Aguilar, P., Andrade, P., de lanuza, G.P.I., 2022. Epistatic interactions between pterin and carotenoid genes modulate intra-morph color variation in a lizard. Integr. Zool. 17, 44–53. https://doi.org/10.1111/1749-4877.12570.
|
Amat, J.A., Garrido, A., Portavia, F., Rendón-Martos, M., Pérez-Gálvez, A., Garrido-Fernández, J., et al., 2018. Dynamic signalling using cosmetics may explain the reversed sexual dichromatism in the monogamous greater flamingo. Behav. Ecol. Sociobiol. 72, 135. .
|
Amundsen, T., 2000. Why are female birds ornamented? Trends Ecol. Evol. 15, 149–155. https://doi.org/10.1016/S0169-5347(99)01800–5.
|
|
Anderson, M.J., 2005. Permutational multivariate analysis of variance. J. Geosci. Environ. Protect. 26, 32–46.
|
Anderson, M.J., Ellingsen, K.E., McArdle, B.H., 2006. Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 9, 683–693. .
|
Andersson, S., 1999. Morphology of UV reflectance in a whistling-thrush: implications for the study of structural colour signalling in birds. J. Avian Biol. 30, 193–204. .
|
|
Andersson, S., Prager, M., 2006. Quantifying colors. In: Hill, G.E., McGraw, K.J. (Eds. ), Bird Coloration, Volume 1: Mechanisms and Measurements. Harvard University Press, Cambridge, pp. 41–89.
|
|
Araújo-Silva, B.M., 2021. Coloração da plumagem, agressividade e dominância social em Sicalis flaveola brasiliensis. Doctoral Thesis. Universidade Vila Velha, Vila Velha.
|
Araújo-Silva, B., Barcellos, M., Duca, C., Diniz, P., 2022. Delayed plumage signals social status in a mutually ornamented bird. J. Ornithol. 164, 417–431. .
|
Barreira, A.S., García, N.C., Lougheed, S.C., Tubaro, P.L., 2016. Viewing geometry affects sexual dichromatism and conspicuousness of noniridescent plumage of Swallow Tanagers ( Tersina viridis). Auk 133, 530–543. .
|
|
Bastien, P., Tenenhaus, M., 2001. PLS generalised linear regression. Application to the analysis of life time data. In: Proceedings of the PLS'01 International Symposium, Paris.
|
Bastien, P., Vinzi, V.E., Tenenhaus, M., 2005. PLS generalised linear regression. Comput. Stat. Data Anal. 48, 17–46. .
|
Benítez Saldivar, M.J., Massoni, V., 2018. Lack of conspecific visual discrimination between second-year males and females in the Saffron Finch. PLoS One 13, e0209549. .
|
Benitez Saldivar, M.J., Miño, C.I., Massoni, V., 2019. Genetic mating system, population genetics and effective size of Saffron Finches breeding in southern South America. Genetica 147, 315–326. .
|
Bergeron, Z.T., Fuller, R.C., 2018. Using human vision to detect variation in avian coloration: how bad is it? Am. Nat. 191, 269–276. .
|
|
Bertrand, F., Meyer, N., Maumy-Bertrand, M., 2014. Partial least squares regression for generalized linear models. R package version.
|
Bleiweiss, R., 2004. Novel chromatic and structural biomarkers of diet in carotenoid-bearing plumage. Proc. R. Soc. B 271, 2327–2335. doi. org/10.1098/rspb. 2004.2868.
|
Bleiweiss, R., 2005. Variation in ultraviolet reflectance by carotenoid-bearing feathers of tanagers (Thraupini: Emberizinae: Passeriformes). Biol. J. Linn. Soc. 84, 243–257. https://doi.org/10.1111/j.1095-8312.2005.00427.x.
|
Bridge, E.S., Hylton, J., Eaton, M.D., Gamble, L., Schoech, S.J., 2008. Cryptic plumage signaling in Aphelocoma scrub-jays. J. Ornithol. 149, 123–130. .
|
Brush, A.H., 1990. Metabolism of carotenoid pigments in birds. Faseb. J. 4, 2969–2977. .
|
Burnham, H., Cruz-Bernate, L., 2020. Parental investment does not directly affect reproductive success in the saffron finch. J. Avian Biol. 51, e02489. .
|
Burns, K.J., 1998. A phylogenetic perspective on the evolution of sexual dichromatism in tanagers (Thraupidae): the role of female versus male plumage. Evolution 52, 1219–1224. .
|
Burns, K.J., Shultz, A.J., 2012. Widespread cryptic dichromatism and ultraviolet reflectance in the largest radiation of Neotropical songbirds: implications of accounting for avian vision in the study of plumage evolution. Auk 129, 211–221. .
|
Butler, M.W., Toomey, M.B., McGraw, K.J., 2011. How many color metrics do we need? Evaluating how different color-scoring procedures explain carotenoid pigment content in avian bare-part and plumage ornaments. Behav. Ecol. Sociobiol. 65, 401–413. .
|
Casalía, B., Vilacoba, E., Lavinia, P.D., Tubaro, P.L., Barreira, A.S., 2020. UV sensitive vision in cardinals and tanagers is ubiquitous. Emu 120, 355–359. .
|
Clements, J.F., Schulenberg, T.S., Iliff, M.J., Billerman, S.M., Fredericks, T.A., Gerbracht, A., et al., 2021. The eBird/Clements Checklist of Birds of the World: v2021. .
|
|
Cuthill, I.C., 2006. Analyzing colors. In: Hill, G.E., McGraw, K.J. (Eds. ), Bird Coloration, Volume 1: Mechanisms and Measurements. Harvard University Press, Cambridge, pp. 3–40.
|
Cuthill, I.C., Partridge, J.C., Bennett, A.T., Church, S.C., Hart, N.S., Hunt, S., 2000. Ultraviolet vision in birds. Adv. Stud. Behav. 29, 159–214. https://doi.org/10.1016/S0065-3454(08)60105–9.
|
Dale, J., Dey, C.J., Delhey, K., Kempenaers, B., Valcu, M., 2015. The effects of life history and sexual selection on male and female plumage colouration. Nature 527, 367–370. .
|
Darwin, C., 1871. The Descent of Man, and Selection in Relation to Sex. Cambridge University Press, Cambridge. .
|
|
de Smith, M.J., 2018. Statistical Analysis Handbook. The Winchelsea Press, Drumlin Publications, Drumlin Security Ltd, UK.
|
Delestrade, A., 2001. Sexual size dimorphism and positive assortative mating in Alpine Choughs ( Pyrrhocorax graculus). Auk 118, 553–556. .
|
|
Delhey, K., 2005. Sexual Selection and Blue Tit (Parus caeruleus) Crown Coloration. Doctoral Thesis. Ludwig-Maximilians-Universität, München.
|
Delhey, K., Szecsenyi, B., Nakagawa, S., Peters, A., 2017. Conspicuous plumage colours are highly variable. Proc. R. Soc. B 284, 20162593. .
|
Demko, A.D., Sosa-López, J.R., Simpson, R.K., Doucet, S.M., Mennill, D.J., 2020. Divergence in plumage, voice, and morphology indicates speciation in Rufous-capped Warblers ( Basileuterus rufifrons). Auk 137, ukaa029. .
|
Diniz, P., Ribeiro, P.H., Rech, G.S., Macedo, R.H., 2016. Monochromatism, cryptic sexual dimorphism and lack of assortative mating in the Rufous Hornero, Furnarius rufus albogularis. Emu 116, 294–300. .
|
Eaton, M.D., 2005. Human vision fails to distinguish widespread sexual dichromatism among sexually "monochromatic" birds. Proc. Natl. Acad. Sci. USA 102, 10942–10946. .
|
Eaton, M.D., 2007. Avian visual perspective on plumage coloration confirms rarity of sexually monochromatic North American passerines. Auk 124, 155–161. .
|
Ellegren, H., 1996. First gene on the avian W chromosome (CHD) provides a tag for universal sexing of non-ratite birds. Proc. R. Soc. B 263, 1635–1641. .
|
|
Espinal, L.S., 1967. Visión ecológica del departamento del Valle del Cauca. Santiago de Cali, Universidad del Valle.
|
|
Espinosa, C., 2019. Relación entre coloración de la coronilla, elección de pareja y dominancia en Sicalis flaveola (Aves: Thraupidae). Universidad del Valle, Colombia.
|
Espinosa, C., Cruz-Bernate, L., Barreto, G., 2017. Biología reproductiva de Sicalis flaveola (Aves: Thraupidae) en Cali, Colombia. Bol. Cient. Mus. His. Nat. 21, 101–114. .
|
|
Figuerola, J., Gutierrez, R., 1998. Sexual differences in levels of blood carotenoids in cirl buntings Emberiza cirlus. Ardea 86, 245–248.
|
Fridolfsson, A.K., Ellegren, H., 1999. A simple and universal method for molecular sexing of non-ratite birds. J. Avian Biol. 30, 116–121. .
|
|
Grubb Jr., T.C., Waite, T.A., Wiseman, A.J., 1991. Ptilochronology: induced feather growth in Northern Cardinals varies with age, sex, ambient temperature, and day length. Wilson Bull. 103, 435–445.
|
Hart, N.S., 2001. Variations in cone photoreceptor abundance and the visual ecology of birds. J. Comp. Physiol. 187, 685–697. https://doi.org/10.1007/s00359-001-0240–3.
|
Hart, N.S., Vorobyev, M., 2005. Modelling oil droplet absorption spectra and spectral sensitivities of bird cone photoreceptors. J. Comp. Physiol. 191, 381–392. https://doi.org/10.1007/s00359-004-0595–3.
|
Hart, N.S., Partridge, J.C., Cuthill, I.C., Bennett, A.T.D., 2000. Visual pigments, oil droplets, ocular media and cone photoreceptor distribution in two species of passerine bird: the blue tit ( Parus caeruleus L. ) and the blackbird ( Turdus merula L. ). J. Comp. Physiol. 186, 375–387. .
|
Håstad, O., Ödeen, A., 2008. Different ranking of avian colors predicted by modeling of retinal function in humans and birds. Am. Nat. 171, 831–838. .
|
Hegyi, G., Laczi, M., Kötél, D., Csizmadia, T., Lőw, P., Rosivall, B., et al., 2018. Reflectance variation in the blue tit crown in relation to feather structure. J. Exp. Biol. 221, jeb176727. .
|
Hegyi, G., Szigeti, B., Török, J., Eens, M., 2007. Melanin, carotenoid and structural plumage ornaments: information content and role in great tits Parus major. J. Avian Biol. 38, 698–708. https://doi.org/10.1111/j.2007.0908-8857.04075.x.
|
Hill, G.E., 1990. Female house finches prefer colourful males: sexual selection for a condition-dependent trait. Anim. Behav. 40, 563–572. .
|
Hill, G.E., 1995. Interspecific variation in plasma hue in relation to carotenoid plumage pigmentation. Auk 112, 1054–1057. .
|
Hill, G.E., 2000. Energetic constraints on expression of carotenoid-based plumage coloration. J. Avian Biol. 31, 559–566. .
|
Hill, G.E., 2011. Condition-dependent traits as signals of the functionality of vital cellular processes. Ecol. Lett. 14, 625–634. https://doi.org/10.1111/j.1461-0248.2011.01622.x.
|
Hill, G.E., Johnson, J.D., 2012. The vitamin A–redox hypothesis: a biochemical basis for honest signaling via carotenoid pigmentation. Am. Nat. 180, E127-E150. .
|
Hill, G.E., McGraw, K.J., 2004. Correlated changes in male plumage coloration and female mate choice in cardueline finches. Anim. Behav. 67, 27–35. .
|
|
Hill, G.E., McGraw, K.J., 2006. Bird Coloration, vol. 2. Function and Evolution. Harvard University Press, Cambridge.
|
Hill, G.E., Montgomerie, R., Inouye, C.Y., Dale, J., 1994. Influence of dietary carotenoids on plasma and plumage colour in the house finch: intra-and intersexual variation. Funct. Ecol. 8, 343–350. .
|
|
Hilty, S.L., Brown, W.L., 2001. Guía de las aves de Colombia. American Bird Conservancy, Cali, Colombia.
|
Hofmann, C., Lo, W.S., Yao, C.T., Li, S.H., 2007. Cryptic sexual dichromatism occurs across multiple types of plumage in the Green-backed Tit Parus monticolus. Ibis 149, 264–270. .
|
|
Horak, P., Surai, P.F., Moller, A.P., 2002. Fat-soluble antioxidants in the eggs of great tits Parus major in relation to breeding habitat and laying sequence. Avian Sci. 2, 123–130.
|
Hung, H.Y., Yeung, C.K., Omland, K.E., Yao, C.T., Yao, C.J., Li, S.H., 2017. Himalayan black bulbuls ( Hypsipetes leucocephalus niggerimus) exhibit sexual dichromatism under ultraviolet light that is invisible to the human eye. Sci. Rep. 7, 43707. .
|
Hunt, S., Bennett, A.T.D., Cuthill, I.C., Griffiths, R., 1998. Blue tits are ultraviolet tits. Proc. R. Soc. B. 265, 451–455. .
|
Irwin, R.E., 1994. The evolution of plumage dichromatism in the New World blackbirds: social selection on female brightness. Am. Nat. 144, 890–907. .
|
Kelber, A., Vorobyev, M., Osorio, D., 2003. Animal colour vision-behavioural tests and physiological concepts. Biol. Rev. 78, 81–118. .
|
Kelly, R.J., Murphy, T.G., Tarvin, K.A., Burness, G., 2012. Carotenoid-based ornaments of female and male American goldfinches ( Spinus tristis) show sex-specific correlations with immune function and metabolic rate. Physiol. Biochem. Zool. 85, 348–363. .
|
Keyser, A.J., Hill, G.E., 2000. Structurally based plumage coloration is an honest signal of quality in male blue grosbeaks. Behav. Ecol. 11, 202–209. .
|
Koch, R.E., Staley, M., Kavazis, A.N., Hasselquist, D., Toomey, M.B., Hill, G.E., 2019. Testing the resource trade-off hypothesis for carotenoid-based signal honesty using genetic variants of the domestic canary. J. Exp. Biol. 222, jeb188102. .
|
Kopp, G., 2023. Daily solar flux as a function of latitude and time. Sol. Energy 249, 250–254. .
|
Kraaijeveld, K., Kraaijeveld-Smit, F.J., Komdeur, J., 2007. The evolution of mutual ornamentation. Anim. Behav. 74, 657–677. .
|
Lee, E., Aoyama, M., Sugita, S., 2009a. Microstructure of the feather in Japanese Jungle Crows ( Corvus macrorhynchos) with distinguishing gender differences. Anat. Sci. Int. 84, 141–147. https://doi.org/10.1007/s12565-009-0022–5.
|
Lee, E., Tanaka, H., Wakamatsu, K., Sugita, S., 2009b. Melanin-based iridescent feather color in the jungle crow. J. Vet. Med. Sci. 71, 1261–1263. .
|
Lou, Y., Chen, L., Zhao, Q., Møller, A.P., Fang, Y., Sun, Y., 2022. Sexual dichromatism and assortative mating by multiple plumage color traits in wild Chestnut Thrush. Avian Res. 13, 100033. .
|
MacDougall, A.K., Montgomerie, R., 2003. Assortative mating by carotenoid-based plumage colour: a quality indicator in American goldfinches, Carduelis tristis. Naturwissenschaften 90, 464–467.
|
Maia, R., Macedo, R.H., Shawkey, M.D., 2012. Nanostructural self-assembly of iridescent feather barbules through depletion attraction of melanosomes during keratinization. J. R. Soc. Interface 9, 734–743. .
|
Maia, R., White, T.E., 2018. Comparing colors using visual models. Behav. Ecol. 29, 649–659. .
|
Maia, R., Eliason, C.M., Bitton, P.P., Doucet, S.M., Shawkey, M.D., 2013. pavo: an R package for the analysis, visualization and organization of spectral data. Methods Ecol. Evol. 4, 906–913. .
|
Maia, R., Gruson, H., Endler, J.A., White, T.E., 2019. Pavo 2: new tools for the spectral and spatial analysis of colour in R. Methods Ecol. Evol. 10, 1097–1107. .
|
Marcondes-Machado, L.O., 1982. Poliginia em Sicalis flaveola brasiliensis (Gmelin, 1789) (Passeriformes, Emberizidae). Rev. Bras. Zool. 1, 95–99. .
|
|
Marcondes-Machado, L.O., 1997. Comportamento social de Sicalis flaveola brasiliensis em cativeiro (Passeriformes, Emberizidae). Iheringia Ser. Zool. 82, 151–158.
|
Martin, T.E., Badyaev, A.V., 1996. Sexual dichromatism in birds: importance of nest predation and nest location for females versus males. Evolution 50, 2454–2460. https://doi.org/10.1111/j.1558-5646.1996.tb03631.x.
|
Martin Schaefer, H., Schaefer, V., Vorobyev, M., 2007. Are fruit colors adapted to consumer vision and birds equally efficient in detecting colorful signals? Am. Nat. 169, S159-S169. .
|
Masello, J.F., Lubjuhn, T., Quillfeldt, P., 2009. Hidden dichromatism in the Burrowing Parrot ( Cyanoliseus patagonus) as revealed by spectrometric colour analysis. Hornero 24, 47–55. .
|
Mason, P., 1985. The nesting biology of some passerines of Buenos Aires, Argentina. Ornithol. Monogr. 36, 954–972. .
|
Mason, N.A., Shultz, A.J., Burns, K.J., 2014. Elaborate visual and acoustic signals evolve independently in a large, phenotypically diverse radiation of songbirds. Proc. R. Soc. B 281, 20140967. .
|
Mays Jr., H.L., McGraw, K.J., Ritchison, G., Cooper, S., Rush, V., Parker, R.S., 2004. Sexual dichromatism in the yellow-breasted chat Icteria virens: spectrophotometric analysis and biochemical basis. J. Avian Biol. 35, 125–134. .
|
McCoy, D.E., Shultz, A.J., Vidoudez, C., van der Heide, E., Dall, J.E., Trauger, S.A., et al., 2021. Microstructures amplify carotenoid plumage signals in tanagers. Sci. Rep. 11, 8582. .
|
|
McGraw, K.J., 2006. Mechanism of carotenoid-based coloration. In: Hill, G.E., McGraw, K.J. (Eds. ), Bird Coloration, Volume 1: Mechanisms and Measurements. Harvard University Press, Cambridge, pp. 177–242.
|
McGraw, K.J., Ardia, D.R., 2003. Carotenoids, immunocompetence, and the information content of sexual colors: an experimental test. Am. Nat. 162, 704–712. .
|
McGraw, K.J., Ardia, D.R., 2004. Immunoregulatory activity of different dietary carotenoids in male zebra finches. Chemoecology 14, 25–29. .
|
McGraw, K.J., Hill. G.E., 2000a. Differential effects of endoparasitism on the expression of carotenoid-and melanin-based ornamental coloration. Proc. R. Soc. A B 267, 1525–1531. .
|
McGraw, K.J., Hill, G.E., 2000b. Carotenoid-based ornamentation and status signaling in the house finch. Behav. Ecol. 11, 520–527. .
|
McGraw, K.J., Hill, G.E., Parker, R.S., 2005. The physiological costs of being colourful: nutritional control of carotenoid utilization in the American goldfinch, Carduelis tristis. Anim. Behav. 69, 653–660. .
|
|
Møller, A.P., Biard, C., Blount, J.D., Houston, D.C., Ninni, P., Saino, N., et al., 2000. Carotenoid-dependent signals: indicators of foraging efficiency, immunocompetence or detoxification ability. Avian Biol. Res. 11, 137–159.
|
Møller, A.P., Erritzøe, J., Saino, N., 2003. Seasonal changes in immune response and parasite impact on hosts. Am. Nat. 161, 657–671. .
|
|
Montgomerie, R., 2006. Analyzing colors. In: Hill, G.E., McGraw, K.J. (Eds. ), Bird Coloration, Volume 1: Mechanisms and Measurements. Harvard University Press, Cambridge, pp. 90–147.
|
Morales, M., Gigena, D.J., Benitez-Vieyra, S.M., Valdez, D.J., 2020. Subtle sexual plumage color dimorphism and size dimorphism in a South American colonial breeder, the Monk Parakeet ( Myiopsitta monachus). Avian Res. 11, 18. .
|
Navara, K.J., Badyaev, A.V., Mendonca, M.T., Hill, G.E., 2006. Yolk antioxidants vary with male attractiveness and female condition in the house finch ( Carpodacus mexicanus). Physiol. Biochem. Zool. 79, 1098–1105. .
|
Norte, A.C., Ramos, J.A., Sousa, J.P., Sheldon, B.C., 2009. Variation of adult great tit Parus major body condition and blood parameters in relation to sex, age, year and season. J. Ornithol. 150, 651–660. .
|
Ödeen, A., Håstad, O., 2003. Complex distribution of avian color vision systems revealed by sequencing the SWS1 opsin from total DNA. Mol. Biol. Evol. 20, 855–861. .
|
Olsson, P., Lind, O., Kelber, A., 2015. Bird colour vision: behavioural thresholds reveal receptor noise. J. Exp. Biol. 218, 184–193. .
|
|
Orozco Valor, P.M., Santillan, M.A., Bragagnolo, L.A., Rebollo, M.E., López, F.G., Martínez, P.A., 2016. Aportes a la biología reproductiva del Chirigüe Azafrán (Sicalis flaveola) en cajas nido en un bosque semiárido del centro de Argentina. Rev. Chi. Ornitol. 22, 165–170.
|
Osorio, D., Smith, A.C., Vorobyev, M., Buchanan-Smith, H.M., 2004. Detection of fruit and the selection of primate visual pigments for color vision. Am. Nat. 164, 696–708. .
|
Owen, J.C., Sogge, M.K., Kern, M.D., 2005. Habitat and sex differences in physiological condition of breeding southwestern willow flycatchers ( Empidonax trailliiextimus). Auk 122, 1261–1270. .
|
|
Palmerio, A.G., 2012. Maduración tardía del plumaje y costo reproductivo en el Jilguero Dorado Sicalis flaveola. Doctoral Thesis. Universidad de Buenos Aires, Buenos Aires.
|
Palmerio, A.G., Massoni, V., 2009. Reproductive biology of female Saffron Finches does not differ by the plumage of the mate. Condor 111, 715–721. .
|
Palmerio, A.G., Massoni, V., 2011. Parental care does not vary with age-dependent plumage in male Saffron Finches Sicalis flaveola. Ibis 153, 421–424. .
|
Parsons, J., Baptista, L.F., 1980. Crown color and dominance in the white-crowned sparrow. Auk 97, 807–815. .
|
Powers, M.J., Hill, G.E., 2021. A review and assessment of the Shared-Pathway Hypothesis for the maintenance of signal honesty in red ketocarotenoid-based coloration. Integr. Comp. Biol. 61, 1811–1826. .
|
Price-Waldman, R.M., Shultz, A.J., Burns, K.J., 2020. Speciation rates are correlated with changes in plumage color complexity in the largest family of songbirds. Evolution 74, 1155–1169. .
|
Pryke, S.R., 2007. Fiery red heads: female dominance among head color morphs in the Gouldian finch. Behav. Ecol. 18, 621–627. .
|
|
Quiroga, M., del Barco, O., Agostelli, F., 2003. First approaches to de reproductive biology of Sicalis flaveola birds: Emberizidae at the alluvial valley of Paraná River, Argentina. FAVE 2, 35–40.
|
R Core Team, 2021. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. .
|
Radford, A.N., Du Plessis, M.A., 2004. Extreme sexual dimorphism in Green Woodhoopoe ( Phoeniculus purpureus) bill length: a case of sexual selection? Auk 121, 178–183. .
|
|
Rising, J.D., Jaramillo, A., Copete, J.L., Ryan, P.G., Madge, S., 2011. Tanagers to new world blackbirds. In: del Hoyo, J., Elliott, A., Christie, D. (Eds. ), Handbook of the Birds of the World, Vol. 16. Lynx Edicions, Barcelona, pp. 381–382.
|
Saks, L., McGraw, K., Horak, P., 2003. How feather colour reflects its carotenoid content. Funct. Ecol. 17, 555–561. https://doi.org/10.1046/j.1365-2435.2003.00765.x.
|
Sánchez, R., Blendinger, P.G., Xuereb, A., Lougheed, S.C., 2018. Extra-pair paternity in a socially monogamous neotropical tanager, the ringed warbling-finch Poospiza torquata. Acta Ornithol. 52, 197–208. .
|
Seutin, G., White, B.N., Boag, P.T., 1991. Preservation of avian blood and tissue samples for DNA analyses. Can. J. Zool. 69, 82–90. .
|
Shultz, A.J., Burns, K.J., 2017. The role of sexual and natural selection in shaping patterns of sexual dichromatism in the largest family of songbirds (Aves: Thraupidae). Evolution 71, 1061–1074. .
|
Siddiqi, A., Cronin, T.W., Loew, E.R., Vorobyev, M., Summers, K., 2004. Interspecific and intraspecific views of color signals in the strawberry poison frog Dendrobates pumilio. J. Exp. Biol. 207, 2471–2485. .
|
Siefferman, L., Hill, G.E., 2003. Structural and melanin coloration indicate parental effort and reproductive success in male eastern bluebirds. Behav. Ecol. 14, 855–861. .
|
Siitari, H., Honkavaara, J., Huhta, E., Viitala, J., 2002. Ultraviolet reflection and female mate choice in the pied flycatcher, Ficedula hypoleuca. Anim. Behav. 63, 97–102. .
|
|
Skutch, A.F., Gardner, D., 1989. Life of the Tanager. Comstock Publishing Associates,
Ithaca.
|
Stoffel, M.A., Nakagawa, S., Schielzeth, H., 2017. rptR: repeatability estimation and variance decomposition by generalized linear mixed-effects models. Methods Ecol. Evol. 8, 1639–1644. .
|
Székely, T., Reynolds, J.D., Figuerola, J., 2000. Sexual size dimorphism in shorebirds, gulls, and alcids: the influence of sexual and natural selection. Evolution 54, 1404–1413. https://doi.org/10.1111/j.0014-3820.2000.tb00572.x.
|
|
Tabachnick, B.G., Fidell, L.S., 2001. Using Multivariate Statistics. Pearson, Boston.
|
Toomey, M.B., Ronald, K.L., 2021. Avian color expression and perception: is there a carotenoid link? J. Exp. Biol. 224, jeb203844. .
|
Toomey, M.B., Collins, A.M., Frederiksen, R., Cornwall, M.C., Timlin, J.A., Corbo, J.C., 2015. A complex carotenoid palette tunes avian colour vision. J. R. Soc. Interface 12, 20150563. .
|
Tubaro, P.L., Lijtmaer, D.A., Lougheed, S.C., 2005. Cryptic dichromatism and seasonal color variation in the diademed tanager. Condor 107, 648–656. .
|
Valdez, D.J., Benitez-Vieyra, S.M., 2016. A spectrophotometric study of plumage color in the Eared Dove ( Zenaida auriculata), the most abundant South American Columbiforme. PLoS one 11, e015550. .
|
Vorobyev, M., Osorio, D., 1998. Receptor noise as a determinant of colour thresholds. Proc. R. Soc. B 265, 351–358. .
|
|
Webb, E.A., 2021. Where Have the Carotenoids Gone? Physiology of Carotenoid Absorption and Distribution in Birds. Doctoral Thesis. Arizona State University, Tempe.
|
Webster, M.S., 1992. Sexual dimorphism, mating system and body size in New World blackbirds (Icterinae). Evolution 46, 1621–1641. https://doi.org/10.1111/j.1558-5646.1992.tb01158.x.
|
Wetmore, A., 1936. The number of contour feathers in passeriform and related birds. Auk 53, 159–169. .
|
Wilkinson, K.E., Palmer, G.C., 2022. Ambient light energy intensity as a trigger for the dawn chorus: Patterns in five common eastern Australian bird species. Aust. Field Ornithol. 39, 82–88. .
|


 DownLoad:
DownLoad:
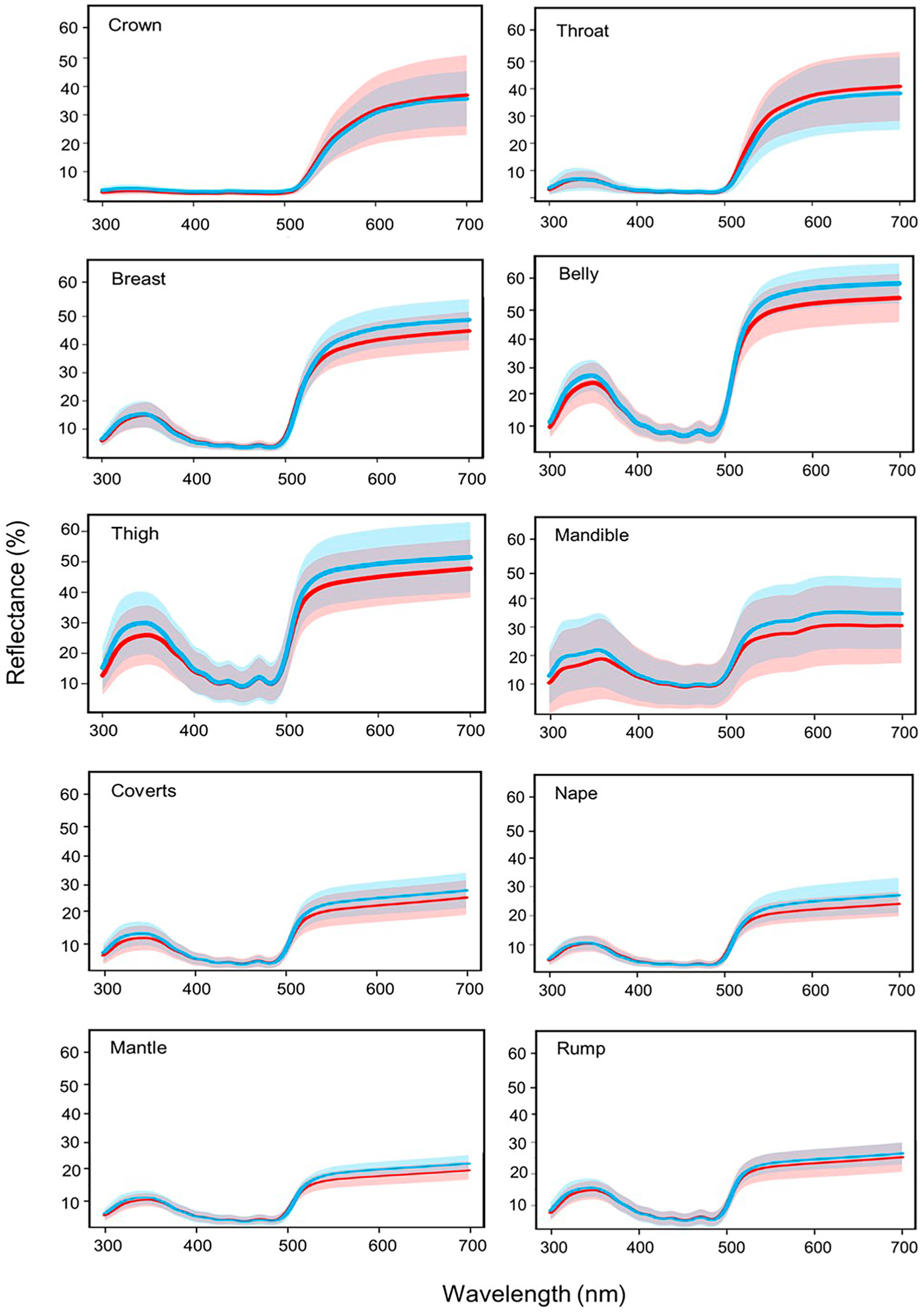
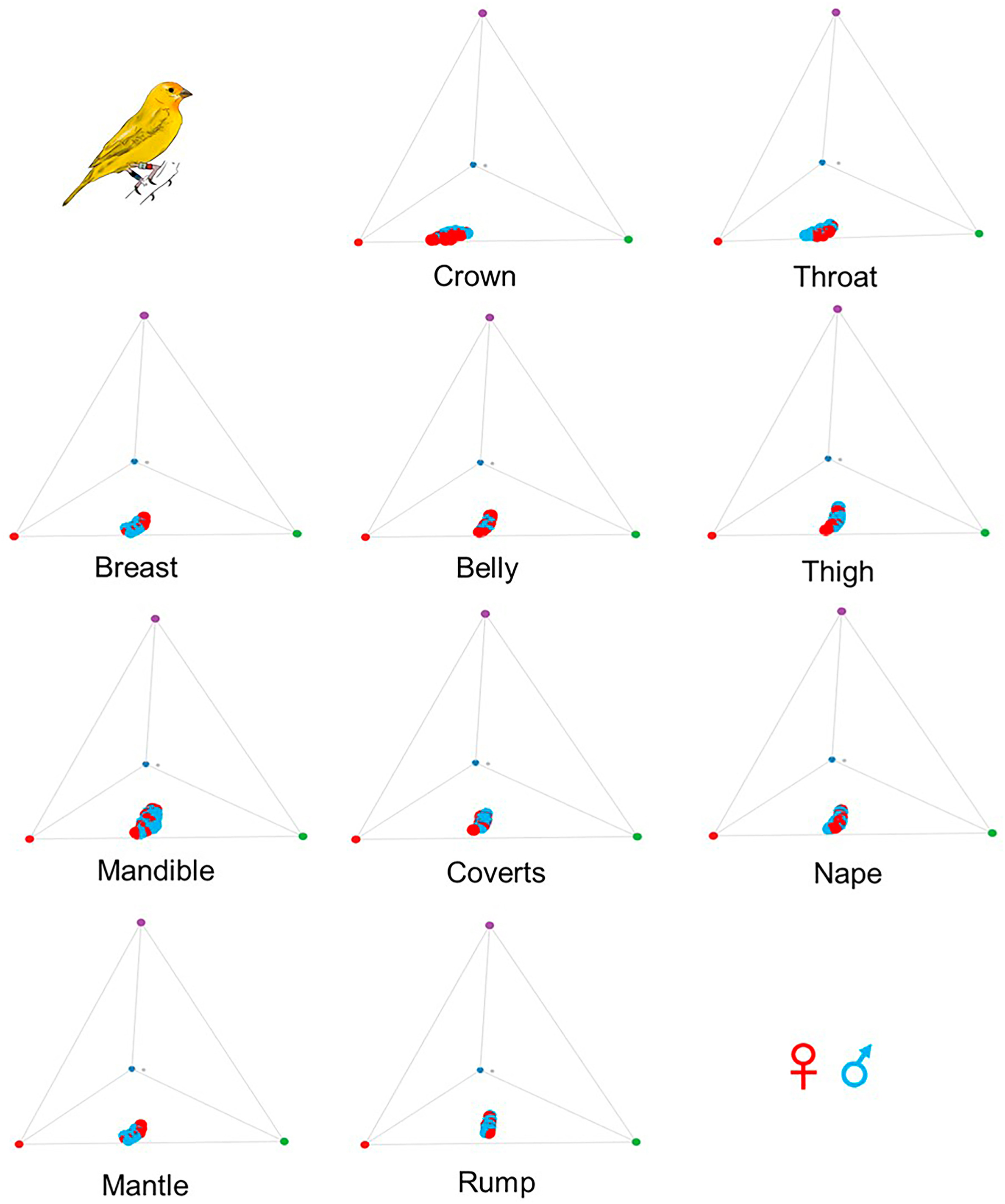
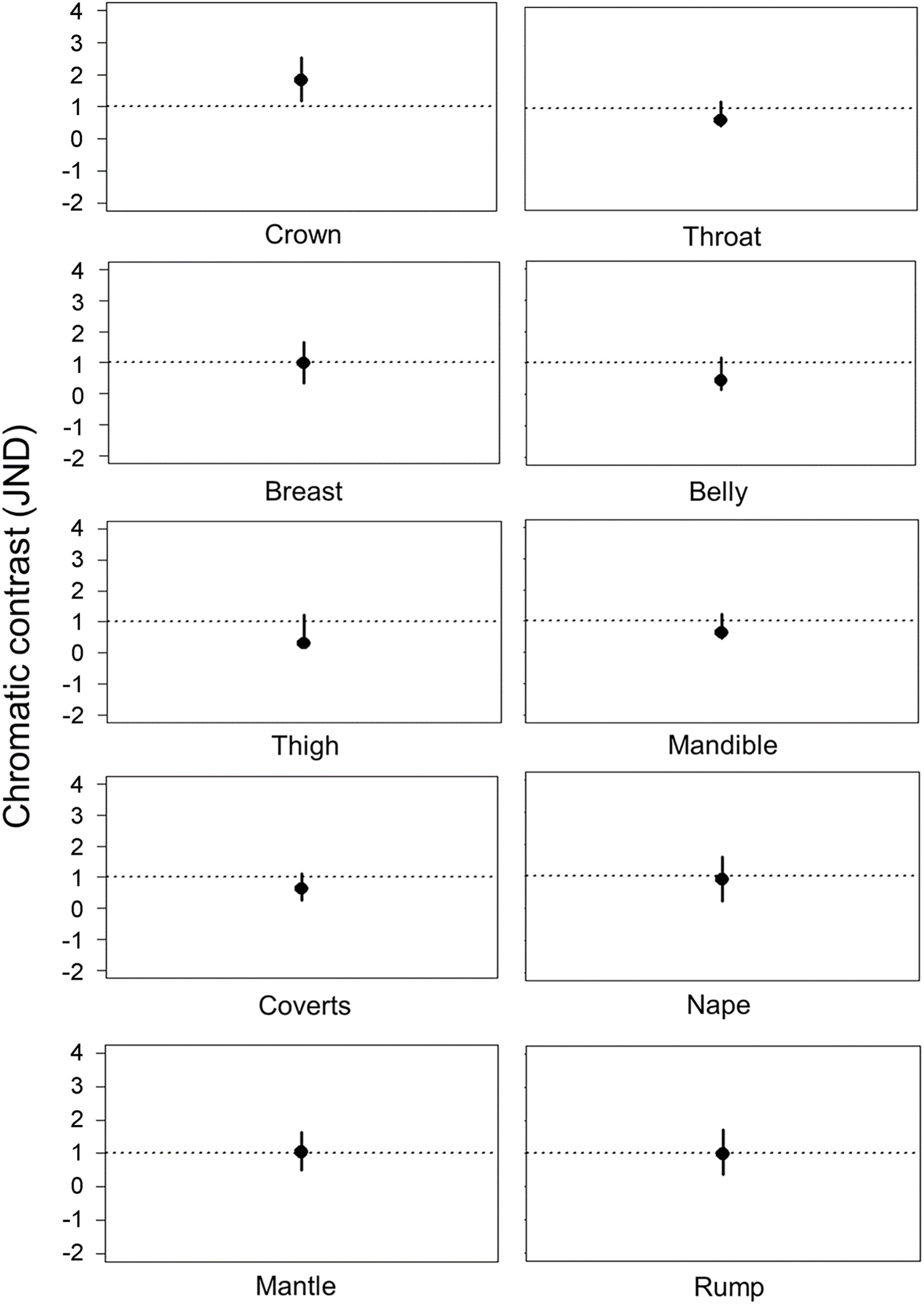





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds