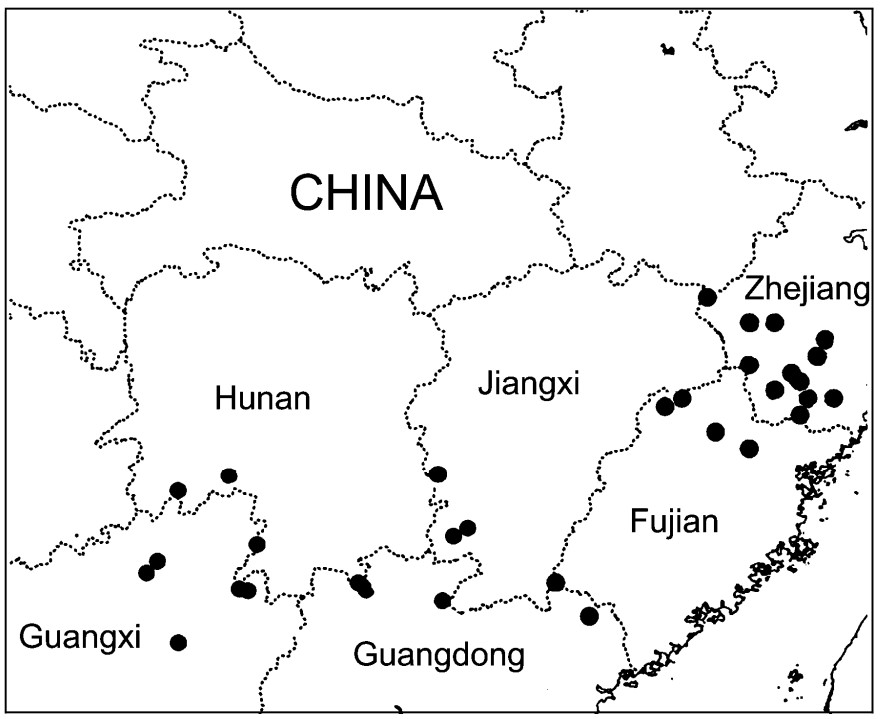
Figure
1.
Distribution areas of Cabot's Tragopan
| Citation: |
Yanyun ZHANG. 2010: Cabot's Tragopan. Avian Research, 1(1): 77-79. DOI: 10.5122/cbirds.2009.0016

|
Cabot's Tragopan (Tragopan caboti) (Galliformes: Phasianidae) is one of the endemic pheasant species in China. It is categorized as Vulnerable in the IUCN Red list (IUCN, 2008) and is also listed in Appendix Ⅰ of CITES (Fuller and Garson, 2000). It is a national first grade protected species in China. Its population size is less than 4000 and this pheasant is only found in several small patches in southeastern China (Zheng and Wang, 1998; Fig. 1).
The typical habitat of Cabot's Tragopan is a broadleaf evergreen and mixed broadleaf coniferous forest at elevations between 800–2000 m (Zhang and Zheng, 2007). Much of its preferred habitat has been destroyed through conversion to agricultural land or for the construction of houses, roads and railways. It is mostly found in forests dominated by Fagaceae, Lauraceae, Theaceae, Aquifoliaceae, Symplocaceae, Rosaceae and Ericaceae families. Habitat use is similar throughout the year and individuals do not move to lower areas during the winter, in contrast with other tragopan species. During the late autumn and winter, habitat use is correlated significantly with the presence of Daphniphyllum macropodum, a tree whose fruits and leaves are an important food for the pheasant during this difficult time. In January 2008, southern China experienced a huge cold snap. Cabot's Tragopan is the most frequently occurring species in the research area (from 700–2000 m), but was not affected by this disaster as were other pheasants, such as the Silver Pheasant and the Chinese Bamboo Partridge, which are found in lower areas.
The home range size varied from 0.029 to 0.39 km2, with a mean of 0.24 km2. The home range of neighboring males always overlaps to some degree (Sun and Zheng, 1992). The furthest distance recorded of an individual moving was less than 3 km. Recent research shows that Cabot's Tragopan cannot adapt well to habitat fragmentation since it hardly passes the mid-level ecological barrier. Because of its weak dispersal capability, habitat fragmentation is the most important factor affecting this pheasant. In recent decades, much of its typical habitat has been destroyed, converted to agricultural use, bamboo forest or separated by anthropogenic activities such as road and railway construction. As a result, the pheasant has been absent from some regions where individuals have been detected in earlier years (Qian and Zheng, 1993).
Cabot's Tragopan built nests in large trees (Fig. 2b), between 2.4 m and 10 m above the ground (mean 4.4±2.1 m), found near ridge tops. Height and diameters at breast height of a tree, nest height, canopy cover, distance to the ridge, direction and slope of the hillside appear to be the main factors affecting nest selection. In the 1980s, all nests discovered were found in broadleaf forests, but since 1990, an increasing number has been detected in coniferous forests (Fig. 2b, c, d). Recent trials with artificial nests in coniferous forests confirm this use of nest sites in coniferous forests, which may be caused by a shortage of sites in broadleaf trees (Deng et al., 2005). Cryptomeria fortunei, a conifer widely planted in the 1980s, may provide adequate nesting sites for the tragopan once the trees reach a certain density or height (Ding and Zheng, 1997).
In the morning during the breeding season, males (Fig. 2a) stand on a large horizontal branch, declaring the territory with loud calls wear, wear, ar… ga-ga-ga. When a rival male appears, the male rushes down and attacks the invader or drives it away with side displays. The male exhibits courtship display by showing his nice horn and colorful lappet. The courtship display is quite different from the side display and is referred to as a type of front display (Zheng, 1987). Courtship lasts from February to May and reaches a peak during March and April. The entire display lasts only 52.15±0.49 s (Zheng et al., 1989). From mid-March, the female begins to lay eggs. The mean clutch size is 3.30±0.22 (from 2 to 6, mainly 3 or 4). The incubation period is about 28 d. Only the female sits on the eggs, while the male is not seen at or around the nest at any stage. The female would leave the nest once a day for 1–2 hours each time. The chicks have longer primaries when they are hatched (Fig. 2e, f) and can fly down to the ground and search for food with the female in two or three days. Since 1983, about 48% of known nests have been destroyed by natural predators such as the Eurasian Jay (Garrulus glandarius), the Yellow-throated Marten (Charronia flavigula) and the Leopard Cat (Felis bengalensis) (Zhang and Zheng, 2007).
On the basis of data obtained over long periods, population viability analysis (PVA) was performed on the Cabot's Tragopan population in southeastern China. The study predicts that there is a 7.3% probability that its population will be extinct in 100 years, with a mean loss of heterozygosity of 14.0%. This analysis also indicates that the nest loss and reduction in the extent of its habitat are the most important factors affecting population size and risk of extinction of the Cabot's Tragopan population in Wuyanling, southwestern China.
The species is divided into two subspecies, caboti and guangxiensis, on the basis of morphological characteristics (Cheng and Wu, 1979). It has been reported that the upper feathers of guangxiensis males are darker and the yellow patches of the feathers lighter than in the caboti subspecies. Guangxiensis birds are only found in northeastern Guangxi, while caboti birds are found in southeastern Hunan, southern Zhejiang, Jiangxi, Fujian and northern Guangdong (Cheng and Wu, 1979; Zheng, 2005).
On the basis of a control region (CR) and the cytochrome b (cyt b) gene of Cabot's Tragopan, a recently completed genetic study showed that this species has a high level of genetic diversity. Its genetic data strongly indicated two phylogenetic clades, which are inconsistent with the current distribution region (Zhang, unpublished data).
I thank professors Guangmei Zheng and Zhengwang Zhang for useful comments and improving the manuscript. I thank all of the doctoral and masters students of Beijing Normal University for their excellent work on Cabot's Tragopan.
|
Cheng TH, Wu MC. 1979. A new subspecies of Tragopan caboti − Tragopan caboti guangxiensis. Acta Ecol Sin, 25(3): 292–294 (in Chinese)
|
|
Ding CQ, Zheng GM. 1997. The nest site selection of the Yellow-bellied Tragopan (Tragopan caboti). Acta Zool Sin, 43(1): 27–33 (in Chinese)
|
|
Fuller RA, Garson PJ. 2000. Pheasants: Status Survey and Conservation Action Plan 2000–2004. IUCN, Gland, Switzerland and Cambridge, UK
|
|
Qian FW, Zheng GM. 1993. The study of habitat used by Cabot's Tragopan. J Beijing Normal Univ Nat Sci, 29(2): 256–264 (in Chinese)
|
|
Sun YH, Zheng GM. 1992. A radiotracking study of home range of Cabot's Tragopan. Acta Zool Sin, 38(4): 385–392 (in Chinese)
|
|
Zheng GM. 1987. Cabot's Tragopan. J Zool, 22(5): 40–43 (in Chinese)
|
|
Zheng GM, Wang QS. 1998. China Red Data Book of Endangered Animals: Aves. Science Press, Beijing, pp 157–159 (in Chinese)
|
|
Zheng GM. 2005. A Checklist on the Classification and Distribution of the Birds of China. Science Press, Beijing, p 57 (in Chinese and English)
|
|
Zheng GM, Yin RL, Zhang ZW, Liu ZX, Zhou HQ. 1989. Behavior of courtship display of Cabot's Tragopan. Acta Zool Sin, 35(3): 328–332 (in Chinese)
|
Figures(2)





 DownLoad:
DownLoad:

 Email Alerts
Email Alerts RSS Feeds
RSS Feeds