
Figure
1.
Location of the Hara Biosphere Reserve in southern Iran
| Citation: |
Ana B. Navarro, Juliano A. Bogoni, Marcelo Z. Moreira, Luís F. Silveira. 2023: Intraguild niche partitioning in granivorous birds from the late past. Avian Research, 14(1): 100075. DOI: 10.1016/j.avrs.2023.100075

|
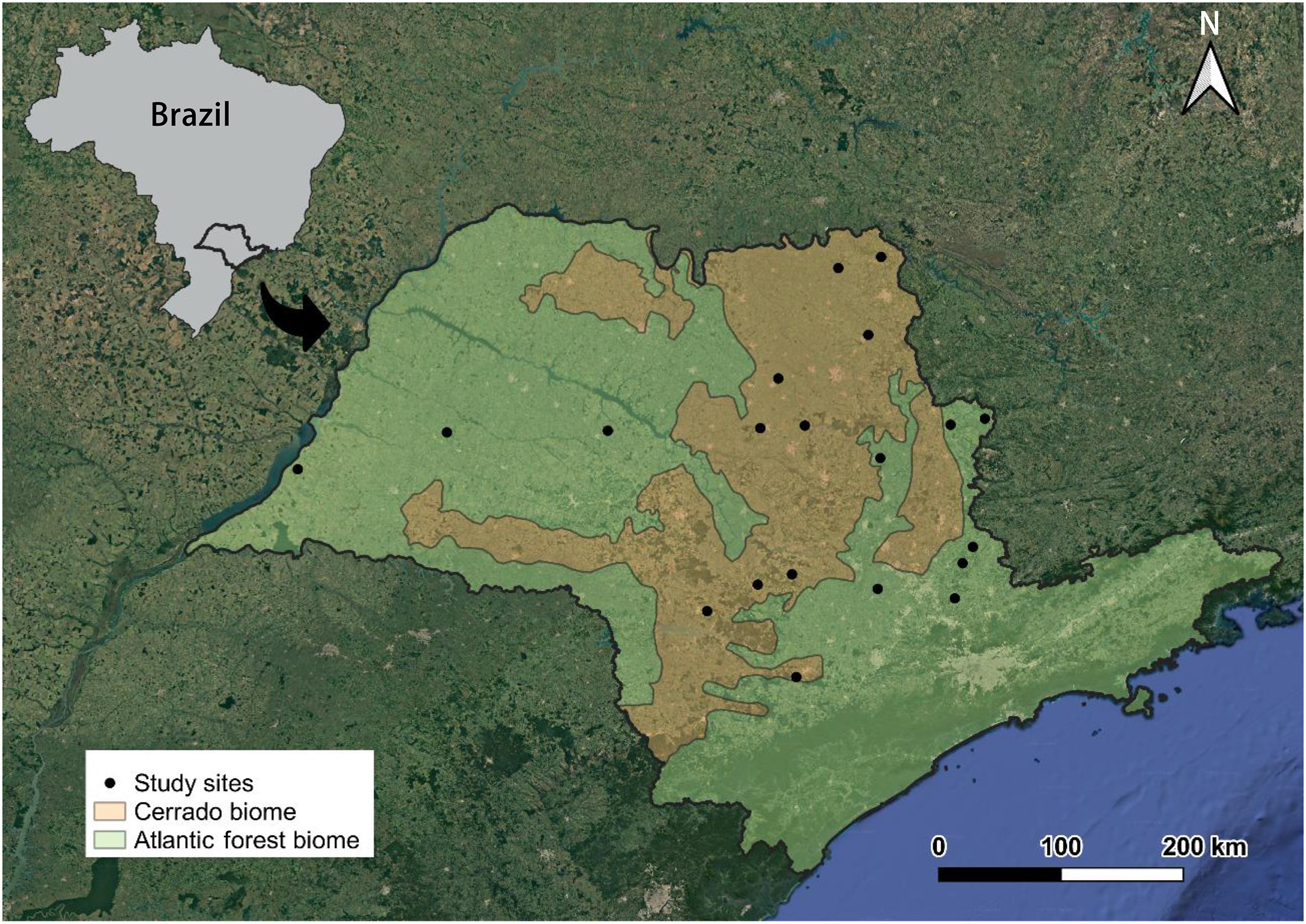
Niche partitioning is a widespread ecological strategy within trophic guilds, ensuring the coexistence of sympatric species by reducing interspecific competition. Stable isotope analysis of carbon and nitrogen and isotopic niche metrics (width and overlap) are used as a proxy to investigate niche partitioning among species of a guild. In our study, we investigated if niche partitioning was an ecological mechanism contributing to the coexistence of granivorous birds from the same geographic region along time, employing an isotopic approach. We sampled and isotopically analyzed (δ13C and δ15N values) wing feathers from 58 specimens of granivorous birds collected between 1900 and 1966 in southeastern Brazil. We grouped birds according to the main habitat type used by them (forest: Cyanoloxia brissonii, Leptotila rufaxilla, and Leptotila verreauxi; and grassland: Columbina squammata and Sicalis flaveola). We used the Isotopic Richness (IRic) metric to measure the isotopic niche width and the Isotopic Similarity (ISim) and Isotopic Nestedness (INes) metrics to measure the niche overlap between groups and species. The results of low isotopic niche overlap suggest that common granivorous birds had a clear niche partitioning in southeastern Brazil in the late past, especially through foraging in distinct habitat types (forest and grassland; ISim = 0.1, INes = 0.2). Niche overlaps in almost all species-by-species comparisons were low (ISim ≤0.3, INes ≤0.4), except between C. brissonii and L. verreauxi (ISim = 0.6, INes = 1.0). These results suggest that these birds, belonging to the same trophic guild, coexisted through niche and resource partitioning. Despite being considered a plastic trophic guild, the isotopic niche partitioning among granivorous birds indicates that each of these species uses a set of specific resources and habitats. This raises an ecological concern about the homogenization of landscapes across the Neotropics, oversimplifying food resources and habitats to granivorous birds.
Pollution of the natural environment by heavy metals is a worldwide problem. Heavy metals enter the aquatic ecosystem through a variety of anthropogenic sources as well as from natural processes (Ebrahimpour and Mushrifah, 2010). They are a serious threat because of their toxicity, bioaccumulation, long persistence and bio-magnification in the food chain (Erdoĝrul and Ates, 2006). The degree of toxic metal uptake, translocation and eventual detoxification within an organism depends on metal speciation, but also differs strongly among organisms (Mukherjee and Nuorteva, 1994; Doyle and Otte, 1997). Assessing ecosystem health adequately by means of biomonitoring requires the selection of representative indicator species. Birds are widely used to biomonitoring variation in environmental levels of anthropogenic pollutants (Furness and Camphuysen, 1997; Barbieri et al., 2009), because they are exposed to a wide range of chemicals and occupy high trophic levels and can therefore provide information on the extent of contamination in the entire food chain (Furness and Camphuysen, 1997; Burger et al., 2007).
According to Burger and Gochfeld(2000a, 2000b) feathers are useful for measuring heavy metal contamination in birds because birds sequester heavy metals in their feathers, where the proportion of the body burden is relatively constant for each metal. In general, metals in breast feathers are representative of circulating concentrations in the blood stream only during the limited time period of feather formation, which in turn represents both local exposure and mobilization from internal tissues (Lewis and Furness, 1991; Monteiro, 1996). Many studies have recommended herons and egrets as bioindicators for heavy metals in aquatic systems and local pollution around breeding sites (Boncompagni et al., 2003; Kim and Koo, 2007). Herons and gulls are high at the top of their food pyramid and can yield information over a large area around each sampling site, not only on bioavailability of contaminants but also on how, where, and when they are transferred within the food web (Battaglia et al., 2005). Hence, the aim of the present study was to investigate the level of nickel in feathers of two bird species, the Western Reef Heron (Egretta gularis) and the Siberian Gull (Larus heuglini), in order 1) to compare metal concentrations between two species with their life strategy and 2) to examine the species and gender related variation in trace nickel accumulations in the Hara Biosphere Reserve of southern Iran.
The Hara Biosphere Reserve (26°40′–27°N, 55°21′–55°52′E) is located in southern Iran, in the Straits of Khuran between Queshm Island and the Persian Gulf (Fig. 1). This area became part of the Man and Biosphere Program (MAB) of UNESCO in 1977 (UNESCO, 2010). As well, this region is one of the protected areas in Iran introduced by the Department of the Environment. The entire region was selected as wetland of international importance under the Wetlands of International Importance category as a Habitat of Aquatic Birds. This region was also introduced as one of the important bird areas by the International Organization of Birdlife (Neinavaz et al., 2010).
During November and December 2010, under license of the Environmental Protection Agency of Hormozgan, a total of thirty birds were shot and removed from throughout the Hara biosphere reserve. The collection included the Western Reef Heron (Egretta gularis) (n = 15) and the Siberian Gull (Larus heuglini) (n = 15). The birds were transported to the laboratory packed in ice. The specimen were killed, weighed, stored in plastic bags and kept at −20℃ until dissection and analysis. We chose breast feathers because they are representatives of the plumage and are less affected by molt compared to flight feathers. All feathers were analyzed in the Laboratory of the Inland Water Aquaculture Research Institute in port Anzali. The feather samples were digested in a nitric acid (HNO3) and perchloric acid (HClO4) mixture. Feathers were then accurately weighed in 150-mL Erlenmeyer flasks, where 10 mL nitric acid (65%) was added to each sample. The samples were left overnight to be slowly digested; thereafter, 5 mL perchloric acid (70%) was added to each sample. Digestion was performed on a hot plate (sand bath) at 200℃. After that, the digested samples were diluted by 25 mL deionized water. The concentration of nickel was estimated using a Shimadzu AA 680 flame atomic absorption spectrophotometer. The accuracy of the analysis was checked by measuring CRM certified reference tissue (DORM-2, NRC Canada). The detection limit for nickel was 0.039 µg·g−1. The results for nickel gave a mean recovery of 98.6%.
A statistical analysis was carried out using SPSS (version 18.0). We used a three-way ANOVA for nickel (sex, age, species, interaction (sex × age × species)). Data were log transformed to obtain normal distributions that satisfied the homogeneity of variance required by ANOVA (Custer et al., 2003; Kim et al., 2009). The nickel concentration in feathers was tested for mean differences between species using Student t tests. The level of significance was set at α = 0.05. The concentration of nickel in feathers was expressed in microgram per gram of dry weight (dw). Values are given in means ± standard errors (SE).
Variations in the nickel concentration of feathers of the Western Reef Heron and Siberian Gull are presented in Table 1. The results show that there was a significant difference between the mean nickel concentrations in the two bird species, i.e. the Western Reef Heron and Siberian Gull, while there was no evidence of significant different accumulation between genders and ages (Table 2). Also, the results indicate that the level of nickel concentration in the Western Reef Heron was higher in females than in males; in contrast, the level of nickel concentration in the Siberian gull was higher in males.
| Species | Male/adult | Male/juvenile | Female/adult | Female/juvenile |
| Western Reef Heron | ||||
| Geometric mean | 2.77 | – | 3.79 | 4.67 |
| Mean ± SE | 3.47±2.7 | – | 4.67±2.7 | 4.67 |
| Number | 9/9 | – | 5/5 | 1 |
| Siberian Gull | ||||
| Geometric mean | 8.89 | 3.87 | 6.72 | 7.24 |
| Mean ± SE | 8.94±1.1 | 4.38±2.5 | 7.6±3.4 | 8.1±4.8 |
| Number | 3/3 | 3/3 | 6/6 | 3/3 |
 DownLoad:
CSV
DownLoad:
CSV
| Source of variation | Mean square | F | p |
| Species | 78.23 | 8.56 | 0.01 a |
| Gender | 9.56 | 1.04 | NS b |
| Age | 7.57 | 0.82 | NS |
| Intercept (species × gender × age) | 572.55 | 62.66 | 0.001 |
|
a p-value for 3-way ANOVA. Interaction term significant as indicated. b NS = not significant at p > 0.05. |
|||
DownLoad:
CSV
Nickel is not an important trace element in organisms, but at high levels, they can cause adverse health effects. Sources of heavy metals vary considerably. It is emitted into the environment by both natural and man-made sources. Once released into the environment, nickel readily forms complexes with many ligands, making it more mobile than most heavy metals (Mansouri et al., 2011). Nickel is related to the pigmentation of feathers in birds and excreted via the feathers by moulting (Honda et al., 1986). Nickel concentrations in the current study (3.47–8.94 µg·g−1) were higher than those in Fulica atra (0.8 µg·g−1), Phalacrocorax carbo (0.5 µg·g−1) and Nycticorax nycticorax (1.2 µg·g−1) from Russia (Lebedeva, 1997), in Egretta alba (0.2 µg·g−1) from Korea (Honda et al., 1986) and Parus major (0.25 µg·g−1) from China (Deng et al., 2007). In our study, the nickel concentrations were similar to those in Bubulcus ibis L. (7.8–9.0 µg·g−1) from Pakistan (Malik and Zeb, 2009).
Few studies have examined the effect of gender on the accumulation of heavy metals in feathers and other tissues (Burger, 1995; Zamani-Ahmadmahmoodi et al., 2010). Several studies reported no significant differences in the heavy metal content of feathers between male and female birds (Hutton, 1981; Zamani-Ahmadmahmoodi et al., 2009a). Similarly, in the present study there was no evidence of significant different accumulations between male and female birds (Table 2), suggesting that both sexes utilize similar foraging strategies in both species (Hindell et al., 1999). While studying heavy metals in the feathers of Larus dominicanus, Barbieri et al. (2009) showed that the levels of nickel concentration were higher in adults (5.92 µg·g−1) than in juveniles (2.23 µg·g−1). Similar levels of nickel have been detected in other seabird species from different parts of the world (Norheim, 1987). Adults have had several years to accumulate metals in their internal tissues; these can be mobilized into the blood and deposited in feathers during their formation (Burger, 1994). Elsewhere, Burger and Gochfeld (1991) pointed to heavy metal concentrations in feathers of adult birds that may reflect exposure obtained at other time of the year, including exposure at non-breeding areas. On the other hand, while they were studying heavy metal concentrations in the feathers of Herring Gulls (Larus argentatus) in Captree, Long Island, they showed that the cadmium concentration was higher in juveniles but the lead concentration was higher in adults (Burger, 1995). Differences in levels of metal concentrations in adults and fledglings might also occur if adults and young eat different types of food during the breeding season, or different-sized food items (Burger, 1996).
Research has indicated that the concentration of heavy metals in the tissues of migratory birds is higher than that in resident birds (Pacyna et al., 2006; Zamani-Ahmadmahmoodi et al., 2009b). Siberian Gulls are winter visitors to Hara Biosphere Reserve, while Western Reef Heron are residents there. The results of the current study show that the amount of nickel in the Siberian Gull feathers is higher than in the Western Reef Heron.
The results of three-way ANOVA showed there were significant differences between the Western Reef Heron and Siberian Gull (p < 0.01). The Siberian Gull showed higher nickel concentrations than the Western Reef Heron. Birds that are large fish eaters should accumulate higher levels than those that eat a range of different foods or smaller fish. Furthermore, levels of metal in birds should reflect the levels in the fish they consume (Burger, 2002). The main difference in nickel levels between Western Reef Heron and Siberian Gull could be the result of different phylogenetic origin and physiology (Teal, 1969; Welty, 1975; Deng et al. 2007), or because they grow their feathers in different breeding areas with different levels of background contamination. Also, metabolic rates vary inversely with body weight and directly with activities such as flight and rest. Being smaller than Western Reef Heron, Siberian Gull was expected to have a higher metabolic rate. Higher metabolic rates may cause fast accumulations of trace nickel in the Siberian Gull. In general, the Siberian Gull eats more invertebrates than the Western Reef Heron, catches some larger fish (between 20–25 cm in size), consumes offal discarded by fishing boats and eats dead fish found along the shore, while the Western Reef Heron eats smaller fish, amphibians and insects.
|
Alexandrino, E.R., Bovo, A.A.A., da Luz, D.T.A., da Costa, J.C., Betini, G.S., de Barros Ferraz, K.M.P.M., et al., 2013. Aves do campus "Luiz de Queiroz" (Piracicaba, SP) da Universidade de São Paulo: mais de 10 anos de observações neste ambiente antrópico. Atual. Ornitol. 173, 40–52.
|
|
Botta, S., Secchi, E.R., Rogers, T.L., Prado, J.H.F., de Lima, R.C., Carlini, P., et al., 2018. Isotopic niche overlap and partition among three Antarctic seals from the western Antarctic Peninsula. Deep-Sea Res. Pt. Ⅱ 149, 240–249.
|
|
Branco, J.O., Keske, B.R., Barbieri, E., 2016. Abundance and potential impact of granivorous birds on irrigated rice cultivation, Itajaí, Santa Catarina, Brazil. Arq. Inst. Biol. 83, e0462014.
|
|
Chase, J.M., Leibold, M.A., 2003. Ecological Niches: Linking Classical and Contemporary Approaches. The University of Chicago Press, Chicago.
|
|
Cotton, P.A., 1998. Temporal partitioning of a floral resource by territorial hummingbirds. Ibis 140, 647–653.
|
|
Destro, G.F.G., Pimentel, T.L., Sabaini, R.M., Borges, R.C., Barreto, R., 2012. Efforts to combat wild animals trafficking in Brazil. In: Akeem, L.G. (Ed.), Biodiversity Enrichment in a Diverse World. IntechOpen, London, pp. 421–436.
|
|
Ingels, J., 1982. Separation in the field of white-fronted and grey-fronted doves (Leptotila verreauxi and Leptotila rufaxilla). Bonner Zool. Beiträge 33, 61–70.
|
|
Klink, C.A., Machado, R.B., 2005. A conservação do Cerrado brasileiro. Megadiversidade 1, 147–155.
|
|
Kronka, F.J.N., Nalon, M.A., Matsukuma, C.K., Kanashiro, M.M., Ywane, M.S.S. -I., Barradas, A.M.F., et al., 2005. Monitoramento da vegetação natural e do reflorestamento no Estado de São Paulo. Anais XII Simpósio Brasileiro de Sensoriamento Remoto 16–21, 1569–1576.
|
|
Morin, P.J., 2011. Community Ecology, second ed. Wiley-Blackwell, Chichester.
|
|
Piratelli, A.J., Siqueira, M.A.C., Marcondes-Machado, L.O., 2000. Reprodução e muda de penas em aves de sub-bosque na região leste de Mato Grosso do Sul. Rev. Bras. Ornitol. 8, 99–107.
|
|
R Core Team, 2022. R: a Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna.
|
|
Revelle, R., Suess, H.E., 1957. Carbon dioxide exchange between atmosphere and ocean and the question of an increase of atmospheric CO2 during the past decades. Tellus 9, 18–27.
|
|
Sano, S.M., Almeida, S.P., Ribeiro, J.F., 2008. Cerrado: Ecologia e Flora. Embrapa Informação Tecnológica, Brasília.
|
|
Silverthorne, K.M., Perrig, P.L., Lambertucci, S.A., Pauli, J.N., 2020. Isotopic niche partitioning in avian scavenger communities of South America. J. Raptor Res. 54, 440–446.
|
|
Tilman, D., 1982. Resource Competition and Community Structure. Princeton University Press, Princeton.
|
|
Victor, M.A.M., Cavalli, A.C., Guillaumon, J.R., Serra Filho, R., 2005. Cem Anos de Devastação: Revisitada 30 Anos Depois. Ministério do Meio Ambiente, Brasília.
|
|
Wiley, A.E., James, H.F., Ostrom, P.H., 2017. Emerging techniques for isotope studies of avian ecology. In: Webster, M.S. (Ed.), The Extended Specimen: Emerging Frontiers in Collections-Based Ornithological Research. CRC Press, Boca Raton, pp. 89–109.
|
|
Wilman, H., Belmaker, J., Simpson, J., de la Rosa, C., Rivadeneira, M.M., Jetz, W., 2014. EltonTraits 1.0: species-level foraging attributes of the world's birds and mammals. Ecology 95, 2027–2027.
|
|
Xu, X., Jiang, J., Lei, Y., Wang, C., Qing, B., Ding, C., 2022. Using stable isotope to compare the habitat use and trophic level between the new and old breeding range of wild Crested Ibis in the early breeding season. Avian Res. 13, 100007.
|
Figures(3) / Tables(2)





| Species | Male/adult | Male/juvenile | Female/adult | Female/juvenile |
| Western Reef Heron | ||||
| Geometric mean | 2.77 | – | 3.79 | 4.67 |
| Mean ± SE | 3.47±2.7 | – | 4.67±2.7 | 4.67 |
| Number | 9/9 | – | 5/5 | 1 |
| Siberian Gull | ||||
| Geometric mean | 8.89 | 3.87 | 6.72 | 7.24 |
| Mean ± SE | 8.94±1.1 | 4.38±2.5 | 7.6±3.4 | 8.1±4.8 |
| Number | 3/3 | 3/3 | 6/6 | 3/3 |
DownLoad:
CSV
| Source of variation | Mean square | F | p |
| Species | 78.23 | 8.56 | 0.01 a |
| Gender | 9.56 | 1.04 | NS b |
| Age | 7.57 | 0.82 | NS |
| Intercept (species × gender × age) | 572.55 | 62.66 | 0.001 |
|
a p-value for 3-way ANOVA. Interaction term significant as indicated. b NS = not significant at p > 0.05. |
|||
DownLoad:
CSV
 Email Alerts
Email Alerts RSS Feeds
RSS Feeds