Figure
1.
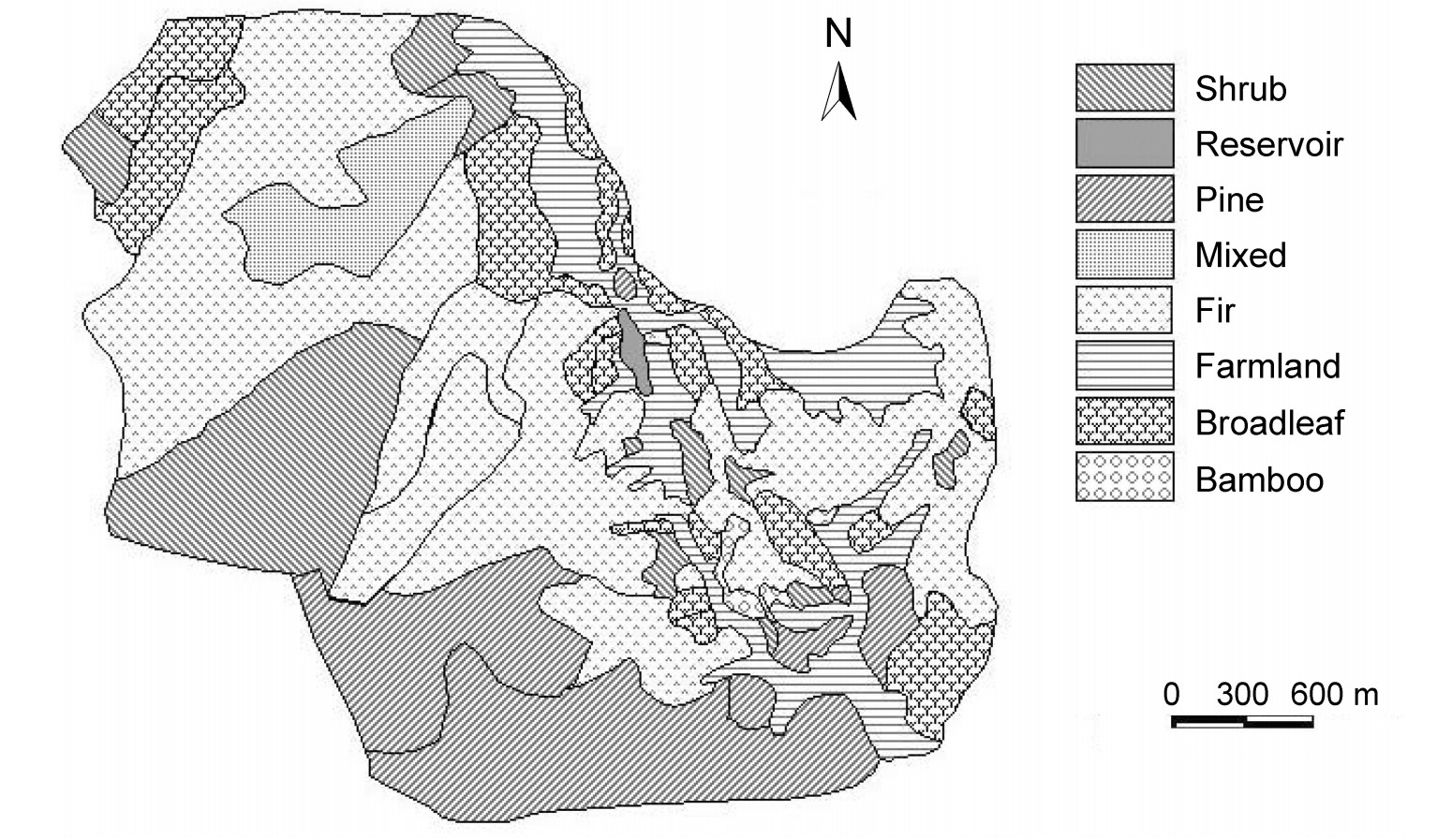
Habitats on Xianjuding Forest Farm in Hubei Province, China
| Citation: |
Jiliang XU, Zhengwang ZHANG. 2011: Home range and habitat composition of male Reeves's Pheasants in an agricultural-forest plantation landscape in central China: a preliminary report. Avian Research, 2(1): 53-58. DOI: 10.5122/cbirds.2011.0006

|
We present a preliminary examination of the home range and habitat use of male Reeves's Pheasants (Syrmaticus reevesii) in an agricultural-forest plantation landscape on the Xianjuding Forest Farm, Hubei Province, central China. Fieldwork was carried out from March to August in 2003. The home range of males averaged 33.17±12.55 ha by MCP (minimum convex polygon) and 21.05±5.61 ha by a 95% fixed kernel estimator. The core area by a 60% fixed kernel estimator was 3.92±0.27 ha. We did not detect significant seasonal variations in home range, core area and movement in this farm. Chinese fir (Cunninghamia lanceolata) plantations were the dominant habitat type within the home ranges and core areas. The males used their habitat non-randomly in spring and summer, preferred the fir plantations and avoided broadleaf forests in both seasons. In addition, the males used shrubs less than were available in the spring. The vegetation structure of different habitats may be the leading factor affecting the use of the habitats.
Habitat selection and availability of suitable habitats differ among populations across spatial and temporal scales as distribution and quality of resources vary over space and time (Boyce, 2006; Avila-Flores et al., 2010). Many widespread species respond differently to local habitat conditions among diverse parts of their populations (Avila-Flores et al., 2010). Edge populations, for instance, are typically restricted to habitat islands immersed in a matrix of unsuitable habitats (Hampe and Petit, 2005). In consequence, regional data on the behavior of some animals are not universally applicable (e.g. pheasants, see Gatti et al., 1989). Therefore, it is necessary to conduct similar studies in a number of different places in order to obtain a complete understanding of the behavior of wildlife, e.g. habitat use and movements.
Reeves's Pheasant (Syrmaticus reevesii) is a threatened species endemic to China (Cheng, 1987) and has been extirpated from much of its historical range in China (Zheng and Wang, 1998). Some of its populations are found in nature reserves and others in areas outside these reserves (Xu et al., 2007) and even in some forest farms subjected to heavy logging. Many investigations have been conducted on the habitat use of this species (Sun et al., 2002; Xu et al., 2006, 2007, 2011), its home range (Sun et al., 2003; Xu et al., 2009), daily movement patterns and site fidelity (Xu et al., 2009) in a national nature reserve; however, little is known about the habitat and home range of this pheasant in areas outside nature reserves, especially on forest farms.
Therefore, we examined home range, daily movement and habitat use of this pheasant using radio-telemetry on a forest farm. Our main objectives were: 1) to estimate home range size and daily movements and 2) to investigate habitat composition of these home ranges.
Field work was conducted from March to August in 2003 on the Xianjuding Forest Farm (114°26′48″E, 31°32′20″N), over an area of about 2162 ha located on the northern slopes of the Dabie mountain range, Hubei Province, central China. The forest farm is situated at the edge of the northern subtropical zone. Its vegetation is characterized by mature plantations (Fig. 1) dominated by Masson pine (Pinus massoniana), Larix spp., Chinese fir (Cunninghamia lanceolata), Metasequoia glyptostroboides and China Wingnut (Pterocarya stenoptera). The shrubby areas are dominated by young oaks (Quercus spp.), young Chinese firs, glaucous allspice (Lindera glauca) and teas (Camellia spp.). The elevation of this forest farm ranges from 300 to 600 m, with the highest peak at 680.2 m. The climate is warm and humid with a mean annual temperature of 15.4℃ and a mean annual precipitation of 1114.9 mm.
We used the method described in Xu et al.(2007, 2009) to capture and track the males on the Xianjuding Forest Farm in March 2003. Birds were located in the spring (from March to May) and summer (from June to August) (Xu et al., 2007, 2009).
With the use of the Animal Movement software and detected locations, the home range was estimated based on a 100% minimum convex polygon (MCP) and a 95% fixed kernel. We defined a 60% fixed kernel as a core area (Xu et al., 2009). All fixed kernel estimations were based on least squares cross validation for deriving a smoothing factor. We used the size of the home range, obtained by the 95% fixed kernel contours, for additional analyses. The average distance between consecutive locations (every 12 hrs) was calculated and used as an index of daily mobility for each individual, referred to as daily movement (Cardinal and Paxton, 2005). First, we assessed the home range, core areas and daily movement of the males in the breeding seasons (including spring and summer) and then made independent assessments in the spring and summer.
A 1:10000 vegetation map supplied by the administration of the forest farm was "ground-truthed" by field observations to digitize and produce a land cover map of the study site. The habitats within the main study area were classified into pine plantations, broadleaf and conifer mixed forests, Chinese fir plantations, farmland, broadleaf forests, bamboo forests and shrubby areas (Fig. 1) according to present plant species and their coverage.
We followed Neu et al. (1974), using a χ2 test of goodness of fit and a Bonferroni Z-statistic to test the habitat use of Reeves's Pheasant in the study area. From the radio locations of these pheasants during each season, a minimum convex polygon (MCP) was obtained and a 100 m buffer established, based on the MCP with some radio locations located on the borderline. After removing the area of the reservoir, the MCP and the buffer comprised the available area. The proportion of radio locations within a habitat type was considered as used, while the proportion of the corresponding habitat type area in the available area as the control.
We used randomized repeated measures (Edgington, 1980) to test the difference in the home range and core areas of this pheasant between spring and summer seasons.
Statistics are provided as mean ± 1 standard deviation (SD). For all statistical tests, a probability of 0.05 or less was considered to be significant. Randomized tests were performed by psychStats (http://www.lcsdg.com/psychStats) online in 2006. Other statistical analyses were carried out using SPSS 10.0.1 for Windows (SPSS Inc., 1999).
We only tracked four male Reeves's Pheasants, given the difficulty in finding the birds (Xu et al., 2007) and the small population size of this pheasant in this area (Zhang, 2002). One male, however, was unfortunately killed by its predator after about two weeks. Therefore, we only included 202 radio locations from three individuals in our analyses.
The average home range size of male Reeves's Pheasants in the breeding seasons was 33.17 ± 12.55 ha based on minimum convex polygons and 21.05 ± 5.61 ha based on the 95% fixed kernel estimator. The average core area was 3.92 ± 0.27 ha. The size of the home range did not vary between spring and summer (test of randomized repeated measures, MCP: p = 0.25; 95% Kernel: p = 0.25; see Table 1). Similar trends prevailed in the core areas (test of randomized repeated measures, p = 0.50; see Table 1) and daily movements (test of randomized repeated measures, p = 1.000).
| Season | Individual code | Home range (ha) | Core area (ha) | Daily movements (m) | |
| MCP | Kernel (95%) | ||||
| Spring | M224 | 8.31 | 11.15 | 1.08 | 137.27 |
| M273 | 26.08 | 24.70 | 6.77 | 212.46 | |
| M793 | 47.56 | 34.84 | 5.55 | 188.48 | |
| 27.31 ± 19.65 | 23.56 ± 11.88 | 4.47 ± 3.00 | 179.40 ± 38.41 | ||
| Summer | M224 | 6.05 | 7.70 | 1.96 | 127.44 |
| M273 | 26.00 | 20.39 | 3.36 | 211.43 | |
| M793 | 14.72 | 24.36 | 4.58 | 205.21 | |
| 15.59 ± 10.00 | 17.48 ± 8.70 | 3.30 ± 1.31 | 181.36 ± 47.00 | ||
 DownLoad:
CSV
DownLoad:
CSV
The habitat composition of home ranges and core areas varied among different individuals. The Chinese fir plantation dominated the home ranges and core areas of the male pheasants (Table 2). In particular, male birds could use part of the bamboo forests which accounted for 26.44% of the home range in the spring and 27.09% in the summer, while it constituted about 50% of the core area in the spring and summer seasons.
| Types | Season | Individual code | Shrub | Fir | Broadleaf | Bamboo | Farmland |
| Home range | Spring | M224 | 2.25 | 47.01 | 11.84 | 26.44 | 12.46 |
| M273 | 7.15 | 66.15 | 16.81 | 0.51 | 9.38 | ||
| M793 | 1.27 | 50.10 | 24.43 | 0.54 | 23.67 | ||
| Summer | M224 | 3.48 | 24.74 | 12.91 | 37.09 | 21.78 | |
| M273 | 8.16 | 65.44 | 16.26 | 3.80 | 6.34 | ||
| M793 | 0.04 | 48.77 | 35.58 | 0 | 14.61 | ||
| Core areas | Spring | M224 | 0 | 43.89 | 1.68 | 49.31 | 5.13 |
| M273 | 21.84 | 70.41 | 5.91 | 0 | 1.84 | ||
| M793 | 0 | 72.07 | 2.72 | 0 | 25.21 | ||
| Summer | M224 | 0 | 16.34 | 18.57 | 51.68 | 13.41 | |
| M273 | 31.67 | 68.33 | 0 | 0 | 0 | ||
| M793 | 0 | 94.88 | 0 | 0 | 5.12 |
DownLoad:
CSV
The male pheasants used these habitat types selectively in spring and summer (Spring: χ2 = 12.99, df = 4, p = 0.011; Summer: χ2 = 11.39, df = 4, p = 0.023). Largely, the males used the Chinese fir plantations and avoided the broadleaf forests in both spring and summer seasons (Table 3). It also avoided shrubby areas in the spring, while these areas were used in proportion to their availability in the summer (Table 3). Moreover, the bird used farmland and bamboo forests in proportion to their availability.
| Habitat type | Availability a | Spring | Summer | |||||||||
| Rate of use b | Number of use c | Confidence interval | U vs. A d | Rate of use | Number of use | Confidence interval | U vs. A | |||||
| Lower | Upper | Lower | Upper | |||||||||
| Shrub | 0.064 | 0.020 | 2 | –0.016 | 0.056 | -- | 0.030 | 3 | –0.014 | 0.073 | ||
| Fir | 0.537 | 0.703 | 71 | 0.586 | 0.820 | ++ | 0.683 | 69 | 0.564 | 0.803 | ++ | |
| Farm | 0.035 | 0.089 | 9 | 0.016 | 0.162 | 0.050 | 5 | –0.006 | 0.105 | |||
| Broadleaf | 0.204 | 0.079 | 8 | 0.010 | 0.149 | -- | 0.059 | 6 | –0.001 | 0.120 | -- | |
| Bamboo | 0.160 | 0.109 | 11 | 0.029 | 0.189 | 0.178 | 18 | 0.080 | 0.276 | |||
|
a Availability is the proportion of area of a type of habitat in the available area; b Rate of use is the proportion of radiolocations in the corresponding habitat; c Number of use is the number of radiolocations in the corresponding habitat; d "U vs. A" means "use vs. availability". | ||||||||||||
DownLoad:
CSV
An understanding of the movement and habitat use of pheasants is fundamental to habitat management regimes. Our findings are the first step towards a more detailed knowledge of the responses of Reeves's Pheasant to habitat conditions in an artificial landscape, i.e. a forest farm. However, caution must be taken when interpreting our results given the small sample size. We also had to limit our results to males of Reeves's Pheasant, for there is little information available on females in these habitats. Nevertheless, given its conservation status and the relatively high proportion of artificial plantations in its range of distribution, the knowledge on the biology of this bird in artificial plantations remains scarce.
The size of the home ranges and core areas of the male pheasants in the Dongzhai National Nature Reserve in Henan Province (Sun et al., 2003; Xu et al., 2009) seemed larger than those we assessed in this landscape. Many factors could have great impacts on the size of the home range, such as body size (Jenkins and Benn, 1998), habitat quality and food abundance (Elchuk and Wiebe, 2003), and human disturbance (Koehler and Pierce, 2003). However, these differences may result from the small sample size on the Xianjuding Forest Farm and it is therefore necessary to track more individuals on this farm in the future.
The habitat composition of the home ranges and core areas of male Reeves's Pheasants varied greatly among different individuals, implying this pheasant could adapt itself to different habitat conditions. As one of the typical plantations, Chinese fir plantations now occupy a larger part, especially on some forest farms (Zhang et al., 2000). This male pheasant prefers Chinese fir plantations and shows no preference to broadleaf forests on the farm. Chinese fir plantations on the Xianjuding Forest Farm have a dense canopy with sparse undergrowth, similar to that in the Dongzhai Nature Reserve (Xu et al., 2007). In contrast, the canopy of broadleaf forests in this area is sparse as is its undergrowth. In particular, Chinese fir plantations in this area should be beneficial to the activities of this pheasant, whereas the broadleaf might limit it (Xu et al., 1991).
The bamboo forests did not account for much on this farm, but occupied a large part of the home range of one bird we tracked. We also found some activity traces of males in bamboo forests in the field. Overall, the structure of bamboo forests is more or less similar to that of Chinese fir plantations and very convenient for this pheasant to move inside. However, Reeves's Pheasants roost in big trees (Sun et al., 2002) and, indeed we found the males we tracked in the bamboo forests to be roosting in big Chinese fir trees or oaks. Therefore, it was reasonable to state that the landscape structure of its habitat might have an effect on the habitat use of this pheasant.
The farmland might provide supplementary food for Reeves's Pheasants in some places, especially in the winter (Wu et al., 1994). However, farmland was not an important habitat for the males in the Xianjuding Forest Farm, a condition also reported in the Dongzhai National Nature Reserve (Xu et al., 2007, 2009) although we observed some non-tracked individuals occasionally foraging in farmland near the forest edge, implying that there was sufficient food within their home ranges, but a lack of cover made them susceptible to predation (Xu et al., 2002).
Funding and support for this study were provided by the Forestry Commonweal Programs of the Ministry of Science & Technology of China (No. 200904003) and the National Natural Science Foundation of China (No. 30800103).
|
Cardinal SN, Paxton EH. 2005. Home range, movement, and habitat use of the Southwestern Willow Flycatcher, Roosevelt Lake, AZ – 2004. U. S. Geological Survey report to the U. S. Bureau of Reclamation, Phoenix, AZ.
|
|
Cheng TH. 1987. A Synopsis of the Avifauna of China. Science Press, Beijing, p 167.
|
|
Edgington ES. 1995. Randomization Tests (3rd ed). Marcel Dekker, New York.
|
|
Jenkins AR, Benn GA. 1998. Home range size and habitat requirements of peregrine falcons on the Cape Peninsula, South Africa. J Raptor Res, 32: 90–97.
|
|
Sun QH, Zhang ZW, Zheng GM, Zhang KY, Ruan XF, Zhu JG. 2003. Ranging behavior of territorial male Reeves's Pheasants in the breeding season. Acta Zool Sinica, 49: 318–324. (in Chinese with English abstract)
|
|
Sun QH, Zhang ZW, Zhu JG, Gao ZJ. 2002. Roosting behavior and factors affecting roost-site used by Reeves's Pheasant (Syrmaticus reevesii). J Beijing Normal Univ (Nat Sci), 38: 108–112. (in Chinese with English abstract)
|
|
Wu ZK, Li ZM, Wang JH. 1994. Progress in research on Reeve's Pheasant in China. Annual Review of the World Pheasant Association, pp 39–43.
|
|
Xu WS, Wu ZK, Li ZM. 1991. Reeves's Pheasant (Syrmaticus reevesii). In: Lu TC, Liu RS, He FQ (eds) The Rare and Endangered Gamebirds in China. Fujian Science and Technology Press, Fuzhou, pp 328–338. (in Chinese)
|
|
Zhang ZW. 2002. Survey of the habitat fragmentation of Reeves's Pheasant. Unpublished Small Grant Report to WWF-China, p 16.
|
|
Zheng GM, Wang QS. 1998. China Red Data Book of Endangered Animals (Aves). Science Press, Beijing. (in Chinese)
|
Figures(1) / Tables(3)





| Season | Individual code | Home range (ha) | Core area (ha) | Daily movements (m) | |
| MCP | Kernel (95%) | ||||
| Spring | M224 | 8.31 | 11.15 | 1.08 | 137.27 |
| M273 | 26.08 | 24.70 | 6.77 | 212.46 | |
| M793 | 47.56 | 34.84 | 5.55 | 188.48 | |
| 27.31 ± 19.65 | 23.56 ± 11.88 | 4.47 ± 3.00 | 179.40 ± 38.41 | ||
| Summer | M224 | 6.05 | 7.70 | 1.96 | 127.44 |
| M273 | 26.00 | 20.39 | 3.36 | 211.43 | |
| M793 | 14.72 | 24.36 | 4.58 | 205.21 | |
| 15.59 ± 10.00 | 17.48 ± 8.70 | 3.30 ± 1.31 | 181.36 ± 47.00 | ||
DownLoad:
CSV
| Types | Season | Individual code | Shrub | Fir | Broadleaf | Bamboo | Farmland |
| Home range | Spring | M224 | 2.25 | 47.01 | 11.84 | 26.44 | 12.46 |
| M273 | 7.15 | 66.15 | 16.81 | 0.51 | 9.38 | ||
| M793 | 1.27 | 50.10 | 24.43 | 0.54 | 23.67 | ||
| Summer | M224 | 3.48 | 24.74 | 12.91 | 37.09 | 21.78 | |
| M273 | 8.16 | 65.44 | 16.26 | 3.80 | 6.34 | ||
| M793 | 0.04 | 48.77 | 35.58 | 0 | 14.61 | ||
| Core areas | Spring | M224 | 0 | 43.89 | 1.68 | 49.31 | 5.13 |
| M273 | 21.84 | 70.41 | 5.91 | 0 | 1.84 | ||
| M793 | 0 | 72.07 | 2.72 | 0 | 25.21 | ||
| Summer | M224 | 0 | 16.34 | 18.57 | 51.68 | 13.41 | |
| M273 | 31.67 | 68.33 | 0 | 0 | 0 | ||
| M793 | 0 | 94.88 | 0 | 0 | 5.12 |
DownLoad:
CSV
| Habitat type | Availability a | Spring | Summer | |||||||||
| Rate of use b | Number of use c | Confidence interval | U vs. A d | Rate of use | Number of use | Confidence interval | U vs. A | |||||
| Lower | Upper | Lower | Upper | |||||||||
| Shrub | 0.064 | 0.020 | 2 | –0.016 | 0.056 | -- | 0.030 | 3 | –0.014 | 0.073 | ||
| Fir | 0.537 | 0.703 | 71 | 0.586 | 0.820 | ++ | 0.683 | 69 | 0.564 | 0.803 | ++ | |
| Farm | 0.035 | 0.089 | 9 | 0.016 | 0.162 | 0.050 | 5 | –0.006 | 0.105 | |||
| Broadleaf | 0.204 | 0.079 | 8 | 0.010 | 0.149 | -- | 0.059 | 6 | –0.001 | 0.120 | -- | |
| Bamboo | 0.160 | 0.109 | 11 | 0.029 | 0.189 | 0.178 | 18 | 0.080 | 0.276 | |||
|
a Availability is the proportion of area of a type of habitat in the available area; b Rate of use is the proportion of radiolocations in the corresponding habitat; c Number of use is the number of radiolocations in the corresponding habitat; d "U vs. A" means "use vs. availability". | ||||||||||||
DownLoad:
CSV
 Email Alerts
Email Alerts RSS Feeds
RSS Feeds