Environment Analysis and Management Unit - Guido Tosi Research Group - Department of Theoretical and Applied Sciences, Università degli Studi dell'Insubria, Via J. H. Dunant 3, 21100, Varese, Italy
b.
CNR-IRSA National Research Council – Water Research Institute, Via del Mulino 19, 20861, Brugherio, MB, Italy
c.
Gruppo Insubrico di Ornitologia, c/o Civico Museo Insubrico di Storia Naturale di Clivio e Induno Olona, Via Manzoni, 21050, Clivio, VA, Italy
d.
Parco Pineta di Appiano Gentile e Tradate, Via Alessandro Manzoni, 11, 22070, Castelnuovo Bozzente, CO, Italy
e.
Dipartimento di Scienze della Vita e Biologia dei Sistemi, Università degli Studi di Torino, Via Accademia Albertina 13, 10123, Turin, Italy
f.
Conservation Department LIPU-Birdlife Italia, Via Udine 3/a, 43122, Parma, Italy
g.
Department of Environmental Science and Policy, Università degli Studi di Milano, Via Celoria 26, 20122, Milan, Italy
* Corresponding author. Environment Analysis and Management Unit - Guido Tosi Research Group - Department of Theoretical and Applied Sciences, Università degli Studi dell'Insubria, Via J. H. Dunant 3, 21100, Varese, Italy.
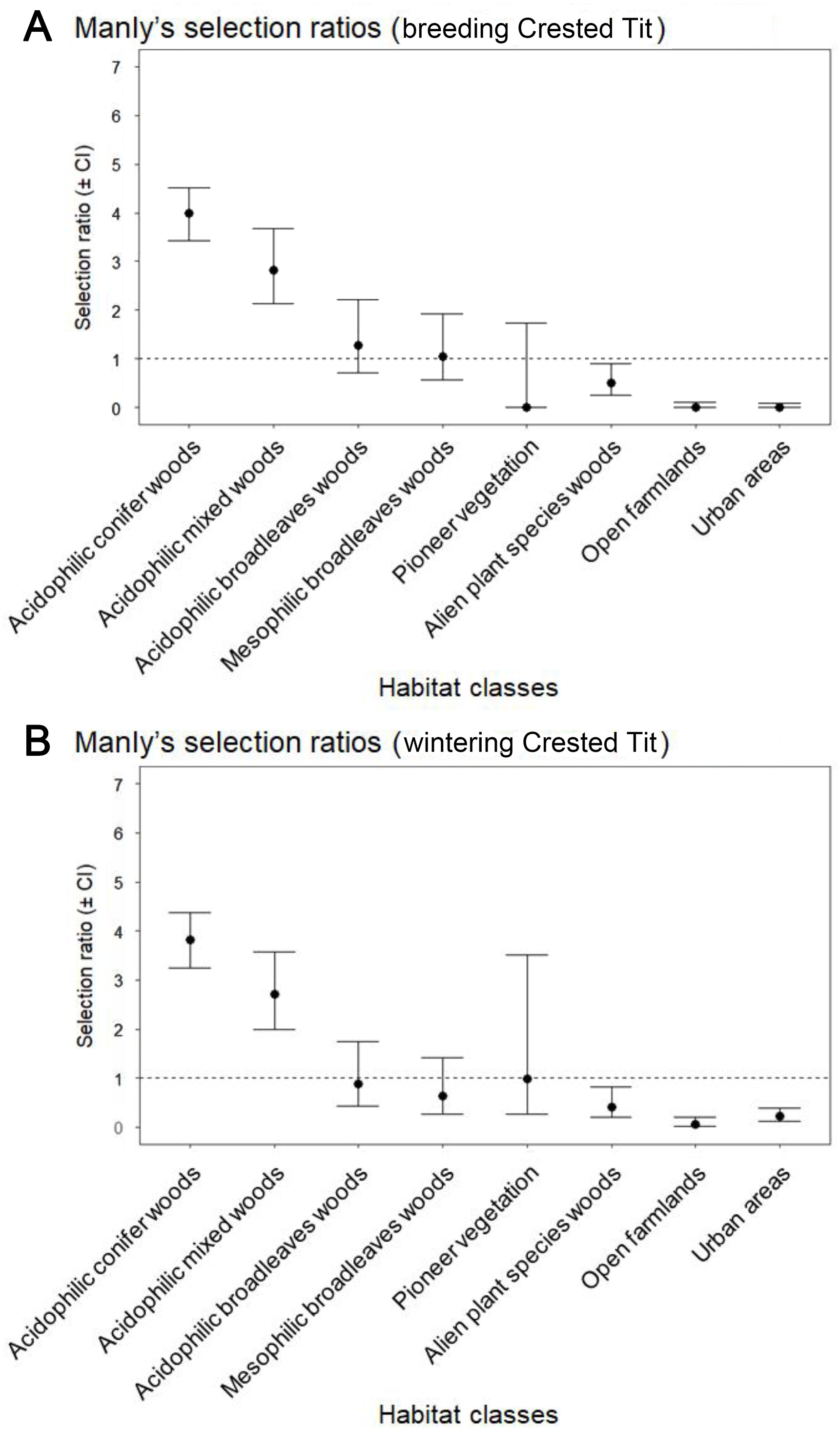
Determining how animals respond to resource availability across spatial and temporal extents is crucial to understand ecological processes underpinning habitat selection. Here, we used a multi-scale approach to study the year-round habitat selection of the Crested Tit (Lophophanes cristatus) in a semi-natural lowland woodland of northern Italy, analysing different habitat features at each scale. We performed Crested Tit censuses at three different spatial scales. At the macrohabitat scale, we used geolocalized observations of individuals to compute Manly's habitat selection index, based on a detailed land-use map of the study area. At the microhabitat scale, the trees features were compared between presence and absence locations. At the foraging habitat scale, individual foraging birds and their specific position on trees were recorded using focal animal sampling. Censuses were performed during both the breeding (March to May) and wintering (December to January) seasons. At the macrohabitat scale, the Crested Tits significantly selected pure and mixed pine forests and avoided woods of alien plant species, farmlands and urban areas. At the microhabitat scale, old pine woods with dense cover were selected, with no significant difference in the features of tree selection between the two phenological phases. At the foraging habitat scale, the species was observed spending more time foraging in the canopies than in the understorey, using mostly the portion of Scots Pine (Pinus sylvestris) canopies closer to the trunk in winter, while during the breeding period, the whole canopy was visited. Overall, breeding and wintering habitats largely overlapped in the Crested Tit. Based on our findings, lowland Crested Tits can be well defined as true habitat specialists: they are strictly related to some specific coniferous woodland features. Noteworthily, compared to other tit species, which normally show generalist habits during winter, the Crested Tit behaves as a habitat specialist also out of the breeding season. Our study stressed the importance of considering multi-scale (both spatial and phenological) habitat selection in birds.
Tits and titmice are small, familiar cavity-nesting songbirds forming the family Paridae which contains 4 genera, 55 to 65 species, depending on different classifications (Harrap and Quinn, 1996; Salzburger et al., 2002; Dickinson, 2003). They are distributed in the entire Holarctic, Oriental and Afrotropical regions with a hot spot of diversity in the Himalayan and Chinese mountain ranges (Päckert et al., 2007). Because of their overall morphological similarity, all but three species have been classified into the genus Parus and the genus is divided into 10 subgenera (Harrap and Quinn, 1996) or 12 species groups (Eck, 1988). The three other genera, Melanochlora, Sylviparus and Pseudopodoces are monotypic. A well-supported, corroborated and presumably accurate phylogeny is essential for an understanding of ecological and behavioral variation within the group. Therefore, in recent years, a plethora of studies using molecular markers, such as allozyme comparisons (Gill et al., 1989), mitochondrial restriction fragment-length polymorphism (RFLP) data (Gill and Slikas, 1992; Gill et al., 1993), DNA-DNA hybridization distances (Sheldon et al., 1992; Slikas et al., 1996) and cytochrome-b sequences (Kvist et al., 1996; Gill et al., 2005; Martens et al., 2006) have been applied to the phylogeny of Paridae. However, the phylogenetic relationship among species tends to become increasingly controversial as different genetic markers emerge. For instance, based on nuclear DNA-DNA hybridization, Parus caeruleus and P. major formed a clade which is the sister group of all other Parids (Sheldon et al., 1992; Slikas et al., 1996), but the phylogeny inferred from cyt b sequences did not support that result (Gill et al., 2005).
Aegithalos, at one time, was classified as a genus in the family Paridae (Gadow, 1883; Hellmayr, 1903; Mayr and Amandon, 1951). However, Snow (1967) considered Aegithalos and a few related genera as a separate family, the Aegithalidae. Phylogenetic analyses based on DNA-DNA hybridization (Sibley and Ahlquist, 1990; Sheldon and Gill, 1996), protein allozyme comparisons (Ohta et al., 2000) and nuclear gene sequences (Barker et al., 2002; Spicer and Dunipace, 2004) suggested that Aegithalos was indeed not Parids and the closest sister group to Parids was the penduline tits, species of Remizidae. However, genera closely related to Aegithalos are still under dispute, as well as the phylogenetic relationships within Aegithalos (Päckert et al., 2010).
In this study, we sampled part of representative taxa of tits, long-tailed tits and penduline tits in an attempt to resolve the relationships among these three groups and also to infer the phylogenetic relationships within tits and long-tailed tits based on two mitochondrial gene sequences, COΙ and cyt b.
Materials and methods
Taxon selection and choice of outgroups
Altogether, twenty-nine individuals of ten species from Paridae, nine individuals of four species from Aegithalos and two Chinese penduline tits were included in this study. Six species had more than one subspecies included in the analysis (Table 1). According to Sibley and Monroe (1990), Sheldon and Gill (1996), Barker et al. (2002), Spicer and Dunipace (2004), an individual of Pica pica, Uragus sibiricus, Garrulax lunulatus and G. ocellatus were selected as outgroups.
Extraction, amplification and sequencing
Genomic DNA was extracted from blood, feathers or tissue specimens using the QIAamp™ DNA Mini Kit as per manufacturer's instructions. Nucleotide sequence data were obtained from the two mitochondrial, cytochrome c oxidase Ⅰ (COI) and cytochrome b (cyt b) genes. PCR amplification and sequencing of cytochrome c oxidase Ⅰ followed the method suggested by Sorenson et al. (1999), while Gill et al. (2005) described protocols for cytochrome b.
For the sequencing reactions, the same primers were used. Both strands of each PCR product were sequenced. For each gene and sample, multiple sequence fragments were obtained by sequencing with different primers. Complete sequences were assembled using Seqman Ⅱ (DNASTAR®). Sequences were compared visually to the original chromatograms to avoid reading errors. Assembled sequences were aligned by eye. All sequences were deposited in GenBank.
Phylogenetic analysis
Based on a priori assumption and partition homogeneity test (p = 0.44), the two mitochondrial genes were analyzed as one data set with a total nucleotide length of 2149 base pairs (bp). The data were analyzed using maximum likelihood (ML, Felsenstein, 1981) and Bayesian inference methods (BI, Rannala and Yang, 1996; Yang and Rannala, 1997; Larget and Simon, 1999). Statistics for nucleotide variation and genetic distance were computed with MEGA 4 (Tamura et al., 2007). A Jukes-Cantor estimate of the number of nucleotide substitutions per site was computed for the cyt b gene (Jukes and Cantor, 1969). Following the suggestion of Nei and Kumar (2000), if the Jukes-Cantor distance is less than 0.05, the p-distance would be used. Otherwise, a more complicated distance model would be employed.
Following alignment, we partitioned the data by genes in order to allow different rates for the various partitions for ML and BI analyses. Nucleotide substitution models were selected separately by genes and then used for different data partitions in reconstruction. However, the TVM + I + G model (−lnL = 15397.6758, K = 9, AIC = 30813.3156) was identified as the best fit for these two genes, using both the likelihood-ratio test (LRT) and Akaike Information Criterion (AIC) implemented in MODELTEST 3.7 (Posada and Crandall, 1998). The parameter values for the model include a symmetric rate matrix specifying relative probabilities for all possible nucleotide changes (Rmatrix = 0.6396 [A-C], 8.0280 [A-G], 1.1611 [A-T], 0.0498 [C-G], 8.0280 [C-T], 1.0000 [G-T]), the proportion of invariant sites (pinvar = 0.6183) and the shape parameter for the gamma distribution of rate variation (shape = 1.2128). The base frequencies were set as follows: A = 0.3263, C = 0.4100, G = 0.1009, T = 0.1628.
ML reconstruction (1000 replicates) was performed in TREEFINDER (Jobb, 2007) and BI performed in MRBAYES 3.1.2 (Ronquist and Huelsenbeck, 2003). In the BI analysis, we ran two analyses of two million generations and trees sampled every 100 generations. One cold and three heated Markov chains were used in our analysis. The trees saved during the "burn-in" phase (the first 200000 generations in our analysis) were discarded. The remaining trees from two runs were used to create a 50% majority rule consensus tree.
Results
Sequence characteristics
We obtained a total of 88 sequences, with their GenBank accession numbers listed in Table 1. No stop codons were identified in a contiguous 1201 bp stretch of the COI gene and 948 bp of cyt b. The overlapping sequences from different PCR products and a single peak in the electropherograms suggest that these sequences do not come from "numts" (nuclear sequences of mitochondrial origin). The average base composition of sequence was skewed, which is similar to that found in previous avian studies (Barhoum and Burns, 2002; Webb and Moore, 2005). The characteristics of these sequences are summarized in Table 2.
Table
1.
List of taxonomic samples and sequences used in the study
Family
Genus
Species and subspecies
Museum No.
Collection locality
GenBank Accession Nos.
cyt b
COΙ
Paridae
Parus
Parus major commixtus
IOZ1254
Guangxi
HM185345
HM185334
Parus major subtibetanus
IOZ5236
Yunnan
HM185346
HM185333
Parus major subtibetanus
IOZ3918
Sichuan
HM185347
HM185332
Parus major artatus
IOZ5661
Jilin
HM185348
HM185331
Parus monticolus yunnanensis
IOZ3633
Sichuan
HM185349
HM185330
Parus monticolus yunnanensis
IOZ2465
Shaanxi
HM185350
HM185329
Parus spilonotus rex
IOZ3026
Fujian
HM185351
HM185328
Parus cyanus
IOZ 781
Xinjiang
HM185352
HM185327
Parus venustulus
IOZ2806
Hubei
HM185353
HM185326
Parus venustulus
IOZ1945
Shaanxi
HM185354
HM185325
Parus ater aemodius
IOZ1094
Gansu
HM185355
HM185324
Parus ater
IOZ2331
Zvolen
HM185356
HM185323
Parus ater ater
IOZ9029
Heilongjiang
HM185357
HM185322
Parus palustris hypermelas
IOZ1239
Gansu
HM185358
HM185321
Parus palustris hypermelas
IOZ7991
Hubei
HM185359
HM185320
Parus palustris brevirostris
IOZ5675
Jilin
HM185360
HM185319
Parus palustris hellmayri
IOZ8680
Shanxi
HM185361
HM185318
Parus palustris hellmayri
IOZ2458
Shaanxi
HM185362
HM185343
Parus montanus baicalensis
IOZ5944
Heilongjiang
HM185363
HM185317
Parus montanus baicalensis
IOZ5945
Heilongjiang
HM185364
HM185316
Parus montanus
IOZ2237
Zvolen
HM185365
HM185315
Parus montanus affinis
IOZ2178
Shaanxi
HM185366
HM185342
Parus dichrous dichrous
IOZ2489
Shaanxi
HM185367
HM185314
Parus dichrous dichrous
IOZ6117
Shaanxi
HM185368
HM185313
Parus cristatus
IOZ2333
Zvolen
HM185369
HM185312
Sylviparus
Sylviparus modestus modestus
KIZglgs1240
Yunnan
HM185370
HM185311
Sylviparus modestus modestus
KIZglgs1241
Yunnan
HM185371
HM185310
Pseudopodoces
Pseudopodoces humilis
IOZ4783
Qinghai
HM185372
HM185309
Pseudopodoces humilis
IOZ4785
Qinghai
HM185373
HM185308
Remizidae
Remiz
Remiz consobrinus
IOZ10640
Liaoning
HM185374
HM185298
Remiz consobrinus
IOZ10652
Liaoning
HM185375
HM185297
Aegithalidae
Aegithalos
Aegithalos caudatus caudatus
IOZ5677
Jilin
HM185376
HM185307
Aegithalos caudatus glaucogularis
IOZ2769
Hubei
HM185377
HM185306
Aegithalos caudatus glaucogularis
IOZ3322
Shaanxi
HM185378
HM185305
Aegithalos concinuus concinnus
IOZ1152
Gansu
HM185379
HM185304
Aegithalos concinuus concinnus
IOZ1499
Hunan
HM185380
HM185303
Aegithalos concinuus concinnus
IOZ3661
Sichuan
HM185381
HM185302
Aegithalos concinuus talifuensis
IOZ5344
Yunnan
HM185382
HM185301
Aegithalos bonvaloti
IOZ3766
Sichuan
HM185383
HM185300
Aegithalos fuliginosus
IOZ2476
Shaanxi
HM185384
HM185337
Timaliidae
Garrulax
Garrulax lunulatus
IOZ2594
Shaanxi
HM185385
HM185341
Garrulax ocellatus
IOZ5083
Yunnan
HM185386
HM185340
Fringillidae
Uragus
Uragus sibiricus
IOZ5870
Heilongjiang
HM185387
HM185344
Corvidae
Pica
Pica pica
IOZ3931
Xinjiang
HM185388
HM185339
Note: IOZ, Institute of Zoology, Chinese Academy of Sciences; KIZ, Kunming Institute of Zoology, Chinese Academy of Sciences.
Because the mean Jukes-Cantor distance was 0.112 ± 0.006 SE (> 0.05) for the cyt b sequence data set, the Tamura-Nei model was chosen to compute the genetic distance (Tamura and Nei, 1993). Sequence divergences within species range from 0% (Parus dichrous) to 2.8% (Aegithalos concinnus), with an average divergence of 1.15%. The mean cyt b sequence distance of Aegithalos was 0.081 ± 0.007 SE, with the lowest 0.002 between A. bonvaloti and A. fuliginosus and the highest 0.118 between A. concinnus and A. caudatus. The mean distance of genus Parus was 0.077 ± 0.005 SE, with the lowest 0.053 between P. major and P. monticolus. Between taxa in Parus and the other two species in the Paridae, average pairwise divergences are as follows: 0.115 ± 0.009 SE to Sylviparus modestus and 0.097 ± 0.007 SE to Pseudopodoces humilis. The distances were 0.16 ± 0.01 SE between Aegithalos and Paridae, 0.18 ± 0.013 SE between Aegithalos and Remizidae, and 0.126 ± 0.01 SE between Paridae and Remizidae.
Phylogenetic relationships
The consensus tree from the Bayesian analysis is identical to the ML tree, except for a few weakly supported nodes. A few nodes that are resolved in the ML tree are polytomies in the Bayesian consensus tree (Fig. 1). In all optimal trees, no species which includes several subspecies were found to be paraphyletic in our study. Paridae, Remizidae, Aegithalos and Garrulax species formed a monophyletic group with high support (bootstrap support = 70%, Bayesian posterior probability = 97%) and branched off to two monophyletic clades. The clade, representing individuals from species of Paridae and Remidae, was highly supported (bootstrap = 98%, BPP = 100%). Remiz consobrinus positions as a sister species to the Paridae, including Sylviparus, Pseudopodoces and the species in Parus.Aegithalos is monophyletic, grouped with two Garrulax species and, with weak support, formed the other clade.
Figure
1.
Trees obtained from the analysis of the combined COΙ and cyt b data: (a) Bayesian and (b) maximum likelihood. The optimal model as estimated by Modeltest following the AIC and LRT was estimated to be TVM + I + G. Support values are indicated to the left of the nodes.
Within Aegithalos, four species are monophyletic and grouped in all optimal trees (Bayesian posterior probability = 100%, bootstrap = 100%). Within the clade, Aegithalos bonvaloti is sister to A. fuliginosus and that pair is sister to A. caudatus. Aegithalos concinnus is sister to these three species. Phylogenetic relationships have strong support (Bayesian posterior probability = 100%, bootstrap > 95%) and are identical in the two phylogenetic trees.
Within the family Paridae, Sylviparus modestus positions as a sister species to the remaining species of Paridae from the two trees. These remaining species form two high-support monophyletic clades in the BI tree (BPP = 99%), but the bootstrap value is rather low (bootstrap = 44%) in the ML tree. One clade includes Parus cyanus, P. monticolus, P. major, P. spilonotus and Ps. humilis. The relationships among these five species are congruent in the two trees except for the position of Ps. humilis. In the two trees, Parus cyanus is a sister species to the remaining species, although the nodal support value is low. Parus major and P. monticolus pair as sister taxa with high support. In the BI tree, P. spilonotus and Ps. humilis pair as sister taxa but in the ML tree, Pseudopodoces humilis is a sister species to P. spilonotus, P. major and P. monticolus. However, the nodal supports are low. Another clade includes P. ater, P. venustulus, P. palustris, P. montanus and two Eurasian crested tits, P. dichous and P. cristatus. Parus palustris and P. montanus, P. ater and P. venustulus, P. dichous and P. cristatus pair as sister taxa with high support. Relationships among these three pairs are unclear because of polytomies in the BI tree and low nodal support value in the ML tree.
Discussion
Phylogenetic relationships among Paridae, Remizidae and Aegithalos
Our data support the classification for the family Paridae proposed by Sibley and Ahlquist (1990). Based on DNA-DNA hybridization, they combined family Remizidae with Paridae, hence the family Paridae includes two subfamilies, Remizinae and Parinae. Sheldon and Gill (1996) studied the phylogeny of song birds using DNA-DNA hybridization and concluded that the sister group of Paridae is the Remizidae. Based on combined myoglobin intron Ⅱ and cytochrome b sequences, Alström et al. (2006) revealed the close relationships between P. major and R. pendulnus. In our study, the Chinese penduline tit (Remizidae) positioned as sister group to the Parids (Paridae) with high support. This may indicate very close relationships between Paridae and Remizidae. Although, in the present study, only one species of Remizidae was tested, the high nodal support (> 95%) signifies that the designated branch is most possibly unaffected by sampling among variation existing in the data (Slikas et al., 1996).
Long-tailed tits, species of Aegithalos, because of their morphological similarity with tits, were earlier believed to be Parids by several authors (Gadow, 1883; Hellmayr, 1903; Mayr and Amandon, 1951). However, Stresemann (1923) described several characteristics of Aegithalos that differ from those of the Parids, such as the presence of a complete juvenile-molt, nest structure and naked hatching. Since then, Aegithalos and two other monotypic or small genera are usually placed in the family Aegithalidae (e.g., Paynter, 1967; Morony et al., 1975; Sibley and Ahlquist, 1990; Sibley and Monroe, 1990). Sturmbauer et al. (1998) suggested a close relationship between Leptopoecile and Aegithalos based on the mitochondrial 16S sequence. Alström et al. (2006) confirmed the close relationship between Aegithalos and Leptopoecile based on the combined myoglobin intron Ⅱ and cytochrome b sequences. Using protein allozyme comparison, Ohta et al. (2000) found the genetic distance between Aegithalos and Parus appears to be at a familial level. In our study, two distinct clades and Aegithalos grouped with two outgroup Garrulax species, in spite of weak nodal support, show distant relationships between Aegithalos and Paridae. In addition, the genetic divergence of cyt b between Aegithalos and Paridae is larger than that between Remizidae and Paridae. Hence, the morphological similarity between tits and long-tailed tits may have resulted from evolutionary convergence.
Phylogenetic relationships within Paridae
Based on plumage pattern, Hellmayr (1903) had recognized eleven subgenera for the genus Parus. However, the delimitation of these subgenera is in dispute. For instance, Wolters (1982) classified the species of Pardaliparus into Pariparus and this classification was supported by molecular analyses (Slikas et al., 1996; Gill et al., 2005). In our study, Parus ater and P. venustulus also formed a closely related group, but the divergence of cyt b is ~7% (uncorrected p-distance). According to Hellmayr's classification, our samples belong to seven subgenera: Cyanistes (P. cyanus), Baeolophus (P. dichous, P. cristatus), Poecile (P. montanus, P. palustrs), Periparus (P. ater), Pardaliparus (P. venustulus), Machlolophus (P. spilonotus) and Parus (P. major, P. monticolus). Similar to previous studies, our data did not reveal phylogenetic relationships among these subgenera. Therefore, the Parus phylogeny needs to be studied further. We intend to add more species and subspecies and new markers, including nuclear loci to classify this challenge in the future.
Although our results came to quite similar conclusions as previous studies, different phylogenetic relationships among species were detected as well. The question of greatest interest is: what is the phylogentically most closely related species to Ps. humilis in Parids? This aberrant and enigmatic Tibetan species, earlier thought to be a corvid, turned out to be a Parid (James et al., 2003). Past phylogenetic analyses based on cyt b gene sequences positioned Ps. humilis as sister species to P. major (James et al., 2003; Gill et al., 2005). However, in optimal trees based on the cyt b and COΙ gene sequences in our study, Ps. humilis is sister to P. spilonotus, although the nodal support is weak. This result may indicate that more data of the species and different markers are needed to determine the phylogenetic status of Parids.
Gill et al. (2005) recommended upgrading six subgenra of Parus into genera for facilitating future evolutionary analyses among Parids. The retained genus Parus only included P. major, P. monticolus, P. xanthogenys, P. spilonotus, P. holsti and the African tits (P. leucomelas, P. niger, P. carpi, P. albiventris, P. leuconotus, P. rufiventris, P. funereus, P. fringillinus, P. fasciiventer, P. thruppi, P. griseiventris, P. cinerascens and P. afer). However, in our present study, we also believe that the genus Machlolophus should be recognized in consideration of the phylogenetic relationships between P. spilonotus and Ps. humilis and distinct plumage pattern between P. major and P. spilonotus (Ohta et al., 2000). Furthermore, in our data, the divergence of the cyt b gene between P. major–P. monticolus group and P. spilonotus (8.4%, uncorrected p-distance) is no less than the divergence among recommended genera (ranging from 7.3% between Pariparus and Baeolophus to 9.7% between Poecile and Machlolophus, uncorrected p-distance).
Intra- and interspecies relationships and history of Aegithalos taxa
On the one hand, intraspecific divergence is significant. The Black-throated Tit (Aegithalos concinnus) includes six highly distinct subspecies. These subspecies are remarkably different in their plumage pattern. Divergence differences of the cyt b gene among these subspecies are also significant. In our data, the pairwise p-distance between specimens from Bengal (India, ssp. iredalei, sequence obtained from GenBank) and Gaoligong (Yunnan, ssp. talifuensis) was 4.5%. p-distance between specimens from Gaoligong and Pingjiang (Hunan, ssp. concinnus) was 6.1%. p-distance between specimens from Bengal and Pingjiang was 5.1%. Eck and Martens (2006) sequenced three specimens, including subspecies ssp. iredalei, ssp. talifuensis and ssp. manipurensis, and they found the pairwise p-distance was 5.1% between ssp. iredalei and ssp. talifuensis, 5.3% between ssp. talifuensis and ssp. manipurensis and 6% between ssp. iredalei and ssp. manipurensis. These four subspecies are reddish-crowned. The other two subspecies A. c. pulchellus and A. c. annamensis are grey-headed. They are obviously different in presence of reddish breast band, the latter of which is absent. Hence, it seems possible that A. concinnus represents an unresolved species swarm (Eck and Martens, 2006).
As for the Long-tailed Tit (Aegithalos caudatus), based on comparative morphology, up to 19 subspecies are currently recognized, divided by Harrap and Quinn (1996) into four groups. Zink et al. (2005) studied the mitochondrial phylogeography, including five subspecies or representatives of A. caudatus group and A. alpinus group of the long-tailed tit. Within their samples, no correspondence between five subspecies and mitochondrial subdivision has been discovered. However, two geographically unsorted lineages were displayed and differentiation seems to exist between the two main groups. In this study, the cyt b divergence of two specimens belonging to the subspecies A. c. caudatus and A. c. glaucogularis, is 1.3%. Thus, phylogeographic structures would be developed if additional subspecies were taken into consideration.
On the other hand, interspecies molecular divergence is small. The morphologically distinct sister species A. fuliginosus and A. bonvaloti are separated by unexpectedly small cyt b divergences (0.2%, uncorrected p-distance) and associated very short branch lengths in the cyt b tree. The pairwise cyt b divergence between the species is comparable to that within populations of the same species in other passerine birds. This could indicate recent separation and divergence from their common ancestor. However, surprisingly, the more slowly evolving nuclear locus β-fibrinogen intron 7 shows relatively greater divergence (2.5%, uncorrected p-distance) than the faster-evolving cyt b (unpublished data). This might be the result of amplification of nuclear pseudogenes instead of mitochondrial DNA (Zhang and Hewitt, 1996; Sorensen and Quinn, 1998). However, our sequences show no evidence of being of nuclear origin. Introgression of mitochondrial DNA seems to be a more likely explanation. A. fuliginosus and A. bonvaloti are not known to hybridize, but their current distributions partly overlap. Hybridization or past hybridization leading to introgression is nevertheless a possibility. Weckstein et al. (2001) argued that introgressive hybridization is the cause of discordant patterns of mitochondrial and allozyme data in the North American sparrows Zonotrichia leucophrys and Z. Atricapilla.
Furthermore, species limits within the A. niveogularis are not, as yet, reliably defined. Harrap and Quinn (1996) proposed the species to be split in two species while Dickinson (2003) proposed a three way split. Therefore, currently, five (Martens and Eck, 1995) or six (Harrap and Quinn, 1996) or seven (Dickinson, 2003) species have been recognized within the genus Aegithalos. Our study includes only a few species and subspecies from Aegithalos because of insufficient or unavailable samples. Recently, Päckert et al. (2010) studied the phylogeny of long-tailed tits and allies based on mitochondrial and nuclear markers and resolved the status of some species on the basis of phylogenetic relationships. However, by considering the long evolutionary time of species and differences within species, both morphological and molecular characters, a study which includes more species and subspecies is necessary to define the phylogeny of Aegithalos.
We suggest that the southwestern mountain ranges of China might be the center of origin of Aegithalos species and that the taxa of this genus colonized new habitats from west to east across China. Supporting this hypothesis are the patterns of taxonomic diversity and endemism. For example, members of all long-tailed tits are found in the areas of southwestern China except A. niveogularis. The oldest linage in the genus in our study is the A. concinnus talifuensis which is distributed in Yunnan Province of China. The historical biogeography can be deduced by the relationships among species and the areas of species distribution (Gill et al., 2005). For instance, the closely related phylogenetic relationships between Aegithalos fuliginosus and A. bonvaloti is accompanied by a partly overlapped distribution. Aegithalos caudatuscaudatus is older than A. c. glaucogularis in our phylogenetic analysis and is thus distributed farther from the original areas than A. c. glaucogularis. Given a rough calibration of 2% divergence per million years, we hypothesize that the separation of A. concinnus may occur ~5.5 Mya, on the basis of the estimate of 11% sequence divergence between A. concinnus and other three species. Then, about 4.5 Mya ago, A. caudatus was divided and colonized the palearctic region. More recently, separation between Aegithalos fuliginosus and A. bonvaloti is hypothesized to have taken place in the late Pleistocene, i.e., ~100000 years ago. However, that break may be earlier than late Pleistocene, for the possible introgression between two species may obscure the real separation time.
Congruent genetic differentiation among co-distributed species
Comparative phylogeography helped elucidate the relative effect of shared historical earth events on current patterns of biodiversity by comparing historical patterns of gene flow and divergence among species that overlap in time and space (Hickerson et al., 2010). In other words, co-distribution species may have similar population genetic differentiation or congruent phylogeographical patterns as a result of sharing common environmental and geological changes. In our study, the Great Tit (Parus major) and the Black-throated Tit (A. concinnus) overlap in southern and southwestern China, the Coat Tit (P. ater), the Willow Tit (P. montanus), the Marsh Tit (P. palustris) and the Long-tailed Tit are distributed in the northeast, center and west of China. Although our sample size was small, we found congruent genetic differentiation among these sympatric distribution taxa. The great tit and the black-throated tit have similar phylogeographical patterns. They differentiated into two clades, one including the samples of Yunnan Province and the other one belonging to the other sample locations in China. Similarly, the coat tit, the marsh tit, the willow tit and the long-tailed tit also divided into two branches, one covering the areas of northeastern China and Europe and the other including the species distribution in central and western China. These phylogeographical patterns or population structures of sympatric species may indicate that the historical earth events they experienced, including climatic and geological, had almost the same effect on these closely related taxa.
Acknowledgements
We are grateful to Tao Li for laboratory assistance and anonymous reviewers who provided helpful comments to the improvement of the manuscript. This work was financially supported by the National Natural Science Foundation of China (Grant No. 30870270) and the National Science Funds for Distinguished Young Scientists (No. 30925008) to Fumin Lei.
Aho, K., Bowyer, R.T., 2015. Confidence intervals for ratios of proportions: implications for selection ratios. Method. Ecol. Evol. 6, 121-132. .
Aho, K.A., 2014. Foundational and Applied Statistics for Biologists using R. CRC Press/Taylor & Francis Group, Boca Raton.
Altmann, J., 1974. Observational study of behavior: sampling methods. Behaviour 49, 227-266. .
Anderson, S.H., Shugart, H.H., 1974. Habitat selection of breeding birds in an East Tennessee deciduous forest. Ecology 55, 828-837. .
Assandri, G., Cecere, J.G., Sarà, M., Catoni, C., de Pascalis, F., Morinay, J., et al., 2022. Context-dependent foraging habitat selection in a farmland raptor along an agricultural intensification gradient. Agr. Ecosyst. Environ. 326, 107782. .
Atiénzar, F., Barba, E., Holleman, L.J.M., Belda, E.J., 2009. Nesting habitat requirements and nestling diet in the Mediterranean populations of Crested Tits Lophophanes cristatus. Acta Ornithol. 44, 101-108. .
Berlusconi, A., Preatoni, D., Assandri, G., Bisi, F., Brambilla, M., Cecere, J.G., et al., 2022. Intra-guild spatial niche overlap among three small falcon species in an area of recent sympatry. Eur. Zool. J. 89, 510-526. .
Bianchi, A., 2002. Piano di settore agro-forestale, piano di indirizzo forestale per la tutela e la gestione delle aree boscate e la difesa dagli incendi boschivi - Parco Pineta di Appiano Gentile e Tradate. Consorzio Forestale del Ticino.
Bibby, C.J., 2000. Bird Census Techniques, 2nd ed. Academic, London; San Diego.
Block, W.M., Brennan, L.A., 1993. The habitat concept in ornithology: theory and applications. In: Power, D.M. (Ed.), Current Ornithology. Springer, Boston, pp. 35-91. https://doi.org/10.1007/978-1-4757-9912-5_2.
Blondel, J., Maistre, M., Perret, P., 1993. Habitat heterogeneity and life-history variation of Mediterranean Blue Tits (Parus caeruleus). Auk 110, 511-520. .
Bowyer, R.T., Kie, J.G., 2006. Effects of scale on interpreting life-history characteristics of ungulates and carnivores. Divers. Distrib. 12, 244-257. .
Brambilla, M., Rubolini, D., Guidali, F., 2006. Factors affecting breeding habitat selection in a cliff-nesting peregrine Falco peregrinus population. J. Ornithol. 147, 428-435. .
Brichetti, P., Fracasso, G., 2011. Ornitologia italiana: identificazione, distribuzione, consistenza e movimenti degli uccelli italiani - vol. 7 Paridae - Corvidae, Vol. 7. Alberto Perdisa editore, Bologna.
Broughton, R.K., Hill, R.A., Freeman, S.N., Bellamy, P.E., Hinsley, S.A., 2012. Describing habitat occupation by woodland birds with territory mapping and remotely sensed data: an example using the Marsh Tit (Poecile palustris). Condor 114, 812-822. .
Brown, J.H., 1984. On the relationship between abundance and distribution of species. Am. Nat. 124, 255-279. .
Caughley, G., Gunn, A., 1996. Conservation biology in theory and practice. Blackwell Science, Cambridge.
Chalfoun, A.D., Martin, T.E., 2007. Assessments of habitat preferences and quality depend on spatial scale and metrics of fitness. J. Appl. Ecol. 44, 983-992. .
Collette, J., 1987. Denombrément des oiseaux nicheurs d'un taillis sous futaie dans le Sud de la Manche. Le Cormoran 3, 77-92.
Cramp, S., Perrins, C.M., 1993. Handbook of the birds of Europe the Middle East and North Africa. 7: Flycatchers to Shrikes. Oxford University Press, Oxford.
Cutini, S., Bagni, L., Campedelli, T., Londi, G., Florenzano, G.T., 2009. Ecologia e possibili linee d'espansione della cincia dal ciuffo Lophophanes cristatus nell'Appennino. Alula 6.
Dayton, P.D., Tegner, M.J., 1984. The importance of scale in community ecology: a kelp forest example with terrestrial analogs. In: Price, P.W., Slobodchikoff, C.N., Gaud, W.S. (Eds.), A New Ecology: Novel Approaches to Interactive Systems. John Wiley and Sons, New York, pp. 457-476.
Denny, R.E., Summers, R.W., 1996. Nest site selection, management and breeding success of Crested Tits Parus cristatus at Abernethy Forest, Strathspey. Bird Study 43, 371-379. .
Díaz, M., Illera, J.C., Atienza, J.C., 2008. Food resource matching by foraging tits Parus spp. during spring-summer in a Mediterranean mixed forest; evidence for an ideal free distribution. Ibis 140, 654-660. .
Djorgova, N., Ragyov, D., Biserkov, V., Biserkov, J., Nikolov, B.P., 2021. Habitat preferences of diurnal raptors in relation to human access to their breeding territories in the Balkan Mountain Range, Bulgaria. Avian Res. 12, 29. .
Domínguez, J., Carbonell, R., Ramirez, A., 2017. Seasonal changes in habitat selection by a strict forest specialist, the Middle Spotted Woodpecker (Leiopicus medius), at its southwestern boundary: implications for conservation. J. Ornithol. 158, 459-467. .
Ekman, J., 1979. Coherence, composition and territories of winter social groups of the Willow Tit Parus montanus and the Crested Tit P. cristatus. Ornis Scand. 10, 56. .
Estrada, J., Pedrocchi, V., Brotons, L., Herrado, S., 2004. Atles dels ocells nidificants de Catalunya 1999-2002, Institut Català d'Ornitologia, Lynx Edicions.
Gagliardi, A., Guenzani, W., Preatoni, D.G., Saporetti, F., Tosi, G., 2007. Atlante Ornitologico Geo-referenziato della Provincia di Varese. Uccelli Nidificanti 2003-2005. Civico Museo Insubrico di Storia Naturale di Induno Olona; Università degli Studi dell'Insubria, sede di Varese.
Hartley, P.H.T., 1987. Ecological aspects of the foraging behaviour of Crested Tits Parus cristatus. Bird Study 34, 107-111. .
Hernández Á., 2021. Habitat use and space preferences of Eurasian Bullfinches (Pyrrhula pyrrhula) in northwestern Iberia throughout the year. Avian Res. 12, 8. .
Hinsley, S.A., Carpenter, J.E., Broughton, R.K., Bellamy, P.E., Rothery, P., Amar, A., et al., 2007. Habitat selection by Marsh Tits Poecile palustris in the UK: habitat selection by Marsh Tits. Ibis 149, 224-233. .
Holbrook, J.D., Olson, L.E., DeCesare, N.J., Hebblewhite, M., Squires, J.R., Steenweg, R., 2019. Functional responses in habitat selection: clarifying hypotheses and interpretations. Ecol. Appl. 29, e01852. .
Horák, J., 2017. Insect ecology and veteran trees. J. Insect Conserv. 21, 1-5. .
Jansson, C., Bromssen, A., 1981. Winter decline of spiders and insects in spruce Picea abies and its relation to predation by birds. Ecography 4, 82-93. .
Jansson, C., Ekman, J., von Brömssen, A., von Bromssen, A., 1981. Winter mortality and food supply in Tits Parus spp. Oikos 37, 313. .
Jedlikowski, J., Brambilla, M., 2017. The adaptive value of habitat preferences from a multi-scale spatial perspective: insights from marsh-nesting avian species. PeerJ 5, e3164. .
Johnson, D.H., 1980. The comparison of usage and availability measurements for evaluating resource preference. Ecology 61, 65-71. .
Jones, J., 2022. Habitat selection studies in avian ecology: a critical review. Auk 118, 557-562.
Krams, I.A., 1996. Predation risk and shifts of foraging sites in mixed willow and Crested Tit flocks. J. Avian Biol. 27, 153. .
Laiolo, P., Caprio, E., Rolando, A., 2003. Effects of logging and non-native tree proliferation on the birds overwintering in the upland forests of north-western Italy. Forest Ecol. Manag. 179, 441-454. .
Lens, L., Adriaensen, F., Dhondt, A.A., 1994a. Age-related hoarding strategies in the Crested Tit Parus cristatus: should the cost of subordination be re-assessed? J. Anim. Ecol. 63, 749. .
Lens, L., Dhondt, A., 1993. Individual variation in mate care by alpha males in crested tit winter flocks. Behav. Ecol. Sociobiol. 33. .
Lens, L., Dhondt, A.A., 1992. Variation in coherence of crested tit winter flocks: an example of multivariate optimization. Acta Oecol. 13, 553-567.
Lens, L., Dhondt, A.A., 2008. Effects of habitat fragmentation on the timing of Crested Tit Parus cristatus natal dispersal. Ibis 136, 147-152. .
Lens, L., Wauters, L.A., Dhondt, A.A., 1994b. Nest-building by crested tit Parus cristatus males: an analysis of costs and benefits. Behav. Ecol. Sociobiol. 35, 431-436. .
Li, N., Wang, Z., Cai, Y., Zhang, L., 2020. Importance of microhabitat selection by birds for the early recruitment of endangered trees in a fragmented forest. Avian Res. 11, 46. .
Maícas, R., Fernández Haeger, J., 2004. Pine plantations as a breeding habitat for a hole-nesting bird species crested tit (Parus cristatus) in southern Spain. Forest Ecol. Manag. 195, 267-278. .
Manly, B.F.J., McDonald, L.L., Thomas, D.L., McDonald, T.L., Erickson, W.P., 2004. Resource Selection by Animals. Kluwer Academic Publisher, New York.
Martin, P., Bateson, P.P.G., 2018. Measuring Behaviour: an Introductory Guide. Cambridge University Press, Cambridge.
Mingozzi, T., Boano, G., Pulcher, C., 1988. Atlante degli Uccelli nidificanti in Piemonte e Val d'Aosta. Monografie Ⅷ, Museo Regionale di Scienze Naturali di Torino.
Mitchell, L.J., Kohler, T., White, P.C.L., Arnold, K.E., 2020. High interindividual variability in habitat selection and functional habitat relationships in European nightjars over a period of habitat change. Ecol. Evol. 10, 5932-5945. .
Morganti, M., Viganò, E., Berlusconi, A., Cioccarelli, S., Fasola, M., 2021. Post-fledging habitat selection of a Purple Heron Ardea purpurea revealed by GPS/GSM telemetry. Avocetta 45, 1. .
QGIS Development Team, 2018. QGIS geographic information system. Open Source Geospatial Foundation Project.
R Core Team, 2020. R: a language and environment for statistical computing. .
Raynor, E.J., Beyer, H.L., Briggs, J.M., Joern, A., 2017. Complex variation in habitat selection strategies among individuals driven by extrinsic factors. Ecol. Evol. 7, 1802-1822. .
Robinson, S.K., Holmes, R.T., 1982. Foraging behavior of forest birds: the relationships among search tactics, diet, and habitat structure. Ecology 63, 1918. .
Summers, R.W., Mavor, R.A., Buckland, S.T., MacLennan, A.M., 1999. Winter population size and habitat selection of Crested Tits Parus cristatus in Scotland. Bird Study 46, 230-242. .
Summers, R.W., Taylor, W., Underhill, L.G., 1993. Nesting habitat selection by Crested Tits Parus cristatus in a pine plantation. Forestry 66, 147-151. .
Thompson, I.D., Hogan, H.A., Montevecchi, W.A., 1999. Avian communities of mature balsam fir forests in Newfoundland: age-dependence and implications for timber harvesting. Condor 101, 311-323. .
Tremblay, I., Thomas, D.W., Lambrechts, M.M., Blondel, J., Perret, P., 2003. Variation in blue tit breeding performance across gradients in habitat richness. Ecology 84, 3033-3043. .
Welsh, D.A., 1987. The influence of forest harvesting on mixed coniferous-deciduous boreal bird communities in Ontario. Acta Oecol. 8, 247-252.

 DownLoad:
DownLoad:






 Email Alerts
Email Alerts RSS Feeds
RSS Feeds