Nest predation is an essential factor affecting bird population density, reproductive ecology and life cycle. However, there are still contradictory results about how nest predation pressure changes with urbanization. One of the reasons for the controversy is that previous studies often use artificial nests to investigate nest predation, but the result of artificial nests is often biased and contrary to that of natural nests. Therefore, it is important to perform nest predation experiments simultaneously with both natural and artificial nests. In this study, we examined the change of nest predation pressure on the Chinese Bulbul (Pycnonotus sinensis) along the urbanization gradient in Hangzhou, China. We detected 148 natural nests of Chinese Bulbuls and continuously monitored them in two continuous breeding seasons from 2012 to 2013. After the breeding season, we placed artificial eggs in natural nests of Chinese Bulbuls to investigate the changes in nest predation pressure and used infrared cameras to record the predators. We then calculated the urbanization synthetic index (USI) as the measure of the degree of urbanization and examined its relationship with nest predation pressure. We found that no matter whether natural nests or artificial eggs were used, the nest predation pressure always decreased significantly with the degree of urbanization. The average height of shrubs within 10 m of the nest differed significantly between the predated and unpredated nests, which also affected the risk of nest predation. Our study highlights the importance of using natural and artificial nests simultaneously to conduct nest predation experiments, which can reduce the bias or errors caused by only using artificial nests. We also recommend the use of infrared cameras in future nest predation studies, which has obvious advantages in monitoring and identifying potential predators.
With industrialization and continuous population growth, we are living in an increasing urbanized world (Johnson and Munshi-South, 2017). Urbanization is the expansion and development of human architecture and manufactured landscapes to natural lands, such as forests, wetlands, and lakes (Marzluff, 2001). However, the expansion of urbanization has considerably impacted local wildlife (Marzluff et al., 2001; Juárez et al., 2020; Yang et al., 2020). Compared with natural habitats, urbanization has changed many environmental factors, such as the heat island effect (Arnfield, 2003), noise and light pollution (Zhang et al., 2020; Joshua et al., 2021), and various human disturbance (Marzluff, 2001), which will affect creatures and pose challenges to the survival of wild animals. However, some species have adapted to urban environments by changing their feeding habits (Fleischer et al., 2003), body size (Liker et al., 2008), vocalization (Wood and Yezerinac, 2006), and breeding behaviors (Wang et al., 2008, 2009).
Predation is one of the primary mechanisms that make up natural communities, shaping the foraging strategies of organisms, reflecting the behavioral response of organisms to the environment (Verdolin, 2006). Human activities can affect wildlife species' population density by altering the predatory pressure in cities (Shochat et al., 2006). For example, artificial night illumination can increase the ability of predators to detect prey in dark conditions (Gaston et al., 2013). Birds are more numerous and easily monitored by observers than other urban animals (Chace and Walsh, 2006), so they are often used as model organisms to study urban predatory ecology (Vincze et al., 2017; Eötvös et al., 2018).
Nest predation, primarily the predation of eggs or nestlings, is often one of the critical causes of reproductive failure in birds (Ricklefs, 1969; Bollmann and Reyer, 2001; Greenwell et al., 2019). As an essential factor influencing bird population dynamics (Jokimäki and Huhta, 2000; Henschke et al., 2016), nest predation can effectively reflect birds' responses to urbanization. In some studies that compared natural environments and artificial landscapes, nest predation rates have been found to increase (López-Flores et al., 2009; Thorington and Bowman, 2003) or decrease (Gering and Blair, 1999; Buxton and Benson, 2015) in urban environments. The "Predator Safe Zone Hypothesis" (Ryder et al., 2010) argues that urban habitats are safer than natural habitats, with fewer nest predation events (Eötvös et al., 2018). However, Jokimäki and Huhta (2000) found that nest predation rates are higher in urbanized centers than in separate residential areas. Sorace (2002) also found a surge in the number of nest predators in cities, leading to increased predation pressure. Therefore, there is still controversy about whether nest predation increases or decreases with urbanization.
Investigations of nest predation often use artificial nests and artificial eggs (e.g., with plasticine or soft pottery; Major et al., 1996), and substitutes (e.g., Japanese Quail (Coturnix japonica; Jokimäki and Huhta, 2000; Borgmann and Rodewald, 2004; Patterson et al., 2016), or farmed Mallard (Anas platyrhynchos; Holopainen et al., 2021)). Since artificial nests are always different from natural nests (Moore and Robinson, 2004; Robinson et al., 2005) in terms of materials, shapes, etc., the conclusions reached with artificial nests are often questioned (Zanette, 2002). Although a few studies have found that nest predation results from artificial nests are sometimes consistent with natural nests (Teglhøj, 2018), others have frequently found that they are different (Sloan et al., 1998; Zanette, 2002; Berry and Lill, 2003; Vincze et al., 2017). According to Vincze's meta-analysis (Vincze et al., 2017), most nest studies either use only natural nests or only artificial nests. Therefore, to reduce such a bias, nest predation experiments should be performed simultaneously with natural nests and artificial nests.
The Chinese Bulbul (Pycnonotus sinensis), a small bird in the order Passeriformes, is one of the most widely distributed birds in southern China and East Asia (Zhuge, 1990). It is an urban adapter that can even inhabit highly urbanized areas (Wang et al., 2009). Previous studies show that Chinese Bulbuls can adapt to urbanization in several ways, such as using artificial human food (Wang, 2003), changing nest materials (Wang et al., 2009), and producing a variety of microgeographic song dialects in the city (Ding and Jiang, 2005). However, it is unclear whether the adaptation of Chinese Bulbuls to the city is related to nest predation.
In this study, we investigated the change of nest predation on the Chinese Bulbul along the urbanization gradient in Hangzhou, China. We first monitored natural breeding nests of Chinese Bulbuls to examine the change in the predation pressure along the urban gradient. We then conducted the artificial nest predation experiment by placing artificial eggs to the natural nests of Chinese Bulbuls immediately after the breeding season. We addressed the following three questions: (1) Does the nest predation pressure of Chinese Bulbuls increase or decrease along the urbanization gradient? (2) Are the results of the natural breeding nests consistent with the artificial nest predation experiment? (3) Which habitat characteristics might affect the nest predation pressure of Chinese Bulbuls?
2.
Materials and methods
2.1
Study area
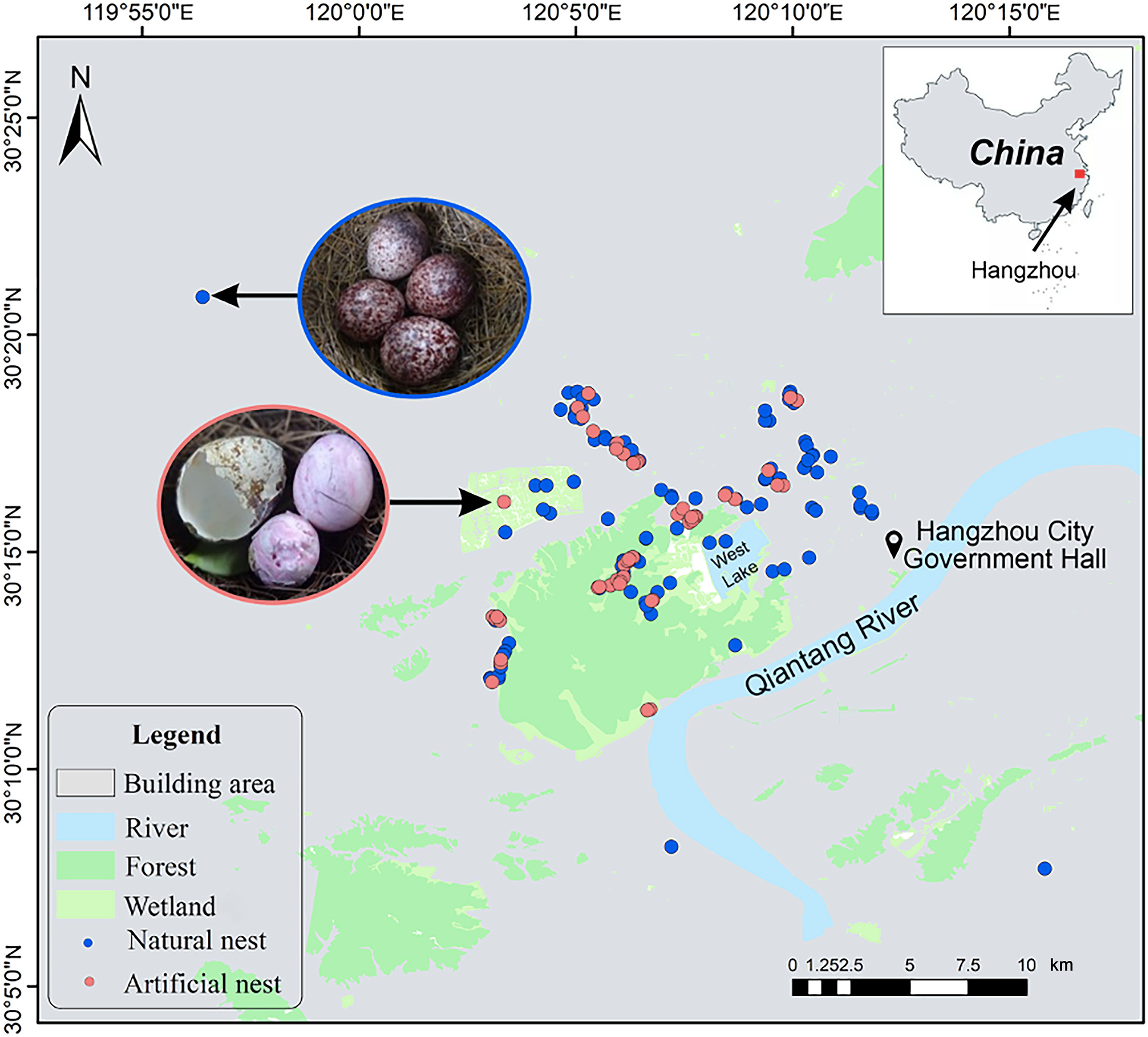
This study was conducted in the Hangzhou city, Zhejiang Province, China (29°11′–30°33′ N, 118°21′–120°30′ E; Fig. 1). As the political, economic and cultural center of Zhejiang Province, Hangzhou is one of the most urbanized cities in the Yangtze River Delta region, China (Wang et al., 2009, 2013). The study area is 774.82 km2 and has a total population of 3.614 million by 2019. As of 2019, the urban population of Hangzhou accounted for 78.5% of the total population (urbanization rate). The city's gross domestic product (GDP) reached RMB 1537.3 billion in 2019. The city is covered with hills, plains and water systems (e.g., rivers, lakes, and ponds), representing 65.6%, 26.4%, and 8.0% of the total area, respectively (Chen et al., 2000).
Figure
1.
Distribution map of the nest sites of the Chinese Bulbul (Pycnonotussinensis) in Hangzhou, China.
From March to July in 2012 and 2013, we searched for natural breeding nests of Chinese Bulbuls along the urbanization gradient in Hangzhou from the urban area with main buildings to the western mountainous region with an excellent natural environment (Fig. 1). We determined whether Chinese Bulbuls begun to breed by listening to the chirping and observing the behavior of carrying nest materials with the telescope (Wang et al., 2009). First, we searched the nests of Chinese Bulbuls based on its nest site selection (Wang et al., 2003; Chen et al., 2006). If there was an old nest, there is a high probability of nesting in the vicinity in the current year. Second, we searched the nests by checking the favorite nesting trees of Chinese Bulbuls, such as the Osmanthus tree (Lan et al., 2013). Third, we observed the emergence of carrying nest materials to locate the nests of Chinese Bulbuls (Wang et al., 2009). After locating a breeding nest of the Chinese Bulbul, we recorded it by a GPS and marked the nest tree for further monitoring. The nests are 2–4 m above the ground and in the shape of a tall cup built from hay stems, fine roots, and plant fibers (Zhuge, 1990).
We monitored the breeding nest every other day with a camera tied to the top of a fishing rod to observe and record the situation inside the nest (Lan et al., 2013). Each nest check confirmed the reproductive status of the Chinese Bulbul, including missing eggs, missing or dead nestlings. Finally, through human observation and multiple re-examinations by using cameras, we considered the loss of eggs and nestlings as nest predation incidents (Zhang, 2014). We monitored the nest in the absence of adult birds to reduce the potential effect of human intervention on breeding nests.
In addition, we measured several nest-site characteristics that might affect the nest predation pressure of Chinese Bulbuls. Some studies found that nest site location (O'hUallachaín, 2014), nest height from the ground (Chen et al., 2006), the visibility of the nest (Grégoire et al., 2003), and vegetation coverage near the nest (Davis, 2005; Malpass et al., 2015) can significantly affect the predation pressure. The coverage of shrubs may also affect the richness of rodent predators of bird nests (Johnson and Karels, 2015). Therefore, we measured the vegetation coverage below the nest by standing directly below the nest to observe the nest being obscured by vegetation (Zhang, 2014). Similarly, the vegetation coverage above the nest was estimated using the camera observation of the shade on the top of the nest (Chen et al., 2006). We measured nest height from the ground and tree height by using fishing rods on the vertical ground (Zhang, 2014). We also measured several related variables within 10 m of the nest site, such as average height of arbors (e.g., Cinnamomum camphora, Acer spp.), the proportion of arbors, average height of shrubs (e.g., Ligustrum quihoui, Euonymus alatus), and the proportion of shrubs (Table 1).
Table
1.
Difference in habitat variables between predated and unpredated nests of the Chinese Bulbul (Pycnonotus sinensis) in Hangzhou, China.
After completing the monitoring and investigation of the breeding nests of Chinese Bulbuls, we immediately conducted the artificial nest predation experiment by using 54 nests in areas with high and low urbanization indices (according to the urbanization synthetic index, see 2.4.). We placed two fake eggs and two Common Quail (Coturnix coturnix) eggs in the natural nest of the Chinese Bulbul because the clutch size was 4 or 3 in our study region (Lan et al., 2013; Zhang, 2014). The fake eggs were kneaded with soft pottery to form a shape that was the same size as the real eggs. The purpose of placing quail eggs was to prevent some predators from having the ability to identify and reject soft pottery simulated eggs (Zhang, 2014). We can judge the predator category by the bite marks left by predators on delicate pottery and quail eggs (Zanette, 2002).
Meanwhile, we selected 20 of the 54 artificial nests to arrange infrared cameras (Fig. 1) along the urbanization gradient to record potential predators. The duration of the experiment was two weeks in June 2013, which is the natural incubation time of Chinese Bulbuls (Zhuge, 1990; Lan et al., 2013). We checked infrared cameras every seven days to record nest predation. Then we took back the infrared camera and simulated eggs on day 14, the last day of the experiment. We determined whether the nests were preyed upon by observation and playback footage, identifying predator species based on the traces on the eggs and the pictures taken by the infrared camera (Zhang, 2014, Fig. 2).
Figure
2.
Predator species of nests of Chinese Bulbuls recorded by infrared cameras. (A) Red-billed Blue Magpie (Urocissa erythrorhyncha); (B) Unidentified mouse (Muroidea sp.); (C) Red-bellied Squirrel (Callosciurus erythraeus); (D) Signs left by unidentified mice. (For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
To provide a quantitative measure of urbanization intensity, we selected and measured four characteristic parameters of urbanization: building index (BI), environmental noise (EN), disturbance index (DI), and distance to the city center (DCC) (Chen et al., 2000; Wang et al., 2008, 2009).
The building index was constructed according to the method of Bolger et al. (1997). We imported the located nest into a GIS (ArcView 3.3) using the background map in 2013 and measured the area covered by buildings around each nest with a radius of 250 m, 500 m, 1000 m, and 2000 m. The proportion of buildings in the 250 m was given a weight of 1.0; the proportion in the 500 m was given a weight of 0.5, in the 1000 m a weight of 0.25, and in the 2000 m a weight of 0.125 (Bolger et al., 1997). The weighted proportions were then summed to form the building index (Wang et al., 2009).
We chose one day on weekdays and one day on weekends to measure the environmental noise during the breeding period. The value of the ambient noise (dBA) was measured using a HS5633A digital sound level meter within 10 m of the nest from 8:00 to 10:00 a.m. (Wang et al., 2008, 2013). We recorded the numerical value every 5 s interval, and a total of 30 times were recorded to calculate the average. The ambient noise value was calculated using the following formula (Zhang, 2014): environmental noise (EN) = average ambient noise during the weekdays × 5/7 + average ambient noise during the weekends × 2/7.
We chose one day on weekdays and one day on weekends to measure the human disturbance. Specifically, we measured the number of vehicles passing by and counted the total number of people who passed within 10 m of the nests in 1 min from 8:00 to 10:00 a.m. in the morning (Wang et al., 2009). Average traffic flow was calculated as: traffic flow on weekdays × 5/7 + traffic flow on weekends × 2/7 (Zhang, 2014). Similarly, average foot traffic = foot traffic on weekdays × 5/7 + foot traffic on weekends × 2/7. Disturbance index (DI) was then calculated as: average traffic flow + average foot traffic (Zhang, 2014).
The Hangzhou City Government Hall (30°14′54.39″ N, 120°12′22.47″ E) is basically located in the city center of Hangzhou (Wang et al., 2008, 2009, Fig. 1). We estimated the distance to city center (DCC), i.e., the distance from the nest to the city center of Hangzhou (km) using the Google map.
To standardize the four urbanization characteristic parameters, we adjusted them to the value that increases from 0 to 100 with the increase of urbanization (Chen et al., 2000; Wang et al., 2008). First, the value of BI was between 0 and 2, so it was multiplied by 100 and divided by 2. Second, the value of EN was between 0 and 100, so it was not adjusted. Third, the value of DI was between 0 and 300, which was divided by 3. In addition, the value of DCC was between 1 and 20, after taking the reciprocal and multiplying by 100, it increased from 0 to 100 with the increase of urbanization (Chen et al., 2000; Wang et al., 2009). Finally, we combined the above four urbanization characteristic variables to construct an urbanization synthetic index (USI) as follows: USI = BI × 100/2 + EN + DI/3 + 1/DCC × 100 (Chen et al., 2000; Wang et al., 2008).
2.5
Data analyses
In a meta-analysis, Vincze et al. (2017) found that all artificial nests were considered as predated when at least one egg/offspring died, while in natural nests usually only complete brood loss was counted as a predation event in 97% of the studies. However, such different definitions of predation for artificial and natural nests may lead to contradictory results on how nest predation pressure changes with urbanization (Vincze et al., 2017). To avoid this bias, we defined nest predation events for artificial and natural nests in the same way. That is, a nest was considered as predated if at least one egg/offspring died. The advantage of the simple definition makes the results of artificial and natural nests comparable.
We then converted the occurrence of nest predation events into a two-valued response variable (no nest predation = 0, nest predation occurred = 1). We performed binary logistic regression to analyze the influence of urbanization synthetic index (USI) on nest predation. The logistic regression is considered as the most appropriate and flexible statistical test for analyzing factors influencing artificial nest losses (Lewis, 2004).
We determined the role of habitat characteristics in affecting the nest predation risk of Chinese Bulbuls in the following three steps. First, prior to parametric tests, we used one sample K–S test to examine whether the eight habitat characteristics were normally distributed (Table 1). Second, for the three normally distributed variables, we used the t-test to compare their differences between predated and unpredated nests (Table 1). By contrast, we used the Mann-Whitney U test to compare the other five variables that were not normally distributed (Table 1). Finally, we included the habitat variables that differed significantly between predated and unpredated nests in the logistic regression model to examine their influences on the risk of nest predation. All analyses were performed in R 4.1.0 (R Core Team, 2021).
3.
Results
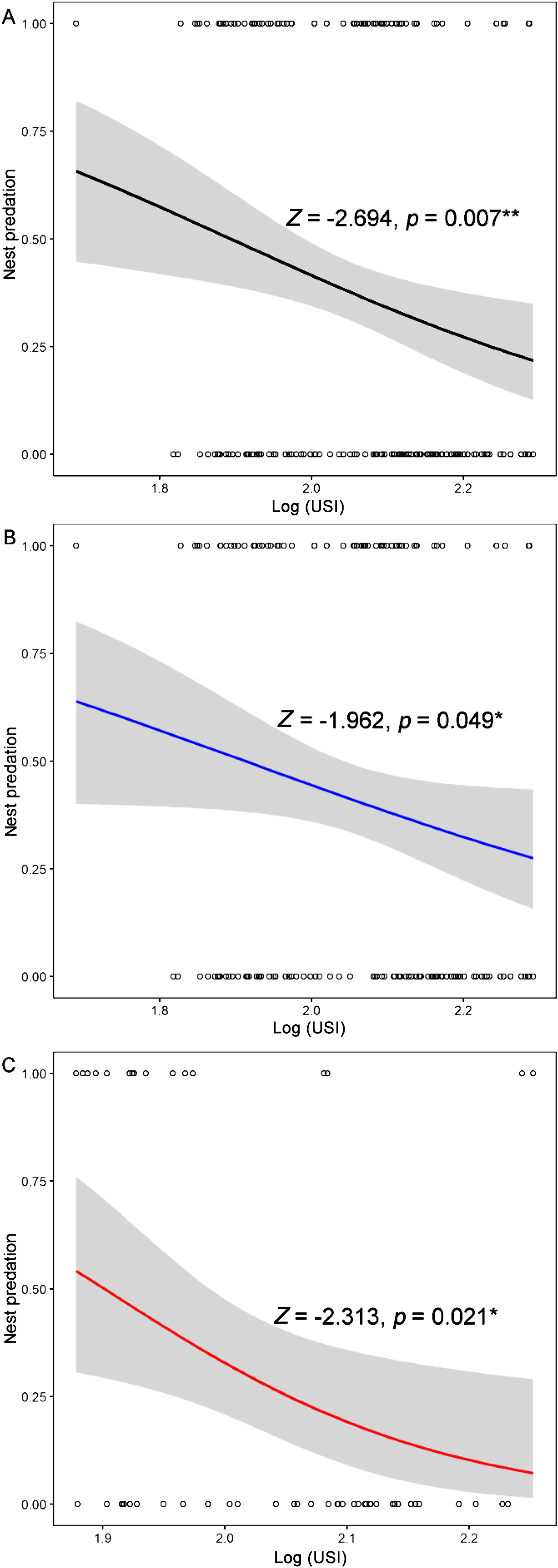
The binary logistic regression analysis showed that the nest predation risk of the Chinese Bulbul decreased significantly with the urbanization synthetic index (USI) for the combined data of natural and artificial nests (Z = −2.694, P = 0.007; Fig. 3A).
Figure
3.
The relationship between the probability of occurrence of nest predation for Chinese Bulbuls and the urbanization synthetic index (USI, log transformed). Open circles represent observed values. (A) Total nest predation; (B) Natural nest predation; (C) Artificial nest predation.
After removing the abandoned nests due to human disturbance, we obtained 148 well-monitored breeding nests of Chinese Bulbuls, among which 61 (41%) were preyed upon by predators. Using the binary logistic regression, we found that the urbanization synthetic index (USI) had a significantly negative effect on the occurrence of nest predation events (Z = −1.962, P = 0.049; Fig. 3B).
3.2
Artificial nest predation
According to infrared camera, of all the nests where artificial eggs were placed, there were 16 nests where predation events occurred in areas with low urbanization levels, accounting for 29.63% of the total experimental nest count (N = 54). The recovery of fake eggs and the traces on eggshells through infrared cameras from 20 nests revealed that the predator species were mainly Red-billed Blue Magpie (Urocissa erythrorhyncha), Red-bellied Squirrel (Callosciurus erythraeus), and unidentified mouse (Muridae sp.) (Fig. 2). However, none of the nests in highly urbanized environments were preyed upon by predators (Fig. 1). The logistic regression analysis showed that the urbanization synthetic index (USI) was significantly and negatively correlated with the predation risk in the artificial nest predation experiment (Z = −2.313, P = 0.021; Fig. 3C).
3.3
Risk of nest predation and habitat characteristics
For the eight habitat variables potentially influencing the nest predation risk of Chinese Bulbuls, only the average height of shrubs within 10 m of the nest differed significantly between predated and unpredated nests (t = −2.179, P = 0.031; Table 1). The logistic regression analysis showed that with the increasing average height of shrubs, the nest is more likely to be predated (Z = 2.333, P = 0.020).
4.
Discussion
In this study, we examined the change of nest predation on the Chinese Bulbul using both natural and artificial nests simultaneously along the urbanization gradient in Hangzhou, China. We found that whether natural nests or artificial eggs were used, the nest predation pressure always decreased with the degree of urbanization. The average height of shrubs within 10 m of the nest also affected the risk of nest predation on Chinese Bulbuls.
Our study differs from most previous nest predation studies in two ways. First, we performed nest predation experiments on Chinese Bulbuls using both natural and artificial nests simultaneously. In previous studies, artificial nests were often used by researchers to study nest predation (Jokimäki and Huhta, 2000; López-Flores et al., 2009; Gómez-Catasús et al., 2021). However, results obtained from artificial nests were often biased and contrary to that of natural nests (Sloan et al., 1998; Berry and Lill, 2003; Vincze et al., 2017). Zanette (2002) even suggests that researchers should avoid inferring about predation on natural nests based only on artificial nest studies. Second, the nests used in the artificial nest predation experiment are the natural nests of Chinese Bulbuls immediately after the breeding season. In contrast, previous studies often used bird nests made entirely of artificial materials (e.g., soils, tennis balls; Gering and Blair, 1999; Zanette, 2002; Vincze et al., 2017; Krüger et al., 2018). The above two advantages of our study can effectively reduce potential biases or errors occurred in previous nest predation studies.
We found that the nest predation pressure of Chinese Bulbuls decreased with the degree of urbanization regardless of whether natural nests or artificial nests were used. The results were consistent with the findings of Gering and Blair (1999), Morgan et al. (2011) and Brahmia et al. (2013) where high predation rates occurred in rural areas. However, our results were contrary to the conclusions of Leston and Rodewald (2006) and Kaisanlahti-Jokimäki et al. (2012), who found that rural areas experienced lower nest predation pressures than urban areas. These contrary results may be related to the use of artificial nests and natural nests (Berry and Lill, 2003; Vincze et al., 2017). Artificial nests may attract different predators than natural nests (Willebrand and Marcstr, 1988). Artificial nests also lack the protection of parent birds (Zanette, 2002; Vincze et al., 2017). Moreover, the use of fake eggs can also affect predator's choices; Haskell et al. (2001) found that the probability of predation in artificial nests containing quail eggs does not change significantly with increasing housing density but is related to the presence of some predators. However, there was no difference in predation pressure when using both natural and artificial nests (Grégoire et al., 2003).
Several reasons may explain why the nest predation pressure of Chinese Bulbuls decreased with the degree of urbanization in our study. First, Gray's increasing disturbance hypothesis (Gray, 1989) posits that there would be a loss of species along the urban-rural gradient, which means the number of predators in the city will decrease. Our artificial nest predation experiments support this hypothesis because no case of nest predation has occurred in highly urbanized areas. The predators have been photographed only in rural areas with a low urbanization index. Meanwhile, our results also support the "urban areas as predator-free zones" theory, where urban areas are refuges for prey against predators. Predators are sometimes more afraid of the human approach (Møller, 2012). Second, increasing urbanization will bring more complementary food to animals (Shochat et al., 2006; Chamberlain et al., 2009), which improves the physical condition of the parent birds and reduces the energy requirements for pre-breeding birds. This may increase the time for the parents to hatch eggs in the nest, shortening the incubation period and thus reducing the risk of nest predation (Lothery et al., 2014). Lastly, urban resources can divert the attention of potential predators, leading to reduced predation pressure in urban areas (Friesen et al., 2013).
We found that the average height of shrub within 10 m of the nest differed significantly between predated and unpredated nests. Nest predation pressure of Chinese Bulbuls had an increasing trend with increasing shrub height. The results can be explained by the following reasons. First, shrubs with higher height can provide more shelter and food for rodents (Lu and Zhang, 2004), one of the primary nest predators of Chinese Bulbuls. Second, shrub height is one of the most important factors determining rodent community structure (Wu et al., 2021). Rodents in low shrubs are more likely to be spotted by their predators, so rodents may tend to stay in tall shrubs. In addition, previous studies have found that Red-billed Blue Magpies, another nest predator of Chinese Bulbuls, tend to stay in the shrubs after feeding (Jin et al., 2021). Overall, taller shrubs may accompany more nest predators of Chinese Bulbuls, so nests with higher shrubs have greater predation pressure.
5.
Conclusions
In the context of increasing urbanization worldwide, nest predation is important in understanding the population dynamics of avian species in urbanized landscape. We found that regardless of whether natural nests or artificial eggs were used, the nest predation pressure of Chinese Bulbuls always decreased with the increase of urbanization. However, experiments using artificial nests are often biased and contrary to the results of natural nests (Sloan et al., 1998; Berry and Lill, 2003; Vincze et al., 2017). Therefore, our study highlights the importance of using natural and artificial nests simultaneously to conduct predatory pressure experiments, which can reduce the bias or errors caused by only using artificial nests. We also recommend the use of infrared cameras in future nest predation studies (Krüger et al., 2018), which has obvious advantages in monitoring and identifying potential predators and investigating the impact of urbanization on nest predation.
Authors' contribution
YW and SC conceived the study; QZ and SL collected data and conducted the research; XC and YW developed methods, analyzed data, and wrote the manuscript. All authors read and approved the final manuscript.
Declaration of competing interest
The authors declare that they have no competing interests.
Acknowledgements
We thank three anonymous reviewers for their helpful comments on the manuscript. We are grateful for Qin Huang, Yiwei Lu, Tengda Zhu, Yafei Ma, Jianwei Zhou, Chao Chen, Xiang Yu, Wen Xu, Mingming Hu and Chenchen Zhu for their field assistance in finding and monitoring bird nests. This study was supported by the National Natural Science Foundation of China (Grant No. 31971545, 31071908).
Arnfield, A.J., 2003. Two decades of urban climate research: a review of turbulence, exchanges of energy and water, and the urban heat island. Int. J. Climatol. 23, 1-26.
Berry, L., Lill, A., 2003. Do predation rates on artificial nests accurately predict predation rates on natural nests? The effects of nest type, egg type and nest-site characteristics. Emu 103, 207-214.
Bolger, D.T., Scott, T.A., Rotenberry, J.T., 1997. Breeding bird abundance in an urbanizing landscape in coastal Southern California. Conserv. Biol. 11, 406-421.
Bollmann, K., Reyer, H.U., 2001. Reproductive success of water pipits in an alpine environment. Condor 103, 510-520.
Borgmann, K.L., Rodewald, A.D., 2004. Nest predation in an urbanizing landscape: the role of exotic shrubs. Ecol. Appl. 14, 1757-1765.
Brahmia, Z., Scheifler, R., Crini, N., Maas, S., 2013. Breeding performance of blue tits (Cyanistes cæruleus ultramarinus) in relation to lead pollution and nest failure rates in rural, intermediate, and urban sites in Algeria. Environ. Pollut. 174, 171-178.
Buxton, V.L., Benson, T.J., 2015. Do natural areas in urban landscapes support successful reproduction by a group of conservation priority birds? Anim. Conserv. 18, 471-479.
Chace, J.F., Walsh, J.J., 2006. Urban effects on native avifauna: a review. Landscape Urban Plan. 74, 46-69.
Chamberlain, D.E., Cannon, A.R., Toms, M.P., Leech, D.I., Hatchwell, B.J., Gaston, K.J., 2009. Avian productivity in urban landscapes: a review and meta-analysis. Ibis 151, 1-18.
Chen, S., Ding, P., Zheng, G., Zhuge, Y., 2000. Effects of urbanization on wetland waterbird communities in Hangzhou. Zool. Res. 21, 279-285.
Chen, W., Wu, Y., Hu, J., Yu, Z., Guo, Z., 2006. Nest site selection and materials of Pycnonotus sinensis in Nanchong, Sichuan. Sichuan J. Zool. 25, 590-593.
Davis, S.K., 2005. Nest-site selection patterns and the influence of vegetation on nest survival of mixed-grass prairie passerines. Condor 107, 605-606.
Ding, P., Jiang, S., 2005. Microgeographic song variation in the Chinese Bulbul (Pycnonotus sinenesis) in urban areas of Hangzhou city. Zool. Res. 26, 453-459.
Fleischer, A.L., Bowman, R., Woolfenden, G.E., 2003. Variation in foraging behavior, diet, and time of breeding of Florida scrub-jays in suburban and wildland habitats. Condor 105, 515-527.
Friesen, L.E., Casbourn, G., Martin, V., Mackay, R.J., 2013. Nest predation in an anthropogenic landscape. Wilson J. Ornithol. 125, 562-569.
Gaston, K.J., Bennie, J.B., Davies, T.W., Hopkins, J., 2013. The ecological impacts of nighttime light pollution: a mechanistic appraisal. Biol. Rev. 88, 912-927.
Gering, J.C., Blair, R.B., 1999. Predation on artificial bird nests along an urban gradient: predatory risk or relaxation in urban environments? Ecography 22, 532-541.
Gómez-Catasús, J., Barrero, A., Reverter, M., Bustillo-de la Rosa, D., Pérez-Granados, C., Traba, J., 2021. Landscape features associated to wind farms increase mammalian predator abundance and ground-nest predation. Biodivers. Conserv. 30, 2581-2604.
Gray, J.S., 1989. Effects of environmental stress on species rich assemblages. Biol. J. Linn. Soc. 37, 19-32.
Greenwell, C.N., Calver, M.C., Loneragan, N.R., 2019. Cat gets its tern: a case study of predation on a threatened coastal seabird. Animals 9, 445.
Grégoire, A., Garnier, S., Dréano, N., Faivre, B., 2003. Nest predation in Blackbirds (Turdus merula) and the influence of nest characteristics. Ornis Fennica 80, 1-10.
Haskell, D.G., Knupp, A.M., Schneider, M.C., 2001. Nest predator abundance and urbanization. In: Marzluff, J.M., Bowman, R., Donelly, R. (Eds. ), Avian Ecology and Conservation in an Urbanizing World. Kluwer Academic Publishers, Boston, pp. 243-258.
Henschke, S.J., Bayne, E.M., Ball, J.R., 2016. Post-predation parental behavior of boreal songbirds. Wilson J. Ornithol. 128, 766-774.
Holopainen, S., Väänänen, V.M., Vehkaoja, M., Fox, A.D., 2021. Do alien predators pose a particular risk to duck nests in Northern Europe? Results from an artificial nest experiment. Biol. Invasions 23, 3795-3807.
Jin, B., Liu, H., Lv, T., Wang Y., Jiang, P., Wang, Z., 2021. Fruits foraging and dispersal of Machilus ichangensis by frugivorous birds in Nanjing Botanical Garden Mem. Sun Yat-Sen. Ecol. Sci. 40, 149-154.
Johnson, A.M., Karels, T.J., 2015. Partitioning the effects of habitat fragmentation on rodent species richness in an urban landscape. Urban Ecosyst. 19, 547-560.
Johnson, M.T.J., Munshi-South, J., 2017. Evolution of life in urban environments. Science 358, eaam8327.
Jokimäki, J., Huhta, E., 2000. Artificial nest predation and abundance of birds along an urban gradient. Condor 102, 838-847.
Joshua, G., Ali, Z., Ayub, M., Nadeem, S.I., 2021. Heavy metal contamination in wild avian species inhabiting human-modified habitats. Environ. Monit. Assess. 193, 588.
Juárez, R., Chacón-Madrigal, E., Sandoval, L., 2020. Urbanization has opposite effects on the territory size of two passerine birds. Avian Res. 11, 11.
Kaisanlahti-Jokimaki, M.L., Jokimaki, J., Huhta, E., Siikamaki, P., 2012. Impacts of seasonal small-scale urbanization on nest predation and bird assemblages at tourist destinations. In: Lepczyk, C.A., Warren, P.S. (Eds. ), Urban Bird Ecology and Conservation. University of California Press, Berkeley, CA, pp. 93-110.
Kruger, H., Väänänen, VM., Holopainen, S., Nummi, P., 2018. The new faces of nest predation in agricultural landscapes-a wildlife camera survey with artificial nests. Eur. J. Wildlife Res. 64, 76.
Lan, S.S., Zhang, Q., Huang, Q., Chen, S., 2013. Breeding ecology of Chinese Bulbul in the urban environment of Hangzhou, China. Zool. Res. 34, 182-189.
Leston, L.F.V., Rodewald, A.D., 2006. Are urban forests ecological traps for understory birds? An examination using northern cardinals. Biol. Conserv. 131, 566-574.
Lewis, K.P., 2004. How important is the statistical approach for analysing categorical data? A critique using artificial nests. Oikos 104, 305-315.
Liker, A., Papp, Z., Bokony, V., Lendvai, A.Z., 2008. Lean birds in the city: body size and condition of house sparrows along the urbanization gradient. J. Anim. Ecol. 77, 789-795.
López-Flores, V., MacGregor-Fors, I., Schondube, J.E., 2009. Artificial nest predation along a neotropical urban gradient. Landscape Urban Plan. 92, 90-95.
Lothery, C.J., Thompson, C.F., Lawler, M.L., Sakaluk, S.K., 2014. Food supplementation fails to reveal a trade-off between incubation and self-maintenance in female House Wrens. PLoS One 9, e106260.
Lu, J.Q., Zhang, Z.B., 2004. Effects of habitat and season on removal and hoarding of seeds of wild apricot (Prunus armeniaca) by small rodents. Acta Oecol. 26, 247-254.
Major, R.E., Gowing, G., Kendal, C.E., 1996. Nest predation in Australian urban environments and the role of the Pied Currawong, Strepera Graculina. Aust. J. Ecol. 21, 399-409.
Malpass, J.S., Rodewald, A.D., Matthews, S.N., 2015. Woody cover does not promote activity of nest predators in residential yards. Landscape Urban Plan. 135, 32-39.
Marzluff, J.M., 2001. Worldwide urbanization and its effects on birds. In: Marzluff, J.M., Bowman, R., Donelly, R. (Eds. ), Avian Ecology and Conservation in an Urbanizing World. Kluwer Academic Publishers, Boston, pp. 19-47.
Marzluff, J.M., Bowman, R., Donelly, R. 2001. A historical perspective on urban bird research: trends, terms, and approaches. In: Marzluff, J.M., Bowman, R., Donelly, R. (Eds. ), Avian Ecology and Conservation in an Urbanizing World. Kluwer Academic Publishers, Boston, pp. 1-17.
Moeller, A.R., 2012. Urban areas as refuges from predators and flight distance of prey. Behav. Ecol. 23, 1030-1035.
Moore, R.P., Robinson, W.D., 2004. Artificial bird nests, external validity, and bias in ecological field studies. Ecology 85, 1562-1567.
Morgan, D.K.J., Waas, J.R., Innes, J., Fitzgerald, N., 2011. Identification of nest predators using continuous time-lapse recording in a New Zealand city. New Zeal. J. Zool. 38, 343-347.
O'hUallachain, D., 2014. Nest site location and success rates of an urban population of Woodpigeon (Columba Palumbus) in Ireland. Biol. Environ. 114B, 13-17.
Patterson, L., Kalle, R., Downs, C., 2016. Predation of artificial bird nests in suburban gardens of Kwazulu-Natal, South Africa. Urban Ecosyst. 19, 615-630.
R Core Team, 2021. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. .
Ricklefs, R., 1969. Analysis of nesting mortality in birds. Smithsonian Contrib. Zool. 9, 1-48.
Robinson, W.D., Styrsky, J.N., Brawn, J.D., 2005. Are artificial bird nests effective surrogates for estimating predation on real bird nests? A test with tropical birds. Auk 122, 843-852.
Ryder, T.B., Reitsma, R., Evans, B., Marra, P., 2010. Quantifying avian nest survival along an urbanization gradient using citizen- and scientist-generated data. Ecol. Appl. 20, 419-426.
Shochat, E., Warren, P.S., Faeth, S.H., McIntyre, N.E., Hope, D., 2006. From patterns to emerging processes in mechanistic urban ecology. Trends Ecol. Evol. 21, 187-191.
Sloan, S.S., Holmes, R.T., Sherry, T.W., 1998. Depredation rates and predators at artificial bird nests in an unfragmented northern hardwoods forest. J. Wildlife Manage. 62, 529-539.
Sorace, A., 2002. High density of bird and pest species in urban habitats and the role of predator abundance. Ornis Fennica 79, 60-71.
Teglhøj, P.G., 2018. Artificial nests for barn swallows Hirundo rustica: a conservation option for a declining passerine? Hous. Theor. Soc. 65, 385-395.
Thorington, K.K., Bowman, R., 2003. Predation rate on artificial nests increases with human housing density in suburban habitats. Ecography 26, 188-196.
Verdolin, J.L., 2006. Meta-analysis of foraging and predation risk trade-offs in terrestrial systems. Behav. Ecol. Sociobiol. 60, 457-464.
Vincze, E., Seress, G., Lagisz, M., Nakagawa, S., Dingemanse, N.J., Sprau, P., 2017. Does urbanization affect predation of bird nests? A meta-analysis. Front. Ecol. Evol. 5, 1-12.
Wang Y., 2003. Study on adaptations of birds in Hangzhou to urbanization. Master's Thesis. Zhejiang University, Hangzhou.
Wang, Y., Chen, S., Ding, P., 2003. Breeding birds and their nests in street tree strips in Hangzhou city. Zool. Res. 24, 259-264.
Wang, Y., Chen, S., Jiang, P., Ding, P., 2008. Black-Billed Magpies (Pica pica) adjust nest characteristics to adapt to urbanization in Hangzhou, China. Can. J. Zool. 86, 676-684.
Wang, Y., Chen, S., Blair, R.B., Jiang, P., Ding, P., 2009. Nest composition adjustments by Chinese Bulbuls (Pycnonotus sinensis) in an urbanized landscape of Hangzhou (E China). Acta Ornithol. (Wars.) 44, 185-192.
Wang, Y., Ding, P., Chen, S., Zheng, G., 2013. Nestedness of bird assemblages on urban woodlots: implications for conservation. Landscape Urban Plan. 111, 59-67.
Willebrand, T., Marcstr, V., 1988. On the danger of using dummy nests to study predation. Auk 105, 378-379.
Wood, W.E., Yezerinac, S.M., 2006. Song Sparrow (Melospiza melodia) song varies with urban noise. Auk 123, 650-659.
Wu, H., Liu, D., Wei, M., Qi, Y., Shao, C., Chu, H., 2021. Community diversities of rodents and its relationship with environmental factors in the Mt. Kalamaili Ungulate Nature Reserve, Xinjiang. Acta Ecol. Sin. 41, 9511-9524.
Yang, X., Tan, X., Chen, C., Wang, Y., 2020. The influence of urban park characteristics on bird diversity in Nanjing, China. Avian Res. 11, 45.
Zanette, L., 2002. What do artificial nests tells us about nest predation? Biol. Conserv. 103, 323-329.
Zhang, Q., 2014. Responses of life history traits of Chinese Bulbul to urbanization. Master's Thesis. Zhejiang Normal University, Jinhua.
Zhang, F.S., Wang, Y., Wu, K., Xu, W.Y., Wu, J., Liu, Y.J., Wang, X.Y., Shuai, L.Y., 2020. Effects of artificial light at night on foraging behavior and vigilance in a nocturnal rodent. Sci. Total. Environ. 724, 138271.
Zhuge, Y., 1990. Fauna of Zhejiang: Aves. Zhejiang Science and Technology Press, Hangzhou.

 DownLoad:
DownLoad:






 Email Alerts
Email Alerts RSS Feeds
RSS Feeds