Key Laboratory of Biodiversity Conservation of National Forestry and Grassland Administration, Ecology and Nature Conservation Institute, Chinese Academy of Forestry, Beijing, 100091, China
b.
Shaanxi Hanzhong Crested Ibis National Nature Reserve, Hanzhong, 723300, China
Funds:
the Fundamental Research Funds of Chinese Academy of ForestryCAFYBB2020SY023
Moult is an essential event in the annual cycle of birds. Moult patterns and strategies of birds are crucial for understanding their life-history trade-offs and adaptations to the environment. However, moult knowledge is still lacking for many Eastern Palaearctic birds, especially large endangered species, mainly because of the difficulty in data collection. Here, we used field photography, a non-invasive method, to detect and score moult in the endangered Crested Ibis (Nipponia nippon) and explored the moult timing variations and moult-breeding trade-offs. The adult Crested Ibis sequentially moults its primaries outward, and secondaries inward from three foci from April to October. Age (adult versus sub-adult) and breeding status (breeding or not) have a significant effect on moult timing. Moult timing does not differ between the sexes, likely because of similar parental investments in breeding. Crested Ibis exhibits significant moult-breeding overlap, because moult is time constrained and needs to finish before the season of food scarcity in winter. Breeding timing (first egg laying date) has a strong impact on moult duration, with late breeders experiencing a longer moult duration, indicating that the moult intensity is slowed down during the breeding period because of trade-offs between moult and breeding. This reflects that energy allocation of late breeders is challenging during extensive periods of moult-breeding overlap, which might partly contribute to their high breeding failure. Our results strongly suggest the existence of a negative relationship between breeding time on one hand, and fitness and moult on the other: early breeders show higher reproductive output and higher quality plumage, whereas late breeders show lower reproductive output and lower quality plumage.
Moult is commonly defined as the natural, periodic, scheduled shedding, and replacement of feathers, which may involve partial or complete renewal of plumage (Humphrey and Parkes, 1959; Jenni and Winkler, 2020), which restores function of damaged plumage caused by several factors, such as mechanical damage, photochemical processes, and parasites (De La Hera et al., 2010; Terrill and Shultz, 2023). Moult incurs very high costs. This includes the energy expended on the growth of new feathers, as well as the cost of maintaining other functions such as temperature regulation and flight, which are impaired when the plumage is incomplete.
In order to reduce moult costs, birds usually replace their remex one by one so as to reduce moult gap in wing, minimize aerodynamic loss and maintain efficient flight performance. Large species require more time to moult due to their long and larger number of remex. As a result, their secondaries may moult from multiple foci so as to balance moult speed and flight costs (Zuberogoitia et al., 2018). Unlike the moult sequence, which is usually very fixed, moult timing is generally more flexible in response to external factors. Some biological factors, such as age and sex, influence the moult progress of birds (Nadal et al., 2021; Wolfe et al., 2021). For example, the Bearded Vulture (Gypaetus barbatus) needs 2–3 years to moult all of its flight feathers, and the timing of remex moult is different across age classes mainly due to their distinct breeding requirements (Zuberogoitia et al., 2013). In addition, the impact of sex on moult timing is often related to their parental investment in breeding. For example, females usually start moult at an early breeding stage to remain their plumage in good condition, thereby facilitating the incubation of eggs and chick rearing (Jenni and Winkler, 2020). Thus, moult sequence and timing are important aspects of birds' life history and have significant implications for their survival, reproduction, and overall fitness.
Since moult and breeding are both energy-costly processes, birds must adequately allocate time and resources between these two crucial life-history events to maximize their lifetime fitness (Jenni and Winkler, 2020). For small birds, moult and breeding are often temporally separated. However, moult-breeding overlap is common in many medium-sized and large altricial bird species, mainly due to time constraints resulted from both long breeding and moult periods (Rohwer, 1999; Zuberogoitia et al., 2018). In some cases, moult-breeding overlap is also because of abundant food availability during breeding season but food scarcity in non-breeding period, and birds may benefit from this overlap (Repenning and Fontana, 2016; Jahn et al., 2017; Tomotani et al., 2018; Kiat and Bloch, 2023). For example, the Hispaniolan Woodpecker (Melanerpes striatus) moults slowly during the breeding season when food is plentiful to minimize post-breeding food shortages, allowing moult and breeding processes to overlap (Garrod and LaPergola, 2018). Thus, moult strategies represent different adaptive outcomes which are key for life-history trade-offs.
The method of data acquisition is a pivotal aspect of moult research, and little change has occurred since the last century (Jenni and Winkler, 2020). The predominant approach involves investigating museum specimens and dead specimens killed accidently, or capturing wild birds in mist nets, particularly when studying passerines (Jenni and Winkler, 2020; Ryan and Adekola, 2023). For large birds, the collection of moult data can be challenging and invasive. To overcome the limitations of traditional methods, field photography, as a non-invasive approach, can provide new insights and data sources for moult research involving elusive, inaccessible, and endangered bird species (Osborne and Ryan, 2021). Moreover, field photography represents a convenient method for reliably and remotely studying moult. Its results are comparable to those obtained through the direct observation of birds (Vieira et al., 2017). This method can provide detailed information on the feathers of birds in wild populations over prolonged periods, especially when the birds are flying in the air (Keijl, 2011; Ramírez and Panuccio, 2019).
The Crested Ibis (Nipponia nippon) is classified as an endangered species (EN) by the IUCN (BirdLife International, 2024) and is also listed as a National First-Class Key Protected Animal in China. Currently, studies of the plumage of the Crested Ibis are limited to colour changes during the breeding season and the timing and sequence of post-juvenile moult in captivity (Li and Gao, 1989; Wingfield et al., 2010; Sun et al., 2020; Liu et al., 2023). However, data on post-breeding moult and the corresponding influential factors are scarce. Since Crested Ibis is a medium-sized altricial bird species with a relatively long breeding period, and food shortage in winter is a crucial limiting factor for population growth (Ding, 2004), we speculate that there is a significant moult-breeding overlap due to both time constraints and restricted period of abundant food, which could have management applications.
In this study, we used field photography to study the moult of a captive Crested Ibis population kept in an acclimation aviary in Central China, with the aim to: (1) determine the remex-moult sequence and understand the moult pattern for subsequent analysis; (2) assess the influence of age, sex, and breeding status (breeding or not) on moult timing; (3) quantify the moult-breeding overlap, providing data basis for the trade-offs evaluation; and (4) evaluate the effect of breeding timing (early versus late breeders) on moult strategy (moult start date and moult speed), so as to understand the moult-breeding trade-offs and furnish implications for the management and conservation efforts.
2.
Materials and methods
2.1
Study area
This study was conducted at the Crested Ibis Breeding and Rewilding Seed Source Base (33.5954° N, 107.5358° E) situated in Huayang, Yangxian County, Shaanxi Province of China. An acclimation aviary of approximately 34, 000 m2 in area and 30–40 m in height was designed to hold a semi-free-ranging Crested Ibis population (Fig. 1); the enclosure was made of nylon net (mesh size: 70 mm × 70 mm) held up by several central and peripheral columns. In the acclimation aviary, there were various habitats, such as trees, wetlands, and grasslands, that enable the imitation of real wildlife monitoring situations to facilitate the acclimatization of the ibises to wild environments before release. Loaches (Misgurnus anguillicaudatus) (450 g per individual) were supplemented in the wetlands in the aviary on a daily basis to ensure sufficient food.
Figure
1.
General overview of the acclimation aviary (left) and the internal environment (right).
In total, 74 Crested Ibises, namely, 60 adults (≥3rd calendar year) and 14 sub-adults (2nd calendar year), were kept in the aviary beginning in early 2023. All individuals were colour-marked with unique numbers, which allowed for continuous monitoring of the moult progress of individuals, and enabled accurate assessment of the moult timing. Furthermore, it enabled precise recording of information regarding the age (n = 65), sex (n = 37), and breeding status (n = 52) of individuals, facilitating a fine-scale assessment of the trade-offs associated with different moult strategies.
2.2
Moult data collection and quantitative evaluation of moult scores
From April to October 2023, we used a camera for random sampling in the acclimation aviary to take clear images of birds in flight during moult. Only one valid image of each individual on each date was selected. Simultaneously, we identified the ring number of each flying individual via a camera or telescope. Moult stage and bird identity (based on the ring number of the individual) were recorded from photographs of birds in flight. Only images clear enough to assess moult scores and identify birds were used for analyses.
We scored growth of each primary using a standard system (Ginn and Melville, 1983): 0 for old feathers, 5 for new feathers, and 1–4 for the intermediate stages of development of the growing feathers. The scores were generally symmetrical between the two wings, so only one wing was analysed. The scores for each individual were combined to establish a summary table of feather replacement, which was used to determine the moult start date and, hence, the pattern of feather replacement (Rohwer et al., 2011).
2.3
Bird identity information and breeding parameters
All individuals in the aviary were tagged with unique numbered ring 20–25 days after hatching. Regular observations were conducted from March to July to determine the breeding information daily, including the ring number of the breeding pair, the laying date of the first egg, the brood size, and the number of fledglings. This enabled us to accurately determine the breeding timing and the reproductive success. Sexing was based on observations of copulation behaviour and egg-laying, a method that has been widely applied for Crested Ibis (Ding, 2004; Dong et al., 2018).
2.4
Statistical analyses
We obtained image documenting the entire moult cycle of primaries for identifiable individuals, enabling us to assess the sequence of remex replacement on the basis of moult scores recorded at different times. To study moult sequence, we used a Bayesian linear regression model (value ~ 1, which estimates the mean value of the dependent variable, i.e., the average pattern of the feather replacement sequence for all individuals) (Bürkner, 2017; Guallar and Quesada, 2023; Guallar, 2024) in the "brms" package in R to analyse the moult scores data. Model convergence was assessed on the basis of the Rhat value (with an ideal value of 1) and the effective sample size.
To assess moult timing (including the moult start date and moult duration) and the influence of age (adults versus sub-adults), sex (breeding male versus breeding female), and breeding status (breeding versus non-breeding), we used the Underhill–Zucchini moult model implemented in the "moult" package (moult.index ~ days) in R (Underhill and Zucchini, 1988; Erni et al., 2013). Two age groups were included in the analysis, namely adult and sub-adult. Adults refer to individuals that are in their third year or older, with full adult plumage; sub-adults refer to individuals that are in their second year, usually with juvenile remiges and rectrices, and adult body and wing coverts. Sex indicates whether a breeding bird is male or female. Breeding status indicates whether an adult bird breeds. We employed the moult index (the ratio of the sum of moult scores at any time to the total moult score of all feathers, with a value between 0 and 1) as the response variable, which requires a vector of corresponding days on which selected individuals were observed (e.g., individuals with known age, sex, and breeding status). We used the discrepancy coefficient generated by the model to reflect the variance in the dataset between groups. We used a t-test analysis to determine whether the regression coefficient was significant.
To quantify moult-breeding overlap, we compared the moult period of paired individuals with their breeding period. We used Underhill–Zucchini moult model to determine the moult timing of paired individuals based on their moult index. The breeding period is defined as the date from the laying of the first egg to the fledging of the last nestling from the nest.
To evaluate the effect of breeding timing (early versus late breeders) on moult strategy, we used a generalized linear model with moult start date and moult duration as the response variable and first egg laying date as predictor. The linear regression model is as follows:
moult start date ∼β0+β1× first egg laying date +ϵ
where β0 represents the baseline value of moult start date when the first egg laying date is zero, β1 reflects the corresponding average change in moult start date for each one-unit increase in first egg laying date (e.g., days), and ϵ represents the unexplained random error in the model.
All the analyses were performed in R version 4.3.2 (R Core Team, 2023). Statistics are reported as the mean ± SD.
3.
Results
A total of 1020 valid images of flight feathers from 65 individuals were recorded (15.8 ± 10.7 images per individual, on average), with 52 adults and 13 sub-adults. Among the 52 adults, 37 breeding individuals and 15 non-breeding individuals were included.
3.1
Moult sequence
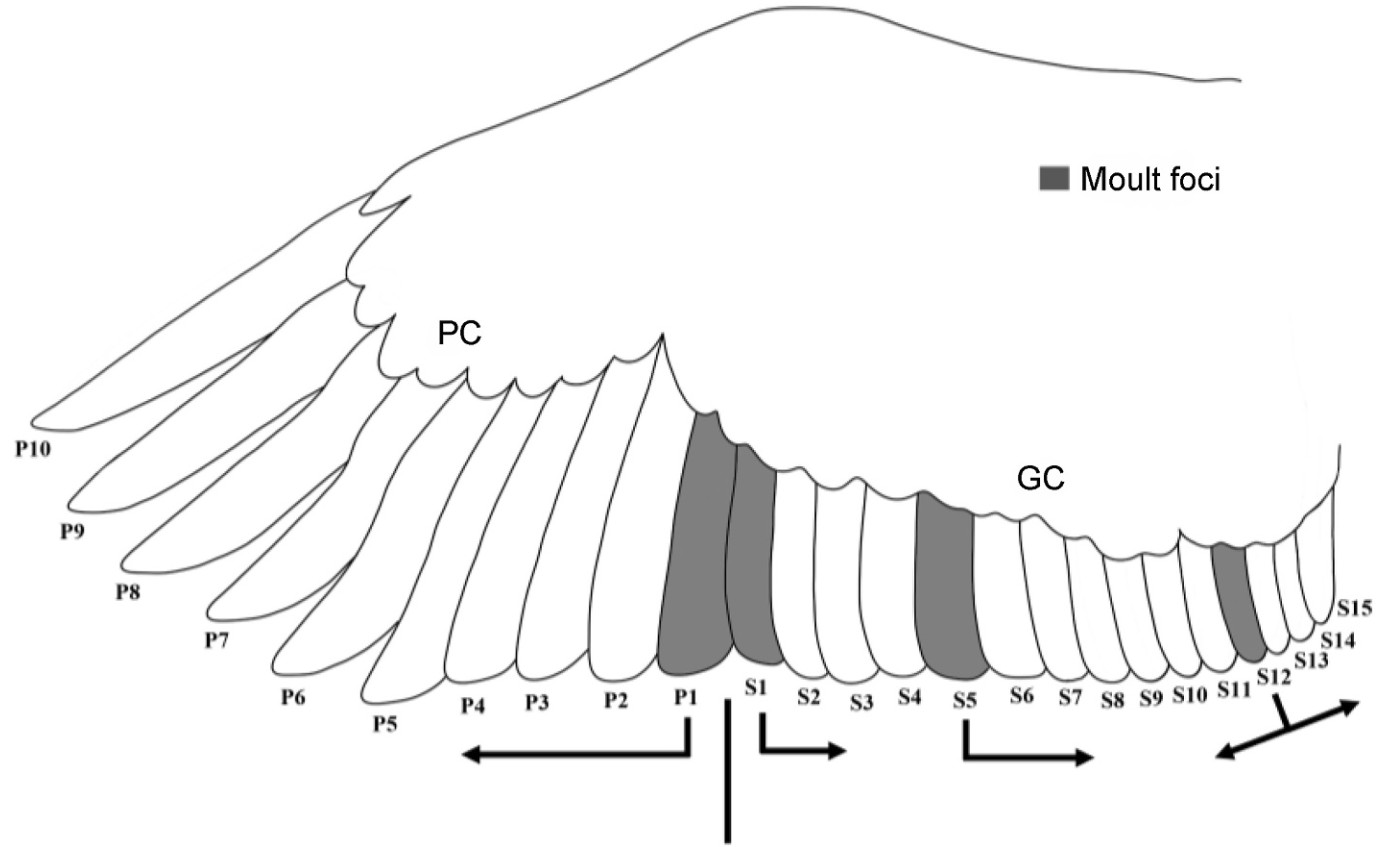
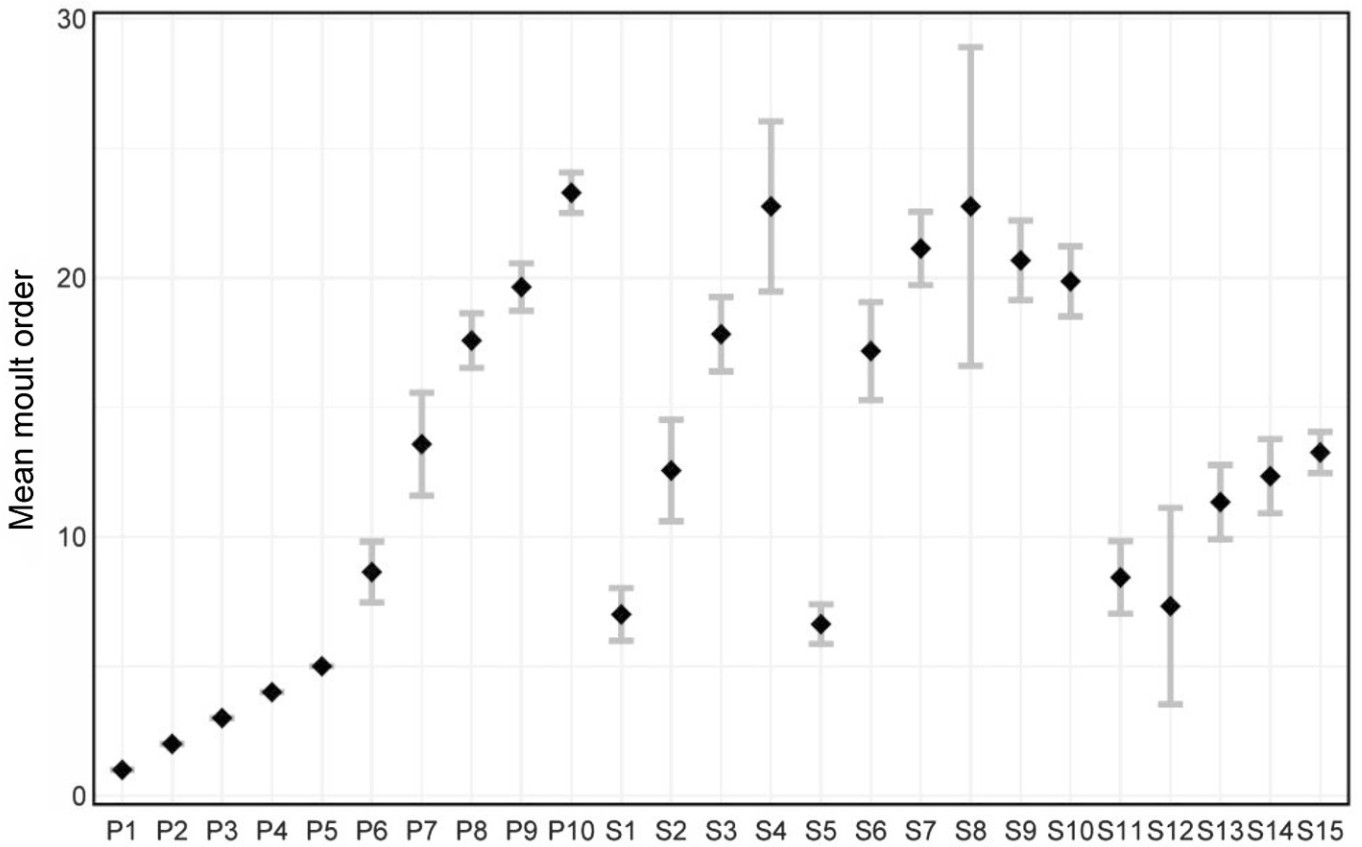
The Crested Ibis has 25 remiges, with 10 primaries and usually 15 secondaries. The Crested Ibis replaces primaries outward from P1. Secondaries tend to show three moult foci almost simultaneously, S1, S5, and S12. The feathers are replaced inward from S1 to S5 and divergently from S12 (Fig. 2). Secondary moult starts when P5 is in an advanced stage of growth or P6 begins growing (n = 60, Rhat = 1.0). Compared with P1–P5 and S13–S15, the other remiges showed much greater moult-sequence variation. P10 is usually the last feather to be moulted (Fig. 3).
Figure
2.
Moult foci in primaries (P1) and secondaries (S1, S5, and S12) for the Crested Ibis.
Figure
3.
Moult sequence (±95% CIs) for the primaries and secondaries of the Crested Ibis. The black dots represent the mean order of average moult activation. Wider grey bars indicate greater variation within the sequence based on confidence intervals.
3.2
Moult timing and the effects of age, sex, and breeding status
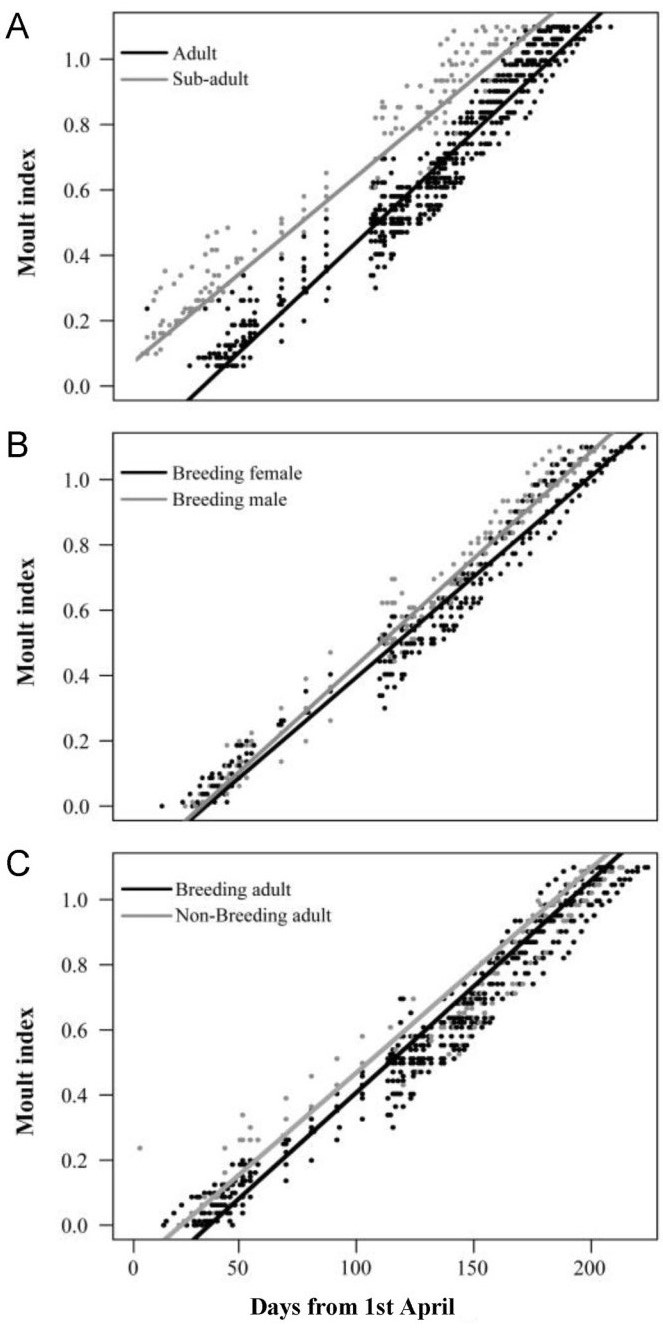
Adult Crested Ibises started remex moult in late April (Table 1). In contrast, sub-adults started moult much earlier (P < 0.001) and has a significantly longer duration (P < 0.001, Table 1 and Table 2; Fig. 4). Although breeding females started moult earlier and had a shorter moult duration compared to breeding males, these differences were not significant. However, breeding status had a significant effect on moult timing, and non-breeding adults moulted earlier (P < 0.001) and for a shorter duration (P < 0.001) than did breeding adults (Table 2).
Table
1.
Moult start date and duration of Crested Ibis with respect to age, sex, and breeding status.
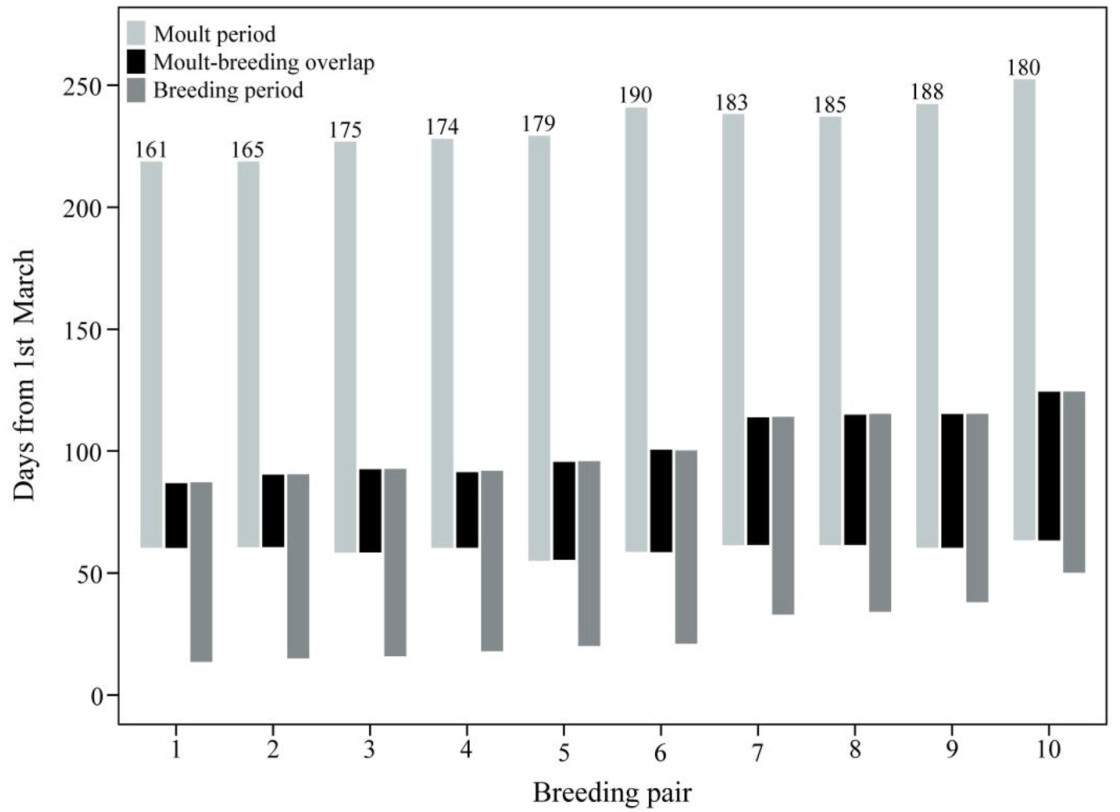
Crested Ibis exhibited significant moult-breeding overlap, averaging 23.0 ± 6.7% of the total moult duration (ranging from 13.6% to 34.1%, n = 10) and 55.5 ± 16.1% of the total breeding period (ranging from 32.9% to 82.2%, n = 10). As the first egg laying date delayed, the moult-breeding overlap gradually increased, from 24 days to 60 days (Fig. 5). For early breeders (breeding pair 1 in Fig. 5), primary moult started when the first nestling was 20 days old, and P1–P2 were under moulting by the end of the breeding period; for late breeders (breeding pair 10 in Fig. 5), primary moult started on the 13th day after the first egg hatched, and P3–P4 were under moulting by the end of the breeding period.
Figure
5.
Relationship between moult duration and moult-breeding overlap. Late breeders had greater moult-breeding overlap, resulting in a longer moult duration. The numbers above bars indicate the moult duration in days.
The linear regression model failed to effectively explain the variability of the moult start date, and the fit was poor (R2 = 0.013), indicating that the first egg laying date had no significant effect on the moult start date in the Crested Ibis population (P = 0.755; Table 3). In contrast, the first egg laying date had a strong effect on moult duration (R2 = 0.495, P = 0.023), with late breeders experiencing a longer moult duration (Fig. 5). Specifically, a one-day delay in breeding resulted in an approximately 0.659 days extension of the moult duration (Table 3).
Table
3.
Effects of the breeding start date (first egg laying date) on moult start date and moult duration (n = 10).
In the Crested Ibis, P1–P5 are moulted independently at the onset of the remex moult sequence, and then secondary moult follows. The staggered replacement of primary and secondary is extremely widespread, which avoid big moult gap in the wing at the very beginning of moult, a strategy likely aimed at minimizing aerodynamic loss and maintaining efficient flight performance (Tucker, 1991; Hedenström and Sunada, 1999; Guallar and Quesada, 2023; Guallar, 2024).
Although we do not have the body moult intensity data, the overlapping moult of secondaries and primaries P6–P10 increases overall wing-feather mass production as well as raggedness (Schmidt-Nielsen, 1984; Hedenström, 1998). This helps maintain flight capacity by balancing feather production with the maximum power available to flight muscles while addressing the time constraints that large birds face to prevent simultaneous moult of many feathers. Moreover, the replacement of secondaries aligns with the end of the breeding period based on our results, allowing the Crested Ibis to allocate more energy resources to moult once breeding is over.
In Crested Ibis, secondaries are moulted from multiple foci. This moult pattern has been documented in many large species, as well as the family Threskionithidae (Zuberogoitia et al., 2018; Heath et al., 2020; Ryan and Adekola, 2023). By dividing moult into different series, several feathers can moult concurrently while separated by non-moulting ones. Therefore, multiple small gaps are produced, in contrast to a normal sequential moult that has only one gap (Rohwer, 1999; Rohwer and Broms, 2013). The multiple small wing-gaps are likely to reduce any aerodynamic disadvantage as they can be covered by closing the wing slightly, thereby maintaining a complete aerofoil (Jenni and Winkler, 2020).
4.2
Moult timing in relation to age and sex
Our results indicate that the moult of sub-adults starts much earlier and lasts for a longer time. This is primarily due to the fact that sub-adults do not partake in breeding activities. Consequently, they are able to allocate surplus energy and thus have more time to complete the moult. As a result, sub-adults typically extend the moult duration by decreasing the moult intensity, so as to reduce the overall aerodynamic costs throughout the moult period (Jenni and Winkler, 2020).
In the Crested Ibis, moult timing exhibits no sexual dimorphism, thus providing additional evidence that sex-related moult timing differences may be linked to disparities in parental investment during breeding (Weimerskirch, 1991; Svensson and Nilsson, 1997). For example, in many raptors, males moult later than females do because males provide most of the food for female and young nestlings during the incubation and brooding periods (Zuberogoitia et al., 2018; Adekola et al., 2021). In some species with biparental care, such as swans and geese, male typically delay his simultaneous remex-moult strategically during the chick-rearing period, thereby balancing parental responsibilities with the energetic demands of plumage renewal (McCleery et al., 2007).
4.3
Moult-breeding overlap
The Crested Ibis typically starts moult during the nestling-rearing stage, resulting in substantial moult-breeding overlap that comprises more than half of the total breeding period (from first egg laying to the fledging of the last nestling). Similar moult-breeding overlap has also been observed in other Threskionithidae species, such as the Sacred Ibis (Threskiornis aethiopicus) and White Ibis (Eudocimus albus) (Low et al., 1985; Heath et al., 2020). Previous studies have indicated that food shortage in winter is the primary cause of mortality for Crested Ibis (Ding, 2004). As a result, the moult-breeding overlap represents an adaptive strategy to deal with the long moult duration, which must to be completed prior to the harsh season of winter.
In larger altricial species that breed in a seasonal environment, moulting commonly commences during the egg-laying or incubation stage (Zuberogoitia et al., 2018; Jenni and Winkler, 2020). Interestingly, the Crested Ibis, on the other hand, starts moult later, during the nestling-rearing phase. We propose that this is due to the fact that the Crested Ibis is a resident bird and thus has relatively more time to balance the processes of moult and preparation of breeding.
4.4
Effect of breeding timing on moult strategy
Our study reveals that in the Crested Ibis, breeding timing, as indicated by first egg laying date, has a significant effect on moult duration. Late breeders exhibit more extensive moult-breeding overlap and experience a longer moult duration. This phenomenon can be attributed to fact that moult is slowed down during the breeding period as a result of the trade-off between moult and breeding. This finding is further supported by the fact that non-breeding adults experience a significantly shorter moult duration compared to breeders. Similar results have also been documented for the male Southern Giant Petrel (Macronectes giganteus) and potentially other birds that regularly commence moult during the early stages of breeding (Osborne and Ryan, 2021), in which moult is significantly slowed or suspended when the nestlings' food demand is greatest during the breeding season (Cieślak and Kwieciński, 2009).
Previous studies have indicated that late breeders of wild Crested Ibis typically exhibit lower productivity or even fail to breed, possibly because of decreased food abundance and higher temperatures compared with early breeders (Ding, 2004; Liu et al., 2020). However, the productivity of late breeders in captivity was much greater than that under natural conditions because enough food was supplemented in this study. This further indicates that food resources may be a limiting factor, as late breeders face significant challenges in energy allocation during the extensive overlapping period of moult and breeding. Improvements in food conditions apparently increase the productivity of late-breeding Crested Ibis in captive reproduction programs. Therefore, our data strongly suggest the existence of a negative relationship between breeding time on one hand, and fitness and moult on the other: early breeders show higher reproductive output and higher quality plumage, whereas late breeders show lower reproductive output and lower quality plumage.
CRediT authorship contribution statement
Jia Li: Writing – review & editing, Writing – original draft, Methodology, Data curation. Dongping Liu: Supervision, Software, Conceptualization. Wei Li: Data curation. Wenbin Duan: Conceptualization. Jie Gao: Data curation. Chao Wang: Data curation.
Declaration of competing interest
All authors have read and approved this version of the article, and due care has been taken to ensure the integrity of the work. The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We thank Ming Zhang, Hongxin Su, Wenming Ren, and Ying Li for their help in data collection and logistical support. We thank two reviewers for their constructive feedback and support during the process, which significantly improved the manuscript.
Adekola, O.E., Allan, D.G., Bernitz, Z., Dlungwana, W., Ryan, P.G., 2021. Extent and symmetry of tail moult in Amur Falcons. J. Ornithol. 162, 655-667.
BirdLife International, 2024. Species factsheet: Nipponia nippon. . (Accessed 28 May 2024).
Bürkner, P.C., 2017. brms: an R package for Bayesian multilevel models using stan. J. Stat. Softw. 80, 1-28.
Cieślak, M., Kwieciński, Z., 2009. Moult and breeding of captive northern hawk owls Surnia ulula. Ardea 97, 571-579.
De La Hera, I., Pérez-Tris, J., Tellería, J.L., 2010. Relationships among timing of moult, moult duration and feather mass in long-distance migratory passerines. J. Avian Biol. 41, 609-614.
Ding, C., 2004. Research on the Crested Ibis. Shanghai Scientific and Technical Publishers, Shanghai.
Dong, R., Ye, X., Lin, Z., Li, X., Li, M., Wang, H., et al., 2018. Effects of breeding success, age and sex on breeding dispersal of a reintroduced population of the Crested Ibis (Nipponia nippon) in Ningshan County, China. Avian Res. 9, 40.
Erni, B., Bonnevie, B.T., Oschadleus, H., Altwegg, R., Underhill, L.G., 2013. moult: an R package to analyse moult in Birds. J. Stat Softw. 52, 123.
Garrod, H.M., LaPergola, J.B., 2018. Overlap in molt and breeding phenology in the hispaniolan wood pecker (Melanerpes striatus). Ornitol. Neotrop. 29, S29-S36.
Ginn, H., Melville, D.S., 1983. Moult in Birds. British Trust for Ornithology, Tring, UK.
Guallar, S., 2024. From partial to complete: wing- and tail-feather moult sequence and intensity depend on species, life-cycle stage, and moult completeness in passerines. Avian Res. 15, 100163.
Guallar, S., Quesada, J., 2023. Moult intensity constraints along the complete moult sequence of the House Sparrow (Passer domesticus). Avian Res. 14, 100125.
Heath, J.A., Frederick, P.C., Kushlan, J.A., Bildstein, K.L., 2020. White Ibis (Eudocimus albus), version 1.0. In: Poole, A.F. (Ed.), Birds of the World. Cornell Lab of Ornithology, Ithaca, NY, USA. .
Hedenström, A., 1998. The relationship between wing area and raggedness during molt in the willow warbler and other passerines. J. Field Ornithol. 69, 103-108.
Hedenström, A., Sunada, S., 1999. On the aerodynamics of moult gaps in birds. J. Exp. Biol. 202, 67-76.
Humphrey, P.S., Parkes, K.C., 1959. An approach to the study of molts and plumages. Auk 76, 1-31.
Jahn, A.E., Bejarano, V., Guzmán, M.B., Brown, L.M., Provinciato, I.C.C., Cereghetti, J., et al., 2017. Molting while breeding? Lessons from new world Tyrannus flycatchers. J. Ornithol. 158, 1061-1072.
Jenni, L., Winkler, R., 2020. The Biology of Moult in Birds. Bloomsbury Publishing Plc, London.
Keijl, G.O., 2011. Sooty shearwaters Puffinus griseus in the North Atlantic – moult studies using digital cameras. Mar. Ornithol. 39, 141-142.
Kiat, Y., Bloch, I., 2023. The relationship of moult timing, duration and sequence to the aerial lifestyle of the Little Swift (Apus affinis). Ibis 165, 1331-1342.
Li, F., Gao, X., 1989. A study of postnatal moult in the Crested Ibis. Wildlife (Lond.) 5, 21-22.
Liu, D., Tong, Y., Dong, R., Ye, X., 2023. A breeding plumage in the making: the unique process of plumage coloration in the Crested Ibis in terms of chemical composition and sex hormones. Animals 13, 3820.
Liu, D., Zhang, G., Wang, C., Qing, B., Lu, J., 2020. Breeding variation in a reintroduced crested ibis Nipponia nippon population in Central China. Pakistan J. Zool. 52, 1595-1598.
Low, K.W., Clark, A., Clark, R.A., 1985. Body measurements, plumage and moult of the sacred ibis in South Africa. Ostrich 56, 111-116.
McCleery, R.H., Perrins, C.M., Wheeler, D., Groves, S., 2007. The effect of breeding status on the timing of moult in Mute Swans Cygnus olor. Ibis 149, 86-90.
Nadal, J., Ponz, C., Margalida, A., 2021. The end of primary moult as an indicator of global warming effects in the Red-legged Partridge Alectoris rufa, a medium sized, sedentary species. Ecol. Ind. 122, 107287.
Osborne, A., Ryan, P.G., 2021. Using digital photography to study moult extent in breeding seabirds. Ostrich 92, 225-228.
R Core Team, 2023. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. .
RamírezJuan, R.J., PanuccioMichele, M.P., 2019. Flight feather moult in western marsh harriers during autumn migration. Avian Res. 10, 7.
Repenning, M., Fontana, C.S., 2016. Seasonality of breeding, moult and fat deposition of birds in subtropical lowlands of southern Brazil. Emu 111, 268-280.
Rohwer, S., 1999. Time constraints and moult-breeding tradeoffs in large birds. In: Adams, N.J., Slotow, R.H. (Eds.), Proceedings of the International Ornithological Congress, Durban, South Africa, 16-22 August 1998, pp. 568-581.
Rohwer, S., Broms, K., 2013. Replacement rules for the flight feathers of yellow-billed cuckoos (Coccyzus americanus) and common cuckoos (Cuculus canorus). Auk 130, 599-608.
Rohwer, S., Viggiano, A., Marzluff, J.M., 2011. Reciprocal trade-offs between molt and breeding in albatrosses. Condor 113, 61-71.
Ryan, P.R., Adekola, O.E., 2023. Pattern and intensity of moult in White-capped/Shy Albatrosses Thalassarche steadi/cauta. Ostrich 92, 292-303.
Schmidt-Nielsen, K., 1984. Scaling: Why is Animal Size so Important? Cambridge University Press, New York.
Svensson, E., Nilsson, J.A., 1997. The trade-off between moult and parental care: a sexual conflict in the blue tit? Behav. Ecol. 8, 92-98.
Terrill, R.S., Shultz, A., 2023. Feather function and the evolution of birds. Biol. Rev. 98, 540-566.
Tomotani, B.M., Muijres, F.T., Koelman, J., Casagrande, S., Visser, M.E., 2018. Simulated moult reduces flight performance but overlap with breeding does not affect breeding success in a long-distance migrant. Funct. Ecol. 32, 389-401.
Tucker, V.A., 1991. The effect of molting on the gliding performance of a harris' hawk (Parabuteo unicinctus). Auk 108, 108-113. .
Underhill, L.G., Zucchini, W., 1988. A model for avian primary moult. Ibis 130, 358-372.
Vieira, B.P., Furness, R.W., Nager, R.G., 2017. Using field photography to study avian moult. Ibis 159, 443-448.
Weimerskirch, H., 1991. Sex-specific differences in moult strategy in relation to breeding in the wandering albatross. Condor 93, 731-737.
Wingfield, J.C., Ishii, S., Kikuchi, M., Wakabayashi, S., Chikatsuji, K., 2010. Biology of a critically endangered species, the toki (Japanese crested ibis) Nipponia nippon. Ibis 142, 1-11.
Wolfe, J.D., Terrill, R.S., Johnson, E.I., Powell, L.L., Ryder, B.T., 2021. Ecological and evolutionary significance of molt in lowland Neotropical landbirds. Ornithology 138, ukaa073.
Zuberogoitia, I., De la Puente, J., Elorriaga, J., Alonso, R., Palomares, L.E., Martínez, J.E., 2013. The flight feathers molt of Griffon Vultures Gyps fulvus and associated biological consequences. J. Rap. Res. 47, 292-303.
Zuberogoitia, I., Zabala, J., Martínez, J.E., 2018. Moult in birds of prey: a review of current knowledge and future challenges for research. ARDEOLA 65, 183-207.


 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds