
Figure
1.
Males conflicting at the border of territories in spring (Photo by Yue-Hua Sun)
| Citation: |
Hongxing JIANG, Fawen QIAN, Chunyue LIU, Xiaoming LI, Yunqiu HOU, Guogang ZHANG, Ming DAI, Dongping LIU. 2012: Impact of marsh changes on breeding cranes in Sanjiang Plain, northeastern China. Avian Research, 3(3): 165-179. DOI: 10.5122/cbirds.2012.0026

|
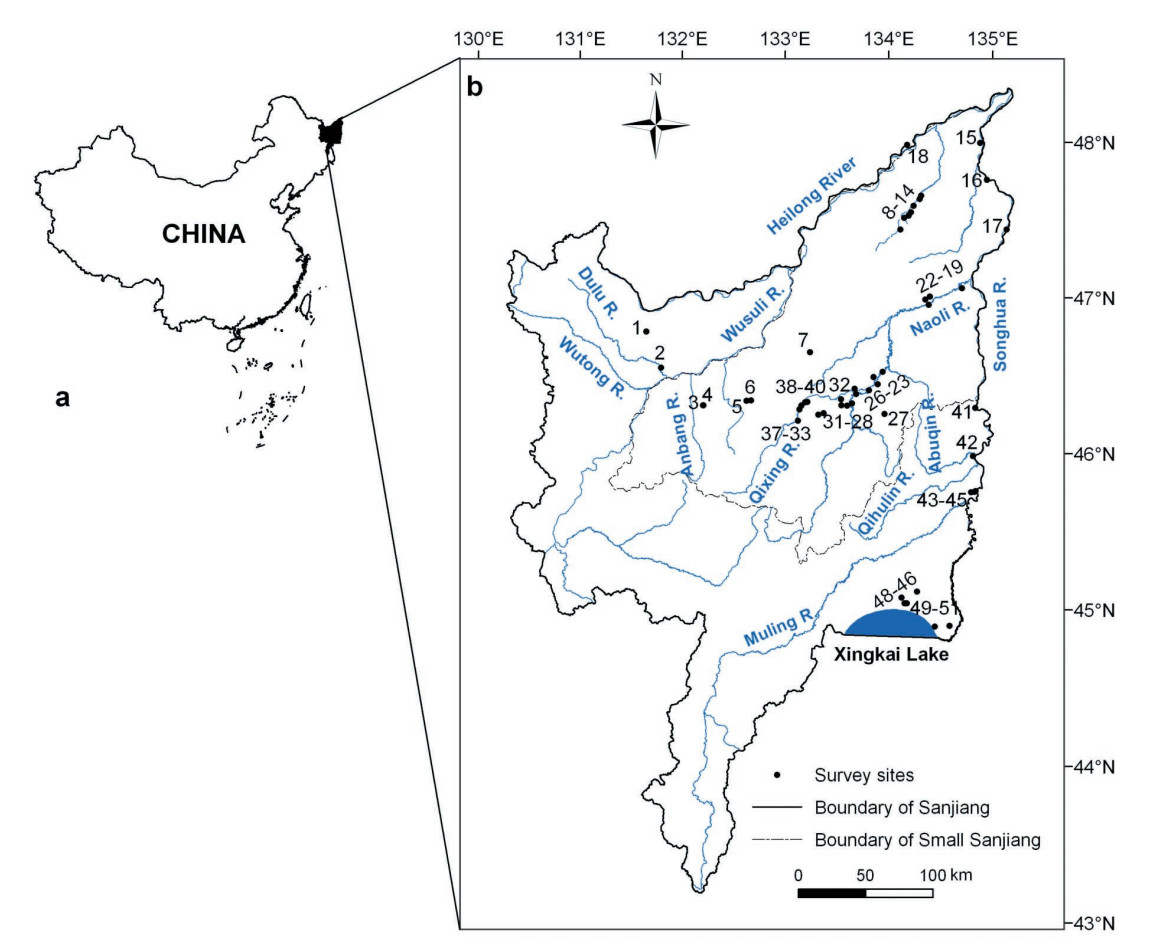
In order to measure the impact of changes in the marsh on breeding Red-crowned Cranes (Grus japonensis) and White-naped Cranes (G. vipio) in the Sanjiang Plain, we conducted a complete crane survey in each of the summers of 2007 and 2008, and obtained information on the changes in its marsh through interpretation of satellite images in 1986, 1995, 2000 and 2005. A comparison between the 1984 and 2008 census shows that: 1) the number of Red-crowned Cranes remained stable at about 300 birds, while the number of White-naped Cranes increased markedly; 2) the estimates of nests of Red-crowned cranes are close to the numbers of 1984, while approximately 90% of the nests of these two crane species were distributed over five national nature reserves (NNRs); 3) the two crane species became more concentrated in three regions, i.e., the Fuyuan Delta, the middle reaches of the Naoli and Qixing rivers, and the Xingkai Lake wetlands, rather as formerly in six regions. The area of the marsh decreased by 45% within the 20-year period from 1986 to 2005. The major loss of the marsh area was in the form of a conversion to paddy fields and dry land, which can be primarily attributed to direct human activity under various economic and policy drivers. The disappearance of cranes in the Dulu and Abuqin rivers was induced by large-scale agricultural development. The fragmentation into and isolation of small areas of the marsh became increasingly more serious, which resulted in unsuitable condtions for crane breeding in small patches of the marsh. The number of cranes declined from 1984-1995 but had recovered remarkably by 2008, which may be due to the establishment of a system of nature reserves, in place since 2000. The land ownership of the reserve plays a key role in maintaining the integrity of the wetland ecosystem, especially in Sanjiang, Honghe, Naolihe and Qixinghe NNRs, which together accommodated approximately 80% of the population of cranes. However, we did not discover any nests at all in the provincial and municipal reserves, which may be attributed to high human disturbances in these areas. In the end, we propose some recommendations on reserve management.
The Chinese Grouse (Bonasa sewerzowi) is a bird endemic to China, distributed in the conifer-dominated mountains between 2400 and 4300 m elevation, living in the most southerly range of grouses in the world (from about 27° to 39°N) and considered to be one of the rarest grouse species in the world, listed as "Lower Risk (near threatened)" by the IUCN in 1996. In China, it has been listed as an endangered species, in Category Ⅰ of nationally protected animals (Zheng and Wang, 1998; Sun, 2000)
Knowledge about the natural history of a bird is essential for its conservation (Crandall et al., 2000); however, little was known before 1995 on the natural history characteristics of the Chinese Grouse (Wang et al., 1987; Liu and Geng, 1994). Since 1995, we started a systematical study on this bird at the Lianhuashan Nature Reserve in the south of Gansu Province, western China (Sun et al., 2003). Here we summarize the main results achieved so far about this endemic species.
The territorial Chinese Grouse population at Lianhuashan is stable with a high density of up to 15 occupied territories per square kilometer (Klaus et al., 1998; Sun et al., 2003). In spring, the Chinese Grouse prefers to inhabit coniferous forests, with a good cover of shrubs as their food. It has been suggested that both trees (spruce and fir) and shrubs (including juvenile willows and arrow bamboos) are important vegetation for the conservation of the habitat of the Chinese Grouse (Ji et al., 2005).
According to field data from the population at Lianhuashan during 1995 to 2001, the body mass of males is significantly lower during the breeding season (309.9 ± 22.4 (SD) g, n = 24) than during the non-breeding season (341.2 ± 27.1 g, n = 16). In contrast to males, females measure a higher body mass during the egg-laying and incubation periods (337.8 ± 26.4 g, n = 31) than during the non-breeding season (314.9 ± 20.7 g, n = 24). Compared with currently available body mass data of other grouse species, the Chinese Grouse is considered to be the smallest grouse in the world (Sun et al., 2005).
Around late March to April, males leave the winter flocks and return to their spring territories earlier than the females (Sun, 2004). Males mark their territories by flutter-jumping and flutter-fly, with the peak of such activities occurring in early May (Sun and Fang, 1997; Klaus et al., 1998). Since the sex ratio of the population is skewed towards males (64%, with 44% of the males unmated in the spring) (Sun et al., 2003), competition on the part of male birds for female partners is intensive (Fig. 1); not all males have the chance to mate. The territory of paired males is smaller in size (2.1 ha vs 2.7 ha) and of better quality than that of unpaired males, with smaller areas of overlap (Sun, 2004; Ji et al., 2005). Pair bonding in Chinese Grouse has proven to be very strong, with 89.9 ± 13.9% (n = 8) of the time pairs observed living together. Males show much more alarming behavior during pairing while females spend more time feeding. Sun (2004) suggested that the Chinese Grouse is strictly a monogamous bird.
It has been reported that the Chinese Grouse nests at the base of trees, in tree stumps or on rocky ledges (Hoyo and Sargatal, 1994). From our observations, all 103 nests at Lianhuashan were found to be at the base of trees during 1995–2004 (Fig. 2), with 56.2% at the base of deciduous trees (mainly birch and willow) and 40.4% at the base of coniferous trees. The average number of cover around a nest was 5.08 ± 2.03 (n = 86). The average distance between nests made by the same female in different years was 154.2 ± 109.0 m (n = 19). Compared with the Hazel Grouse (B. bonasia), the Chinese Grouse seems to prefer the base of tree stems as nesting sites (Sun et al., 2007). Females lay eggs every two days. Of 26 first clutches observed, the average clutch size was 6.19 ± 0.16 eggs (ranging from 5–8), and the hatching success of 206 eggs from successful nests was 93% (Sun et al., 2003). The 156 eggs from 26 nests, including those that had been incubated, were measured 20.5 ± 1.62 g in mass, 42.9 ± 0.1 mm in length and 30.4 ± 0.08 mm in width (Sun et al., 2005). Overall incubation constancy was 93.5%. Females took 4.95 ± 0.99 (ranging from 3 to 7) recesses each day during incubation with each recess for 19.3 ± 7.2 min on average. The length of incubation period varied from 27–29 days (Sun, 2004). The downy chicks hatched in late June or early July. Normally, after hatching chicks, the hens lead the broods leaving the nest area, while their paired males continued to live in or around their territories. After living together with hens for around 2–3 months, the chicks dispersed in August or September within distances from 1 to 5 km, according the limited data from Lianhuashan (Y.-H. Sun, unpublished data).

Although less frequent than in spring, the territorial behavior of male Chinese Grouse also occurs in autumn, with noisy flutter-flights, flutter jumps and agonistic behavior of uttering two types of aggressive songs. Fighting is observed mostly in spring and less in autumn. The occupancy of a territory in autumn with a good supply of deciduous trees, which provide winter food sources as well as conifer cover, is assumed to be crucial for the survival of the Chinese Grouse (Klaus et al., 2009). After late October when frost and snow prevail in Lianhuashan, the grouse form into flocks and move to their preferred winter habitats. Most females and all the yearlings move from the breeding areas and form mixed-sex flocks. Males adopt one of three types of social behavior and movement patterns, i.e., staying in the territory, moving to their winter flocking area, or commuting between the spring territory and the wintering area. Food, cover, topography and future breeding opportunity are major variables determining the social behavior of the Chinese Grouse in winter. Flock size might increase to 13 or 14 in early December, varying with habitat and food availability. According to radio-tracking data from 25 male Chinese Grouses, their home range in winter changes from 6.04 ± 1.60 ha (n = 9) to 15.98 ± 2.61 (n = 10) ha (Sun, 1997; Yang et al., 2010). The winter flocks of the Chinese Grouse dissolve around early April and the birds will start a new yearly round of natural history (Table 1).
| Month (Illustrations) | Male | Female |
| April (Flock dissolution and pair bond formation) | Hold territory by flutter-jump and flutter-fly displays (adults leave the flock and return to their territories earlier) | Pair with males with better territory quality (adults leave the winter flock area and return to their breeding area, all females breed) |
| May (Living in pairs) | Hold territory and guard their mates, and juveniles normally have no chance to breed | Feed to gain nutrition in order to lay clutches of 5–8 eggs |
| June (Incubation) | Stay around nests | Incubation, hatching around the end of the month |
| July–August (Brood time) | Stay in or around territory, recovering from heavy investment | Lead the brood leaving the nest area, raising the chicks themselves |
| September (Brood dissolution) | Stay in or around territory | Brood dissolution, chick dispersal for 1–5 km |
| October (Autumn territoriality) | Defend their territories | Wander in territories of males, join small groups of 3–4 birds |
| November-March (Winter flock) | Adopt three strategies: stay in the territory, move between flock area and territory or stay in the flock (juveniles all in flock) | Move back to their winter flock area, most remain in flock with maximum size of 12–13 birds |
 DownLoad:
CSV
DownLoad:
CSV
The primary food source of the Chinese Grouse in winter consists of willow buds and twigs (83% of forage) (Fig. 3). Among the six main willow species, the Kangding Willow (Salix paraplesia) is the most preferred and the Sichuan Willow (S. hylonoma) the least (χ2 = 4.5, p = 0.49), with feeding preference (used/available) more significantly related to the content of tannin (r = –0.83, p = 0.04) than to that of protein, lipid, phosphorous, calcium, ash, fiber, tannin and energy in buds. We suggest that the major determinant of the diet of this herbivorous bird is the maximization of the intake of nutrients while simultaneously minimizing the intake of digestion inhibitors, rather than the rate of intake (Wang et al., in press). The ratio of cecum to small intestine is higher than that of other grouse species of similar weight, such as the Hazel Grouse and the White-tailed Ptarmigan (Lagopus leucurus), possibly due to its adaptation to a fibrous diet (Wang et al., 2010b).

Analysis of crop contents from six birds during the breeding season indicated that willow (Salix spp.) is the primary food for males, whereas both willow and Dragon Spruce (Picea asperata) seeds constitute the primary food for females. Dragon Spruce seeds, invertebrates (mainly ants) and forbs on the ground are frequently consumed by females, but seldom by males, possibly to meet the nutrient constraints of egg formation. This sex-specific diet in this monogamous species might result from the fact that females allocate more time searching for scarce, nutritious food, whereas males spend more time being vigilant (Wang et al., 2010a).
Statistics from 73 radio-marked Chinese Grouses during 1995–2000 show that the annual survival rate of the bird is 0.636 for adults and 0.174 for chicks from birth to 13 weeks. The rate of reproduction is 1.81 young females per female in the spring of each year. Chick survival is generally low and varies greatly among years. Although we found no difference in adult survival between both sexes, factors associated with habitat fragmentation may have negative effects on chick survival and contribute to the skewed adult sex ratio (Sun et al., 2003).
The habitat (2600–3600 m elevations) of the Chinese Grouse in the Lianhuashan Reserve is characterized as a mixture of conifers which provide cover and deciduous trees as food resources (willow and birch). Logging of the virgin forest in this area in the past 30 to 40 years has caused serious loss of the coniferous forests. Using digitalized SPOT satellite images, we identified the distribution of coniferous forests, the key habitat of the Chinese Grouse. The population of the Chinese Grouse at the reserve is an isolated one and is now split into two sub-populations. Within a study area of 120000 ha, we found that 77.3% of forest patches were smaller than 10 ha, indicating a high level of habitat fragmentation. From a survey of 31 forest patches where Chinese Grouse were present, we found that forest patches occupied by this bird were much larger (102 vs. 8.6 ha) and closer to occupied ones (583 m vs. 2325 m), compared to empty patches. Suggestions for conservation have been proposed to local governments to protect the entire habitat of this isolated population of Chinese Grouse (Klaus et al., 2001; Sun et al., 2006).
At Lianhuashan, local people found an estimated 10–29% of the nests, from which they took eggs as food (Sun et al., 2003). According to a population viability analysis (PVA), the predicted population trend at Lianhuashan is most sensitive to chick mortality, offspring per female per year and male adult mortality. Among these three variables, the first two are markedly correlated with human activity, such as egg collection by local people. If chick mortality declines by 5%, or the number of offspring produced per female increases by 5% (i.e. reduced nest loss) under current conditions, the size of the local reserve and its current population would form into a minimum viable population (MVP) of 2500 birds. Therefore, the most practical and simple conservation management tool would be to assure the increase in breeding success of the Chinese Grouse, especially limiting the negative effect of anthropological activities during the incubation period (Lu and Sun, 2010).
Finally, we suggest that future research be concentrated on habitat requirements and the impacts of habitat fragmentation and isolation in order to outline a protocol to guide ongoing reforestation in habitat areas of the Chinese Grouse.
The work on the Chinese Grouse at Lianhuashan Nature Reserve has been continuously supported by the National Natural Science Foundation of China (Grants 39400016, 30370223, 30620130110, 31071931) since 1995. Our study has also been supported by the Chinese Academy of Sciences, the Deutsche Forschungs- Gemeinschaft (DFG), the BP Conservation Programme, the World Pheasant Association, the Wildlife Conservation Society and the Martin Wills Trust. We also thank people in the Lianhuashan Nature Reserve for their great help in our work.
|
Cheng L, Li XM. 2006. The status and conservation of Cranes in Heilongjiang Province. Territ Nat Resour Study, 1: 90-91. (in Chinese with English abstract)
|
|
Feng KM, Li JL. 1985. Aerial surveys of the Red-crowned Cranes and other waterflowls in China. J Northeast Forest Univ, 13(1): 80-87. (in Chinese with English abstract)
|
|
Fujita G, Harris J, Bold A, Tseveenmayadag N, Chuluunbatar S. 1994. Habitat preference of Demoiselle and White-napped Cranes, Anthropoides virgo and Grus vipio, breeding in Mongolia. In: Higuchi H, Minton J (eds) The Future of Cranes and Wetlands. Wild Bird Society of Japan, Tokyo, Japan, pp 93-96.
|
|
Han YL, Ge DN, Zhang YX. 2003. The research about birds resource and conservation in Sanjiang Plain. Territ Nat Resour Study, 1: 76. (in Chinese with English abstract)
|
|
Harris J. 1994. Cranes, people and nature: preserving the balance. In: Higuchi H, Minton J (eds) The Future of Cranes and Wetlands. Wild Bird Society of Japan, Tokyo, Japan, pp 1-14.
|
|
Hilton-Taylor C. 2000. IUCN Red List of Threatened Species. Gland, Switzerland and Cambridge, UK: IUCN.
|
|
Hua Y, Li XM, Liu XC, Sun ZY. 2006. Comparison of bird diversity during spring and autumn in Honghe Nature Reserve. J Northeast Forest Univ, 34(3): 23-28. (in Chinese with English abstract)
|
|
Jiang HX, Liu CY, Qian FW, Li CY, Qiu FC. 2009. A model of nest-site selection of Red-crowned crane based on RS, GIS and GPS techniques at Zhalong wetland, China. Sci Silv Sin, 45(7): 76-83. (in Chinese with English abstract)
|
|
Lee SD, Jablonski PD, Higuchi H. 2007. Winter foraging of threatened cranes in the Demilitarized Zone of Korea: behavioral evidence for the conservation importance of unplowed rice fields. Biol Conserv, 139(1-2): 286-289.
|
|
Li FM, Li PX. 1998. A comparative study on territories of White-napped Crane and Red-crowned Crane. Acta Zool Sin, 44(1): 109-111. (in Chinese with English abstract)
|
|
Li FM, Li PX. 1999. A comparative study on territories of White-napped Crane and Red-crowned Crane. China J Ecol, 18(6): 33-37. (in Chinese with English abstract)
|
|
Li XM, Hu YM, Ma YJ, Jia JL. 2003. Status and conservation of cranes in Sanjiang Plain. Territ Nat Resour Study, 1: 74-75. (in Chinese with English abstract)
|
|
Liu HY, Li ZF. 2008. Effects of landscape change of wetlands on habitats of waterfowls within Honghe Nature Reserve by its surrounding area. Acta Ecol Sin, 28(10): 5011-5019. (in Chinese with English abstract)
|
|
Liu HY, Zhang SK, Li ZF, Lv XG, Yang Q. 2004. Impacts on wetlands of large-scale land-use changes by agricultural development: The small Sanjiang Plain, China. Ambio, 33(6): 284-288. (in Chinese with English abstract)
|
|
Liu XT. 1997. Marsh resource and its sustainable utility in the Songnen-Sanjiang Plain. Sci Geogr Sin, 16(Suppl. ): 451-460. (in Chinese with English abstract)
|
|
Liu XT. 2000. Influence of large-scale reclamation on natural environment and regional environmental protection in the Sanjiang Plain. Sci Geogr Sin, 20: 14-19. (in Chinese with English abstract)
|
|
Liu XT, Ma XH. 2002. Natural Environmental Changes and Ecological Protection in the Sanjiang Plain. Science Press, Beijing. (in Chinese)
|
|
Ma YQ, Jin LR. 1987. The numerical distribution of the Red-crowned Crane in Sanjiang Plain area of Heilongjiang Province. Acta Zool Sin, 33(1): 82-87. (in Chinese with English abstract)
|
|
Masatomi H, Ogawa S, Miyama K. 1990. The relations between land cover monitored by satellite remote sensing and distribution of the Tancho's nests in Kushiro Marsh, Hokkaido. J Hokkaido Coll, Sensu Univ, 23: 209-223.
|
|
Pang SL. 2000. The report on the aerial survey of Red-crowned Crane in the Zhalong Nature Reserve. J Sci Teach Coll Univ, 20(2): 59-60. (in Chinese)
|
|
Piao RZ, Han AH, Zhang MH. 2000. Remote sensing and geographic information system to estimate habitat selection of red-crowned crane in Sanjiang Plain of Northeastern China. In: China Ornithology Association, Wild Bird Society of Taipei and China Wildlife Conservation Association (eds) Studies on Chinese Ornithology — Proceedings of the 4th Ornithological Symposium of Mainland and Taiwan, China. China Forestry Publishing House, Beijing, pp 170-197. (in Chinese with English abstract)
|
|
Prugh LR, Hodges KE, Sinclair ARE, Brashares JS. 2008. Effect of habitat area and isolation on fragmented animal populations. PNAS, 105(52): 20770-20775.
|
|
Rodgers JA, Kubilis PS, Nesbitt SA. 2005. Accuracy of aerial surveys of waterbird colonies. Waterbirds, 28(2): 230-237.
|
|
Schmidt BD, Roland J. 2006. Moth diversity in a fragmented habitat: importance of functional groups and landscape scale in the boreal forest. Ecol Popul Biol, 99: 1110-1120.
|
|
Smirenski SM. 1988. Chick relationships and brood sizes in Red-crowned (Grus japonensis) and White-napped (Grus vipio) Cranes. In: Litvinenko NM, Neufeldt IA (eds). The Palearctic Cranes. Amur-Ussuri Branch of the USSE Ornithological Society, Vladivostok. USSR, pp 49-53.
|
|
Smirenski SM, Harris J, Gray C, Strom K, Ichida N. 1995. Report of the Amur program of the Socio-Ecological Union. In: Halvorson CH, Harris JT, Smirenski SM (eds) Cranes and storks of the Amur River: the Proceedings of the International Workshop. Arts Literature Publishers, Moscow, Russia, pp 205-212.
|
|
Song KS, Liu DW, Wang ZM, Zhang B, Jin C, Li F, Liu HJ. 2008. Land use change in Sanjiang Plain and its driving forces analysis since 1954. Acta Geogr Sin, 63(1): 94-104. (in Chinese with English abstract)
|
|
Su LY, Xu J, Zhou DS. 1991. Breeding habitats of White-napped Cranes at Zhalong Nature Reserve. In: Harris J (eds) Prodeedings of 1987 International Crane Workshop. International Crane Foundation, Baraboo, Wisconsin, USA, pp 51-57. (in Chinese)
|
|
Su LY, Wang QS. 2010. The mainland population of the red-crowned crane faces growing threat. China Crane News, 14(1): 3-8.
|
|
Swift TL, Hannon SJ. 2010. Critical thresholds associated with habitat loss: a review of the concepts, evidence, and applications. Biol Rev, 85: 35-53.
|
|
Temple SA. 1986. The problem of avian extinctions. Curr Ornithol, 3: 453-485.
|
|
Wang QS. 2008. Threats for Red-crowned Crane. China Crane News, 12(2): 7-12.
|
|
Yang XJ, Zhang SM, Zhang XX, Feng WY. 2005. The composing and analysis of large waterbirds diversity in the Sanjiang Plain. J Northeast For Univ, 33(3): 56-58. (in Chinese with English abstract)
|
|
Zhang GC, Deng W, Lu XG, Song KS, Li HD, Zhang HY. 2007. The dynamic change of wetland landscape patterns in Xinkai River Basin. J Nat Resour, 22(2): 204-210. (in Chinese with English abstract)
|
|
Zhang SQ, Na XD, Kong B, Wang ZM, Jiang HX, Yu H, Zhao ZC, Li XF, Liu CY, Patricia D. 2009. Identifying wetland change in China's Sanjiang Plain Using Remote Sensing. Wetlands, 29(1): 302-313.
|
|
Zhao KY (ed). 1999. Chinese Mire Records. Science Press, Beijing. (in Chinese)
|
|
Zhou DS, Su LY. 1990. Preliminary study on the territory behavior of Red-crowned Crane. In: Heilongjiang Forestry Department (ed) Conservation and Research of International Cranes. China Forestry Publishing House, Beijing, pp 54-58. (in Chinese)
|
|
Zou HF, Wu QM, Ma JZ. 2003. The nest-site selection of Red-crowned crane in Zhalong Nature Reserve after burning and irrigating. J Northeast Norm Univ, 35(1): 54-59. (in Chinese with English abstract)
|
Figures(4) / Tables(6)





| Month (Illustrations) | Male | Female |
| April (Flock dissolution and pair bond formation) | Hold territory by flutter-jump and flutter-fly displays (adults leave the flock and return to their territories earlier) | Pair with males with better territory quality (adults leave the winter flock area and return to their breeding area, all females breed) |
| May (Living in pairs) | Hold territory and guard their mates, and juveniles normally have no chance to breed | Feed to gain nutrition in order to lay clutches of 5–8 eggs |
| June (Incubation) | Stay around nests | Incubation, hatching around the end of the month |
| July–August (Brood time) | Stay in or around territory, recovering from heavy investment | Lead the brood leaving the nest area, raising the chicks themselves |
| September (Brood dissolution) | Stay in or around territory | Brood dissolution, chick dispersal for 1–5 km |
| October (Autumn territoriality) | Defend their territories | Wander in territories of males, join small groups of 3–4 birds |
| November-March (Winter flock) | Adopt three strategies: stay in the territory, move between flock area and territory or stay in the flock (juveniles all in flock) | Move back to their winter flock area, most remain in flock with maximum size of 12–13 birds |
DownLoad:
CSV
 Email Alerts
Email Alerts RSS Feeds
RSS Feeds