Nest site selection is a vital component of bird reproduction success, and an adaptive behavior conducted to decrease nest predation risk with avoiding external disturbances. Understanding patterns of nest site selection can provide insights into how species adapt to changes in their habitat and has important conservation implications. In this study, we used microhabitat variables and multi-scale data with a field survey of nest occurrence to determine nest site selection patterns and adaptive strategies of the breeding Oriental Storks (Ciconia boyciana) in different nest areas. Results demonstrate that the nest site microhabitat characteristics of the breeding Oriental Storks significantly differed among the three nesting areas, and nest height was higher in the middle and lower Yangtze River floodplain than in the Northeast China and Bohai Bay nest areas. The food resources and intensity of human disturbance had the greatest effects on the nest site selection of the breeding Oriental Storks. The intensity of human disturbance was positively correlated with the nest height of the breeding Oriental Storks in Bohai Bay and the middle and lower Yangtze River floodplain; however, nest height decreased with the abundance of food resources in the Northeast China nest area. Our findings indicate that the nest site selection patterns of Oriental Storks showed flexible adaptive strategies. In safer environments, nests were lower and closer to food resources, which allows parent storks to invest more in the nestlings. However, in areas where human activity was intense, nests were higher to ensure the safety of their offspring. Some measures that could be taken to improve the breeding habitat of Oriental Storks include increasing the percentage of wetland areas in nesting areas to enhance food resources availability and setting artificial nests at suitable heights in potential nesting grounds to encourage nesting. Finally, the establishment of soft barriers around the nesting areas could increase the safety of nests.
Nest site selection is a vital component of individual reproduction, which has been considered as an adaptive behavior conducted by parent birds to increase reproduction success (Zhao et al., 2020). This behavior can also affect population dynamics, given that nesting failure is the main cause of unsuccessful reproduction in birds (Martin et al., 1988; Clark and Shutler, 1999; Cheng et al., 2020; Zhao et al., 2020). The “predation avoidance hypothesis” of nest site selection predicts that birds should adjust their nest site preferences according to variations in predation risks to maximize nesting success (Guan et al., 2018). Factors affecting nest site selection in birds can be classified into five groups: a) food resources, such as the abundance, availability, and spatial distribution of food resources (Bradter et al., 2007; Rather et al., 2020); b) nest resources, such as the presence or absence of nest supports, nest support structure, nest stability, nest space, and nest age (Vergara and Aguirre, 2006; Smith and Wilson, 2010; Onmuş et al., 2012); c) nest predation risk, such as the intensity and type of human disturbance, as well as the identity, abundance, and activity patterns of nest predators (Wegge and Storaas, 1990; Burr et al., 2017); d) environmental conditions, such as habitat characteristics, climate conditions, and topographic features (Whittaker et al., 2005; Onmuş et al., 2012; Kou et al., 2020); and e) breeding status of the parent birds, such as the time of arrival at the breeding site, age, and breeding experience (Linz et al., 2008). Food resources are important factors that affect individual reproduction success and offspring survival, and thus affect population dynamics and structures (Martin et al., 1988; Cheng et al., 2020; Rather et al., 2020). Meanwhile, the presence of suitable nest support is a key factor affecting avian nest site selection, especially for tree-nesting waterbird species (Vergara and Aguirre, 2006; Onmuş et al., 2012). The interactions between these factors also affect the nest site selection of birds.
Previous studies have focused on measuring habitat characteristics in small areas around nest sites, but larger-scale characteristics have often been overlooked (Kim and Koo, 2009; Zhang et al., 2017). Global positioning system (GPS) technology has confirmed that the foraging trips of large waterbirds are usually within a radius of 3–5 km around the nest sites (Johst et al., 2001). Given the powerful flight ability of large waterbirds, large-scale habitat features in a radius of several kilometers around the nest site are likely important factors affecting nest site selection (Onmuş et al., 2012; Xu et al., 2021). Several in situ and ex situ conservation measures, habitat restoration, and species re-introductions, have increased the population sizes of endangered species, promoted the recolonization of traditional nesting areas, and expanded the breeding ranges of species (Yu et al., 2006; Ding, 2010; Marucco and McIntire, 2010). Consequently, understanding how individuals adapt to new breeding habitats and how changes in habitat characteristics affect the distribution of nest sites is key to the success of species recovery programs and habitat management plans. (Whittaker et al., 2005).
The Oriental Stork (Ciconia boyciana) is a large migratory wading waterbird species, which is listed as Endangered (EN) on the International Union for Conservation of Nature's (IUCN) Red List of threatened species (BirdLife International, 2018) and the first-class level of the national key protected wildlife species in China. According to the latest results of a recent wintering waterbirds survey in the middle and lower Yangtze River floodplain, the population of wintering storks in China is approximately 7500 individuals (Data from China Wildlife Conservation Association, CWCA, unpublished data). Historically, Oriental Storks nested widely in Northeast Asia, including the Sanjiang Plain of Northeast China and the Amur and Ussuri basins along the China-Russia border in Elm (Ulmus pumila) and Birch Trees (Betula platyphylla) (Liu et al., 2007; Zhou et al., 2013). Their main wintering sites were located in wetlands in the middle and lower Yangtze River floodplain and Southern China (Wang and Yang, 1995; BirdLife International, 2018). In the 1990s, widespread deforestation and habitat loss in the Sanjiang Plain led to a decrease in the population size and nesting habitat of Oriental Storks in Northeast China (Zheng et al., 2016). During this period, Oriental Storks began to nest in electricity pylons in densely populated stopover and wintering areas in China, such as Bohai Bay coast and the middle and lower Yangtze River floodplain (Zhou et al., 2013; Cheng et al., 2020). Similar changes in nesting behavior have been observed in White Storks (Ciconia ciconia) in Europe (Tryjanowski et al., 2009). Oriental Storks were first recorded to breed outside their traditional breeding areas in 1999. Since then, storks have been reported to breed in Wangjiang of Anhui in 2001, Gaoyou of Jiangsu in 2002, Dongying of Shandong in 2003, Poyang of Jiangxi in 2004, Dafeng of Jiangsu in 2009 (Zhou et al., 2013), Yangxin of Hubei in 2016, and Caofeidian of Heibei in 2018. Following the success of the wetland recovery project, artificial nests have been established in the Yellow River Delta, and this provided sufficient nesting sites and space for breeding Oriental Storks. The number of nest sites has increased yearly, and the geographical distribution of nesting sites has expanded continuously (Cheng et al., 2020). In this context, changes in nesting areas and nest site selection patterns of Oriental Storks in new breeding habitats have received increased interest from conservation researchers.
Studies that investigate the effects of various factors on nest site selection can provide valuable information for conservation management. Most previous studies have examined the effects of microhabitat factors on nest site selection, and the effects of large-scale habitat factors on nest site selection have not been widely examined, yet these factors might affect the nest site selection patterns of Oriental Storks, given their strong flight ability (Zhou et al., 2013; Duan et al., 2015; Zhang et al., 2017). Based on the various habitat characteristics that are potentially important to nest site selection of the breeding Oriental Storks, we have the following three predictions. First, Oriental Storks breeding in the Northeast China would prefer to nest near wetlands and that nest height would be lower, while in Bohai Bay and the middle and lower Yangtze River floodplain, the nest height would be higher to ensure the safety of the broods. Second, the storks would prefer to nest in sites near abundant food resources but far from human disturbance. Third, nest height of storks would increase with the intensity of human disturbance and the abundance of food resources around the nest. The nest site selection patterns and adaptive strategies observed in this study will aid the conservation of Oriental Storks and the management of their habitats.
2.
Methods
2.1
Study area and nest sample data
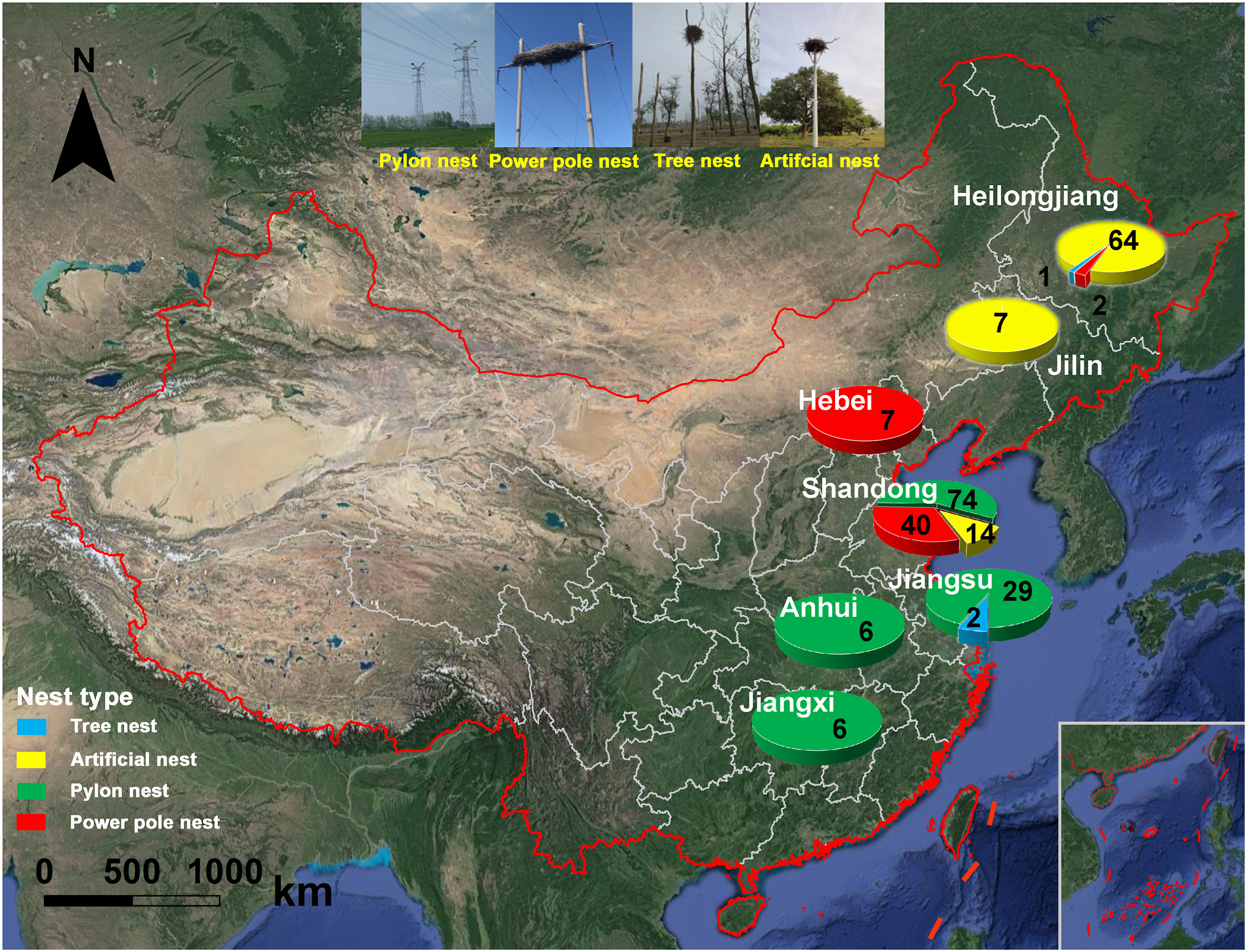
The current breeding ranges of Oriental Storks are mainly located on the Sanjiang Plain and Yellow River Delta, respectively, with a partial distribution near lakes in the middle and lower Yangtze River floodplain in China. Our study areas cover the traditional and new expansion nest areas (Northeast China, Bohai Bay, and the middle and lower Yangtze River floodplain) of Oriental Storks, which incorporates parts of several adjacent provinces (Heilongjiang, Jilin, Hebei, Shandong, Jiangsu, Anhui, and Jiangxi). We obtained the geographical coordinates of 252 nest sites' location for four different nest types of Oriental Storks in breeding areas from 2017 to 2020, including tree nest, power pole nest, pylon nest, and artificial nest (Fig. 1). The Northeast China nest area of Oriental Storks is mainly distributed in the Sanjiang Plain in Heilongjiang and western Jilin, and the habitats of nest sites are mainly marsh and meadow wetlands. As stopover and wintering areas, the Bohai Bay coast and middle and lower Yangtze River floodplain are attracting more and more Oriental Storks breeding in recent years (Zhou et al., 2013). These storks mainly nest on power poles and pylons in mudflat and farmland habitats, as well as in artificial nests (Zhou et al., 2013; Duan et al., 2015; Cheng et al., 2020).
Figure
1.
The nest sample size of the breeding Oriental Storks in study areas during the period from 2017 to 2020.
Based on the previous studies, we evaluated wetland resource variables within a radius of 3 km around the nest sites (Johst et al., 2001). We downloaded Landsat 8 OLI_TIRS images with little or no cloud cover (<10%) for the study area. We interpreted images to obtain the area of different habitats by supervised classification.
Oriental Storks’ food is primarily fish, but it is difficult to accurately estimate the fish biomass. Normally, suitable wetland areas are positively correlated with the abundance of food resources for Oriental Storks (Duan et al., 2015). Therefore, we used the percentage of wetland areas to all habitat areas to characterize the abundance of food resources, such as shallow water areas, farmland, and rivers.
During the field survey, we measured nest height and location with Nikon Laser Rangefinder and GPS respectively which are in the base of a nest support structure. Meanwhile, we collected the intensity of human disturbance to each nest for 15 min along fixed sample lines in the nesting period of storks. Finally, a large quadrat of 10 m × 10 m was centered on each nest, and five 1 m × 1 m small quadrats were set up at four corners and the center of the large one. Habitat characteristics data were collected from all five small quadrats after the parent birds leave the nest. These variables were determined in Table 1. Meanwhile, a quadrat of 10 m × 10 m size was set up randomly as control plot in the study area, and the same variables as those for the nest sites were measured.
Table
1.
Nest site microhabitat variables of the breeding Oriental Storks.
Variables (abbr.)
Definition
Unit
Nest resources
-Nest support structure (NSS)
Power pole/Artificial nest/Pylon pole/Arbor tree
None
-Nest height (NH)
The vertical distance from the nest to the ground
m
Food resources
-The food resources of wetland (FR)
Wetland areas percentage in all habitat areas (Shallow water/Farmland/Rivers)
Percent
Nesting safety
-The intensity of human disturbance (IHD)
The sum of vehicles, humans, and honking
Degree
Habitat characteristic
-Vegetation height (VH)
Mean height of the vegetation per small quadrat
m
-Vegetation density (VD)
Mean density of the vegetation per small quadrat
Ind./m2
-Vegetation coverage (VC)
The ratio of the vertical projection areas of the vegetation to the sample quadrat areas
The factors from a total of 252 nest sites were used for the nest site selection analyses. The overall differences in habitat variables around the nest site among three nest areas were used by analysis of similarity (ANOSIM; permutations = 999) using the vegan package (Version 2.2–0; Oksanen et al., 2014). Furthermore, we used the one-sample Kolmogorov-Smirnov Test to examine whether each variable followed a normal distribution with the statistical software SPSS 20.0. For data that followed a normal distribution, we used the one-way ANOVA to analyze the significance of the difference between the nest site variables; otherwise, the nonparametric test (Kruskal-Wallis H test) was used. We assessed the contribution of each habitat variable around the nest site selection within different nest areas using the random forest models. Then, to test the effects of habitat factors on the nest height selection, we used the logit link functions to fit the generalized linear mixed models (GLMMs) with gamma distributions in all our candidate models. The model analysis of multivariate data was performed in R software using the MuMIn package (Fränzi et al., 2015). Based on the nest height data, we assessed the effects of habitat factors on nest height selection by using three metrics: food resources, intensity of human disturbance, and nest support structure. These variables were determined in Table 1. Finally, to understand the effect of the habitat variables around the nest site on the nesting strategies of the breeding storks, we used linear models (LMs) to analyze the relationships between nest height and explanatory variables.
Before the model analysis of the nest height selection, we used a multicollinearity test to calculate the Variance Inflation Factor (VIF) among the independent variables in R software. If the VIF value of a variable was larger than 2, there were correlations among variables. The VIF values of all independent variables were < 2.0, and thus we considered that none of these variables were correlated with each other and included them all in the following analysis. Then, all factors of metrics fitted the generalized linear mixed models (GLMMs) with gamma distributions. These models included the vegetation height, density, distance to the open water areas, water area percentage in the quadrat, and the intensity of human disturbance as fixed factors, and the nest support structure was used as a random effect. We used Akaike's Information Criterion (AICc) as the evaluation criteria to select the best model. The most parsimonious model with the smallest AICc value was the best model. Further, when the 95% confidence interval for the above coefficients did not cross zero, it provides further evidence that their effects on variables in a model have statistical significance (Arnold, 2010; Umer et al., 2017).
All statistical analyses were performed in the R software 3.6.1 (R Core Team, 2019) and SPSS 22.0 (IBM SPSS Statistics, 2013). A significance level of P = 0.05 was used for all statistical tests and the results were shown as Mean ± SE.
3.
Results
3.1
Nest site microhabitat characteristics
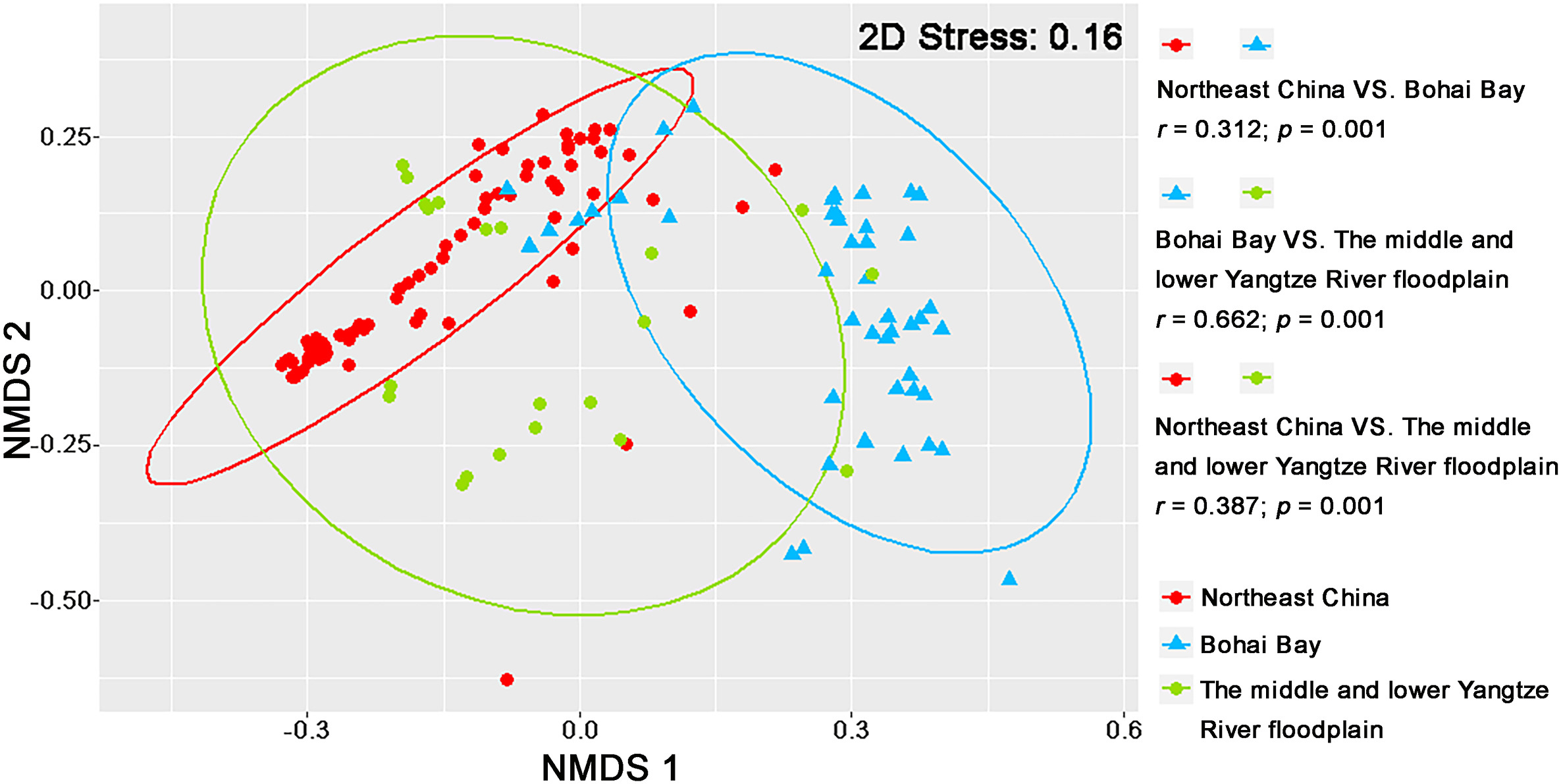
The nest site microhabitat characteristics of the breeding Oriental Storks significantly differed among the three nest areas (Fig. 2). All three vegetation factors, including vegetation height, density, and coverage, were significantly lower in the Northeast China nest area than in the Bohai Bay and the middle and lower Yangtze River floodplain (χ2 = 62.771, P < 0.001; χ2 = 75.894, P < 0.001; χ2 = 26.820, P < 0.001; Table 2). Furthermore, food resources were significantly more abundant in the middle and lower Yangtze River floodplain than in Northeast China and Bohai Bay (χ2 = 78.376, P < 0.001; Table 2); the intensity of human disturbance was also higher in the middle and lower Yangtze River floodplain than in Northeast China and Bohai Bay (Table 2).
Figure
2.
Non-metric multidimensional scaling (NMDS) showing nest site microhabitat characteristics of the breeding Oriental Storks among the three nest areas.
Table
2.
Nest site characteristics of the breeding Oriental Storks among the three nest areas (Abbreviations see Table 1).
Northeast China
Bohai Bay
The middle and lower Yangtze River floodplain
χ2
P
NH
7.47 ± 0.70c
13.94 ± 0.62b
45.87 ± 1.51a
106.602
< 0.001
VH
0.55 ± 0.10b
1.46 ± 0.06a
0.60 ± 0.076b
62.771
< 0.001
VD
91.87 ± 12.87c
280.39 ± 10.05a
148.44 ± 10.95b
75.894
< 0.001
VC
50.42 ± 4.98c
74.55 ± 2.55a
60.67 ± 4.35b
26.820
< 0.001
WA
7.92 ± 2.62b
20.26 ± 2.44a
10.89 ± 1.93b
8.880
< 0.05
DW
248.29 ± 75.87a
210.65 ± 22.23b
170.62 ± 35.98c
9.067
< 0.05
DT
561.46 ± 144.85a
369.00 ± 39.45b
266.58 ± 48.57c
8.770
< 0.05
IHD
0.17 ± 0.08c
1.10 ± 0.21b
4.36 ± 0.35a
69.048
< 0.001
FR
48.96 ± 7.99b
27.03 ± 2.09c
85.33 ± 1.39a
78.376
< 0.001
Note: If two nest areas have the same lowercase letters, they were not statistically significantly different (P > 0.05), according to the Kruskal–Wallis H test.
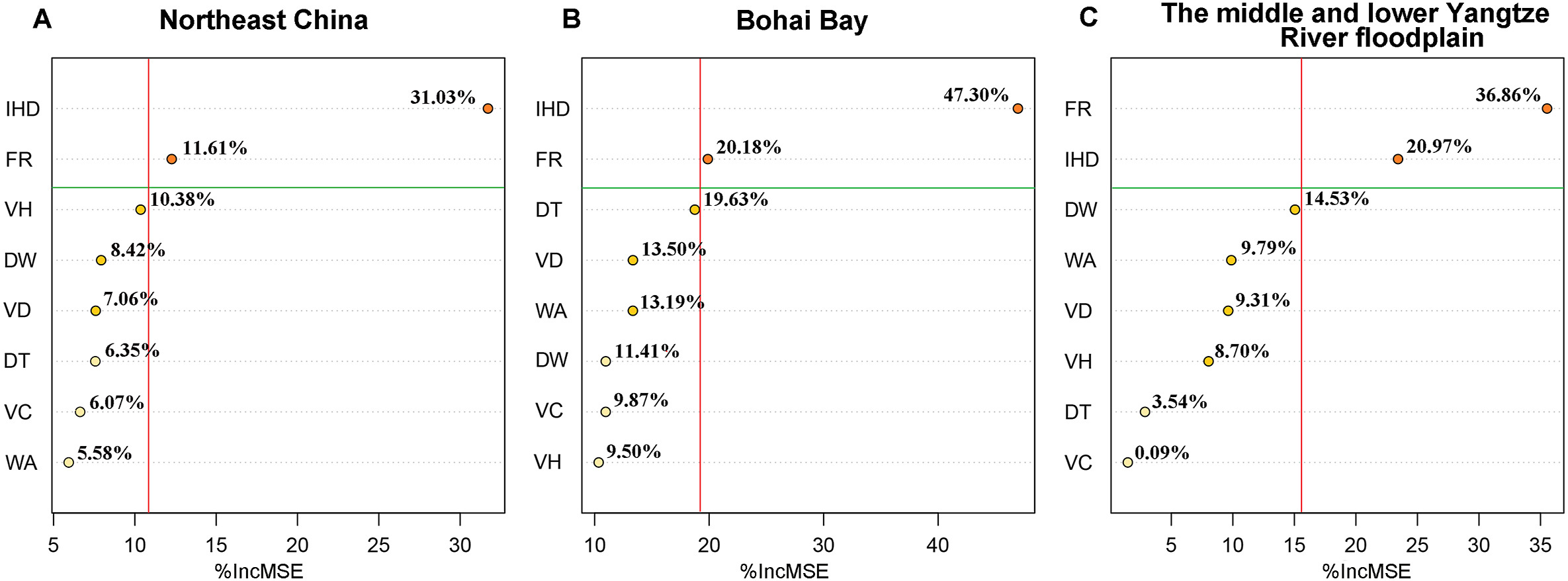
In Northeast China, Oriental Storks mainly nested in areas with low levels of human activities, such as the marsh wetlands. Open reed wetlands were important nesting habitats for the breeding storks in the Bohai Bay. The storks that bred in the middle and lower Yangtze River floodplain preferred to nest in farmland and aquaculture ponds. Specifically, the random forest model showed that the intensity of human disturbance and food resources explained 31.03% and 11.61% of the variation in nest site selection in Northeast China, respectively (Fig. 3A). Similarly, in Bohai Bay, the intensity of human disturbance and food resources made a larger contribution to nest site selection compared with other factors (47.30% and 20.18%, respectively; Fig. 3B). Finally, the abundance of food resources (36.86%) was the most important factor affecting the nest site selection of Oriental Storks in the middle and lower Yangtze River floodplain, followed by the intensity of human disturbance (20.97%; Fig. 3C).
Figure
3.
The relative importance of habitat characteristics in nest site selection among the three nest areas using the random forest model. A, B, and C represent Northeast China, Bohai Bay, and the middle and lower Yangtze River floodplain nest areas, respectively (Abbreviations see Table 1).
3.3
Nest site selection patterns and adaptive strategies
There were significant differences in nest height among the three nest areas. The nest height of the breeding storks was significantly higher in the middle and lower Yangtze River floodplain (NHfloodplain: 45.87 ± 1.51 m) than in the other two nest areas (NHBohai Bay: 13.94 ± 0.62 m; NHNortheast China: 7.47 ± 0.70 m) (χ2 = 106.602, P < 0.001; Table 2). In Northeast China, there were four candidate models, and the best models only included the food resources variable; the support for the combined model was 51.50%. In the Bohai Bay nest area, the best models included food resources and the intensity of human disturbance, and the support for the combined model was 97.00%. In the middle and lower Yangtze River floodplain, the best models only included the intensity of human disturbance, and the support for the combined model was 74.10% (Table 3). The GLMM models were bootstrapped using a resampling approach to evaluate the stability of the models and estimate CIs for all parameters. Food resources (a fixed factor) had a significant effect on nest height in Northeast China (βfood resources = −0.067, 95% CI = −0.109 to −0.002) and Bohai Bay (βfood resources = −0.103, 95% CI = −0.151 to −0.054). The intensity of human disturbance (a fixed factor) significantly affected nest height in Bohai Bay (βdisturbance = 0.905, 95% CI = 0.346 to 1.575) and the middle and lower Yangtze River floodplain (βdisturbance = 2.094, 95% CI = 1.209 to 3.064). Nest support structure (a random factor) affected nest height in Bohai Bay (Std.Devstructure = 3.174, 95% CI = 1.245 to 7.210) and the middle and lower Yangtze River floodplain (Std.Devstructure = 17.300, 95% CI = 4.791 to 51.724) (Table 4).
Table
3.
Results of the generalized linear mixed models (GLMMs) for food resources, nesting safety, and nest resources influencing nest height selection of the breeding Oriental Storks among the three nest areas (Abbreviations see Table 1).
Table
4.
Effects of food resources, nesting safety, and nest resource on the nest height selection of the breeding Oriental Storks among the three nest areas (Abbreviations see Table 1).
Model
Northeast China
Bohai Bay
The middle and lower Yangtze River floodplain
Coefficients (95% credible intervals) for fixed effects
Intercept
10.084 (6.246, 14.551)
16.507 (12.634, 20.601)
20.219 (−12.411, 52.719)
FR
−0.067 (−0.109, −0.002)
−0.103 (−0.151, −0.054)
0.066 (−0.143, 0.278)
IHD
0.396 (−2.949, 3.580)
0.905 (0.346, 1.575)
2.094 (1.209, 3.064)
Standard deviation (95% credible intervals) of random effects
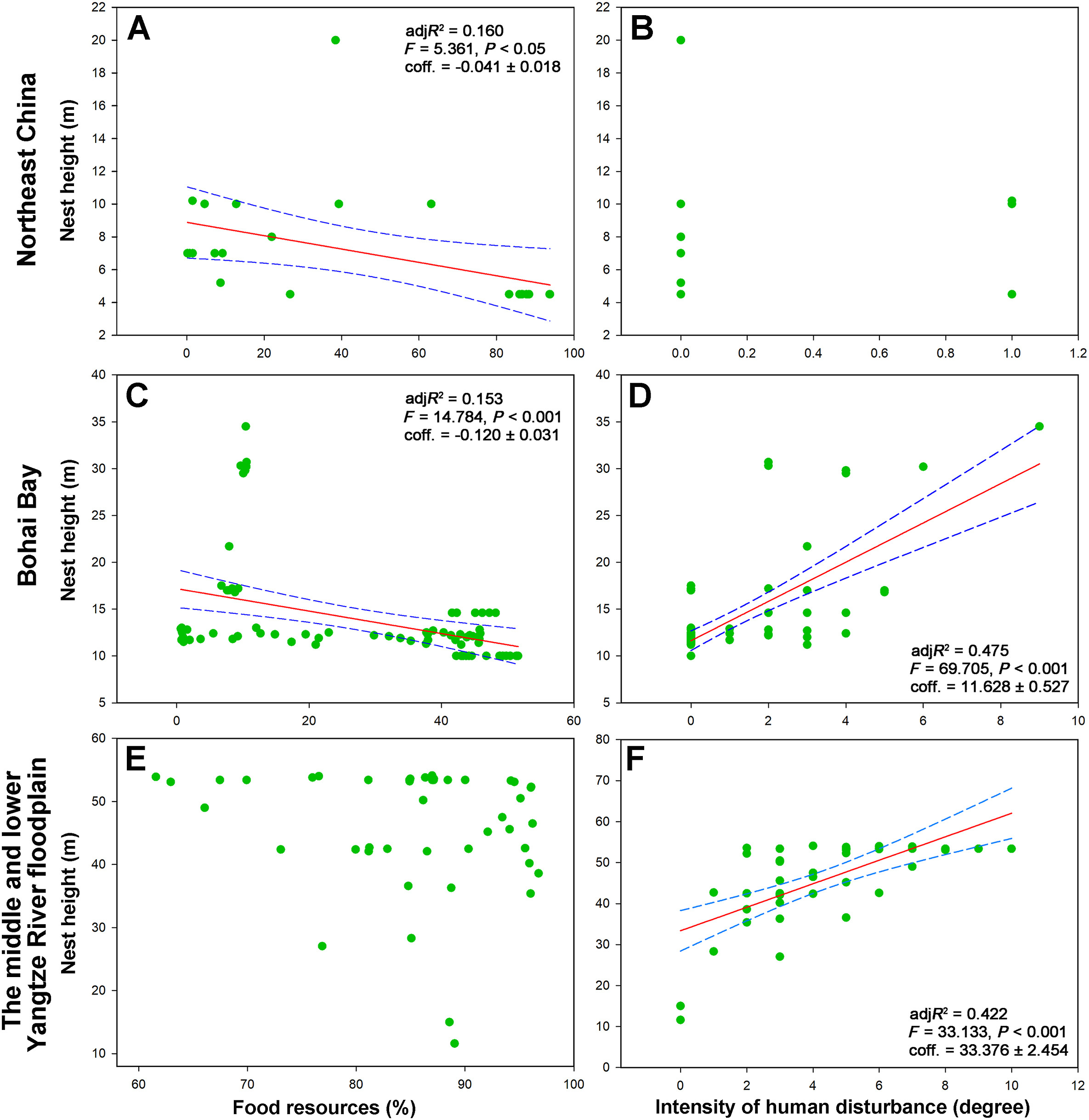
In Northeast China, food resources negatively affected the nest height of the breeding Oriental Storks (P < 0.05, adjR2 = 0.160, coff. = −0.041 ± 0.018); however, the intensity of human disturbance was not correlated with the nest height (P = 0.856) (Fig. 4A, B). In Bohai Bay, nest height decreased with the food resources (P < 0.001, adjR2 = 0.153, coff. = −0.120 ± 0.031) but increased with the intensity of human disturbance (P < 0.001, adjR2 = 0.475, coff. = 11.628 ± 0.527) (Fig. 4C, D). In the middle and lower Yangtze River floodplain, the nest height of the breeding Oriental Storks was positively correlated with the intensity of human disturbance (P < 0.001, adjR2 = 0.422, coff. = 33.376 ± 2.454) (Fig. 4F). We found that food resources had a more significant effect on the nest height of breeding Oriental Storks in Bohai Bay than in Northeast China (Fig. 4A, C). Moreover, the effect of human disturbance on nest height was stronger in the middle and lower Yangtze River floodplain than in Bohai Bay (Fig. 4D, F).
Figure
4.
Food resources and the intensity of human disturbance with the nest height of the breeding Oriental Stork among the three nest areas.
The present study aimed to reveal the effects of habitat characteristics relative to the nest site selection patterns among the three nest areas of Oriental Storks and the underlying adaptive mechanisms. We found that the nest site selection preference of storks showed flexible adaptive strategies because of human activities and food resources. The intensity of human disturbance was positively correlated with the nest height of the breeding Oriental Storks in Bohai Bay and the middle and lower Yangtze River floodplain. Due to the low level of human disturbance in Northeast China, nest height decreased as the abundance of food resources increased. Our results indicate that both microhabitat characteristics and multi-scale habitat features in different nesting areas need to be considered when developing habitat conservation strategies for Oriental Storks.
The nest site selection and reproductive success of Oriental Storks depend on the conditions of wetlands, where high levels of food resources (fish and aquatic plant roots; Zheng et al., 2016) are available. Storks are highly dependent on rivers and marshlands, given that over 70% of their food comprises fish, amphibians, and aquatic invertebrates (Cheng, 1994). Aside from the food resources in the wetlands, aquaculture ponds near the areas of shallow water also provided abundant food sources for the breeding storks, such as the Red Swamp Crayfish (Procambarus clarkii), which is similar to previous studies of White Storks (Ferreira et al., 2019; Burdett et al., 2022). Research has shown that when the areas of suitable habitat (i.e., the food resources of wetlands) are not enough to meet the needs of nesting birds, artificial nests are not effective for attracting birds (Zheng et al., 2016). Consequently, freshwater bodies (water bodies or rivers), aquaculture ponds, and farmland are important foraging habitats for Oriental Storks because they provide sufficient food resources to support breeding.
The nest site selection patterns of individuals vary among breeding environments. Typically, birds are expected to actively select new breeding habitats or alter their nesting behavior to avoid potential threats when resource conditions change and breeding success cannot be guaranteed in their original nesting sites (Hunter et al., 2016; Ma et al., 2019). A previous study had shown that some bird species build nests in areas with high levels of disturbance, indicating that they exhibit plasticity in nest-building behavior, and parental birds' experience might play a key role in enhancing the efficiency of nest building (Clauser and Mcrae, 2016). For example, breeding King Rail (Rallus elegans) construct significantly taller nests at sites with greater variation in water level (Clauser and Mcrae, 2016). Similarly, Clapper Rail (Rallus crepitans) parents construct nests far away from the tidal influences to avoid nest inundation (Gaines et al., 2003). Our results indicated that nests of Oriental Storks in Northeast China and Bohai Bay were mostly distributed in areas with abundant vegetation and far away from human activities, and those in the middle and lower Yangtze River floodplain were near villages and aquaculture ponds. Although human activities might negatively affect breeding performance (Vergara and Aguirre, 2006), foraging in human-mediated landscapes might be more time and energy efficient than in natural habitats. These findings are consistent with observations of White Storks in Europe, which efficiently use foraging opportunities provided by anthropized environments such as landfills under the background of expansion and population growth (Catry et al., 2017; Soriano-Redondo et al., 2021). Although the Oriental Storks have not been verified for this with our data, we will focus on this question in the future.
Several studies have shown that vehicle horns and noises reduce the use of nests closer to highways because of transport level development (Shu et al., 2006; Parris and Schneider, 2009). Meanwhile, with the popularity and growth of birding tourism, bird photographers tried to approach the nesting areas on foot to take more clear pictures (Tan et al., 2022). However, the activities of birders can have negative effects on parenting storks, such as inducing some storks to temporarily flee or even abandon their nests (Cheng et al., 2020). Hence, storks breeding in the middle and lower Yangtze River floodplain, as well as other ecotourism areas, have begun to nest at higher sites to avoid disturbance in areas with high densities of humans (Li and Zhang, 2018).
Alternatively, the height of the nest support structure is also a key factor affecting the nesting position and space for breeding (Vergara and Aguirre, 2006; Onmuş et al., 2012). Although the height of the nest support structure is fixed, storks alter the height of their nests when the intensity of human disturbance is high. Storks might abandon their nests if they are unable to establish nests at heights needed to ensure breeding success. In other words, a safer nest to reduce nesting risk may be a trade-off with avoiding external disturbances (Clauser and Mcrae, 2016; Hunter et al., 2016; Zhao et al., 2020). Meanwhile, having a safe and stable nest support structure is a key factor affecting nest site selection in birds, especially for large tree-nesting waterbirds (Vergara and Aguirre, 2006; Onmuş et al., 2012).
Overall, the relationships between the nest sites of storks and human disturbance reflect the tendency of storks to avoid anthropogenic pressure in disturbed and human-dominated landscapes. Enhancing nesting habitat quality via wetland recovery, establishing artificial nests in suitable areas, and maintaining food resources at high densities are particularly important measures for ensuring the long-term survival of Oriental Storks. Meanwhile, soft barriers should be established between humans and nesting habitats, such as fences and ditches, to mitigate the effect of human disturbance on storks. Our results highlight the importance of taking a holistic perspective that considers both microhabitat variables and larger-scale habitat characteristics of nesting areas when developing conservation strategies for birds, especially for endangered species.
5.
Conclusion
Our study revealed the highly flexible variation in nest site selection patterns and adaptive strategies of Oriental Storks as their breeding areas expand. Our results showed that nest site microhabitat characteristics of the storks significantly differed among nest areas. The nest height of individuals breeding in the middle and lower Yangtze River floodplain and Bohai Bay was high, which was presumably to ensure safety from human disturbance, and the nest height of birds in Northeast China was lower than that of birds in new breeding areas. These results are related to the condition of local food resources, nesting resources, and human disturbance during the breeding seasons. Our analysis of microhabitat variables and larger-scale habitat characteristics provided new insights into the characteristics of nest sites and adaptive nesting strategies of Oriental Storks. The results have implications for the conservation management of other endangered waterbirds.
Authors' contributions
LC and LZ conceived the study. LC and CY contributed to the data collection; LC, ZW, and CL conducted the experiments and analyzed the data; LC, CY, ZW, and CL led the writing and revision of the manuscript. All authors read and approved the final manuscript.
Ethics statement
We obtained approval from all the reserve management offices for the fieldwork. All the observations were conducted under the natural status of Oriental Storks and have no influence on the breeding of storks. All the field observations conducted in this study were performed in accordance with the current laws of China.
Declaration of competing interest
The authors declare that they have no competing interests.
Acknowledgements
We appreciate the staff of the Yellow River Delta National Nature Reserve Management Bureau, Xianghai National Nature Reserve Management Bureau, Dafeng Milu National Nature Reserve Management Bureau, and Honghe National Nature Reserve Management Bureau for their help in the fieldwork. We are grateful to Lianshan Li, Baoguang Zhu, Kai Shan, Lixin Wu, and Guanghai Feng for their assistance at the field site. The manuscript benefited from helpful suggestions by Xi Wang. We especially express appreciation to Zhiwen Yuan, Wanxiang Wu, Weibao Xue, Jiangong Xu, and Yiqun Zhang for helping with the data analyses of the manuscript. This work was supported by the National Natural Science Foundation of China (Grant No. 32171530 and 31472020).
Arnold, T.W., 2010. Uninformative parameters and model selection using Akaike’s information criterion. J. Wildlife Manage. 74, 1175-1178. .
BirdLife International, 2018. Ciconia boyciana. The IUCN Red List of Threatened Species 2018: e.T22697695A131942061. doi: . (Accessed 14 February 2023).
Bradter, U., Gombobaatar, S., Uuganbayar, C., Grazia, T.E., Exo, K.M., 2007. Time budgets and habitat use of White-naped Cranes Grus vipio in the Ulz river valley, north-eastern Mongolia during the breeding season. Bird. Conserv. Int. 17, 259-271. .
Burdett, E.M., Muriel, R., Morandini, V., Kolnegari, M., Ferrer, M., 2022. Power lines and birds: drivers of conflict-prone use of pylons by nesting White Storks (Ciconia ciconia). Diversity 14, 984. .
Burr, P.C., Robinson, A.C., Larsen, R.T., Newman, R.A., Ellis-Felege, S.N., 2017. Sharp-tailed grouse nest survival and nest predator habitat use in North Dakota’s Bakken oil field. PLoS One 12, e0170177. .
Catry, I., Encarnacao, V., Pacheco, C., Catry, T., Tenreiro, P., da Silva, L.P., et al., 2017. Recent changes on migratory behavior of the White Stork (Ciconia ciconia) in Portugal: towards the end of migration? Airo 24, 28-35.
Cheng, L., 1994. Diet analysis of the Oriental White Stork in the middle reaches of Heilongjiang River. Chinese J. Wildlife 78, 29-32. (In Chinese)
Cheng, L., Zhou, L.Z., Wu, L.X., Feng, G.H., 2020. Nest site selection and its implications for conservation of the endangered Oriental Stork Ciconia boyciana in Yellow River Delta, China. Bird. Conserv. Int. 30, 323-334. .
Clark, R.G., Shutler, D., 1999. Avian habitat selection: pattern from process in nest-site use by ducks? Ecology 80, 272-287. .
Clauser, A.J., McRae, S.B., 2016. King Rails (Rallus elegans) vary building effort and nest height in relation to water level. Waterbirds 39, 268-276. .
Ding, C.Q., 2010. Crested Ibis. Chinese Birds 1, 156-162. .
Duan, Y.B., Tian, X.H., Ma, J.Z., Zhu, S.Y., Shan, K., 2015. Foraging habitat use of the oriental white stork during its breeding season. Acta Ecol. Sin. 35, 2628-2634.. (In Chinese)..
Ferreira, E.M., Grilo, F., Mendes, R.C., Lourenço, R., Santos, S.M., Petrucci-Fonseca, F., 2019. Diet of the White Stork (Ciconia ciconia) in a heterogeneous Mediterranean landscape: the importance of the invasive Red Swamp Crayfish (Procambarus clarkii). Airo 26, 33-47. .
Fränzi, K.N., Tobias, R., Stefanie, V.F., Jérôme, G., Bettina, A., Pius, K.N., 2015. Bayesian Data Analysis in Ecology Using Linear Models with R, BUGS, and Stan. Academic Press, London.
Gaines, K.F., Cumbee, J.C., Stephens, W.L., 2003. Nest characteristics of the Clapper Rail in coastal Georgia. J. Field Ornithol. 74, 152-156. .
Guan, H.H., Wen, Y., Wang, P.C., Lei, L., Xu, J.L., Li, J.Q., 2018. Seasonal increase of nest height of the Silver-Throated Tit (Aegithalos glaucogularis): can it reduce predation risk? Avian Res. 9, 42. .
Hunter, E.A., Nibbelink, N.P., Cooper, R.J., 2016. Threat predictability influences seaside sparrow nest site selection when facing trade-offs from predation and flooding. Anim Behav. 120, 135-142. .
IBM SPSS Statistics, 2013. SPSS for Windows Release 22.0. IBM Corporation, Armonk. .
Johst, K., Brandl, R., Pfeifer, R., 2001. Foraging in a patchy and dynamic landscape: human land use and the White Stork. Ecol. Appl. 11, 60-69. .
Kim, J.S., Koo, T.H., 2009. Nest site selection and reproductive success of herons and egrets in Pyeongtaek Heronry, Korea. Waterbirds 32, 116-122. .
Kou, J., Wang, T., Yu, F., Sun, Y., Feng, C., Shao, X., 2020. The moss genus Didymodon as an indicator of climate change on the Tibetan Plateau. Ecol. Indicat. 113, 106204. .
Li, S., Zhang, X., 2018. Land use-based human activity intensity along the Yangtze River economic belt, China (1970s–2015). China Sci. Data 3, 1–8. (In Chinese) doi: .
Linz, G.M., Sawin, R.S., Lutman, M.W., 2008. The influence of breeding experience on nest success in Red-winged Blackbird. West. N. Am. Naturalist 74, 123-129. .
Liu, H.Y., Li, Z.F., Li, X.M., 2007. Ecological effects on Oriental White Stork (Ciconia boyciana) with habitat loss in sub-Sanjiang Plain, China. Acta Ecol. Sin. 27, 2678-2683.. (In Chinese).
Ma, L.K., Zhang, J.W., Liu, J.P., Yang, C.W., Liang, W., Møller, A.P., 2019. Adaptation or ecological trap? Altered nest-site selection by Reed Parrotbills after an extreme flood. Avian Res. 10, 2. .
Martin, T.E., Roper, J.J., Thomas, E., Roper, J.J., 1988. Nest predation and nest-site selection of a western population of the Hermit Thrush. Condor 90, 51-57. .
Marucco, F., McIntire, E.J.B., 2010. Predicting spatio-temporal recolonization of large carnivore populations and livestock depredation risk: wolves in the Italian Alps. J. Appl. Ecol. 47, 789-798. .
Oksanen, J., Blanchet, F.G., Kindt, R., Legendre, P., Minchin, P.R., O’Hara, R.B., et al., 2014. Vegan: Community Ecology Package. Version 2.2-0. .
Onmuş, O., Ağaoğlu, Y., Gül, O., 2012. Environmental factors affecting nest-site selection and breeding success of the White Stork (Ciconia ciconia) in western Turkey. Wilson J. Ornithol. 124, 354-361. .
Parris, K.M., Schneider, A., 2009. Impacts of traffic noise and traffic volume on birds of roadside habitats. Ecol. Soc. 14, 1698-1707. .
R Core Team, 2019. R: a Language and Environment for Statistical Computing. .
Rather, T.A., Kumar, S., Khan, J.A., 2020. Multi-scale habitat modelling and predicting change in the distribution of tiger and leopard using random forest algorithm. Sci. Rep. 10, 11473. .
Shu, Y., Hu, Y.M., Leng, W.F., Zhu, S.Y., Shan, K., 2006. Habitat selection of Red Crowned Crane (Grus japonensis) in Yellow River Delta. Chinese J. Ecol. 8, 954-958. (In Chinese).
Smith, P.A., Wilson, S., 2010. Intraseasonal patterns in shorebird nest survival are related to nest age and defence behaviour. Oecologia 163, 613-624. .
Soriano-Redondo, A., Franco, A.M.A., Acácio, M., Martins, B.H., Moreira, F., Catry, I., 2021. Flying the extra mile pays-off: foraging on anthropogenic waste as a time and energy-saving strategy in a generalist bird. Sci. Total Environ. 782, 146843. .
Tryjanowski, P., Kosicki, J.Z., Kuzniak, S., Sparks, T.H., 2009. Long-term changes and breeding success in relation to nesting structures used by the White Stork, Ciconia ciconia. Ann. Zool. Fenn. 46, 34-38. .
Umer, A., Kelley, G.A., Cottrell, L.E., Giacobbi Jr, P., Innes, K.E., Lilly, C.L., 2017. Childhood obesity and adult cardiovascular disease risk factors: a systematic review with meta-analysis. Publ. Health 17, 683. .
Vergara, P., Aguirre, I.J., 2006. Age and breeding success related to nest position in a White Stork Ciconia ciconia colony. Acta Oecol. 30, 414-418. .
Wang, Q.S., Yang, Z.F., 1995. Current studies on Oriental White Stork. J. Anhui Univ. 19, 82-99. (In Chinese).
Wegge, P., Storaas, T., 1990. Nest loss in Capercaillie and Black Grouse in relation to the small rodent cycle in Southeast Norway. Oecologia 82, 527-530. .
Zhang, L.X., An, B., Shu, M.L., Yang, X.J., 2017. Nest-site selection, reproductive ecology and shifts within core-use areas of Black-necked Cranes at the northern limit of the Tibetan Plateau. PeerJ 5, e2939. .
Zhao, J.M., Yang, C., Lou, Y.Q., Shi, M., Fang, Y., Sun, Y.H., 2020. Nesting season, nest age, and disturbance, but not habitat characteristics, affect nest survival of Chinese Grouse. Curr. Zool. 66, 29-37. .
Zheng, H.F., Shen, G.Q., Shang, L.Y., Lv, X.G., Wang, Q., McLaughlin, N., et al., 2016. Efficacy of conservation strategies for endangered Oriental White Storks (Ciconia boyciana) under climate change in Northeast China. Biol. Conserv. 204, 367-377. .
Zhou, L.Z., Xue, W.W., Zhu, S.Y., Shan, K., Chen, J.L., 2013. Foraging habitat use of Oriental White Stork (Ciconia boyciana) recently breeding in China. Zool. Sci. 30, 559-564. .
Table
2.
Nest site characteristics of the breeding Oriental Storks among the three nest areas (Abbreviations see Table 1).
Northeast China
Bohai Bay
The middle and lower Yangtze River floodplain
χ2
P
NH
7.47 ± 0.70c
13.94 ± 0.62b
45.87 ± 1.51a
106.602
< 0.001
VH
0.55 ± 0.10b
1.46 ± 0.06a
0.60 ± 0.076b
62.771
< 0.001
VD
91.87 ± 12.87c
280.39 ± 10.05a
148.44 ± 10.95b
75.894
< 0.001
VC
50.42 ± 4.98c
74.55 ± 2.55a
60.67 ± 4.35b
26.820
< 0.001
WA
7.92 ± 2.62b
20.26 ± 2.44a
10.89 ± 1.93b
8.880
< 0.05
DW
248.29 ± 75.87a
210.65 ± 22.23b
170.62 ± 35.98c
9.067
< 0.05
DT
561.46 ± 144.85a
369.00 ± 39.45b
266.58 ± 48.57c
8.770
< 0.05
IHD
0.17 ± 0.08c
1.10 ± 0.21b
4.36 ± 0.35a
69.048
< 0.001
FR
48.96 ± 7.99b
27.03 ± 2.09c
85.33 ± 1.39a
78.376
< 0.001
Note: If two nest areas have the same lowercase letters, they were not statistically significantly different (P > 0.05), according to the Kruskal–Wallis H test.
Table
3.
Results of the generalized linear mixed models (GLMMs) for food resources, nesting safety, and nest resources influencing nest height selection of the breeding Oriental Storks among the three nest areas (Abbreviations see Table 1).
Table
4.
Effects of food resources, nesting safety, and nest resource on the nest height selection of the breeding Oriental Storks among the three nest areas (Abbreviations see Table 1).
Model
Northeast China
Bohai Bay
The middle and lower Yangtze River floodplain
Coefficients (95% credible intervals) for fixed effects
Intercept
10.084 (6.246, 14.551)
16.507 (12.634, 20.601)
20.219 (−12.411, 52.719)
FR
−0.067 (−0.109, −0.002)
−0.103 (−0.151, −0.054)
0.066 (−0.143, 0.278)
IHD
0.396 (−2.949, 3.580)
0.905 (0.346, 1.575)
2.094 (1.209, 3.064)
Standard deviation (95% credible intervals) of random effects

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds