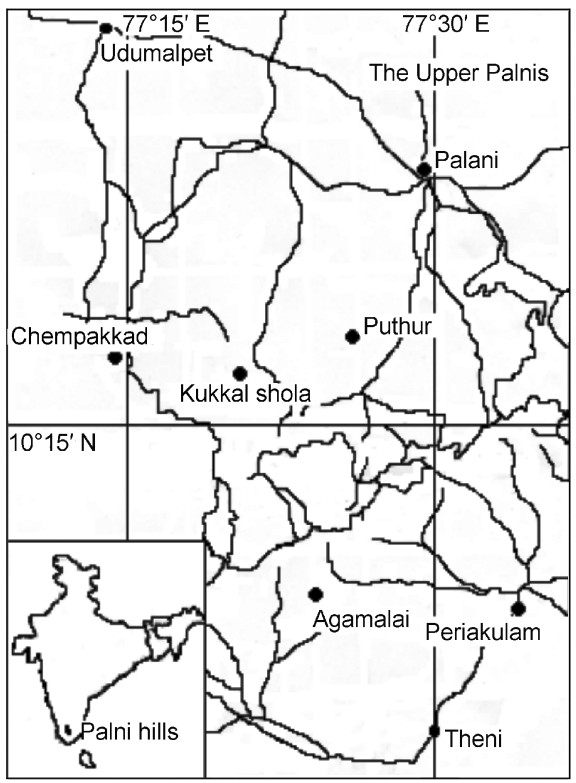
Figure
1.
Map of the Palni Hills with the location of Kukkal shola (scale 1:250000)
| Citation: |
Sellamuthu SOMASUNDARAM, Lalitha VIJAYAN. 2010: Foraging ecology of the globally threatened Nilgiri Wood Pigeon (Columba elphinstonii) in the Western Ghats, India. Avian Research, 1(1): 9-21. DOI: 10.5122/cbirds.2009.0017

|
Foraging ecology of the globally endangered Nilgiri Wood Pigeon (Columba elphinstonii) was investigated in the Western Ghats,India,using faecal sampling and direct observations. The birds fed on fruits of 39 plant species,on seeds of 11 species,flowers and leaf buds of four species and a few ground invertebrates. Direct observations across seasons showed nine plant species were selected more often (47% of observations),with fruits of the members of the family Lauraceae the most preferred. Gleaning accounted for 76% of feeding methods throughout the year. Feeding was mainly from twigs at the edges on the upper and middle canopy (> 6 m). Frequency of feeding on fruits correlated significantly with fruit abundance,which depended on rainfall. The present study indicates that the pigeon requires a wide variety of diets from different forest environments. Conservation of this species depends on preserving all habitat types required to meet their resource needs throughout the year.
Columbids are the fundamental agents in the regeneration of non-wooded areas and are among the most important agents in the colonization and building of wooded habitats in tropical forests. There are 318 species of pigeons and doves found in the world, divided into 42 genera (Gibbs et al., 2001). Of the 318 species of columbid, 13 are already extinct and 59 (19%) are categorized as Threatened and a further 40 (13%) as Near Threatened (BirdLife International, 2008). Overall, nearly one third of extant columbids face, to some degree, a threat of extinction. In Asia, two species, i.e., the Silvery Wood-Pigeon (Columba argentin) and the Negros Fruit Dove (Ptilinopus arcanus) disappeared during the last decade. Both were frugivores and forest dwellers. Virtually all threatened pigeons and doves that are confined to the tropics, are dependent on forests and have restricted ranges with over three-quarters inhabiting the tropical forests of Asia, Australasia and Oceania (Gibbs et al., 2001). In India, 29 species of columbids are found, of which three are listed as Threatened. In the Western Ghats, 12 species are reported, of which one, the Nilgiri Wood Pigeon (Columba elphinstonii), is endemic and also Threatened (Ali and Ripley, 1987; BirdLife International, 2001).
Information on the basic ecology of most Columbidae families in the tropics is very limited, especially on food and feeding (Recher and Date, 1988). In India, no such studies have been conducted for columbids. Food is often considered to be the most important factor affecting the production of offspring (Lack, 1968; Martin, 1987). The relationship between bird and food abundance has been relatively well studied in the tropics, particularly in frugivores (Levey, 1988; Loiselle and Blake, 1991). Most tropical rain forest trees produce fruits that are consumed by animals and rely on frugivores for their seed dispersal (Frankie et al., 1974; Howe, 1986). Concomitantly, a large proportion of the vertebrates in tropical forests are frugivorous (Terborgh, 1986a, 1986b). Food is the major factor determining habitat use (Kwit et al., 2004). However, food shortages can act globally by limiting population levels and the reproductive success of a number of tropical birds (e.g. Martin, 1987; Thiollay, 1989; Robinet et al., 2003).
Phenological patterns can be described as periodic variations in the flowering and fruiting patterns of a species. It can be defined as variations in a) the number of species in flower or fruit, b) the proportion of plants bearing flowers or fruits and c) the abundance of flowers or fruits over time (Blake et al., 1990). Monitoring plant phenology is imperative for the description of temporal variation in resource availability for frugivores. Since the Nilgiri Wood Pigeon is a frugivorous bird, information on the pattern of availability of fruit resources would give a better insight into the response of this species to phenological changes and the vagaries of food abundance.
In the case of the Nilgiri Wood Pigeon an endemic, globally endangered species of the Western Ghats (BirdLife International, 2008), no detailed information about the food and feeding habits is available for evolving a long-term conservation program. However, stomach contents of the Nilgiri Wood Pigeon showed fruits, mainly from Randia dumetorum, and snails (Ali and Ripley, 1987). Hence the present study deals with the foraging ecology of the Nilgiri Wood Pigeon in relation to food availability, which should assist in better management for this globally threatened species.
The study was conducted in a 20-ha plot, from April 2002 to May 2004, in the wet montane temperate (Shola) forests at Kukkal in the Palni hills (10°1′– 10°26′N; 77°14′–77°52′E), a range of hills in the Western Ghats, Tamil Nadu (Fig. 1). The Palni hills consist of two well marked topographic divisions, i.e., the Upper and Lower Palnis. The Upper Palnis, with elevations ranging between 1500 and 2450 m, have a moderate climate with mean temperatures of 12 to 23℃ in summer and 8.3 to 17.3℃ in winter. This area is subject to high winds (Rawat et al., 2003). The annual rainfall averages 1650 mm. The vegetation is predominantly of the wet montane temperate forest type or Shola (Champion and Seth, 1968). The profile of the forest is stunted forests with a canopy height of approximately 15 m (Ramesh and Pascal, 1998). Common plants include species of Syzygium, Ternstroemia, Sideroxylon, Meliosma, Elaeocarpus, Symplocos, Eurya, Litsea and Rhododendron. The forest has adjacent agricultural fields and exotic plantations of Acacia, Eucalyptus and Pinus species (Mathew, 1996). A total of 83 species of birds was recorded from the Kodaikanal area, which has a variety of habitats (Somasundaram and Vijayan, 2004) of which seven are endemic (Inskipp et al., 1996; Grimmett et al., 1998; Somasundaram and Vijayan, 2008).
Foraging ecology of the Nilgiri Wood Pigeon was studied by both direct and faecal sampling methods. In the direct method, birds were observed by using a focal animal sampling or scan sampling method (Altmann, 1974), where the availability of food was assessed by direct observation (Blake et al., 1990). To confirm the direct observation, faecal sampling method was adopted as suggested by Ralph et al. (1985) and Oliveira et al. (2002). In the faecal sampling method, faecal pellets were collected in the resting, roosting and nesting places and were examined.
Foraging activity was recorded throughout the day every five minutes with one minute interval to cover four full days in a month for an average of 50 hours of observations per month. Whenever a foraging bird was observed, only one foraging record (initial record) was made for any individual encountered (MacNally, 1994) and maximum effort was taken to avoid repeated sampling of the same individual. This was continued till the bird was visible. During each foraging observation, the following details were recorded: 1) foraging method, 2) foraging substrate, 3) foraging height and 4) other behavior, if any.
We recognized two major foraging methods: 1) gleaning and 2) leaping. Gleaning consists of picking food items from nearby substrates, including the ground, which can be reached without full extension of legs or neck; no acrobatic movements are involved. Leaping means launching into the air to reach a food item too far for a “reach, “but too close for a “sally”. This differs from “sally” in that the upward thrust seems to come mostly from leg movements rather than wing movements. Other methods include less frequently used manoeuvres such as “sally” and “hang down”.
The location of the food items taken by the birds was considered as a foraging substrate and was further classified into: 1) trunk/main branches, with trunk and main branches of the tree supporting the canopy; this area consists largely of many epiphytes; 2) twigs, i.e., small branches < 3 cm in diameter to which leaves were attached and 3) ground, which includes the forest floor. The foraging location of the birds was divided into eight height classes based on vegetation strata, i.e., 0 m (ground), 0–2 m, 2.1–4 m (shrubs), 4.1–6 m, 6.1–8 m (sub-canopy), 8.1–10 m, 10.1–12 m (canopy) and > 12 m (above canopy). Markings indicating the height classes were made on trees for reference.
Feeding behavior recorded included species of tree where feeding occurred, features and size of the fruits, number of fruits ingested, duration of visit, number of birds feeding at the time and percentage of fruit cover on the particular tree.
In order to investigate the availability of food sources in the study area, phenology of the food plants and fruiting rates of the trees were studied in a 6-ha plot. Trees used by the Nilgiri Wood Pigeon were recorded from the first six months of observations. They accounted for 23 species, of which 10 individuals from each were selected and phenological events recorded every fortnight, following Frankie et al. (1974), Guy et al. (1979), Wheelwright (1988) and Maheswaran (2002). The phenological events were divided into vegetative and reproductive phases. Assigning values ranging from 0–100% for each phase, an approximate proportion of flowers, fruits and young leaves were recorded (Prasad and Hegde, 1986). Fruits were classified as ripe or unripe (Guy et al., 1979).
Faecal samples of the Nilgiri Wood Pigeon were collected mainly from nesting and feeding sites. Plant food items were often unidentifiable macroscopically, so microhistological methods were used based on plant epidermal resistance to herbivore digestion (Chapus, 1979). Prior to analysis, every faecal sample was mixed in water and cleaned by the addition of two drops of sodium hydrochloride. From this mixture a few drops were placed on a glass slide and viewed under the microscope. Epidermal tissues were identified by comparison with a reference collection of leaves, flowers, fruits and seeds of dominant trees and shrubs of the Kukkal shola.
Analysis of variance (ANOVA) was used to compare foraging methods of the Nilgiri Wood Pigeon in different seasons and Student’s t tests were performed to discover the significance in the differences of foraging heights of the Nilgiri Wood Pigeon. Regression analysis was employed to determine the relationship between fruiting phenology and environmental factors. Niche breadth of Nilgiri Wood Pigeon on each foraging dimension such as height, method and substrate was calculated using the Shannon-Wiener index (Shannon and Wiener, 1949). H' = –∑pilnpi (where H' = diversity and pi = the proportion of observation in subset i) as carried out by Recher (1985). These values were then converted to a standardized range using the formula: J' = H' / Hmax (where J' = niche breadth or specialization and Hmax = the H' value obtained when the observations were distributed equally across all subsets of the foraging dimension). J' values range between one and zero, with foraging specialization increasing as J' falls. It was used as an index of foraging specialization by Crome (1978) and Gokula and Vijayan (2000).
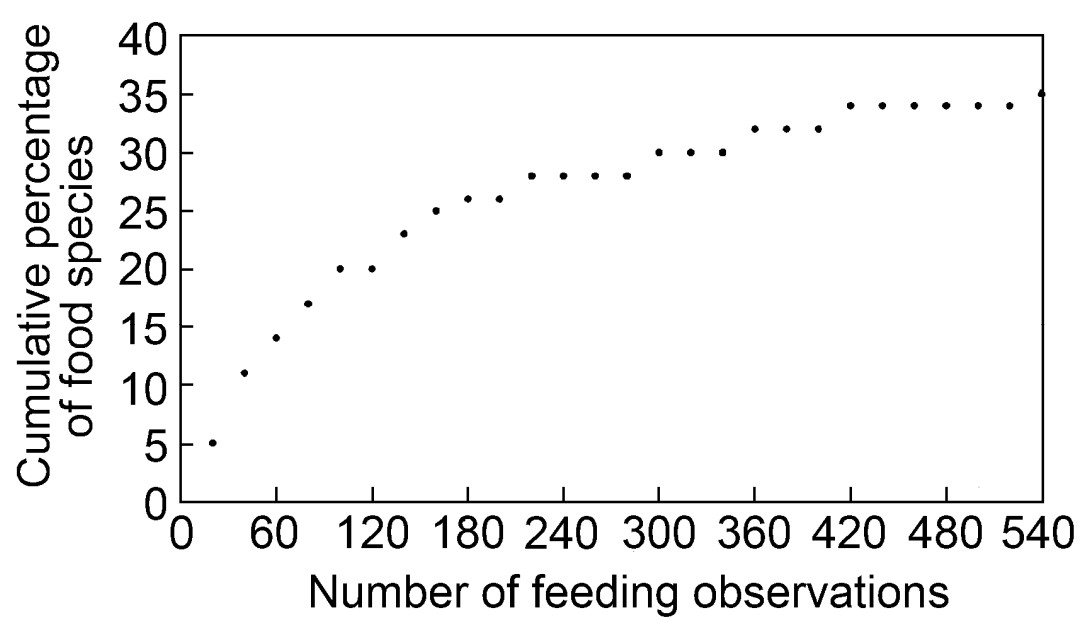
In all, 1520 foraging observations were made during the two-year study. The Nilgiri Wood Pigeon is primarily a frugivore, even though it feeds on leaf buds, flowers, ground invertebrates such as snails and coleopteran grubs and soil. The food of the Nilgiri Wood Pigeon include 39 species (Fig. 2; Appendix 1) belonging to 19 families of plants with fruits and flowers, leaf buds (four species) and three taxa of ground invertebrates. More than 47% of the food digested came from only nine species during all of the seasons and a few were common both in pre-breeding and breeding seasons. Lauraceae contributed the maximum number of species, i.e., 13. Myrsinaceae, Myrtaceae, Rosaceae and Rubiaceae contributed equally, three species each, while 14 species from other families were also used occasionally.
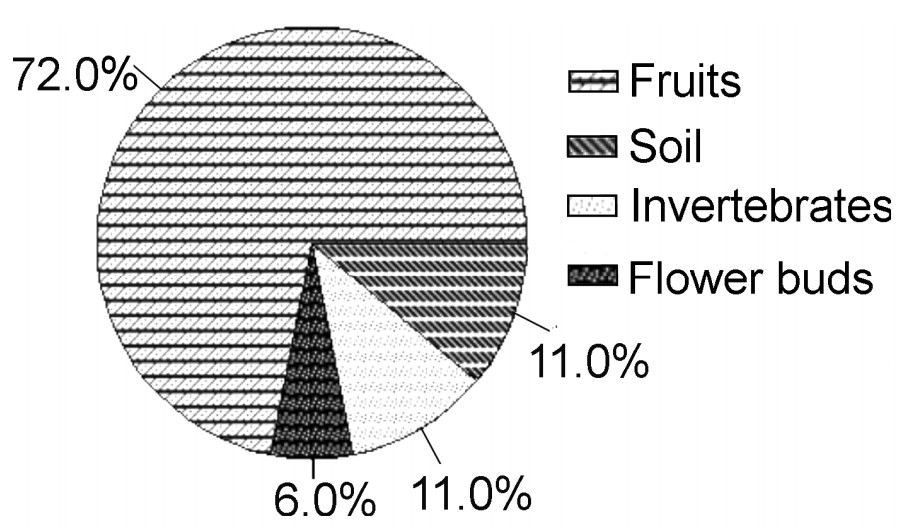
Based on direct feeding observations, more than 70% of food intake of the Nilgiri Wood Pigeon consisted of fruits, followed by soil, invertebrates and buds (Fig. 3). Fruit was the major food in all the seasons. Apart from the fruits, they were found feeding on soil during summer and on soil and flower buds during winter. Invertebrates were also consumed in sizable quantities during the monsoons (Table 1). Food composition was checked between seasons within a year and between the same seasons in the following year. It shows that there was no significant difference (F = 0.5, 0.8; p = 0.50).
| Food item | Summer | South-west monsoon | North-east monsoon | Winter | Total/means |
| Fruit | 71.5 | 72.5 | 70.0 | 70.0 | 71.0 |
| Flower buds | 4.5 | 1.5 | 7.0 | 12.5 | 6.4 |
| Invertebrates | 9.0 | 18.5 | 14.5 | 3.5 | 11.4 |
| Soil | 15.0 | 7.5 | 8.5 | 14.0 | 11.3 |
| Total No. of observations | 350 | 430 | 361 | 379 | 1520 |
 DownLoad:
CSV
DownLoad:
CSV
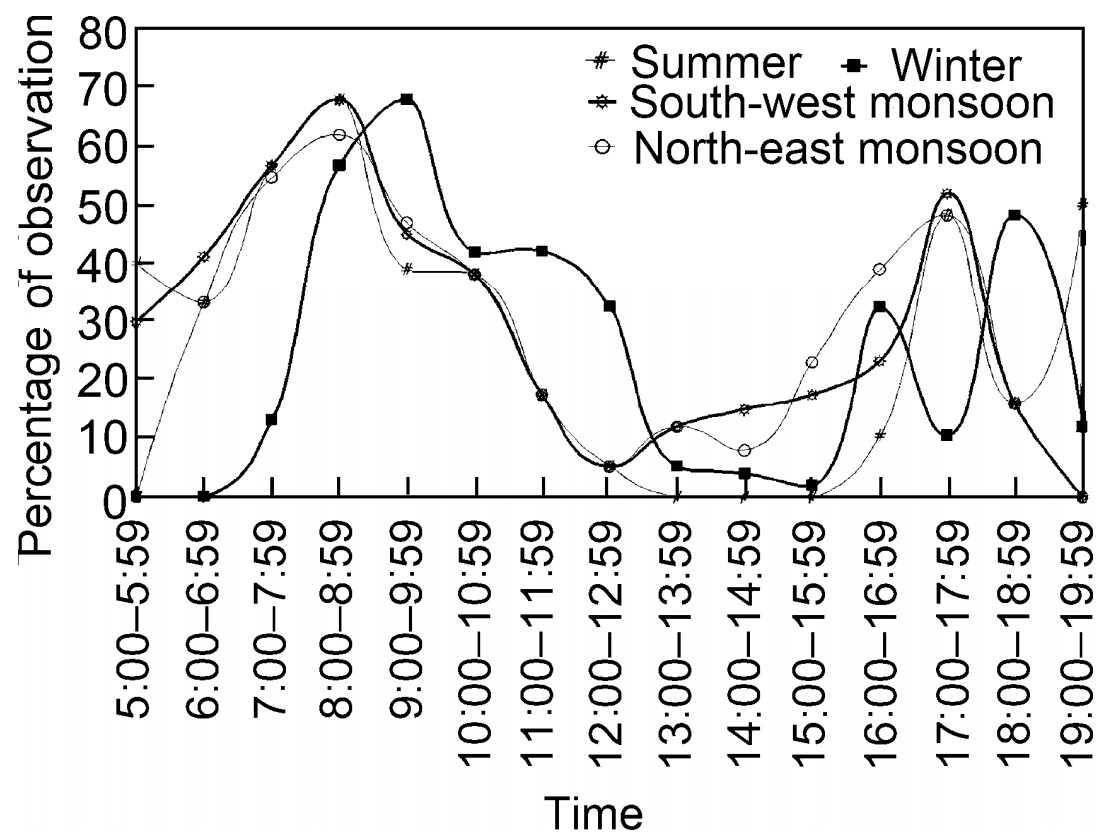
The Nilgiri Wood Pigeon started foraging from 05:00 hour and continued up to 18:00 hour with a major peak during the forenoon and sub-peak in the afternoon (Fig. 4). The major peak was between 08:00 and 08:59 hours in all seasons, except in the winter and the sub-peak occurred during the evening between 17:00–17:59 hours. In all the seasons, feeding was low or absent in the afternoon. In the north-east monsoon and winter, the peak was observed a little later based on environmental conditions. In summer, active feeding was observed after even 18:30 hour.
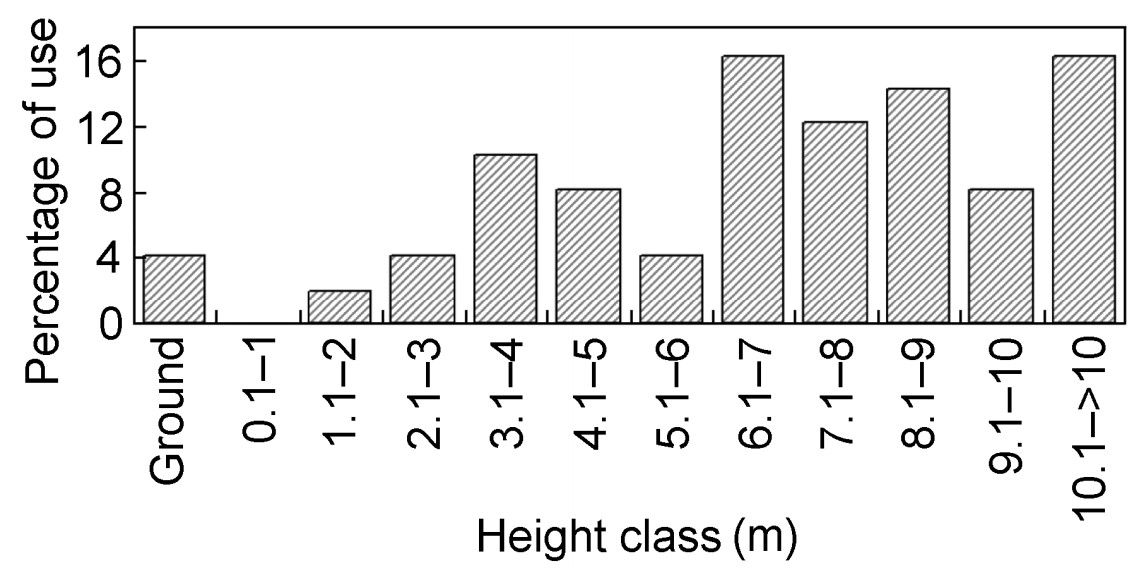
Vertical foraging locations of the Nilgiri Wood Pigeon were categorized into 12 levels (Fig. 5). Predominantly the middle and upper canopy height classes were used, with shrubs and sub-canopy the least. Maximum foraging took place in the upper canopy from 6 to 10 m and above (Fig. 5). The overall mean height utilized by the Nilgiri Wood Pigeon was 4.08±2.71 m. The ANOVA showed that foraging height of the Nilgiri Wood Pigeon varied significantly between the seasons (F = 15.85, p < 0.001).
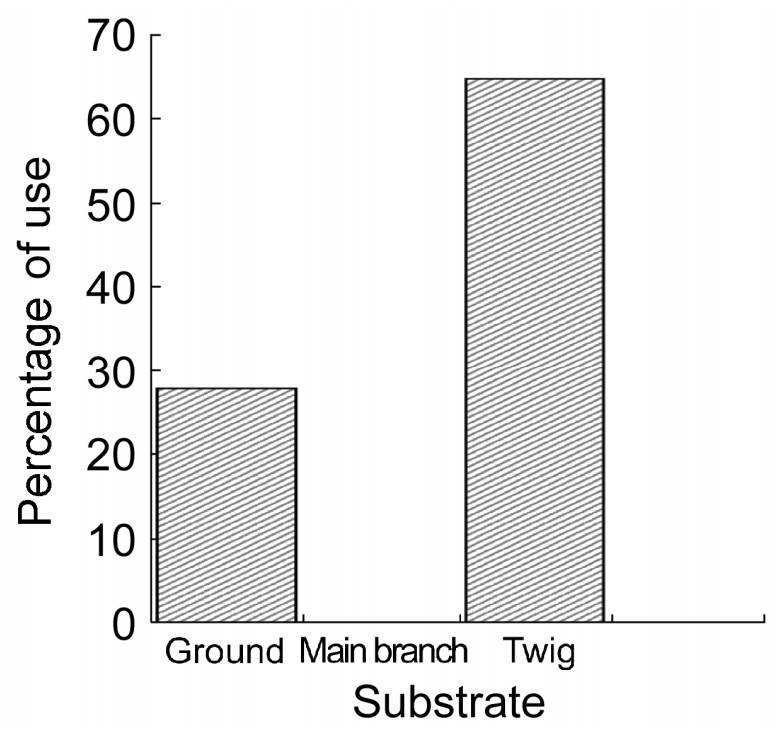
The Nilgiri Wood Pigeon is basically a canopy frugivore. However, occasionally, flocks of birds came, settled on the ground and started feeding on the fallen fruits, snails and soil. The major feeding site was twigs (64.95%) followed by the ground (27.85%) (Fig. 6). Foraging on the ground was maximum during the winter followed by the south-west monsoon. However, the level of use was not significantly different among the seasons (ANOVA; F = 2.95, p > 0.05). There was no record of feeding at the main branches and trunks, since most of the feeding occurred at the twigs where a large number of fruits were available.
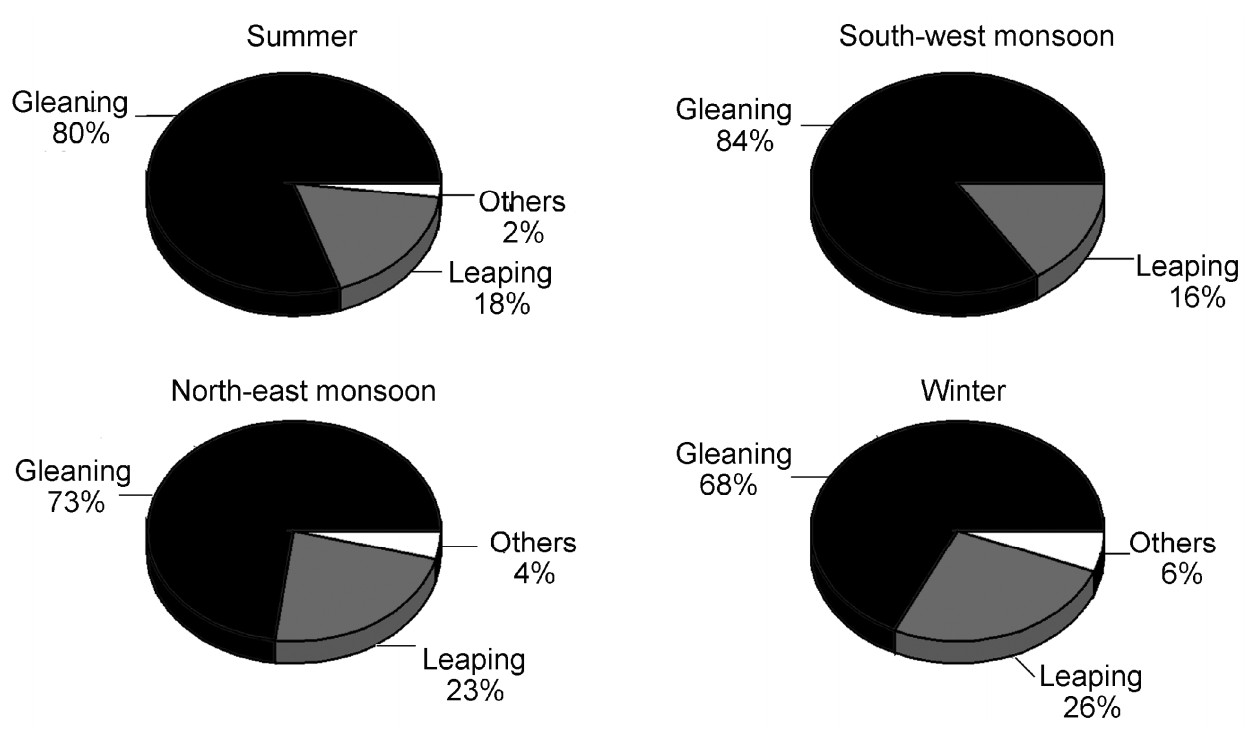
The Nilgiri Wood Pigeon used largely two foraging methods, i.e., gleaning and leaping. The former was more common, with frequencies varying from 68% in the winter to 84% during the south-west monsoon (Fig. 7). No significant difference was observed between the seasons within the year and between the years.
Niche breadth of the Nilgiri Wood Pigeon was calculated based on three dimensions, i.e., foraging substrate, method and height. The Nilgiri Wood Pigeon is more specialized in foraging method (0.19), followed by substrate (0.30). The mean value of J' in the three dimensions was 0.42. J' values range between one and zero, with foraging specialization increasing as the J' value falls.
One hundred and thirty four faecal samples of the Nilgiri Wood Pigeon were collected and analyzed during the two-year study. Of these, 30 samples contained only soil particles, 95 contained only seeds and the remaining plant matter such as tender shoots and flower buds and, as well, snail shells, while 11 species of seeds were identified from the faecal samples. The largest number of samples (17) contained seeds of Viburnum cylindricum, followed by Ilex wightiana (12), Casearia zeylanica, Olea paniculata, Beilschmiedia wightii and Daphniphyllum neilgherrense (Table 2). Seeds found intact in the faecal pellets were considered to be fit for dispersal (Table 2). Fruits were identified by seeds as well as by epicarp or skin, present in faecal matter. Seeds up to 35 mm in size were intact and hence dispersible, whereas those of 50 to 125 mm were slightly damaged and hence, considered non-dispersible.
| Species | Occurrence (%) |
Dispersible (%) |
| Viburnum cylindricum | 17.89 | 25 |
| Ilex wightiana | 12.63 | 100 |
| Casearia zeylanica | 9.47 | 0 |
| Olea paniculata | 11.58 | 0 |
| Daphniphyllum neilgherrense | 9.47 | 0 |
| Ficus drupacea | 8.42 | 100 |
| Beilschmiedia wightii | 6.32 | 25 |
| Syzygium densiflorum | 5.26 | 0 |
| Syzygium lanceolatum | 9.47 | 0 |
| Syzygium tamilnadensis | 5.26 | 0 |
| Maesa indica | 4.21 | 75 |
DownLoad:
CSV
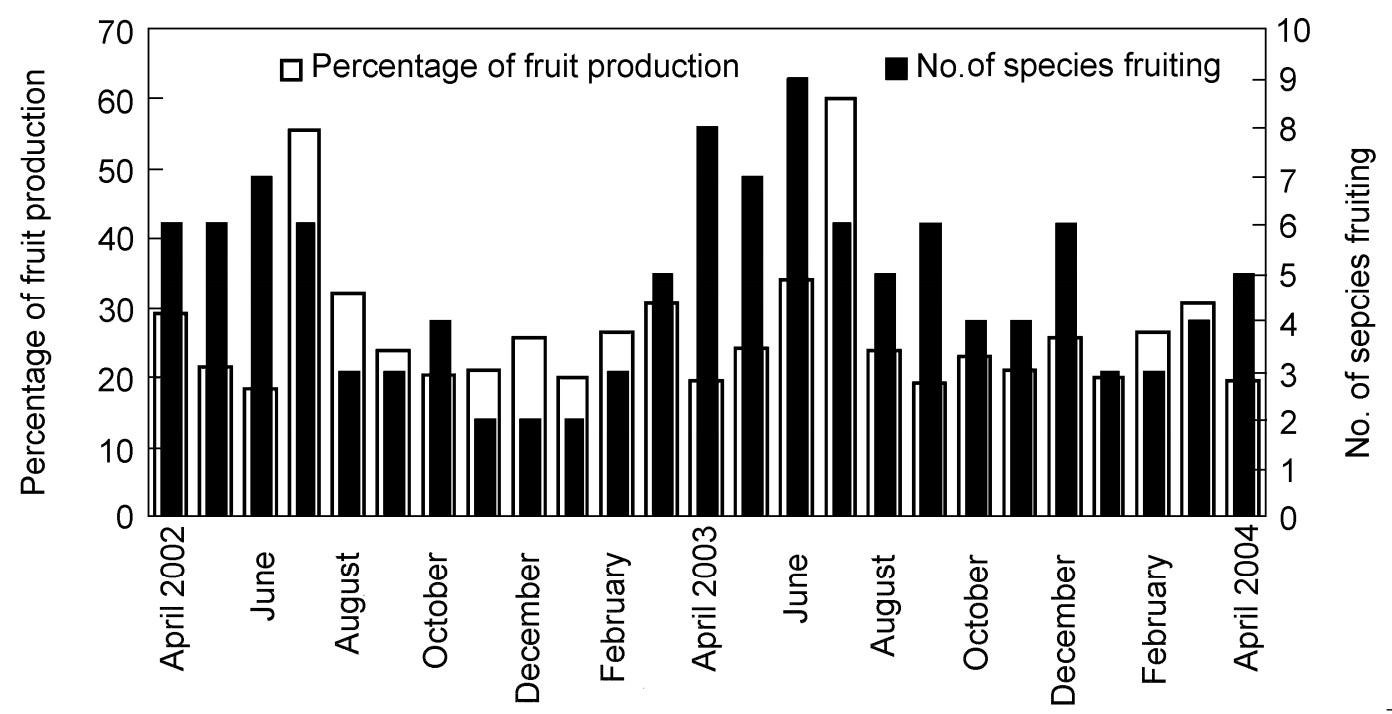
The phenology of 39 tree species was observed during the two-year field study from April 2002 to May 2004. Fruiting was at a peak in July 2003 and least in June 2002 (Fig. 8). During the peak period, fruits of 85 individual trees of six species were consumed by the Nilgiri Wood Pigeon. Syzygium tamilnadensis, Ilex wightiana and Beilschmiedia wightii fruited only once during the two years of study. In August, only Trichillia connoroides were in fruit, whereas in December only Olea paniculata and Maesa indica had fruits, and Maesa indica contributed only 10% of the total fruit production. The number of fruiting species appeared to have no correlation with rainfall (r = 0.26, p = 0.2) while fruit abundance did show a correlation (r = 0.40, p < 0.05). Out of the 23 species, fruits of only eight species were mostly consumed in different seasons.
The peak fruit abundance had a significant correlation with rainfall (r = 0.40, p < 0.05), while there was no correlation between fruit abundance and the number of species in fruit (r = 0.22, p = 0.2), fruit abundance and the number of rainy days (r = 0.16, p = 0.44). The maximum number of species was in fruit only when the number of rainy days was less than six. Total fruit production did not correlate with the monthly mean maximum temperature, while the number of species in fruit was significantly correlated (r = 0.39, p < 0.05). Minimum temperature and number of species fruiting also showed a similar trend (r = 0.58, p < 0.01). Total fruit production and relative humidity were not correlated, whereas the number of species in fruit was significantly correlated with humidity (r = 0.32, p = 0.05).
The Nilgiri Wood Pigeon is largely a frugivorous bird, with its animal component (invertebrates) in the diet small enough to be neglected in our analyses. It used 34 species of plants for fruits, buds and flowers and three taxa of invertebrates. Such diverse diet has been reported in various species of pigeons (Neff, 1947; Crome, 1975a; McEwen, 1978; Zino and Zino, 1986; Snow and Snow, 1988; Pearson and Climo, 1993; Bancroft and Bowman, 1994; Brooke and Jones, 1995; Oliveira and Jones, 1995; Powlesland et al., 1997; Hernandez et al., 1999; Martín et al., 2000; Oliveira et al., 2002).
Direct observation of feeding, widely used to assess pigeon diets elsewhere (Snow and Snow, 1988; Rosenberg and Cooper, 1990; Oliveira et al., 2002), showed that the diet of the Nilgiri Wood Pigeon contained principally fruits while other items were less frequent as secondary diet, observed in several tropical frugivores (Kannan and James, 1999). Recent studies on the nutritional value of fruits and frugivoros birds indicate that most of the birds are unable to subsist on an exclusive diet of fruits (Herrera, 1984), since fruits have low protein content. Hence, the frugivores were moving on to mixed diet with invertebrates (Levey and Karasov, 1992). The proportion of invertebrate diet may vary depending upon the energy requirements of a species, such as breeding and moulting (Kannan and James, 1999). In summer, most of the rain forest birds start breeding and once the breeding is over they undergo moulting and hence, during this period they need protein-rich food such as invertebrates. The behavior of the Nilgiri Wood Pigeon is the same. Pigeon diets contain soil and grit (Griminger, 1983; Ali and Ripely, 1987; Jarvis and Passmore, 1992; Sanders and Jarvis, 2000; Dhondt and Hochachka, 2001). Frequency of feeding on soil was high in the breeding season, when they require more calcium for eggshell and crop milk production (March and Sadleir, 1975; Griminger, 1983; Sanders and Jarvis, 2000). The present observations on the diet of the Nilgiri Wood Pigeon confirm earlier findings on the pigeons. Nutrient intake would be better balanced if the birds were to feed on several fruit species, rather than on a single species, since a mixed diet increases the possibility of extracting specific nutrients (Jordano, 1992).
The vertical feeding strata varied according to the abundance of fruit. However, the Nilgiri Wood Pigeon prefers the upper canopy as do the rain forest pigeons of Australia (Crome, 1975a, 1975b). The Nilgiri Wood Pigeon fed mostly on the edges of the tree canopy mainly because of the availability of fruits and foliage cover to avoid predators. It should be mentioned that tropical forest trees contain fruits along their small axes (twigs). During food scarcity, flocks of birds feed on wind fallen fruits and snails as noted by Ali and Ripely (1987). Apart from this, the ground stratum was used to feed on invertebrates or soil to compensate nutritional and calcium requirements during the breeding season. This is especially true for the development of the gonad, the egg shell (Kannan and James, 1999) and the production of crop milk in pigeons (March and Sadleir, 1975; Griminger, 1983; Sanders and Jarvis 2000).
The foraging method used is important in determining the size of the food consumed (Remsen and Robinson, 1990). As in many other tropical bird species, the Nilgiri Wood Pigeon prefers the gleaning method for foraging (Innis, 1989; Recher et al., 1995). Ford et al. (1986) found that more than 50% of bird species in Australia used gleaning as the predominant method, since it involves the least energy expenditure (Moermond and Denslow, 1983; Remsen and Parkerii, 1984).
Faecal analysis, a more reliable evidence of food consumption (McEwen, 1978), shows that the Nilgiri Wood Pigeon feeds on fruits of different sizes, leaf buds, flowers, snails, coleopteran grubs and soil. Eighty five samples showed that the Nilgiri Wood Pigeon acted as a seed predator for many plants, since the seeds were crushed. A sizeable number of intact seeds were also obtained, suggesting that they also act as a disperser for a large number of plant species. The pigeon has a muscular gizzard adapted for grinding food and a long narrow gut, unlike some genera (example; Ptilinopus, and Ducula; Goodwin, 1983). However, in the pellets of the Nilgiri Wood Pigeon, intact seeds were noticed as in the pellets of the Maderia Laurel Pigeon (Columba trocaz) (Oliveira et al., 2002).
Plant phenological studies are fundamental to understand the forest as a resource base for dependent species, populations and communities. Tropical plant communities display conspicuous seasonal patterns in vegetative and reproductive phenophases at both community and species levels (Williams-Linera, 2003). Hence, we emphasized phenological studies at Kukkal. Ripe fruits were present the year round with a peak in July in both years, after the onset of the south-west monsoon. Similar patterns were reported in many tropical forests (Sun et al., 1997; Kannan and James, 1999; Griz and Machado, 2001; Kimura et al., 2001; Bleher et al., 2003; Sundarapandian et al., 2005). After the heavy rains, the number of species fruiting decreased, hence also total fruit production. In the drier months, fruit abundance was mainly due to Olea paniculata and Daphniphyllum neilgherrense, which served as keystone species for the Nilgiri Wood Pigeon. Rainfall had a significantly positive correlation with fruiting at Kukkal, as also reported for several other tropical forests (Murali and Sukumar, 1993; Kannan and James, 1999; Sundarapandian et al., 2005). The total monthly rainfall and the number of rainy days, when considered together, was only weakly correlated with the number of species fruiting, whereas total fruit production was significantly correlated. In the case of dry forests, total rainfall and number of rainy days stimulated fruiting (Murali and Sukumar, 1993; Sundarapandian et al., 2005), whereas in the case of wet, montane temperate forests, as in Kukkal, fruiting was close to the onset of rainfall or at the onset of an early rainy season (Sun et al., 1997; Griz and Machado, 2001; Kimura et al., 2001). These phenomena serve to enhance dispersal, escape predation and avoid pathogen infection (Prasad and Hegde, 1986; Sundarapandian et al., 2005).
The total number of species in fruit at Kukkal was significantly correlated with both maximum and minimum temperatures. However, in the wet evergreen forests of the southern Western Ghats, this number was only correlated with minimum temperature (Ganesh et al., 1996). The number of species in fruit was significantly correlated with relative humidity. But total fruit production had only a weak correlation with relative humidity as in the evergreen forests in the southern Western Ghats (Ganesh et al., 1996).
A universal tendency for tropical forests to fluctuate seasonally in fruit production with abundance peaks and periods of scarcity has been established in South-east Asia (Leighton and Leighton, 1983; Kannan and James, 1999; Maheswaran, 2002). Fruiting plants that provide a critical resource for frugivores during periods of scarcity are called keystone plant resources (Leighton and Leighton, 1983; Terborgh, 1986a, 1986b). They are of great ecological importance, because they appear to establish the carrying capacity of the frugivores (Terborgh, 1986a, 1986b). In most tropical forests, the Moraceae family, Ficus spp., act as keystone species (Kannan and James, 1999; Maheswaran, 2002), whereas in the montane forest only a single species of Ficus (Ficus drupacea), was present and its fruiting did not coincide with the lean season. In the montane shola forest, its place is taken by Olea paniculata (Oleaceae) and Trichillia connoroides (Meliaceae), which act as keystone species. Meliaceae was identified as a keystone resource for hornbills in the lowland rainforests of Borneo (Leighton and Leighton, 1983; White, 1994). Lauraceae and Daphniphyllaceae are plant families with lipid-rich fruits. These species act as an important source of food during the breeding season for various frugivores in India (Kannan and James, 1999; Maheswaran, 2002). In most of the Lauraceae family, fruiting was observed from April to June, which coincided with the breeding of birds (Powlesland et al., 1997). Frugivores that rely on these keystone resources may be vulnerable to changes in fruit supply resulting from the effects of climatic change on plant phenology (Corlett and LaFrankie, 1999), increased deforestation and fragmentation (Bleher et al., 2003).
Frugivorous birds move and migrate, tracking fruit sources (Leighton and Leighton, 1983; van Schaik et al., 1993). In temperate regions, migratory frugivorous birds which have finished breeding in the north arrive when fruits are abundant (Herrera, 1984). In subtropical and tropical regions, where most of the temperate migrants overwinter, fruiting occurs during this period. However, these migrants also move locally, tracking food sources (Loiselle and Blake, 1991). Local movement of resident frugivorous birds within their habitats has already been mentioned (Leighton and Leighton, 1983; Loiselle and Blake, 1991; van Schaik et al., 1993), although the destinations, seasonality and periodicity have not been clarified for the most part. In South-east Asia, tropical montane areas have been observed as destinations for migratory frugivorous birds (Corlett, 1998). Leighton and Leighton (1983) reported that the number of inhabiting hornbills and pigeons decreased with decreasing numbers of fruiting species in a lowland forest of Borneo. They suggested montane regions as one of the destinations of the disappearing birds. During the prolonged periods of dryness and food scarcity, Columbiforms might easily disperse to neighboring forests in the ecotone of the dry and moist zones (Rivera-Milan, 1992). Wells (1985) noted that in the Greater Sunda region, frugivorous birds including temperate migrants, often flocked to montane regions and suggested that fruit resource conditions might be more favorable in montane regions. However, we know of no reliable study associating seasonal changes in fruiting conditions with seasonal dynamics of frugivorous birds in montane regions of South-east Asia (Corlett, 1998).
The resource use pattern suggests that the Nilgiri Wood Pigeon, which lives in a limited area and habitat, are flexible and exploit each food resource as available. This flexibility in diet is demonstrated by the strong seasonal changes in niche breadth. The optimal foraging theory predicts “less energy spent with maximum gain” (Goldstein, 1990). Hence, selection of foraging method is important for long-term survival of any species. Niche breadth of the Nilgiri Wood Pigeon narrows with their diets in response to high availability of fruits and they broaden their diet to include other plant parts when fruits are scarce. These observations suggests that the diet of the Nilgiri Wood Pigeon is quite flexible, adapted to exploit the available resources as in the Maderia Laurel Pigeon (Oliveira et al., 2002). Such evolutionary adaptations make the species more successful, although it is a habitat specialist. This flexibility is shown within their given restricted habitat.
During low fruit abundance in the study area, the Nilgiri Wood Pigeon moved to other areas where fruits were abundant (personal observation). Frequency of their sightings was significantly correlated with fruit abundance. Fruit eating birds generally have to deal with strong spatio-temporal patterns that will affect the way they use their habitat (Herrera, 1984), while suitable changes in resources such as berry density may promote changes in habitat use (Cody, 1985). Seasonal patterns were observed in the diet of Maderia Laurel Pigeon; when total abundance of fruits in the forest decreased, birds shifted from trees to the ground and shrub layers (Oliveira et al., 2002). Innis (1989) showed that foraging habits of pigeons in subtropical forests in Australia were largely opportunistic; the birds used whatever fruits were available. Crome (1975b) identified the same pattern for several Australian columbid species. Usually, the Nilgiri Wood Pigeon forages in flocks, varying from 6 to 24 individuals and sometimes as many as 45. Such flocking for feeding has also been reported in other tropical pigeons (Recher and Date, 1988), since flocking may reduce predators.
The findings of the present study on the Nilgiri Wood Pigeon provide an understanding of the relationship between this endemic, threatened species and its relict forest habitat. Conservation of the species depends on preserving all habitat types, required to meet their resource needs throughout the year. Habitat loss and degradation are the major threats to this species. Moreover, the knowledge emanated from the study that pigeons use many flowering plants which occur more abundantly on the edges of the forest can be used to redefine the conservation value of these areas. Illegal hunting and lack of awareness of this species are other threats to the species as in other pigeons (Crome, 1975b; Oliveira et al., 2002).
We thank V.S. Vijayan, founder director, Sálim Ali Centre for Ornithology and Natural History for encouragement. This study was conducted as a part of a project on endemic birds in the Western Ghats, funded by the Ministry of Environment and Forest, Government of India (23–1/2001–RE). We are grateful to the Chief Wildlife Warden of the Tamil Nadu Forest Department and the DFO and other staff of the Kodaikanal forest division for their support in the field.
| Species | Family |
| Psychotria nilgiriensis var. astephana (Hook. f.) Deb & Gang. | Rubiaceae |
| Maesa indica (Roxb.) DC. | Myrsinaceae |
| Phoebe paniculata Nees. | Lauraceae |
| Neolitsea cassia (L.) Kosterm. | Lauraceae |
| Lasianthus acuminatus Wight. | Rubiaceae |
| Beilschmiedia wightii (Nees) Benth. ex Hook. f. | Lauraceae |
| Turpinia nepalensis Wall. & Wight & Arn. | Staphyleaceae |
| Symplocos cochinchinensis (Lour.) Moore subsp. laurina (Retz.) | Symplocaceae |
| Viburnum cylindricum Buch.-Ham. ex D. Don | Caprifoliaceae |
| Daphniphyllum neilgherrense Thw. | Euphorbiaceae |
| Vaccinium neilgherrense Wight. | Vacciniaceae |
| Casearia zeylanica Thwaites | Flacourtiaceae |
| Ardisia rhomboidea Wight. | Myrsinaceae |
| Olea paniculata R. Br. | Oleaceae |
| Syzygium densiflorum Wall. ex Wight & Arn. | Myrtaceae |
| Myrsine wightiana (Wall. ex DC) | Myrsinaceae |
| Syzygium tamilnadensis Rathakr. & Chithra nom. | Myrtaceae |
| Litsea glabrata (Wall. ex Nees) Hook. f. | Lauraceae |
| Eurya nitida Korth. | Theaceae |
| Neolitsea scrobiculata (Meisner) Gamble. | Lauraceae |
| Cassine paniculata (Wight & Arn.) Lobr.-Callen | Celastraceae |
| Mahonia leschenaultii (Wall. ex Wight & Arn) | Berberidaceae |
| Cinnamomum perrottetii Meisner | Lauraceae |
| Litsea wightiana (Nees) Hook. f. | Lauraceae |
| Trichilia connaroides (Wight & Arn.) Bent var. connaroides | Meliaceae |
| Neolitsea fischeri Gamble. | Lauraceae |
| Rubus ellipticus Smith. | Rosaceae |
| Cinnamomum malabatrum (Burm. F.) Blume | Lauraceae |
| Cinnamomum wightii Meisner | Lauraceae |
| Coffea arabica L. | Rubiaceae |
| Cryptocarya bourdillonii Gamble | Lauraceae |
| Cryptocarya neilgherrensis Meisner | Lauraceae |
| Litsea floribunda (Blume) Gamble | Lauraceae |
| Photinia integrifolia Lindl. var. sub-lanceolata Miq. | Rosaceae |
| Syzygium lanceolatum (Lam.) Wight & Arn. | Myrtaceae |
| Elaeocarpus glandulosus Wall. | Elaeocarpaceae |
| Ficus drupacea Thunb. | Moraceae |
| Rubus racemosus Roxb. | Rosaceae |
| Ilex wightiana | Aquifoliaceae |
DownLoad:
CSV
|
Ali S, Ripley SD. 1987. Handbook of the birds of India and Pakistan, compact edition. Oxford University Press, Delhi
|
|
BirdLife International. 2001. Threatened Birds of the World. Lynx Edicions and BirdLife International, Barcelona and Cambridge
|
|
Blake JG, Loiselle BA., Moermond TC, Levey DJ, Denslow JS. 1990. Quantifying abundance of fruits for birds in tropical habitats. Stud Avian Biol, 13:71–77
|
|
Brooke MDEL, Jones P. 1995. The diet of the Henderson Fruit Dove Ptilinopus insularis. I. Field observations of fruit choice. Biol J Linn Soc, 5656:149–165
|
|
Champion HG, Seth SK. 1968. A Revised Survey of the Forest Types of India. Government of India Press, New Delhi
|
|
Chapus JL. 1979. Evolution saisanniere du regime alimentaire d’Oryctolagus cuniculus dans differents types d’habitats. en France. In: Mayers K, Maclnnes CD (eds) Proceedings of the World Lagomorph Conference. University of Guelph. on Canada, pp 743–761
|
|
Cody ML. 1985. Habitat Selection in Birds. Academic Press, New York
|
|
Corlett RT, LaFrankie JV. 1999. Potential impacts of climate change on tropical Asian forests through an influence on phenology. Clim Change, 39:439–453
|
|
Ganesh T, Ganesan R, Soubadradevy M, Davidar P, Bawa KS. 1996. Assessment of plant biodiversity at a mid-elevation evergreen forest of Kalakad-Mundanthuari Tiger reserve, Western Ghats, India. Curr Sci, 71:379–392
|
|
Gibbs D, Barnes E, Cox J. 2001. Pigeons and Doves of the World. Pica Press, Mountifield, UK
|
|
Gokula V, Vijayan L. 2000. Foraging pattern of birds during the breeding season in thorn forest of Mudumalai wildlife sanctuary, Tamil Nadu, South India. Trop Ecol, 41:195–208
|
|
Goldstein DL. 1990. Energetics of activity and free living in birds. Stud Avian Biol, 13:423–426
|
|
Goodwin D. 1983. Pigeons and Doves of the World, 3rd edn. Cornell University Press, New York
|
|
Griminger P. 1983. Digestive system and nutrition. In: Abs M (ed) Physiology and Behaviour of the Pigeon. Academic Press, San Francisco, pp 19–39
|
|
Grimmett R, Inskipp C, Inskipp T. 1998. Birds of the Indian Subcontinent. Oxford University Press, Delhi
|
|
Guy PR, Mahlangu ZA, Claridza H. 1979. Phenology of some trees and shrubs in the Sengwa Wildlife Research Area, Zimbabwe-Rhodesia. S Afr J Wildl Res, 9:47–54
|
|
Hernandez MA, Martin A, Nogales M. 1999. Breeding success and predation on artificial nests of the endemic pigeons Bolles Laurel Pigeon Columba bollii and White-tailed Laurel Pigeon Columba junoniae in the laurel forest of Tenerife (Canary islands). Ibis, 141:52–59
|
|
Howe HF. 1986. Seed dispersal by fruit-eating birds and mammals. In: Murray DR (ed) Seed Dispersal. Academic Press, New York, pp 123–190
|
|
Inskipp T, Lindsey N, Duckworth W. 1996. An Annotated Checklist of the Birds of the Oriental Region. Oriental Bird Club, Sandy, Bedfordshire, UK
|
|
Jarvis RL, Passmore MF. 1992. Ecology of Band-tailed Pigeons in Oregon. U.S. Dept Inter. Fish and Wildl. Serv., Rep. 6. Washington DC
|
|
Jordano P. 1992. Fruits and Frugivory. In: Fenner M (ed) Seeds: The Ecology of Regeneration in Plant Communities. CAB International, G. Britain, pp 105–156
|
|
Kannan R, James DA. 1999. Fruiting phenology and the conservation of the Great Pied Hornbill (Bucerors bicornis) in the Western Ghats of Southern India. Biotropica, 31:167–177
|
|
Kimura K, Yumoto T, Kikuzawa K. 2001. Fruiting phenology of fleshy-fruited plants and seasonal dynamics of frugivorous birds in four vegetation zones on Mt. Kinabalu, Bo rneo. J Trop Ecol, 17:833–857
|
|
Lack D. 1968. Ecological Adaptations for Breeding in Birds. Methuen, London
|
|
Leighton M, Leighton DR. 1983. Vertebrate response to fruiting seasonality within a Bornean rain forest. In: Sutton SL, Whitemore TC, Chadwick AC (eds) Tropical Rain Forests: Ecology and Management. Blackwell Scientific Publications, Oxford, pp 181–209
|
|
Levey DJ, Karasov WH. 1992. Digestive modulation in a seasonal frugivore, the American Robin (Turdus migratorius). Am J Physiol, 262:G711–G718
|
|
Maheswaran B. 2002. Habitat utilization by Malabar Grey Hornbill at Mudumalai Wildlife Sanctuary, Western Ghats. Ph.D thesis. Bharathiar University, Coimbatore
|
|
Martin A, Hernandez MA, Lorenzo JA, Nogales M, Gonzalez C. 2000. Las palomas endemicas de Canarias. Consejeria de Gobierno de Canarias and SEO/Birdlife. Santa Cruz de Tenerife, Spain (in Spanish)
|
|
Mathew KM. 1996. Illustrations on the Flora of the Palni Hills. The Rapinat Herbarium, St. Joseph’s College, Tiruchirapalli, India
|
|
McEwen WM. 1978. The food of the New Zealand Pigeon (Hemiphaga novaeseelandiae novaeseelandiae). New Zealand J Ecol, 1:99–108
|
|
Murali KS, Sukumar R. 1993. Reproductive phenology of a tropical dry forest in Mudumalai, Southern India. J Ecol, 82: 759–767
|
|
Neff JA. 1947. Habitats, food and endemic status of the Band-tailed Pigeon USDI Fish and Wildlife Service. North American Fauna 58. Washington, DC
|
|
Oliveira P, Jones M. 1995. Population numbers habitat preferences and the impact of the Longtoed Pigeon Columba trocaz on agriculture. Boletim do Museu Municipal do Funchal 4:531–542.
|
|
Pearson PE, Climo GC. 1993. Habitat use by Chatham Island Pigeons. Notornis, 40:45–54
|
|
Powlesland RG, Dilks PJ, Flux A, Grant AD, Tisdall CJ. 1997. Impact of food abundance, diet and food quality on the diet and food quality on the breeding of the Fruit pigeon Parea hemiphaga novaeseeladiae chathamensis on Chatham island, New Zealand. Ibis, 139:353–365
|
|
Prasad SN, Hedge M. 1986. Phenology and seasonality in the tropical deciduous forest of Bhandipur, South India. Proc Indian Acad Sci Plant Sci, 96:121–133
|
|
Ralph CP, Nagata SE, Ralph CJ. 1985. Analysis of droppings to describe diets of small birds. J Field Ornithol, 56:165–174
|
|
Ramesh BR, Pascal JP. 1998. Atlas of Endemics of the Western Ghats (India). Institut Francais De Pondichery, India
|
|
Rawat GS, Karunakaran PV, Uniyal VK. 2003. Shola Grasslands of Western Ghats – Conservation status and management needs. ENVIS Bull Grassland Ecosyst Agroforest, 1(1):57–64
|
|
Recher HF, Date EM, Ford HA. 1995. The biology and management of rainforest pigeons in New South Wales. Species management report No 16. National parks and wildlife services (NWS), Hurstville
|
|
Recher HF. 1985. Synthesis: A model of forest and woodland bird communities. In: Keast A, Recher HF, Saunders D (eds) Birds of the Eucalypt Forests and Woodlands: Ecology, Conservation and Management. Surrey-Beatty, Sydney, pp 129–135
|
|
Recher HF, Date EM. 1988. Distribution and abundance of rainforest pigeons in New South Wales. Report to National parks and wildlife services (NWS)
|
|
Remsen JV, Robinson SK. 1990. A classical scheme for the foraging behaviour of birds in terrestrial habitats. Stud Avian Biol, 13:144–160
|
|
Robinet O, Barre N, Salas M. 2003. Population estimate for the Ouvea Parakeet Eunymphicus cornutus uvaeensis: its present range and implications for conservation. Emu, 96: 151–157
|
|
Rosenberg KV, Cooper RJ. 1990. Approaches to avian diet analysis. Stud Avian Biol, 13:80–90
|
|
Shannon CE, Wiener W. 1949. The Mathematical Theory of Communication. University of Illinois Press, Urbana, USA
|
|
Snow B, Snow D. 1988. Birds and Berries. T & AD Poyser, London
|
|
Somasundaram S, Vijayan L. 2008. Foraging behavior and guild structure of birds in montane wet temperate forest of the Palni Hills, South India. Podoces, 3(1/2):79–91
|
|
Somasundaram S, Vijayan L. 2004. Avifauna of Palni Hills: a conservation perspective. In: Muthuchelian K (ed) Proceeding of National Workshop on “Biodiversity Resources Management and Sustainable Use”. Centre for Biodiversity and Forest Studies, School of Energy, Environment and Natural Resources, Madurai Kamraj University, Madurai, India, pp 318–322
|
|
Sundarapandian SM, Chanderasekaran S, Swamy PS. 2005. Phenological behaviour of selected tree species in tropical forests at Kodayar in the Western Ghats, Tamil Nadu, India. Curr Sci, 88:805–810
|
|
Terborgh J. 1986a. Keystone plant resource in tropical forest. In: Soule ME (ed) Conservation Biology: The Source of Scarcity and Diversity. Sinaur Associates, Sunderland, Massachusetts, pp 330–344
|
|
Terborgh J. 1986b. Community aspects of frugivory in tropical forests. In: Estradon A, Fleming T (eds) Frugivores and Seed Dispersal, Junk, Pordrecht, The Netherlands, p 371
|
|
Thiollay JM. 1989. Area requirements for the conservation of rain forest raptors and game birds in French Guiana. Conserv Biol, 3:128–137
|
|
van Schaik, Terborgh CPJ, Wright SJ. 1993. The phenology of the tropical forests, adaptive significance and consequences for primary consumers. Annu Rev Ecol Syst, 24: 353–377
|
|
Wells DR. 1985. The forest avifauna of western Malesia and its conservation. Intl Council Bird Preserv Tech Publ, 4: 213–232
|
|
Wheelwright NT. 1988. Fruit-eating birds and bird dispersed plants in the tropics and temperate zone. Tree, 3(10):270–274
|
|
White LJT. 1994. Patterns of fruit-fall phenology in the Lopé Reserve, Gabon. J Trop Ecol, 10:289–312
|
|
Williams-Linera G. 2003. Phenology of deciduous and broad- leaved-evergreen tree species in a Mexican tropical lower montane forest. Global Ecol Biogeogr Lett, 6:115–127
|
|
Zino E, Zanio PA. 1986. An account of the habitat, feeding habits, density, breeding and need of protection of the Long-toed Pigeon, Columba trocaz. Bocagiana, 88:1–16
|
Figures(8) / Tables(3)





| Food item | Summer | South-west monsoon | North-east monsoon | Winter | Total/means |
| Fruit | 71.5 | 72.5 | 70.0 | 70.0 | 71.0 |
| Flower buds | 4.5 | 1.5 | 7.0 | 12.5 | 6.4 |
| Invertebrates | 9.0 | 18.5 | 14.5 | 3.5 | 11.4 |
| Soil | 15.0 | 7.5 | 8.5 | 14.0 | 11.3 |
| Total No. of observations | 350 | 430 | 361 | 379 | 1520 |
DownLoad:
CSV
| Species | Occurrence (%) |
Dispersible (%) |
| Viburnum cylindricum | 17.89 | 25 |
| Ilex wightiana | 12.63 | 100 |
| Casearia zeylanica | 9.47 | 0 |
| Olea paniculata | 11.58 | 0 |
| Daphniphyllum neilgherrense | 9.47 | 0 |
| Ficus drupacea | 8.42 | 100 |
| Beilschmiedia wightii | 6.32 | 25 |
| Syzygium densiflorum | 5.26 | 0 |
| Syzygium lanceolatum | 9.47 | 0 |
| Syzygium tamilnadensis | 5.26 | 0 |
| Maesa indica | 4.21 | 75 |
DownLoad:
CSV
| Species | Family |
| Psychotria nilgiriensis var. astephana (Hook. f.) Deb & Gang. | Rubiaceae |
| Maesa indica (Roxb.) DC. | Myrsinaceae |
| Phoebe paniculata Nees. | Lauraceae |
| Neolitsea cassia (L.) Kosterm. | Lauraceae |
| Lasianthus acuminatus Wight. | Rubiaceae |
| Beilschmiedia wightii (Nees) Benth. ex Hook. f. | Lauraceae |
| Turpinia nepalensis Wall. & Wight & Arn. | Staphyleaceae |
| Symplocos cochinchinensis (Lour.) Moore subsp. laurina (Retz.) | Symplocaceae |
| Viburnum cylindricum Buch.-Ham. ex D. Don | Caprifoliaceae |
| Daphniphyllum neilgherrense Thw. | Euphorbiaceae |
| Vaccinium neilgherrense Wight. | Vacciniaceae |
| Casearia zeylanica Thwaites | Flacourtiaceae |
| Ardisia rhomboidea Wight. | Myrsinaceae |
| Olea paniculata R. Br. | Oleaceae |
| Syzygium densiflorum Wall. ex Wight & Arn. | Myrtaceae |
| Myrsine wightiana (Wall. ex DC) | Myrsinaceae |
| Syzygium tamilnadensis Rathakr. & Chithra nom. | Myrtaceae |
| Litsea glabrata (Wall. ex Nees) Hook. f. | Lauraceae |
| Eurya nitida Korth. | Theaceae |
| Neolitsea scrobiculata (Meisner) Gamble. | Lauraceae |
| Cassine paniculata (Wight & Arn.) Lobr.-Callen | Celastraceae |
| Mahonia leschenaultii (Wall. ex Wight & Arn) | Berberidaceae |
| Cinnamomum perrottetii Meisner | Lauraceae |
| Litsea wightiana (Nees) Hook. f. | Lauraceae |
| Trichilia connaroides (Wight & Arn.) Bent var. connaroides | Meliaceae |
| Neolitsea fischeri Gamble. | Lauraceae |
| Rubus ellipticus Smith. | Rosaceae |
| Cinnamomum malabatrum (Burm. F.) Blume | Lauraceae |
| Cinnamomum wightii Meisner | Lauraceae |
| Coffea arabica L. | Rubiaceae |
| Cryptocarya bourdillonii Gamble | Lauraceae |
| Cryptocarya neilgherrensis Meisner | Lauraceae |
| Litsea floribunda (Blume) Gamble | Lauraceae |
| Photinia integrifolia Lindl. var. sub-lanceolata Miq. | Rosaceae |
| Syzygium lanceolatum (Lam.) Wight & Arn. | Myrtaceae |
| Elaeocarpus glandulosus Wall. | Elaeocarpaceae |
| Ficus drupacea Thunb. | Moraceae |
| Rubus racemosus Roxb. | Rosaceae |
| Ilex wightiana | Aquifoliaceae |
DownLoad:
CSV
 Email Alerts
Email Alerts RSS Feeds
RSS Feeds