Jiliang XU, Zhengwang ZHANG. 2011: Home range and habitat composition of male Reeves's Pheasants in an agricultural-forest plantation landscape in central China: a preliminary report. Avian Research, 2(1): 53-58. DOI: 10.5122/cbirds.2011.0006
Citation:
Jiliang XU, Zhengwang ZHANG. 2011: Home range and habitat composition of male Reeves's Pheasants in an agricultural-forest plantation landscape in central China: a preliminary report. Avian Research, 2(1): 53-58. DOI: 10.5122/cbirds.2011.0006
Jiliang XU, Zhengwang ZHANG. 2011: Home range and habitat composition of male Reeves's Pheasants in an agricultural-forest plantation landscape in central China: a preliminary report. Avian Research, 2(1): 53-58. DOI: 10.5122/cbirds.2011.0006
Citation:
Jiliang XU, Zhengwang ZHANG. 2011: Home range and habitat composition of male Reeves's Pheasants in an agricultural-forest plantation landscape in central China: a preliminary report. Avian Research, 2(1): 53-58. DOI: 10.5122/cbirds.2011.0006
College of Nature Conservation, Beijing Forestry University, Beijing 100083, China
2.
Ministry of Education Key Laboratory for Biodiversity Sciences and Ecological Engineering, College of Life Sciences, Beijing Normal University, Beijing 100875, China
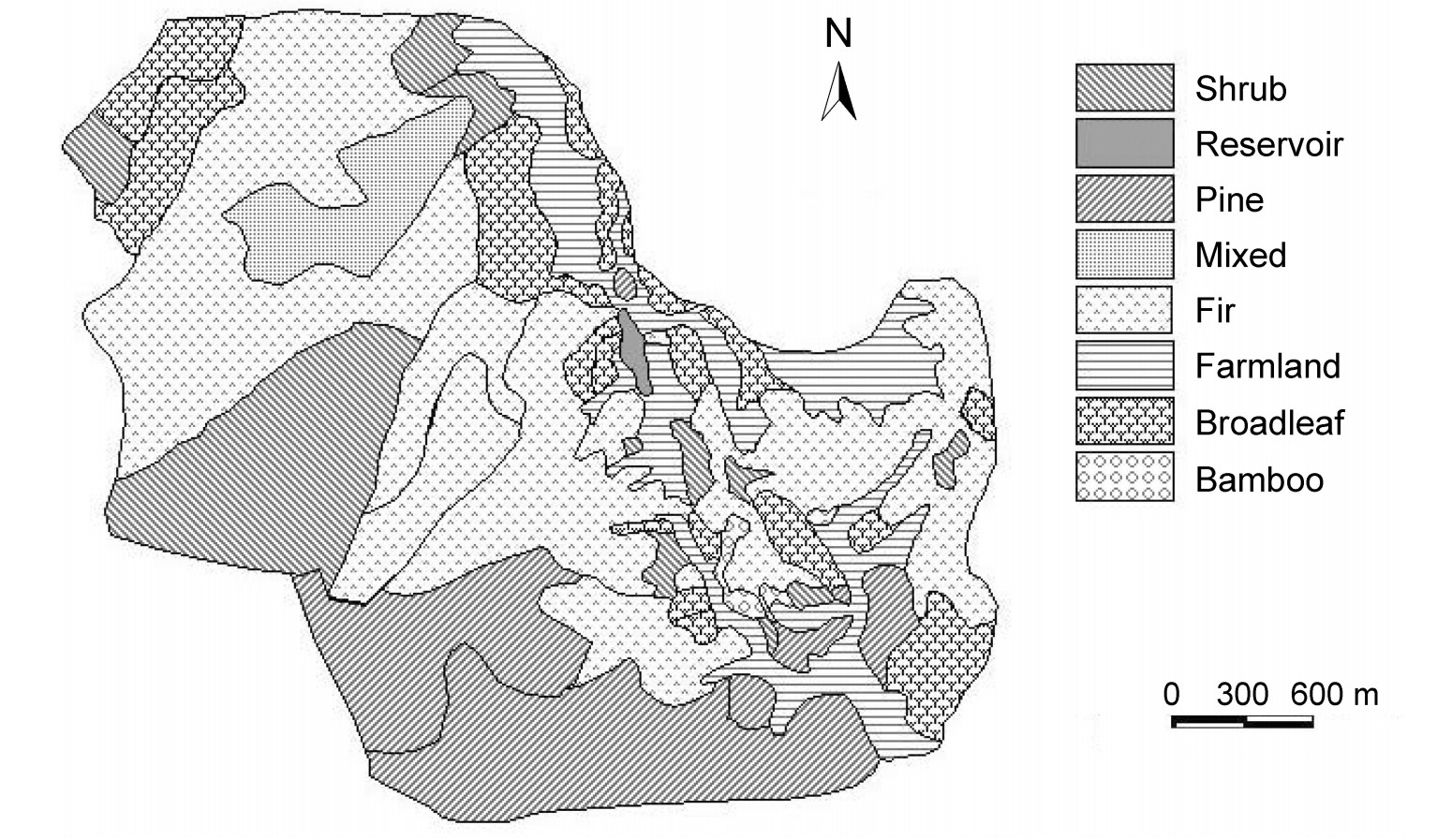
We present a preliminary examination of the home range and habitat use of male Reeves's Pheasants (Syrmaticus reevesii) in an agricultural-forest plantation landscape on the Xianjuding Forest Farm, Hubei Province, central China. Fieldwork was carried out from March to August in 2003. The home range of males averaged 33.17±12.55 ha by MCP (minimum convex polygon) and 21.05±5.61 ha by a 95% fixed kernel estimator. The core area by a 60% fixed kernel estimator was 3.92±0.27 ha. We did not detect significant seasonal variations in home range, core area and movement in this farm. Chinese fir (Cunninghamia lanceolata) plantations were the dominant habitat type within the home ranges and core areas. The males used their habitat non-randomly in spring and summer, preferred the fir plantations and avoided broadleaf forests in both seasons. In addition, the males used shrubs less than were available in the spring. The vegetation structure of different habitats may be the leading factor affecting the use of the habitats.
The Malayan Night Heron (Gorsachius melanolophus) inhabits a range of environments from India and Sri Lanka to Southeast Asia, southern China, Malaysia, Thailand, Taiwan, Indonesia, the Philippines and Japan (del Hoyo et al., 1992; The Ornithological Society of Japan, 2000). This species prefers tropical and subtropical wet forests as its breeding habitats (Hancock et al., 1978; del Hoyo et al., 1992). While it is a migratory bird in some areas (Hancock et al., 1978), it is a resident species in Japan (The Ornithological Society of Japan, 2000). Its distribution in Japan is usually restricted to the Yaeyama and Miyako islands, Okinawa Prefecture (Kawakami and Fujita, 2005). This heron is considered to be uncommon throughout much of its range (Kushlan and Hancock, 2005) and is classified as a vulnerable species in the Japanese Red List released in 2006. Understanding the diet of such a species is a very important issue for conservation because it relates closely to a species' ecological niche.
Little is known about the ecology of this heron species and only piecemeal information on its diet has been available, as this species breeds singularly, does not make a colony or communal roost, and inhabits shadowy forests. Shen and Chen (1996) described that 93% of the diet of the Malayan Night Heron were earthworms based on direct observation. Kushlan and Hancock (2005) estimated that the herons specialized in capturing frogs and very large earthworms, and also preyed on beetles, mollusks, crabs, snakes, lizards, birds and small fish. Regarding to the Japanese population, their diet has been seldom documented except for occasional reports (e.g. Makihara and Kawakami, 2005). However, no quantitative information of their food items has been available. The results of direct observations might bias towards large and noticeable objects at well-lighted area, and the proportion of inconspicuous items is very likely to be underestimated. To show a detailed dietary composition, we collected pellets regurgitated by the herons and stomach contents of dead individuals, because these analyses could provide quantitative estimation without the influence of ease of observation. In this paper, the dietary composition of the heron is described and its foraging microhabitats and circadian activity are discussed in light of dietary preference.
Methods
Pellets cast by Malayan Night Herons were collected under their roosts with the aid of mesh nets. Plastic mesh nets of 2 m × 2 m were located under the roost at a 1 m height from the ground and the pellets cast on the nets were collected after a period of days. The herons roost alone on trees at deciduous forests in the islands. The samples were kept in a freezer until the subsequent analysis. Collections were conducted in natural dense humid forests (Mt. Banna-dake) and riparian forests (Miyara River) on Ishigaki Island in March and April 2002 and in a secondary forest on Kohama Island in March 2003 (Fig. 1). The vegetation was mainly deciduous evergreen forest consisting of Castanopsis sieboldii, Quercus Miyagii, Hibiscus tiliaceus and Ficus trees. Additionally, the stomach contents of five dead birds were analyzed, four of them found on Ishigaki Island and one on Panari Island (Table 1). Three carcasses collected on Ishigaki Island were roadkill and one was rescued infirm. We extracted fragmented animal remains from the pellets and stomach contents, identified them to the lowest possible taxonomic level with the stereomicroscope, and counted the minimum number of each species found in each pellet.
Figure
1.
Map of the Yaeyama Islands, Japan. The black circles show the locations of pellet collection survey.
Thirty-eight pellets were collected on Ishigaki Island and five on Kohama Island, respectively. All pellets included a mass of sands and gravels, which were estimated to be swallowed with soil animals. The undigested remains of land snails, chilopods, arachnids, crabs, insects and reptiles were found among them (Table 2). Forty-three individuals of snails were found in 47% of the analyzed pellets and all of them were Acusta despecta. Two individuals of Chilopoda were found in one pellet on Ishigaki Island. Eight whip scorpions, nine spiders and unidentified arachnids were found in 30% of the pellets. The whip scorpions were certain to be Typopeltis crucifer, as no other species has been observed on the island. Twenty-two crabs were detected from 47% of the pellets, while all of the identified crabs were freshwater and 19 of them were of the family Potamidae. At least 69 individuals of 21 species of insects were detected from 65% of the pellets. Blattaria, Elateridae and Scarabaeidae were dominant and covered 55% of the total number of individuals. Nine reptiles were detected in 19% of all pellets and the dominant species was an agama lizard, Japalura polygonata ishigakiensis.
Table
2.
The species composition found in the pellets and stomach contents of Malayan Night Herons. Each number shows the number of individuals for each food item. The number in brackets is the percent occurrence of samples containing items for each taxon.
In the stomach contents of five carcasses, an arm of a frog Fejervarya sakishimaensis and several insects were detected (Table 2). The frog was estimated to be an adult and about 10 cm in its entire length. The dominant insects were cockroaches and grasshoppers, which were each detected in four of the five carcasses.
Discussion
Land snails, freshwater crabs and insects were dominantly detected in the diet of Malayan Night Herons. In insect species, cockroaches, click beetles and scarabs were the major prey organisms found in the pellets, whereas the proportions of cockroaches and grasshoppers detected were high in the stomach contents of carcasses. Most of the carcasses were collected in open areas, while pellets were found in forests. As grasshoppers frequently inhabit grasslands, the differences among them were consistent with their habitat uses.
The land snails found in the pellets were major terrestrial species found on the forest floor of Ishigaki Island. Freshwater crabs inhabit streams and the ground nearby. The main microhabitats of the detected cockroaches, click beetles, whip scorpions, spiders, and Scolopendra were litter, soil, and fallen and decaying trees, indicating that the herons prefer to forage on soil animals on the forest floor. Though the agama lizard is arboreal, they often use ground for movement. The scarabs are arboreal and aerial, but observing them walking along the ground is not uncommon. Since the night herons were unlikely to forage on trees, they probably occasionally preyed on scarabs on the ground. Such arboreal animals are suggested to contribute the diet menu of the birds as well as soil animals. Especially, the contribution of the lizards ought to be larger than the other species due to their larger body masses.
Acusta despecta detected in the pellets has nocturnal activity as with other various terrestrial mollusks (Kurozumi, 1997; Cook, 2001). Freshwater crabs are primarily nocturnal and remain hidden during the day in sheltered places (Yeo et al., 2008). The identified Coleoptera, Orthoptera and Blattaria are also nocturnal, which get active from evening. On the other hand, the agama lizards are diurnal (Tanaka, 1986). The herons had been considered to be generally crepuscular or nocturnal, while diurnal activities have been frequently observed (Brazil, 1991; del Hoyo et al., 1992). The current results implied that the herons spend both day and night times for foraging in response to the prey activities.
Although Kushlan and Hancock (2005) suggested that the herons foraged on fish in aquatic environments, distinct aquatic animals were not detected in the current study, while freshwater crabs and partial aquatic species were frequently found. The beak of this heron is shorter and thicker than that of typical herons, while other heron species generally have straight and pointed bills (del Hoyo et al., 1992), which are considered to be adapted to preying on fish. The bill morphology of the Malayan Night Heron is similar to that of the Japanese Night Heron (Gorsachius goisagi), which is considered to be adapted for foraging on soil animals by digging (Kawakami et al., 2005). Freshwater crabs are frequently found in litter layers around streams. Chang (2000) also noted that the Malayan Night Heron rarely foraged on fish. The current result and the morphological character imply that the dominant diet of the Malayan Night Heron consists of small animals in litter layers.
On the other hand, some distinct items, such as aquatic animals, might have been overestimated in their diet menu in the previous studies. The main component of their pellets was sand and gravels, which made the pellets breakable and indistinctive. When the pellets would go into the ground, they ought to be easy to break into pieces. It becomes hard to detect the inconspicuous components such as soil insects due to their naturalness on the ground. Conversely, remains of aquatic animals should be distinct, because it is unnatural that such items are there. In order to avoid overestimation, pellet traps with plastic mesh nets are effective.
The herons appear to prefer wet ground, sometimes near streams, as foraging habitat, where they can capture land snails and freshwater crabs. Since we could not detect fully aquatic animals, the heron was suggested not to directly utilize aquatic habitats. They are believed to prefer moist forests associated with watery environments such as streams and swamps because of the availability of soil animals. These results are consistent with the foraging preferences of the congeneric Japanese Night Heron, which breeds on the mainland of Japan (Kawakami et al., 2005). To conserve the Malayan Night Heron, forests with well developed litter layers and fallen and decaying trees, which are generally rich in the biomass of soil animals, must be maintained.
Malayan Night Herons were described to mainly forage on earthworms based on direct observation (Shen and Chen, 1996; Chang, 2000), although we did not detect any because they have few indigestible parts. However, earthworms were also not observed in the carcasses analyzed in this study. Though the night herons should presumably forage on earthworms, the frequency is likely to vary according to habitat. Confirmation is needed as to how important earthworms are in the diet of the Malayan Night Heron. Combination of multiple methods is required in order to show the actual diet composition.
Frogs were not frequently detected in the current study, although Kushlan and Hancock (2005) estimated that they were one of the heron's main diet sources in forests. This might be caused by the above-mentioned overestimation of eye-catching items, while it is possibly attributed to seasonal variation of food availability. Herons generally have variable diets related to availability of prey in their habitat (del Hoyo et al., 1992) and can seasonally change their diet menus (e.g., Marquiss and Leitch, 1990; Rodriguez et al., 2007). Frogs become active during and after rain events in general and the biomass seasonally undergoes a complete change. Thus, although the herons may often forage on frogs in rainy seasons, all of the pellets analyzed in the current study were collected during the same period, i.e., spring. In addition to the seasonal variation of diet menu, the potential for variation exists depending on location. The dietary menu of the Malayan Night Heron in other seasons and places should be systematically investigated as the next step.
Acknowledgments
We are especially grateful to Taiji Kurozumi for identifying the snails, Tetsuya Watanabe and Yasuhisa Henmi for identifying crabs, Yuichi Ohba and Hitoo Ohira for identifying elaterids, and Akio Tanigawa for identifying arachnids, and for providing corresponding ecological information. We also thank Takashi Fujii, Takashi Hiraoka and the Yamashina Institute for Ornithology for providing the carcass samples, Ako Sukegawa, Minako Murakami, Nanami Kawamura, Ryoko Kanzawa, Yoko Hashiguchi and Yoichiro Sakiyama for collecting the pellets in the field, and Miyuki Mashiko for analyzing the samples.
Avila-Flores R, Boyce MS, Boutin S. Habitat selection by prairie dogs in a disturbed landscape at the edge of their geographic range. J Wildl Manage, 74: 945–953.
Cardinal SN, Paxton EH. 2005. Home range, movement, and habitat use of the Southwestern Willow Flycatcher, Roosevelt Lake, AZ – 2004. U. S. Geological Survey report to the U. S. Bureau of Reclamation, Phoenix, AZ.
Cheng TH. 1987. A Synopsis of the Avifauna of China. Science Press, Beijing, p 167.
Elchuk CL, Wiebe KL. 2003. Home-range size of northern flickers (Colaptes auratus) in relation to habitat and parental attributes. Can J Zool, 81: 954–961.
Gatti RC, Dumke RT, Pils CM. 1989. Habitat use and movements of female ring-necked pheasants during fall and winter. J Wildl Manage, 53: 462–475.
Hampe A, Petit RJ. 2005. Conserving biodiversity under climate change: the rear edge matters. Ecol Lett, 8: 461–467.
Jenkins AR, Benn GA. 1998. Home range size and habitat requirements of peregrine falcons on the Cape Peninsula, South Africa. J Raptor Res, 32: 90–97.
Koehler GM, Pierce DJ. 2003. Black bear home-range sizes in Washington: climatic, vegetative, and social influences. J Mammal, 84: 81–91.
Neu CW, Byers CR, Peek JM. 1974. Technique for analysis of utilization — availability data. J Wildl Manage, 38: 541–545.
Sun QH, Zhang ZW, Zheng GM, Zhang KY, Ruan XF, Zhu JG. 2003. Ranging behavior of territorial male Reeves's Pheasants in the breeding season. Acta Zool Sinica, 49: 318–324. (in Chinese with English abstract)
Sun QH, Zhang ZW, Zhu JG, Gao ZJ. 2002. Roosting behavior and factors affecting roost-site used by Reeves's Pheasant (Syrmaticus reevesii). J Beijing Normal Univ (Nat Sci), 38: 108–112. (in Chinese with English abstract)
Wu ZK, Li ZM, Wang JH. 1994. Progress in research on Reeve's Pheasant in China. Annual Review of the World Pheasant Association, pp 39–43.
Xu JL, Zhang XH, Sun QH, Zheng GM, Wang Y, Zhang ZW. 2009. Home range, daily movements and site fidelity of male Reeves's pheasants Syrmaticus reevesii in the Dabie Mountains, central China. Wildl Biol, 15: 338–344.
Xu JL, Zhang XH, Zhang ZW, Zheng GM, Ruan XF, Zhu JG. 2006. Multi-scale analysis on wintering habitat selection of Reeves's Pheasant (Syrmaticusreevesii) in Dongzhai National Nature Reserve, Henan Province, China. Acta Ecol Sinica, 26: 2061–2067. (in Chinese with English abstract)
Xu JL, Zhang ZW, Wang Y, Connelly JW. 2011. Spatio-temporal responses of male Reeves's Pheasants Syrmaticus reevesii to forest edges in the Dabie Mountains, central China. Wildl Biol, 17(1): 16–24.
Xu JL, Zhang ZW, Zheng GM, Zhang XH, Sun QH, McGowan P. 2007. Habitat use of Reeves's Pheasant (Syrmaticus reevesii) in the protected areas created from forest farms in the Dabie Mountains, central China. Bird Conserv Int, 17: 319–330.
Xu WS, Wu ZK, Li ZM. 1991. Reeves's Pheasant (Syrmaticus reevesii). In: Lu TC, Liu RS, He FQ (eds) The Rare and Endangered Gamebirds in China. Fujian Science and Technology Press, Fuzhou, pp 328–338. (in Chinese)
Zhang PC, Shao GF, Zhao G, Le Master DC, Parker GR, Dunning JB Jr, Li QL. 2000. China's Forest Policy for the 21st Century. Science, 288: 2135–2136.
Zhang ZW. 2002. Survey of the habitat fragmentation of Reeves's Pheasant. Unpublished Small Grant Report to WWF-China, p 16.
Zheng GM, Wang QS. 1998. China Red Data Book of Endangered Animals (Aves). Science Press, Beijing. (in Chinese)
Table
2.
The species composition found in the pellets and stomach contents of Malayan Night Herons. Each number shows the number of individuals for each food item. The number in brackets is the percent occurrence of samples containing items for each taxon.


 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds