, A.L., Crozier, L.G., Rissler, L.J., Gilman, S.E., Tewksbury, J.J., Chunco, A.J., 2011. Do species' traits predict recent shifts at expanding range edges? Ecol. Lett. 14, 677-689. .
Baker, David J, Maclean, Ilya, MD, Goodall, Martin, Gaston, Kevin J, 2022. Correlations between spatial sampling biases and environmental niches affect species distribution models. Glob. Ecol. Biogeogr. 31 (6), 1038–1050.
Balmori, A., 2019. Endangered bird mortality by gunshots: still a current problem. Biodivers. Conserv. 28, 2555-2564. .
Barrowclough, G. F, Cracraft, J., Klicka, J., Zink, R.M., 2016. How many kinds of birds are there and why does it matter? PLoS One 11, e0166307. .
Bonier, F., Martin, P.R., Wingfield, J.C., 2007. Urban birds have broader environmental tolerance. Biol. Lett. 3, 670-673. .
Borchtchevski, V.G., 2017. Does beginning of spring represent a critical period in annual cycle of capercaillie (Tetrao urogallus, Tetronidae, Galliformes)? Biol. Bull. 44, 751-760. doi: ISSN:1062-3590.
Boucher-Lalonde, V., Currie, D.J., 2016. Spatial autocorrelation can generate stronger correlations between range size and climatic niches than the biological signal—a demonstration using bird and mammal range maps. PLoS One 11, e0166243. .
Boutan, E., 1993. Le nuage et la Vitrine: Une vie de Monsieur David. Editions R, Chabaud.
Bowler, D.E., Heldbjerg, H., Fox, A.D., O'Hara, R.B., Böhning‐Gaese, K., 2018. Disentangling the effects of multiple environmental drivers on population changes within communities. J. Anim. Ecol. 87, 1034-1045. https://www.jstor.org/stable/2461494.
Brown, J.H., 1984. On the relationship between abundance and distribution of species. Am. Nat. 124, 255-279. .
Buckley, L.B., Kingsolver, J.G., 2012. Functional and phylogenetic approaches to forecasting species' responses to climate change. Annu. Rev. Ecol. Evol. Syst. 43, 205-226. https://www.jstor.org/stable/23323132.
Burton, A.C., 2012. Critical evaluation of a long-term, locally-based wildlife monitoring program in West Africa. Biodivers. Conserv. 21, 3079-3094. doi: https://doi.org/ .
Ceballos, G., Ehrlich, P.R., Barnosky, A.D., García, A., Pringle, R.M., Palmer, T.M., 2015. Accelerated modern human–induced species losses: entering the sixth mass extinction. Sci. Adv. 1, e1400253.
Chen, C.W., Zeng, D., Zhao, Y.H., Wu, Y.R., Xu, J.F., Wang, Y.P., 2019. Correlates of extinction risk in Chinese endemic birds. Avian Res. 10, 8. .
Coetzee, B.W.T., Chown, S.L., 2016. Land‐use change promotes avian diversity at the expense of species with unique traits. Ecol. Evol. 6, 7610-7622. .
Cohen, A.S., Johnston, M.R., 1987. Speciation in brooding and poorly dispersing lacustrine organisms. Palaios 2, 426-435. .
Collen, B., McRae, L., Deinet, S., De Palma, A., Carranza, T., Cooper, N., et al., 2011. Predicting how populations decline to extinction. Philos. Trans. R. Soc. B 366, 2577-2586. .
Cotgreave, P., 1994. The relation between body size and abundance in a bird community: the effects of phylogeny and competition. Philos. Trans. R. Soc. B 256, 147-149. .
Croci, S., Butet, A., Clergeau, P., 2008. Does urbanization filter birds on the basis of their biological traits. Condor 110, 223-240. .
Dallas, T., Decker, R.R., Hastings, A., 2017. Species are not most abundant in the centre of their geographic range or climatic niche. Ecol. Lett. 20, 526-1533. .
Daru, B.H., Rodriguez, J., 2023. Mass production of unvouchered records fails to represent global biodiversity patterns. Nat. Ecol. Evol. 7, 816-831. .
Davey, C.M., Devictor, V., Jonzén, N., Lindström, Å., Smith, H.G., 2013. Impact of climate change on communities: revealing species' contribution. J. Anim. Ecol. 82, 551-561. .
de Araujo, M.L., Ramos, F.N., 2021. Targeting the survey efforts: gaps and biases in epiphyte sampling at a biodiversity hotspot. For. Ecol. Manag., 498, 119544 .
Delany, S., Haanstra, L., Thorup, O., 2003. How many of the world's wader species are declining, and where are the globally threatened species. WSG Bull. 101, 13.
Ding, Z.F., Feeley, K.J., Wang, Y.P., Pakeman, R.J., Ding, P., 2013. Patterns of bird functional diversity on land-bridge island fragments. J. Anim. Ecol. 82, 781-790. .
Drummond, A.J., Rambaut, A., 2007. BEAST: bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 7, 214. .
Engemann, K., Enquist, B.J., Sandel, B., Boyle, B., Jorgensen, P.M., Morueta-Holme, N., et al., 2015. Limited sampling hampers "big data" estimation of species richness in a tropical biodiversity hotspot. Ecol. Evol. 5, 807-820. .
Estrada, A., Morales-Castilla, I., Caplat, P., Early, R., 2016. Usefulness of species traits in predicting range shifts. Trends Ecol. Evol. 31, 190-203. .
Gardner, J.L., Amano, T., Sutherland, W.J., Joseph, L., Peters, A., 2014. Are natural history collections coming to an end as time-series? Front. Ecol. Environ. 12, 436-438.
Gaston, K., 1996. The multiple forms of the interspecific abundance-distribution relationship. Oikos 76, 211-220. https://www.jstor.org/stable/26530250.
Gupta, G., Dunn, J., Sanderson, R., Fuller, R., McGowan, P.J.K., 2020. A simple method for assessing the completeness of a geographic range size estimate. Global Ecol. Biogeogr. 21, e00788 .
Graham, C.H., Ferrier, S., Huettman, F., Moritz, C., Peterson, A.T., 2004. New developments in museum-based informatics and applications in biodiversity analysis. Trends Ecol. Evol. 19, 497-503.
Gregory, R.D., 1995. Phylogeny and relations among abundance, geographical range and body size of British breeding birds. Philos. Trans. R. Soc. B 349, 345-351. .
Hallfors, M.H., Heikkinen, R.K., Kuussaari, M., Lehikoinen, A., Luoto, M., Pöyry, J., et al., 2023. Recent range shifts of moths, butterflies, and birds are driven by the breadth of their climatic niche. Evol. Lett. 2023, qrad004.
Harnik, P.G., Simpson, C., Payne, J.L., 2012. Long-term differences in extinction risk among the seven forms of rarity. P. Natl. Acad. Sci. USA 279, 4969-4976. https://www.jstor.org/stable/41727774.
He, P., Chen, J., Kong, H.Z., Cai, L., Qiao, G.X., 2021. Important supporting role of biological specimen in biodiversity conservation and research. Bull. Chin. Acad. Sci. 36, 425-435.
Hope, A.G., Sandercock, B.K., Malaney, J.L., 2018. Collection of scientific specimens: benefits for biodiversity. Sciences and limited impacts on communities of small mammals. Bioscience 68, 35-42. .
Hromada, M., Čanády, A., Mikula, P., Peterson, A.T., Tryjanowski, P., 2015. Old natural history collections for new millennium–birds and mammals in the collection of PhMr. Tibor Weisz in Sarisske Museum Bardejov, Slovakia. Acta Univ. Presov. 7, 115-141.
Inman, R., Franklin, J., Esque, T., Nussear, K., 2021. Comparing sample bias correction methods for species distribution modeling using virtual species. Ecosphere 12, e03422. .
Isaksson, C., 2018. Impact of urbanization on birds. In: Tietze, D. (Ed.), Bird Species. Fascinating Life Sciences. Springer, Cham, pp. 235–257. .
Jetz, W., Thomas, G.H., Joy, J.B., Redding, D.W., Hartmann, K., Mooers, A.O., 2014. Global distribution and conservation of evolutionary distinctness in birds. Curr. Biol. 24, 919-930. .
Johnston, A., Moran, N., Musgrove, A., Fink, D., Baillie, S.R., 2020. Estimating species distributions from spatially biased citizen science data. Ecol. Model. 422, 108927. .
Karell, P., Ahola, K., Karstinen, T., Valkama, J., Brommer, J.E., 2011. Climate change drives microevolution in a wild bird. Nat. Commun. 2, 208. doi: https://doi.org/ .
Keck, F., 2020. Biosecurity and the ecologies of conservation. An anthropology of collecting practices among virus hunters and birdwatchers. Horizontes Antropol. 26, 93-114. .
Kemp, C., 2015. The endangered dead. Nature 518, 292-294. .
Killen, S.S., Marras, S., Metcalfe, N.B., McKenzie, D.J., Domenici, P., 2013. Environmental stressors alter relationships between physiology and behaviour. Trends Ecol. Evol. 28, 651-658. .
Köckemann, Benjamin, Buschmann, Holger, Leuschner, Christoph, 2009. The relationships between abundance, range size and niche breadth in Central European tree species. J. Biogeogr. 36 (5), 854–864.
Lino, A., Fonseca, C., Rojas, D., Fischer, E., Pereira, M.J.R., 2019. A meta-analysis of the effects of habitat loss and fragmentation on genetic diversity in mammals. Mamm. Biol. 94, 69-76. .
Lundblad, C.G., Conway, C.J., 2021. Ashmole's hypothesis and the latitudinal gradient in clutch size. Biol. Rev. 96, 1349-1366. .
Lyon, B.E., 2003. Egg recognition and counting reduce costs of avian conspecific brood parasitism. Nature 422, 495-499. .
Maclean, S.A., Beissinger, S.R., 2017. Species' traits as predictors of range shifts under contemporary climate change: a review and meta‐analysis. Global Change Biol. 23, 4094-4105. .
Martin, L.B., Boruta, M., 2013. The impacts of urbanization on avian disease transmission and emergence. In: Diego, G., Henrik, B. (Eds.), Avian Urban Ecology. Oxford University Press, Oxford, pp. 116–128.
Maurer, B.A., 1999. Untangling Ecological Complexity: the Macroscopic Perspective. University of Chicago Press, Chicago. .
McGill, B.J., Enquist, B.J., Weiher, E., Westoby, M., 2006. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 21, 178-185. .
McLean, B.S., Bell, K.C., Dunnum, J.L., Abrahamson, B., Colella, J.P., Deardorff, E.R., et al., 2016. Natural history collections-based research: progress, promise, and best practices. J. Mammal. 97, 287-297. .
Meineke, E.K., Daru, B.H., 2021. Bias assessments to expand research harnessing biological collections. Trends Ecol. Evol. 36, 1071-1082. .
Miller, J.A., Miller, J.H., Pham, D.S., Beentjes, K.K., 2014. Cyberdiversity: improving the informatic value of diverse tropical arthropod inventories. PLoS One 9, e115750.
Monsarrat, S., Boshoff, A.F., Kerley, G.I.H., 2019. Accessibility maps as a tool to predict sampling bias in historical biodiversity occurrence records. Ecography 42, 125-136. .
Newbold, T., Gilbert, F., Zalat, S., El-Gabbas, A., Reader, T., 2009. Climate-based models of spatial patterns of species richness in Egypt's butterfly and mammal fauna. J. Biogeogr. 36, 2085-2209.
Page, L.M., MacFadden, B.J., Fortes, J.A., Soltis, P.S., Riccardi, G., 2015. Digitization of biodiversity collections reveals biggest data on biodiversity. Bioscience 65, 841-842. https://www.jstor.org/stable/90007349.
Parnell, J.A.N., Simpson, D.A., Moat, J., Kirkup, D.W., Chantaranothai, P., Boyce, P.C., et al., 2003. Plant collecting spread and densities: their potential impact on biogeographical studies in Thailand. J. Biogeogr. 30, 193-209. .
Patterson, B.D., 2002. On the continuing need for scientific collecting of mammals. Mastozool. Neotrop. 9, 253–262. ISSN 1666-0536.
Peterson, A.T., Navarro-Sigüenza, A.G., Benítez-Díaz, H., 1998. The need for continued scientific collecting; a geographic analysis of Mexican bird specimens. Ibis 140, 288-294.
Prather, L.A., Alvarez-Fuentes, O., Mayfield, M.H., Ferguson, C.J., 2004. The decline of plant collecting in the United States: a threat to the infrastructure of biodiversity studies. Syst. Bot. 29, 15-28. .
Price, S.A., Gittleman, J.L., 2007. Hunting to extinction: biology and regional economy influence extinction risk and the impact of hunting in artiodactyls. Proc. R. Soc. B 274, 1845-1851. https://www.jstor.org/stable/25249258.
Purvis, A., Gittleman, J.L., Cowlishaw, G., Mace, G.M., 2000. Predicting extinction risk in declining species. Proc. R. Soc. B 267, 1947-1952. .
Remsen, J.V., 1995. The importance of continued collecting of bird specimens to ornithology and bird conservation. Bird. Conserv. Int. 5, 146-180. .
Rocha, L.A., Aleixo, A., Allen, G., Almeda, F., Baldwin, C.C., Barclay, M.V., et al., 2014. Specimen collection: an essential tool. Science 344, 814-815.
Sánchez-Fernández, D., Lobo, J.M., Abellán, P., Ribera, I., Millán, A., 2008. Bias in freshwater biodiversity sampling: the case of Iberian water beetles. Divers. Distrib. 14, 754-762. .
Schmitt, C.J., Cook, J.A., Zamudio, K.R., Edwards, S.V., 2019. Museum specimens of terrestrial vertebrates are sensitive indicators of environmental change in the Anthropocene. Philos. T. Roy. Soc. B 374, 20170387. .
Spooner, F.E.B., Pearson, R.G., Freeman, R., 2018. Rapid warming is associated with population decline among terrestrial birds and mammals globally. Global Change Biol. 24, 4521-4531. .
Supahan, N., 2022. Avian assemblage during the development of rice in organic and inorganic rice paddies and its relation to insect pests. Curr. Appl. Sci. Technol. 22, 10.55003. .
Tilman, D., Clark, M., Williams, D.R., Kimmel, K., Polasky, S., Packer, C., 2017. Future threats to biodiversity and pathways to their prevention. Nature 546, 73-81.
Vargas, C.A., Bottin, M., Särkinen, T., Richardson, J.E., Raz, L., Garzon-Lopez, C.X., et al., 2022. Environmental and geographical biases in plant specimen data from the Colombian Andes. Bot. J. Linn. Soc. 200, 451-464. .
Wang, Y.P., Si, X.F., Bennett, P.M., Chen, C.W., Zheng, D., Zhao, Y.H., et al., 2018. Ecological correlates of extinction risk in Chinese birds. Ecography 41, 782-794. .
Wellborn, G.A., Langerhans, R.B., 2015. Ecological opportunity and the adaptive diversification of lineages. Ecol. Evol. 5, 176-195. .
Wilman, H., Belmaker, J., Simpson, J., de la Rosa, C., Rivadeneira, M.M., Jetz, W., 2014. EltonTraits 1.0: species‐level foraging attributes of the world's birds and mammals. Ecology 95, 2027-2027. .
Winker, K., Fall, B.A., Klicka, J.T., Parmelee, D.F., Tordoff, H.B., 1991. The importance of avian collections and the need for continued collecting. LOON 63, 238-246.
Xu, A.C., Zhang, J., Li, Q., Li, Z.Q., Zhu, Q., 2023. The benefits of being smaller: consistent pattern for climate-induced range shift and morphological difference of three falconiforme species. Avian Res. 14, 100079 .
Yang, W., Ma, K., Kreft, H., 2014. Environmental and socio-economic factors shaping the geography of floristic collections in China. Global Ecol. Biogeogr. 23, 1284-1292. .
Yang, X.T., Wang, Y.P., Si, X.F., Feng, G., 2020. Species traits linked with range shifts of Chinese birds. Glob. Ecol. Conserv. 21, e00874. .
Zheng, G.M., 2023. Checklist on the Classification and Distribution of the Birds of China. Science Publishing House, Beijing.
Zizka, A., Antonelli, A., Silvestro, D., 2021. Sampbias, a method for quantifying geographic sampling biases in species distribution data. Ecography 44, 25-32. .
Table
1.
Results of single variable phylogenetic generalized linear mixed models (PGLMM) and generalized linear models (GLM). The standardized coefficient (Coef) of PGLMM and GLM and Akaike's information criterion (AIC) of PGLMM and GLM were listed. **p < 0.01; *p < 0.05.
Table
2.
Results of multiple variables generalized linear models (GLM). The standardized coefficient and Akaike's information criterion (AIC) of GLM were listed. **p < 0.01; *p < 0.05.
GLM
Model description
AIC
Geographic range size (0.58**) + Habitat specificity (0.12**) + Hunting vulnerability (0.35**) + Body size (−0.39**)
7456
Geographic range size (0.58**) + Habitat specificity (0.13**) + Hunting vulnerability (0.36**) + Body size (−0.38**) + Clutch size (−0.04)
7457
Geographic range size (0.61**) + Hunting vulnerability (0.35**) + Body size (−0.40**)
7468
Geographic range size (0.55**) + Habitat specificity (0.12**) + Body size (−0.26**)
7537
Geographic range size (0.55**) + Habitat specificity (0.12**) + Clutch size (0.02) + Body size (−0.26**)
7540
Geographic range size (0.58**) + Body size (−0.26**)
7548
Geographic range size (0.58**) + Hunting vulnerability (0.23**)
7554
Hunting vulnerability (0.29**) + Body size (−0.31**)

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds