Iftikhar HUSSAIN, Asmat-un-NISA, Sangum KHALIL. 2012: Population biology of the Grey Francolin (Francolinus pondicerianus) in an agro-ecosystem of the Pothwar Plateau, Pakistan. Avian Research, 3(2): 91-102. DOI: 10.5122/cbirds.2012.0009
Citation:
Iftikhar HUSSAIN, Asmat-un-NISA, Sangum KHALIL. 2012: Population biology of the Grey Francolin (Francolinus pondicerianus) in an agro-ecosystem of the Pothwar Plateau, Pakistan. Avian Research, 3(2): 91-102. DOI: 10.5122/cbirds.2012.0009
Iftikhar HUSSAIN, Asmat-un-NISA, Sangum KHALIL. 2012: Population biology of the Grey Francolin (Francolinus pondicerianus) in an agro-ecosystem of the Pothwar Plateau, Pakistan. Avian Research, 3(2): 91-102. DOI: 10.5122/cbirds.2012.0009
Citation:
Iftikhar HUSSAIN, Asmat-un-NISA, Sangum KHALIL. 2012: Population biology of the Grey Francolin (Francolinus pondicerianus) in an agro-ecosystem of the Pothwar Plateau, Pakistan. Avian Research, 3(2): 91-102. DOI: 10.5122/cbirds.2012.0009
The field biology of the Grey Francolin (Francolinus pondicerianus) was studied in 2009 in an agro-ecosystem of the Pothwar area, an important range of its distribution in Pakistan. The bird density was estimated as 1.59±0.39 (mean±standard error) birds per ha in crop fields and 0.87±0.14 birds per ha in forest tracks, with some seasonal fluctuations in its habitat. All bird nests were located on the ground in vegetation mainly comprised of Desmostachia bipinnata, Acacia modesta, Imperata cylindrical, Zizipus jujuba and Euphorbia spp. Egg laying was recorded during the spring and summer months, with a mean egg laying span of 6±0.36 days (range 5-7 days) and a mean clutch size of 7±0.36 (range of 6-8 eggs). The mean incubation period was 20.6±0.50 days (range 19-22 days). Out of a total number of 42 eggs recorded, 32 hatched (76.19% success) with a mean hatching rate of 5.33±1.22 eggs per clutch. The fledgling success was estimated as 3.83±0.83 per clutch (63.08%). The species was found to be omnivorous and euryphagus in its feeding habit. There were 10 identifiable food items in the diet of the species, consisting of seven plant species (Pennisetum typhoideum, Sorghum bicolor, S. halepense, Prosopis juliflora, Phaseolus radiates, Carthemus axycantha and Acacia sp.), two insect species (Coptotermes formosanus and Formica rufa) and one grit.
In Pakistan, the Grey Francolin (Francolinus pondicerianus) is widely associated with the drier regions of the Indus plains and has penetrated the Thar desert in Sindh, as well as the Thal and Chohlistan deserts in Punjab. The species also occurs in the lower hills of the Makran and Lasbela districts in Balochistan, the Cherat and Kohat districts of Khyber Pakhtunkhwa Province, the salt range and agro-forestry tracks of the Pothwar Plateau in the Punjab and in the Margalla hills of Islamabad (Roberts, 1991; Mahmood et al., 1997; Mann and Chaudhry, 2000). The bird is normally found foraging in open cultivated tracks and grasslands intermixed with scrub forests and is rarely observed above an elevation of 1200 m in Pakistan. The calls of the Grey Francolin, uttered mostly at dawn and at dusk, are considered one of the most familiar bird songs of countryside (Roberts, 1991). In the Indian subcontinent, the Grey Francolin has remained a favorite game bird and continues to be hunted for food since it is considered a cheap source of meat to local inhabitants (Long, 1981). This bird is also used for cockfighting at village fairs (Javed et al., 1999). The species is a friend of farmers who believe this bird consumes large numbers of harmful insects, as well as their eggs and larvae and serves therefore as a biological control agent of insect pests in Pakistan (Beg and Qureshi, 1972; Qureshi, 1972; Mian, 1995).
Although the bird is intensively hunted and captured in its native range in Pakistan, owing to which local populations could be declining, but the overall status of the species is regarded as stable (del Hoyo et al., 1994; Islam, 1999; Richard et al., 2002). Therefore, it is not believed that its populations are approaching the threshold criteria set for population decline in the IUCN Red List (i.e. declining more than 30% in ten years or in three generations) and thus the conservation status of this species has been rated as of Least Concern (IUCN, 2011). However, the Grey Francolin has a protected status in Punjab Province and falls in Schedule-I of the Punjab Wildlife Acts and Rules 1974. This law permits restricted hunting of the species during the non-breeding winter months, i.e. from 15 November to 15 January, with a limit to hunting not more than six birds per day per person (Shafiq, 2005). Such legal instruments and general public perception regarding current populations of the Grey Francolin have led us to believe that the species is under pressure, presumably due to illegal hunting, urbanization, expansion and intensification of agricultural practices and other pressures on scrub forests by local populations for their use in fodder, timber and fire wood needs.
Rather than providing a complete picture of the field biology of this species in any particular habitat/area, most of the existing country level studies have reported different and exclusive life history traits (Faruqui et al., 1960; Ullah, 1991; Chaudhry and Bhatti, 1992; Mian and Wajid, 1994; Mahmood et al., 1997; Kamal, 2000; Mann and Chaudhry, 2000). However, a recent investigation by Khan (2010) has covered breeding and dietary aspects of a population inhabiting a protected area of desert ecology, about 500 km towards the southeast of the Pothwar Plateau. The Pothwar Plateau is one of the major distribution ranges of the Grey Francolin (Roberts, 1991). The plateau is characterized by broken terrain, with a total area of 1.82 million ha, out of which only 0.61 million ha is under cultivation and the remainder is occupied by scrub forests (Ahmad, 1990). The available literature provides limited information on the biology of the species in the Pothwar range of its distribution. Hence, in 2009, we explored some factors related to population, habitat use, diet composition and breeding potential of this elegant galliform in a representative site of the agro-ecosystem of the Pothwar Plateau.
Methods
Population and breeding studies were conducted in the area of Jhungal village (33°26.754 N, 73°07.110 E), in the Gujar Khan district of Rawalpindi. The land utilization of Gujar Khan consists for 46% of cultivation and 26% is covered by natural vegetation and forest tracks. Its main crops are wheat, maize, millet, mustard and groundnut (Helders, 2007). The annual rain fall of the area ranges between 500 and 1000 mm. Major tree species, composed of wild flora, are Phulai (Acacia modesta), Kikar (Acacia nilotica), Toon (Cedrala toona), Dharek (Melia azederach), Ber (Zizyphus mausitiana) and Shisham (Dalbergia sissoo). A conspicuous species of grass called Dub (Desmostachia bipinnata) has been reported in abundance (Nizami et al., 2004). The landscape of the study area is represented by the typical characteristics of Pothwar's agro-ecology, consisting of undulated open land used for rain-fed agriculture. These tracks are inter-dispersed with gullies and low hills/ranges covered by dry sub-tropical forests. Generally, winter cultivation (November to April) consists of wheat with intercropping of grams (chickpeas), lentils and mustards whereas the summer (May to October) crops include sorghum, millet and groundnuts or peanuts (Beg et al., 1985; Ahmad, 1990).
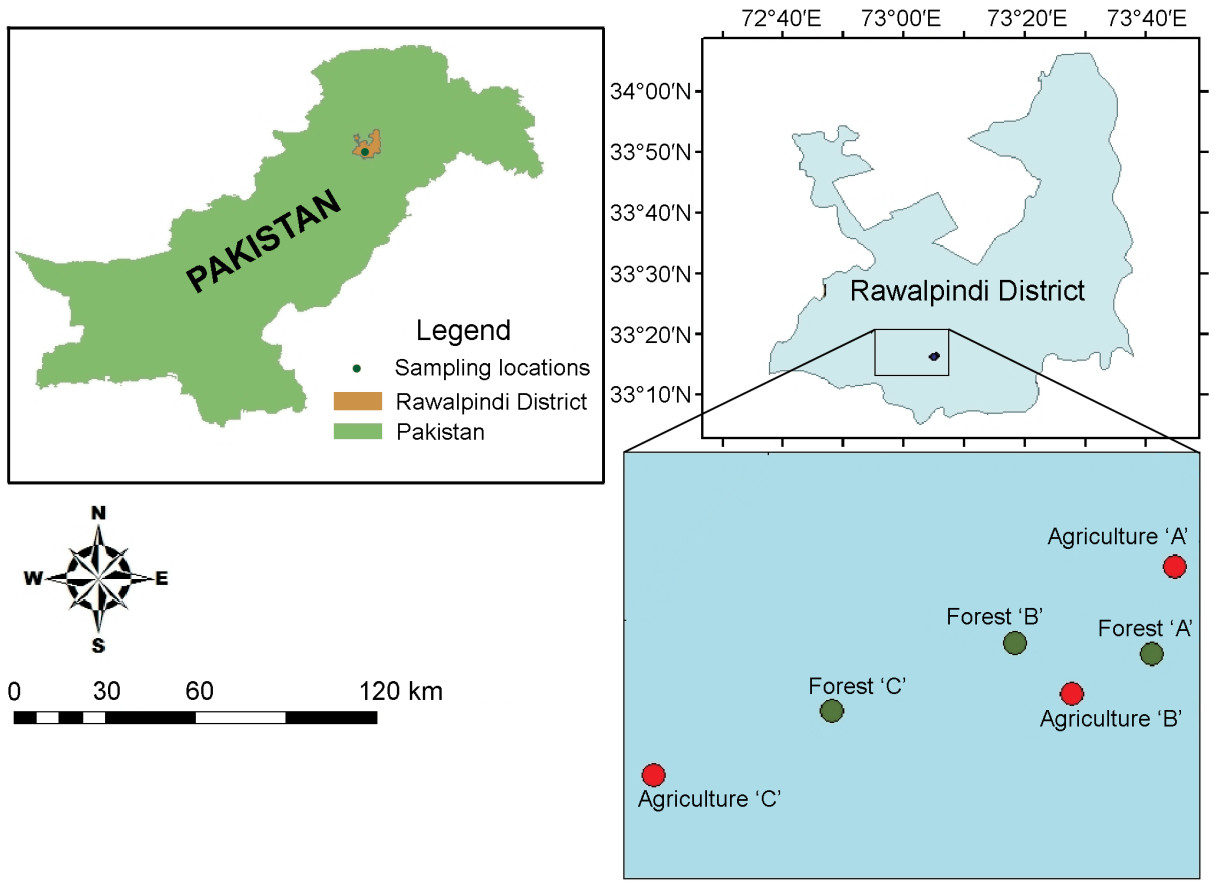
Estimation of the size of the Grey Francolin population was carried out using a line-transect method (Burnham et al., 1980). The study area (9 km2) included two representative sampling sites, i.e., cropland and scrub forest habitats. In both habitats three isolated permanent line transects, with a fixed width and length (500 m × 50 m), were established to monitor the direct and indirect evidence of the presence of the francolin species (Fig. 1). For direct sighting in each of the sampling sites, the observer walked at a moderate speed (2–3 km·hour–1) taking normal paces to record birds within the transect width. The observations were recorded once a month in the morning, the peak activity hours of the bird (Gould, 1966; Mahmood et al., 2010). Only direct sighting data were used for population assessment. Density (number of birds per ha) was calculated by dividing the number of sighted birds by the transect area (2.5 ha = 500 m × 50 m, length × width of the transect). Indirect evidence such as nests, droppings, foot prints, feathers and vocal calls were used to search the bird nests for an in-situ breeding study and to identify its roosting and feeding areas. Similarly, the vegetation associated with nesting sites were also observed and identified.
Figure
1.
Maps of study site and transect lines established for monitoring Grey Francolin (Francolinus pondicerianus)populations in agro-ecosystems in the Rawalpindi district, Pakistan
Assuming equal potential of both habitats for hosting and breeding activities of the Grey Francolin, investigations of the study location for nests were made during March, April and May. This period coincides with the main breeding season of this bird as reported by Roberts (1991) for March and April and by Khan (2010) for March through May. Their habitat were regularly visited and observed for any signs of nest construction before the start of the expected breeding season i.e., the first week of March 2009. Local hunters assisted in locating bird nests. Search of these nests was carried out by walking on existing pathways across a forest plantation, while randomly encountered female birds were followed to reach their nests. After confirming an active nest, it was marked and allotted a specific number. Nest location, general appearance and its structure were noted. Marked nests were visited twice a week with maximum precaution to minimize disturbance to the nesting birds. The frequency of observations/visits was increased (on alternate days) after egg laying in order to record incubation periods and time of hatching. The information recorded include dates of laying first and last eggs, the number of eggs, the date of hatching and the number of hatched eggs.
In order to determine diet composition of the Grey Francolin, 13 (10 males and 3 females) freshly hunted birds were procured from local hunters from the Jhelum and Rawalpindi districts of the Pothwar Plateau. Simultaneously, surveys of the habitat of the hunted birds were carried out to collect reference vegetation, e.g. grasses, shrubs, trees and farm crops, for use to identify the gut contents. The crops and gizzards were dissected to observe their contents in situ and subsequently the contents were segregated into identifiable components. Seeds and plant parts were identified from the Weed Management Programme of the National Agricultural Research Center (NARC), Islamabad, while the insect samples were identified with technical assistance from the Biosystematics Laboratory, Department of Entomology, PMAS Arid Agriculture University Rawalpindi and the National Insect Museum, NARC, Islamabad. All the constituents of the crops and gizzards contents were quantified by measuring their dry weights.
The transect data recorded during this nine-month study period are grouped into climatically distinctive seasons of the area, i.e., winter (mid-November through February), spring (March through April), summer including the monsoon season (May through mid-September) and autumn (mid-September through mid-November). Mean values mentioned in the text are given with their standard errors (SE). We used two-sample t-tests or one-way analyses of variance by employing Minitab-16 Statistical Software for analysis of differences between any two data sets.
Results
Population
Records of bird sighting data from the line-transects revealed a mean perpendicular distance of birds from the observer in the cropland habitat of 5.7 ± 0.5 m (n = 34, range = 0.6–9.4 m) and in the forest habitat of 6.3 ± 0.5 m (n = 23, range = 0.6–9.1 m). Non-significant differences between these two data sets (t = 0.85, df = 55, p = 0.398) suggest that we can assume that the birds were equally detectable in both habitats. The observed flock sizes (n) with frequencies of their occurrence (given in parenthesis) are as follows: 1(20), 2(20), 3(7), 4(1), 5(5), 6(1), 7(2) and 9(1).
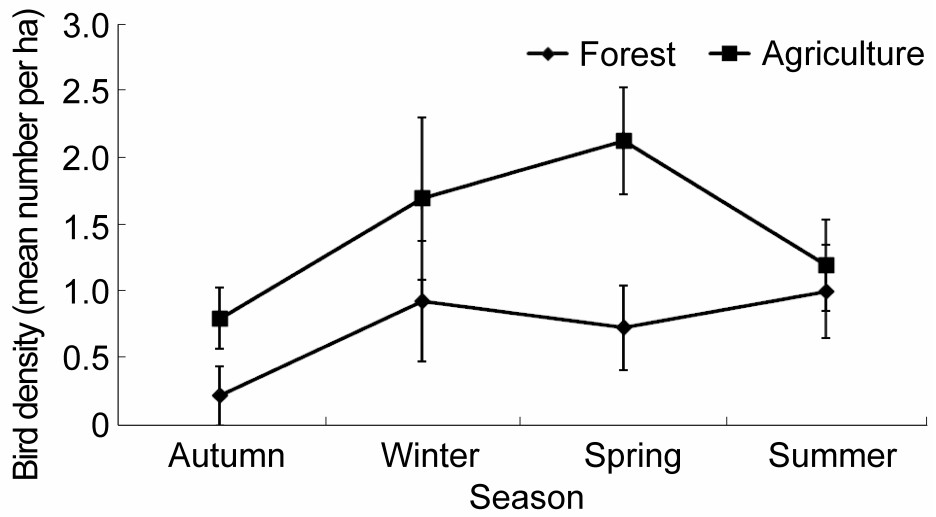
The monthly/seasonal data of the Grey Francolin recorded by direct sightings from all the six transects (three each in the forest and agriculture habitats) are detailed in Tables 1 and 2. The density was estimated as 1.59 ± 0.39 (mean ± standard error) birds per ha in the cropland habitat and 0.87 ± 0.14 birds per ha in the forest track. Although a higher density was evident in the agricultural transects compared to the forest area, a comparison of the overall number of birds seen in these two habitats showed non-significant difference at the 0.05 level of significance (t = 1.82, df = 16, p = 0.098). Season-wise segregation of data indicated the highest population estimates in the cropland areas during the spring season, whereas in the forest habitat the population was (more or less) equally high during the winter and summer months. However, there were minimum sightings during the autumn season in both habitats (Fig. 2).
Table
1.
Record of population densities of the Grey Francolin (Francolinus pondicerianus)in cropland habitats of the Gujar Khan area in the Rawalpindi district, Pakistan. Each transect has a length of 500 m and a width of 50 m with an area of 2.5 ha.
Table
2.
Record of population densities of the Grey Francolin (Francolinus pondicerianus)in scrub forest habitats of the Gujar Khan area in the Rawalpindi district, Pakistan. Each transect has a length of 500 m and a width of 50 m with an area of 2.5 ha.
Season
Month
Transect A
Transect B
Transect C
Mean density(n per ha)(Mean ± SE)
n
Crop and vegetation
n
Crop and vegetation
n
Crop and vegetation
Monthly
Seasonal
Autumn
Oct
–
Acacia modesta (dominant), A. nilotica (rare)
–
Acacia modesta (dominant), Zizyphus mausitiana, Typha angustata (common), Dalbergia sissoo (rare), Desmostachia bipinnata (scattered in patches) along a small wetland
5
Acacia modesta (dominant), Dalbergia sissoo, A. nilotica, Zizyphus mausitiana and Desmostachiabipinnata (common)
Figure
2.
Seasonal variation in population densities of the Grey Francolin (Francolinus pondicerianus)in cropland and forest tracks of agro-ecosystem of the Pothwar Plateau, Pakistan
We assume that the variable distribution of birds within the agriculture habitat (Table 1) is steered by the prevalence of cultivation. The agricultural fields occurring in transect A were recorded as uncultivated during most of the study period, except in October and June, whereas the crop fields of transects B and C remained under seasonal crops except during the periods of land preparation for sowing the next crop. Transect-wise densities of the Grey Francolin in these agricultural areas were recorded at 0.71 ± 0.15, 2.00 ± 0.63 and 2.04 ± 0.54 birds per ha in transects A, B and C, respectively. The breakup of data by season showed estimates in the following descending order: 2.13 ± 0.40 birds per ha during spring, 1.70 ± 0.61 in winter, followed by 1.20 ± 0.34 in the summer and the lowest estimates, i.e. 0.80 ± 0.23 birds per ha during the autumn season (Table 1).
The record of population densities of the Grey Francolin in scrub forest tracks are presented in Table 2. The estimates of bird populations in the first two transects (A and B) were 0.36 ± 0.23 and 0.31 ± 0.21 birds per ha, respectively, whereas the third transect, C, had the maximum population i.e. 1.96 ± 0.44 birds per ha. The vegetation of transect A was that of a monoculture, i.e., Acacia modesta with a rare occurrence of A. nilotica. The flora of transects B and C had an almost similar inventory of plant species, except for the presence of a small wetland surrounded by Typha angustata along transect B. However, there were higher densities of Desmstachia bipinnata and Dalbergia sissoo on transect C where the highest population of Grey Francolins was recorded (Table 2). Although these plant species do not contribute to the food supply of the Grey Francolin (Table 5), the relationship can presumably be attributed to meet the roosting and sheltering needs of the species. The seasonal estimates of the Grey Francolin in the scrub forest habitat revealed occurrences of the birds at 0.22 ± 0.22, 0.93 ± 0.45, 0.73 ± 0.32 and 1.0 ± 0.35 birds per ha during the autumn, winter, spring and summer seasons, respectively.
Breeding
In total, six active nests were found, four in forest tracks A, B, C and D and two in wild vegetation on the boundaries of cultivated fields (E and F). All the nests were situated on the ground in vegetation mainly comprised of Desmostachia bipinnata, Acacia modesta, Imperata cylindrical, Zizipus jujuba and Euphorbia spp. (Table 3, Fig. 3). The mean egg laying period was estimated at 6 ± 0.36 days (range 5–7 days), which corresponds to the recorded average clutch size of 7 ± 0.36 (range of 6–8 eggs), indicating that this population was probably laying eggs daily.
Table
3.
Location and nest material of the Grey Francolin (Francolinus pondicerianus)found in an agro-ecosystem of the Pothwar Plateau, Pakistan
The average incubation period recorded in the present study was of 20.6 ± 0.50 days (range 19–22 days). Out of a total of 42 eggs recorded, 32 hatched (75.39% success) with a mean number of 5.33 ± 1.22 eggs hatched per clutch. The mean number of fledglings was estimated as 3.83 ± 0.83 per clutch, indicating an overall success of 63.08% (Table 4).
Table
4.
Breeding performance of the Grey Francolin (Francolinus pondicerianus)in an agro-ecosystem of the Pothwar Plateau, Pakistan
The results of the gut content analysis suggest it an omnivorous and euryphagus species, with a limited range in food selection (Table 5). The overall diet of this population consisted of ten identifiable food items, consisting of seven plant species, two insects and one grit. However, not more than five identifiable food items were found in any of the individual gut samples. The main components of the diet were seeds of cultivated crops, wild vegetation and arthropods. In total, 42 incidences of occurrence of different identifiable food contents were recorded. Among these were 14 records (33.3%) of cultivated field crops, 10 records (23.8%) of wild vegetation, 7 records (16.7%) of insects and 11 records (26.2%) of grits. Proportions of different food items (by weight) were estimated as 45.6% plant seeds, 12.4% plant leaves, 20.5% insects, 15.7% grits and 5.7% of unidentified material. The proportion of food contents derived from cultivated crops and wild vegetation was found to be in the ratio of 61:39.
Table
5.
Composition of different food items recovered from crops and gizzards of the Grey Francolin (Francolinus pondicerianus) inhabiting an agro-ecosystem of the Pothwar Plateau, Pakistan
Sr. No.
Timeof hunt
Month (Season)a
Habitat
Body wt. (g)
Dry wt. of food contents (g)
Wt. of food contents per 100 g body wt.
Millet (seeds)
Sorghum(seeds)
Vilayati Kikar (leaves)
Mung Beans(seeds)
Acaciasp. (seeds)
Baru (seeds)
Carthemus oxycantha(seeds)
Termites
Black Ant
Grit
Unidentified
1
Evening
Nov (W)
Cropland
250
7.0
2.80
2.5
–
2.5
–
–
–
–
–
–
2.0
–
2
Evening
Dec (W)
Forest
263
12.0
4.56
1.5
2.5
–
2.0
–
2.5
–
–
–
3.0
0.5
3
Evening
Dec (W)
Cropland
210
9.0
4.29
1.7
–
–
3.2
–
–
–
1.3
1.0
1.5
0.3
4
Evening
Dec (W)
Cropland
242
6.0
2.48
–
2.5
–
1.5
–
–
–
–
1.0
0.8
0.2
5
Morning
Jan (W)
Forest
200
3.0
1.50
–
–
–
–
–
–
1.7
–
–
0.9
0.4
6
Evening
Jan (W)
Cropland
267
13.0
4.87
–
5.0
5.5
1.0
–
–
–
–
–
1.0
0.5
7
Evening
Feb (W)
Cropland
247
13.0
5.26
–
5.5
2.0
–
–
–
–
1.2
4.0
–
0.3
8
Evening
May (S)
Cropland
118
1.0
0.85
0.6
–
0.25
–
–
–
–
–
–
–
0.15
9
Morning
May (S)
Cropland
168
3.0
1.79
–
–
0.55
0.5
–
–
–
–
–
1.0
0.95
10
Morning
May (S)
Cropland
155
2.0
1.29
0.8
–
–
–
–
–
–
–
–
0.5
0.7
11
Evening
July (S)
Forest
257
8.5
3.31
–
–
–
–
1.5
–
–
5.5
–
1.0
0.5
12
Evening
July (S)
Forest
240
7.5
3.13
–
–
–
–
2.0
–
–
3.8
–
1.5
0.2
13
Morning
July (S)
Forest
235
2.0
0.85
–
–
–
–
1.2
–
–
–
–
0.5
0.3
Frequency of occurrence of identifiable food items n (%)
–
–
–
5 (11.9)
4 (9.5)
5 (11.9)
5 (11.9)
3 (7.1)
1 (2.4)
1 (2.4)
4 (9.5)
3 (7.1)
11 (26.2)
12
Mean ± SE
219.4 ± 12.9
6.7 ± 1.2
2.8 ± 0.4
1.4 ± 0.3
3.9 ± 0.8
2.2 ± 0.9
1.6 ± 0.5
1.6 ± 0.2
2.5 ± 0.0
1.7 ± 0.0
3.0 ± 1.2
2.0 ± 1.0
1.2 ± 0.2
0.4 ± 0.07
Total weight (% proportion) b
–
–
–
7.1(8.2%)
15.5(17.8%)
10.8(12.4%)
8.2(9.4%)
4.7(5.4%)
2.5(2.9%)
1.7(2.0%)
11.8(13.6%)
6.0(6.9%)
13.7(15.7%)
5.0(5.7%)
a W = winter; S = summer. b Totally, seeds = 45.6%; leaves = 12.4%; insects = 20.5%. Latin names of some of food items: Millet, Pennisetum typhoideum; Sorghum, Sorghum bicolor; Vilayati Kikar, Prosopis juliflora; Mung Beans, Phaseolus radiatus; Baru, Sorghum halepense; Termites, Coptotermes formosanus; Black Ant, Formica rufa.
Adjusted mean weight (g per 100 g body weight) of food contents gleaned from crops and gizzards of hunted birds in the morning (1.36 ± 0.2, n = 4) was significantly less (F1, 12 = 8.83, p = 0.013) than that of birds hunted in the evening (3.51 ± 0.46, n = 9). The increased weight of food content in birds shot in the evening suggests that the birds fill their guts before sunset in order to meet the energy requirements during the non-feeding overnight period.
Weights of the gut contents (g per 100 g body weight) of the birds collected from cropland and forest habitats, respectively, were recorded as 2.95 ± 0.59 g (n = 8) and 2.67 g ± 0.67 (n = 5). The difference between the two habitats was not significant (F1, 12 = 0.10, p = 0.76). These figures do not indicate any quantitative difference in the availability of food sources between both habitats.
Discussion
The pattern of habitat preference recorded for the Grey Francolin in the present study area is supported by a similar study conducted in a scrub habitat of the Pothwar area (Mahmood et al., 2010), suggesting that this species selects habitats dominated by trees and shrubs. The asymmetrical prevalence of the Grey Francolin in the study transects in both habitats (cropland and scrub forest) could be attributed to variable vegetation/crops providing food and shelter to the birds. The data from transect C in the forest habitat show that large numbers of birds in vegetation dominated by Acacia modesta, Dalbergia sissoo and D. bipinnata. Wijeyamohan et al. (2003) in Sri Lanka reported that low bushes and thorn scrubs, especially Acacia eburnean, provided the most favorite habitats of the Grey Francolin. The activity of the bird was lower in the area (transect B in the forest) along a small area of wetland, although it had a good diversity of natural vegetation containing Acacia modesta (dominant), Zizyphus mausitiana, Typha angustata (common), Dalbergia sissoo (rare) and Desmostachia bipinnata (scattered in patches). Some earlier reports have also shown that these birds do not prefer habitats/vegetation close to water sources and that they meet their water requirement from succulent vegetation, dew and/or insects (Yeatter, 1934; Mc Crow, 1982; Ullah, 1991; Mian and Wajid, 1994; Kamal, 2000).
The relatively higher density estimated in the cropland habitat (2.13 ± 0.40 birds per ha) during the spring season, corresponding with the maturity of wheat and related crops, is supported by the report of Mian and Ghani (2007) that population density of the Grey Francolin increases with an increase in vegetation cover. The higher estimates in the cropland is also supported by Roberts (1991) who stated that the bird restricts its movement to areas rich in crop grains, weed seeds, succulent vegetation and grits. Relatively higher populations in the forest habitat during summer and winter months could be attributed to the availability of cover for protection from extreme weather conditions. However, populations of this species in the larger scrub-forests on the sub-mountainous tracks of Kheri Murat, Fateh Jang and Attock districts (Mann and Choudhry, 2000) and the Lehri Wildlife Park, in the Jhelum district of the Pothwar Plateau (Mahmood et al., 2010), were estimated at 0.05 and 0.47 bird per ha, respectively. The current population is higher than those reported (Kamal, 2000) in the different agro-ecological sub-habitats of the Mardan district in Khyber Pakhtunkhwa Province, Pakistan (i.e., 0.98 bird per ha in the scrub forest, 0.28 in an irrigated forest plantation, 1.3 in a cropland adjoining a scrub forest, 0.25 in a cropland, 0.1 in a cropland adjoining a wetland and 0.48 in an alkaline waste/cropland). It is also higher than that in a tropical thorn forest and irrigated forest of a protected area (Lal Suhanra National Park) located in the desert ecology of Chohlistan, Bahawalpur (Khan, 2010), i.e., 0.06 bird per ha.
The higher population density of the Grey Francolin in our cropland area during the winter and spring could also be related to the abundance of food in this habitat, because at that time the crops of Brassica campestris (sarsoon), Triticum aestivum (wheat), Lens culinaris (masoor) and Eruca sativa (taramira)were at their different growth stages and had become sources of succulent leaves, small grains as well as hosting high populations of insects. The diet of the Grey Francolin during the winter months has reportedly been dominated by mustard seeds and leaves in Layyah, Punjab (Mian and Wajid, 1994). Our findings are also supported by Kaiser (1998), who stated that Grey Partridges (Perdix perdix) occur in large numbers on farmlands and fields with a high diversity of crops. Similarly, maize or oil seeds, winter cereals and beans have been reported as preferred habitat of the Grey Partridge in Europe (Bro et al., 2004).
The current data on breeding biology provide only a limited scope from which to draw any conclusion but, as a first report on this aspect of the species in the Pothwar Plateau, it justifies its documentation. Our limited data is being reviewed in the light of published information, with the aim to consider it in designing any large-scale study.
In the present study, all nests were found on the ground in vegetation mainly comprising Desmostachia bipinnata, Acacia modesta, Imperata cylindrical, Zizipus jujuba and Euphorbia spp. These records agree with the observations by Islam (1999) on the location of the Grey Francolin in North America and that of Johnsgard (1973) who reported the occurrence of francolin nests under some protective cover for temperature modulation. However, Bump and Bump (1964) did find a Grey Francolin nest in a stacked pile of sorghum < 1.5 m above the ground in India. According to Sharma (1983), grasslands and plowed fields, as well as Euphorbia spp. were selected as nesting sites by Grey Francolins during the breeding season in Rajasthan, India.
The recorded egg-laying span (from 23 March to 7 May) of the current population coincided with earlier observations, i.e., for March to September (Ali and Ripley, 1969), March to April (Roberts, 1991) and mid-March to early May (Khan, 2010). The present study does not support the report by Waite (1948) from the salt-range part of the Pothwar Plateau suggesting that breeding may occur in December. The mean egg laying period of 6 ± 0.36 days and the average clutch size of 7 ± 0.36 eggs suggest that this population was probably laying eggs daily. The breeding behavior of this species studied in the Pakistan desert (Khan, 2010) has revealed that the number of days required to complete the clutch was 50% larger than clutch size, suggesting that the birds were not laying eggs daily. The average clutch size recorded in the present study is more or less within the range of clutch sizes reported by Baker (1921) of 4–10 eggs, Clark (1901), 8–10 eggs, Sharma (1983), 6–7 eggs. However, Ullah (1991) recorded clutch sizes in the range of 4–16 eggs and a study by Khan (2010) indicated an average clutch size of 7.22 ± 1.23 eggs (range 2–13, n = 76). A variation in clutch size from 4 to 10 eggs also has also been reported by Baker (1935), Long (1981) and Bump and Bump (1964). The maximum number of 8 eggs recorded in nests situated in wild vegetation on field boundaries of cropland habitats suggests that birds, living near food sources, have higher clutch sizes. The relationship between clutch size and food availability has also been documented for agricultural farms in Rajasthan, India (Sharma, 1983). The average incubation period (20.6 ± 0.50 days, range 19–22 days) recorded in the present study falls within the range of this duration as reported by Khan (2010) in a desert population (17.68 ± 2.10 days, range 16–21 days, n = 54). The current estimate also agrees with the values of this parameter in a range of 18–21 days, reported by Bump and Bump (1964), Ali and Ripley (1969) and Roberts (1991). The estimated hatching success (75.39%) is evidently higher than that (44.0 ± 3.36%, n = 85 clutches, 598 eggs) reported by Khan (2010) but any conclusion could not be drawn due to the low number of clutches available for this study. The same limitation may apply to the estimated overall breeding/fledging success (63.08%) of this study against the estimated (37.0 ± 3.25%) breeding success reported by Khan (2010). Khan (2010) has also reported a direct impact of rainfall on overall breeding parameters, indicating that the average clutch size and number of nestlings and fledgings increases with higher rainfall. The apparently higher breeding success in the Pothwar Plateau population could be attributed to the higher mean rainfall than that found in the desert of Bahawalpur, Pakistan.
The present study could identify only seven plant species and two insect species in the gut contents of the Grey Francolin hunted in agro-ecosystems of the Pothwar Plateau. The small sample size of the current study does not allow a meaningful comparison with any other study. However, an overview suggests that the number of identifiable food species is less than that reported (35 plant species and 16 animal species) from the Lal Suhanra National Park, located in a desert ecosystem of Bahawalpur, Pakistan (Khan, 2010). These are also smaller than the wide food base of this species reported by other investigators (Faruqui et al., 1960; Ullah, 1991; Chaudhry and Bhatti, 1992; Mian and Wajid, 1994). The present analysis suggests that insects, constituting 20.5% of the diet, is the only animal based food consumed by this group of francolins. The occurrence of only insects in the present samples is supported by Mian and Wajid (1994), but contrasts with the reports by Ali and Riplay (1983), Roberts (1991) and Choudhry and Bhatti (1992) who have reported the presence of reptiles and earthworm along with insects.
Our study shows that the Grey Francolin of the Pothwar agro-ecosystem is mainly associated with wild vegetation, inter-dispersed with cultivated tracks. The diet composition of the species suggests that it has little preference for any specific food species; rather, it consumes/prefers different food items depending upon their temporal and spatial availability in its habitat. Existing populations can be sustained provided the traditional system of cultivation and natural vegetation is maintained. The current rate of expansion in land use for urbanization and agriculture to meet fast growing human needs warrants extensive surveys of the Grey Francolin in the Pothwar ecology to assess its status, studying its feeding and breeding habit requirements, thus allowing the formulation of an effective conservation plan for the species.
Ahmad M. 1990. Groundnut in the Dry Land Agriculture of Pothwar. Barani Agricultural Research and Development Project, PARC, Islamabad, Pakistan, p 43.
Ali S, Ripley SD. 1983. A Pictorial Guide to the Birds of the Indian Subcontinent. Oxford University Press, New York, p 35.
Ali S, Ripley SD. 1969. Handbook of the Birds of India and Paksitan. Vol. 3. Oxford University Press, Bombay, India.
Baker EC. 1921. Indian Game Birds, Pheasants, Bustard, Quail. Bombay Nat Hist Soc, London, UK.
Baker ECS. 1935. The Nidification of Birds of the Indian Empire. Taylor and Francis, London, p 4.
Beg AR, Baig MS, Ali Q, Khan CMA. 1985. Agro-ecological zonation of Potwar: a-wheat, b-maize. National Agricultural Research Centre, Islamabad, Pakistan Forest Institute, Peshawar and Soil Survey of Pakistan, Lahore.
Beg MA, Qureshi JI. 1972. Birds and their habitats in the cultivated areas of Lyallpur district and vicinity. Pakistan J Agric Sci, 9: 161–166.
Bro E, Mayot P, Corda E, Reitz F. 2004. Impact of habitat management on grey partridge populations: Assessing wildlife cover using a multisite BACI experiment. J Appl Ecol, 41: 846–857.
Bump G, Bump JW. 1964. A study and review of the Black Francolin and the Grey Francolin. Special Scientific Report Wildlife (81): Bureau of Sport Fisheries and Wildlife, Washington D.C., USA 81: 53–57.
Burnham KP, Anderson DR, Laake JL. 1980. Estimation of density from line transects sampling of biological populations. Wildl Monogr, 72: 1–202.
Chaudhry AA, Bhatti MN. 1992. Biology of grey francolin (Francolinus pondicerianus) in the central Punjab plains. Abstract Proc. 12th Pakistan Congress of Zoology, Lahore, pp 161–162.
Clark A. 1901. Sport in the Low-country of Ceylon. Tisara Prakasakayo, Dehiwela.
del Hoyo J, Elliott A, Sargatal J (eds). 1994. Handbook of the Birds of the World. Volume 2. New World Vultures to Guineafowl. Lynx Edicions, Barcelona, Spain, pp 412–567.
Faruqui SA, Bump G, Nanda PC, Christensen GC. 1960. A study of the seasonal food of black francolin {Francolinus francolinus (Linnaeus)}, the gray francolin {Francolinus pondicerianus (Gmelin)} and the common sandgrouse {Pterocles exustus (Temminck)} in India and Pakistan. J Bombay Nat Hist Soc, 57: 354–361.
Gould J. 1966. Birds of Europe. Methuen, London, pp 49–50.
Helders S. 2007. Gujar Khan World Gazetteer. . Accessed on 5 September 2011
Islam K. 1999. Erckel's francolin (Francolinus erckelii), black francolin (Francolinus francolinus), and grey francolin (Francolinus pondicerianus). In: Poole A, Gill F (eds) The Birds of North America. The Birds of North America, Inc., Philadelphia, pp 394–396.
IUCN. 2011. IUCN Red List of Threatened Species. Version 2011.1. . Accessed on 05 September 2011.
Javed S, Qureshi Q, Rahmani AR. 1999. Conservation status and distribution of swamp francolin in India. J Bombay Nat Hist Soc, 96: 16–23.
Johnsgard PA. 1973. Grouse and quails of North America. University of Nebraska, Lincoln, p 553.
Kaiser W. 1998. Autumn winter habitat used by radio-tagged grey partridge (perdix perdix) in north Bavaria. Game Wildlife Sci, 15: 471–479.
Kamal S. 2000. Population status, distribution pattern and habitat of grey partridge (Francolinus pondicerianus) in district Mardan. Dissertations, Pakistan Forest Institute, Peshwar, p 50.
Khan WA. 2010. Studies on the comparative ecology of the south Persian black francolin, Francolinus francolinus henrici, and northern grey partridge, Francolinus pondicerianusinterpositus, in Lal Suhanra National Park, Bahawalpur, Punjab, Pakistan. Dissertations, Pir Mehr Ali Shah Arid Agriculture University, Rawalpindi, p 218.
Long JL. 1981. Introduced Birds of the World. David and Charles, London, p 89.
Mahmood K, Ahmad T, Khan A, Mahmood A, Mahmood W. 1997. Some notes on avifauna of Mangla reservoir, AJK, Pakistan. Pak J Ornith, 1: 1–2.
Mahmood S, Mahmood T, Rais M, Qureshi IZ, Nadeem MS. 2010. Comparative study on population and habitat of the grey and black francolins; Francolinus pondicerianus and Francolinus francolinus in Lehri Nature Park, Punjab, Pakistan. Podoces, 5(1): 42–53.
Mann MA, Chaudhry AA. 2000. Francolins in irrigated forest plantations and sub-mountainous tract of the Punjab, Pakistan. Pak Veterin J, 20(3): 118–122.
Mc Crow VP. 1982. Grey partridge habitat use and nesting biology in north central Iowa. Dissertations, Iowa State University, Ames, p 239.
Mian A, Ghani UI. 2007. Macrobiota variation under human protection in desert ecosystem of Cholistan, Pakistan. J Human Ecol, 21 (3): 163–172.
Mian A, Wajid MM. 1994. Food of grey francolin (Francolinus pondicerianus) in Layyah District (southern Punjab, Pakistan). Pakistan J Zool, 26: 185–187.
Nizami MI, Shafiq M, Rashid A, Aslam A. 2004. The soils and their agricultural development potential in Pothwar. WRRI and NARC, Islamabad, Pakistan, p 5–7.
Qureshi JI. 1972. Notes on the useful feeding activities of birds in field areas of Lyallpur. Pakistan J Agr Sci, 9 (3): 35–40.
Richard AF, Caroll JP, McGown JPK. 2002. Partridges, Quails, Francolins, Snowcocks, Guineafowl and Turkeys. Status survey and conservation. World Pheasant Associ. BirdLife International /SSC Partridges, Quails and Francolin specialist group. IUCN: The World Conservation Union. Gland, Switzerland, p 58.
Roberts TJ. 1991. The Birds of Pakistan, Non-passeriformes. Oxford University Press, New York, pp 232–233.
Shafiq MM. 2005. Wildlife Acts and Rules of Pakistan. unpublished report. Pakistan Forest Institute, Peshawar, p 297.
Sharma IK. 1983. The grey partridge (Francolinus pondicerianus) in the Rajasthan desert. Ann Arid Zone, 22: 117–120.
Ullah H. 1991. Studies on the biology, habitat, distribution pattern, and food of the grey partridge (Francolinus pondicerianus) in tehsil, Faisalabad. Dissertations, University of Agriculture, Faisalabad, p 182.
Waite HW. 1948. The birds of the Punjab salt range (Pakistan). J Bombay Nat Hist Soc, 48(1): 97–117.
Wijeyamohan S, Vandercone R, Santiapillai C. 2003. Observations on the grey francolin (Francolinus pondicerianus) in the vicinity of Giant's Tank, Sri Lanka. PQF News. 19: 11–14.
Yeatter RE. 1934. The Hungarian Partridge in Great Lakes Region. University of Michigan Press, Michigan, p 92.
Table
1.
Record of population densities of the Grey Francolin (Francolinus pondicerianus)in cropland habitats of the Gujar Khan area in the Rawalpindi district, Pakistan. Each transect has a length of 500 m and a width of 50 m with an area of 2.5 ha.
Table
2.
Record of population densities of the Grey Francolin (Francolinus pondicerianus)in scrub forest habitats of the Gujar Khan area in the Rawalpindi district, Pakistan. Each transect has a length of 500 m and a width of 50 m with an area of 2.5 ha.
Season
Month
Transect A
Transect B
Transect C
Mean density(n per ha)(Mean ± SE)
n
Crop and vegetation
n
Crop and vegetation
n
Crop and vegetation
Monthly
Seasonal
Autumn
Oct
–
Acacia modesta (dominant), A. nilotica (rare)
–
Acacia modesta (dominant), Zizyphus mausitiana, Typha angustata (common), Dalbergia sissoo (rare), Desmostachia bipinnata (scattered in patches) along a small wetland
5
Acacia modesta (dominant), Dalbergia sissoo, A. nilotica, Zizyphus mausitiana and Desmostachiabipinnata (common)
Table
5.
Composition of different food items recovered from crops and gizzards of the Grey Francolin (Francolinus pondicerianus) inhabiting an agro-ecosystem of the Pothwar Plateau, Pakistan
Sr. No.
Timeof hunt
Month (Season)a
Habitat
Body wt. (g)
Dry wt. of food contents (g)
Wt. of food contents per 100 g body wt.
Millet (seeds)
Sorghum(seeds)
Vilayati Kikar (leaves)
Mung Beans(seeds)
Acaciasp. (seeds)
Baru (seeds)
Carthemus oxycantha(seeds)
Termites
Black Ant
Grit
Unidentified
1
Evening
Nov (W)
Cropland
250
7.0
2.80
2.5
–
2.5
–
–
–
–
–
–
2.0
–
2
Evening
Dec (W)
Forest
263
12.0
4.56
1.5
2.5
–
2.0
–
2.5
–
–
–
3.0
0.5
3
Evening
Dec (W)
Cropland
210
9.0
4.29
1.7
–
–
3.2
–
–
–
1.3
1.0
1.5
0.3
4
Evening
Dec (W)
Cropland
242
6.0
2.48
–
2.5
–
1.5
–
–
–
–
1.0
0.8
0.2
5
Morning
Jan (W)
Forest
200
3.0
1.50
–
–
–
–
–
–
1.7
–
–
0.9
0.4
6
Evening
Jan (W)
Cropland
267
13.0
4.87
–
5.0
5.5
1.0
–
–
–
–
–
1.0
0.5
7
Evening
Feb (W)
Cropland
247
13.0
5.26
–
5.5
2.0
–
–
–
–
1.2
4.0
–
0.3
8
Evening
May (S)
Cropland
118
1.0
0.85
0.6
–
0.25
–
–
–
–
–
–
–
0.15
9
Morning
May (S)
Cropland
168
3.0
1.79
–
–
0.55
0.5
–
–
–
–
–
1.0
0.95
10
Morning
May (S)
Cropland
155
2.0
1.29
0.8
–
–
–
–
–
–
–
–
0.5
0.7
11
Evening
July (S)
Forest
257
8.5
3.31
–
–
–
–
1.5
–
–
5.5
–
1.0
0.5
12
Evening
July (S)
Forest
240
7.5
3.13
–
–
–
–
2.0
–
–
3.8
–
1.5
0.2
13
Morning
July (S)
Forest
235
2.0
0.85
–
–
–
–
1.2
–
–
–
–
0.5
0.3
Frequency of occurrence of identifiable food items n (%)
–
–
–
5 (11.9)
4 (9.5)
5 (11.9)
5 (11.9)
3 (7.1)
1 (2.4)
1 (2.4)
4 (9.5)
3 (7.1)
11 (26.2)
12
Mean ± SE
219.4 ± 12.9
6.7 ± 1.2
2.8 ± 0.4
1.4 ± 0.3
3.9 ± 0.8
2.2 ± 0.9
1.6 ± 0.5
1.6 ± 0.2
2.5 ± 0.0
1.7 ± 0.0
3.0 ± 1.2
2.0 ± 1.0
1.2 ± 0.2
0.4 ± 0.07
Total weight (% proportion) b
–
–
–
7.1(8.2%)
15.5(17.8%)
10.8(12.4%)
8.2(9.4%)
4.7(5.4%)
2.5(2.9%)
1.7(2.0%)
11.8(13.6%)
6.0(6.9%)
13.7(15.7%)
5.0(5.7%)
a W = winter; S = summer. b Totally, seeds = 45.6%; leaves = 12.4%; insects = 20.5%. Latin names of some of food items: Millet, Pennisetum typhoideum; Sorghum, Sorghum bicolor; Vilayati Kikar, Prosopis juliflora; Mung Beans, Phaseolus radiatus; Baru, Sorghum halepense; Termites, Coptotermes formosanus; Black Ant, Formica rufa.

 DownLoad:
DownLoad:






 Email Alerts
Email Alerts RSS Feeds
RSS Feeds