Fenliang Kuang, Wei Wu, David Li, Chris J. Hassell, Grace Maglio, Kar-Sin K. Leung, Jonathan T. Coleman, Chuyu Cheng, Pavel S. Tomkovich, Zhijun Ma. 2022: Detecting the non-breeding region and migration route of Whimbrels (Numenius phaeopus rogachevae) in the East Asian–Australasian Flyway. Avian Research, 13(1): 100011. DOI: 10.1016/j.avrs.2022.100011
Citation:
Fenliang Kuang, Wei Wu, David Li, Chris J. Hassell, Grace Maglio, Kar-Sin K. Leung, Jonathan T. Coleman, Chuyu Cheng, Pavel S. Tomkovich, Zhijun Ma. 2022: Detecting the non-breeding region and migration route of Whimbrels (Numenius phaeopus rogachevae) in the East Asian–Australasian Flyway. Avian Research, 13(1): 100011. DOI: 10.1016/j.avrs.2022.100011
Fenliang Kuang, Wei Wu, David Li, Chris J. Hassell, Grace Maglio, Kar-Sin K. Leung, Jonathan T. Coleman, Chuyu Cheng, Pavel S. Tomkovich, Zhijun Ma. 2022: Detecting the non-breeding region and migration route of Whimbrels (Numenius phaeopus rogachevae) in the East Asian–Australasian Flyway. Avian Research, 13(1): 100011. DOI: 10.1016/j.avrs.2022.100011
Citation:
Fenliang Kuang, Wei Wu, David Li, Chris J. Hassell, Grace Maglio, Kar-Sin K. Leung, Jonathan T. Coleman, Chuyu Cheng, Pavel S. Tomkovich, Zhijun Ma. 2022: Detecting the non-breeding region and migration route of Whimbrels (Numenius phaeopus rogachevae) in the East Asian–Australasian Flyway. Avian Research, 13(1): 100011. DOI: 10.1016/j.avrs.2022.100011
Ministry of Education Key Laboratory for Biodiversity Science and Ecological Engineering, National Observations and Research Station for Wetland Ecosystems of the Yangtze Estuary, Institute of Eco-Chongming, School of Life Sciences, Fudan University, Shanghai, 200433, China
b.
School of Resources Environment and Chemistry, Chuxiong Normal University, Chuxiong, Yunnan, 675000, China
c.
Shanghai Chongming Dongtan National Nature Reserve, Shanghai, 202183, China
d.
National Parks Board, Singapore, 718925, Singapore
e.
Global Flyway Network, PO Box 3089, Broome, WA, 6725, Australia
f.
Australasian Wader Studies Group, Broome, WA, 6725, Australia
g.
Hong Kong Waterbirds Ringing Group, Hong Kong, 999077, China
h.
Queensland Wader Study Group, 22 Parker Street, Shailer Park, QLD, 4128, Australia
i.
Zoological Museum, Lomonosov Moscow State University, Bolshaya Nikitskaya Str. 2, Moscow, 125009, Russia
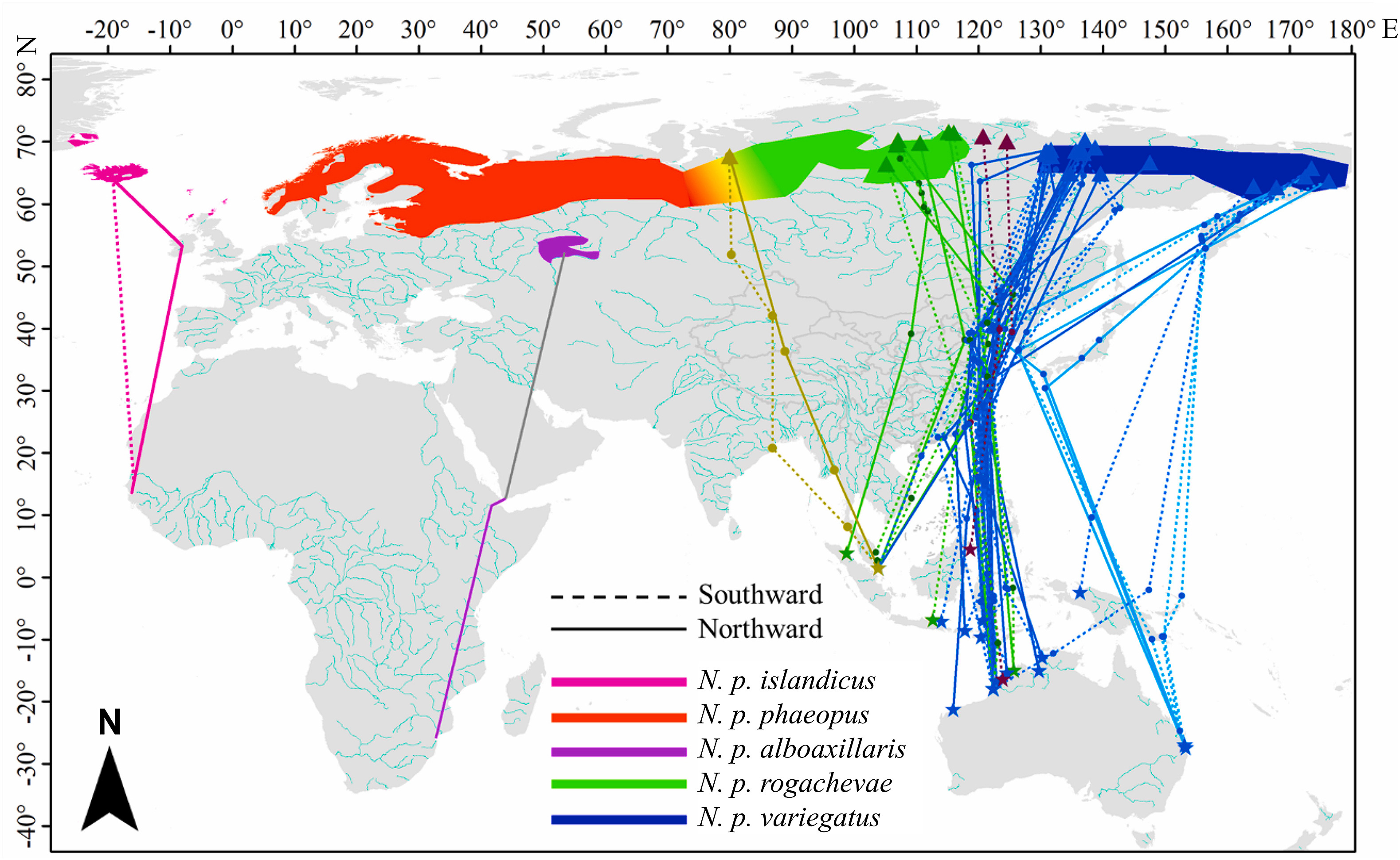
Determining the migration routes and connections of migratory birds at the population level helps clarify intraspecific differences in migration. Five subspecies have been recognized in the Whimbrel (Numenius phaeopus) in Eurasia. Ssp. rogachevae is the most recently described subspecies. It breeds in Central Siberia, while its non-breeding region and migration routes are still unclear. We tracked the migration of Eurasian Whimbrels captured at three non-breeding sites (Moreton Bay in east coast of Australia, Roebuck Bay in Northwest Australia and Sungei Buloh Wetland in Singapore) and two migration stopover sites (Chongming Dongtan and Mai Po Wetland in China). We determined the breeding sites and inferred the subspecies of the tagged birds in the East Asian – Australasian Flyway (EAAF) based on the known breeding distribution of each subspecies. Of the 30 tagged birds, 6 and 21 birds bred in the breeding range of ssp. rogachevae and variegatus, respectively; one bred in the presumed transition area between the breeding range of ssp. phaeopus and rogachevae, and two bred in the region between the breeding range of ssp. rogachevae and variegatus. The birds that bred in the ssp. rogachevae breeding range spent their non-breeding season in the northern Sumatra, Singapore, East Java and Northwest Australia and mainly stopped over along China's coasts during migration. None of our birds bred in the exclusive breeding range of the phaeopus subspecies. Previous studies have predicted that rogachevae whimbrels migrate along the Central Asian Flyway and spend the non-breeding season in West India and East Africa. We found that at least some rogachevae whimbrels migrate along the EAAF and spend the non-breeding season in Southeast Asia and Australia. The ssp. phaeopus is at best sparsely distributed in the EAAF in the west region, or possibly does not occur at all.
Revealing bird migration patterns is crucial to understanding the factors limiting populations of long-distance migratory species. Migrations are considered the most dangerous part of the annual cycle and mortality at that time may exceed that of the breeding and wintering periods even six-fold (Klaassen et al. 2014). The differentiation of migration routes between populations is known to affect individuals' survival and, effectually, also whole population trends (Hewson et al. 2016). Therefore, it is justified to investigate migratory behaviour of selected, especially declining, populations of species, although they already seem sufficiently studied. One such extensively studied species is the Osprey (Pandion haliaetus; see Bierregaard et al. 2014 for summary of migration studies). It is a true cosmopolitan species, considered a long-distance migrant in most of the Northern Hemisphere (Martell et al. 2011; MacKrill 2017), but also a sedentary or partial-migratory species in lower latitudes (Monti et al. 2018a). A great body of studies have shown that Ospreys can migrate in broad-front. A study by Østnes et al. (2019) can serve as a good example of such a flexible migration pattern, where juveniles dispersed in different directions and crossed the Mediterranean Sea at known bottlenecks but also through open sea. On the other hand, depending on the geographical location of populations, Ospreys tend to show some general "highways" and minor paths (Fig. 1). Individuals from western Europe (United Kingdom, Germany, Norway and Sweden) were shown to most frequently use (but not exclusively) the western-most Mediterranean passage through the Iberian Peninsula (Alerstam et al. 2006; Klaassen et al. 2011; Mackrill 2017; Meyburg et al. 2018). This seems logical and, consequently, the Ospreys from eastern Europe (Russia, Baltic countries and some from Finland) passed the Mediterranean Sea along its eastern shore or the Balkan Peninsula (Väli and Sellis 2015; Babushkin et al. 2019; LUOMUS 2020). The central Mediterranean passage through Corsica and Italian Peninsula were used by the Ospreys from central (e.g. Germany) and both, eastern and western Europe, but to a lesser degree. Finally, crossing the sea at its full width without any insurance of peninsulas and islands is also a common phenomenon in the case of this species. However, taking into account that migratory behaviour of soaring raptors, such as the Osprey, is most likely governed by innate mechanism (Väli et al. 2018), resembling the one called "clock and compass" known from passerines (Helbig 1996), we can expect that migration direction differs between populations. This assumption is supported by the philopatry phenomenon known to occur in Ospreys (Monti et al. 2018b). In this context, it might be interesting to investigate migration routes of populations in the middle of the range to check if the migration direction exhibits a continuous range or a migratory divide between the eastern and western populations of the species.
Figure
1.
Main migration paths of Ospreys from northern Europe and predicted migratory division (dashed line) between Osprey populations in eastern (red) and western (blue) Poland. Thin and thick lines show relative importance of migration paths and refer to studies by: (1) Mackrill 2017, (2) Østnes et al. 2019, (3) Alerstam et al. 2006, Klaassen et al. 2011, (4) Väli and Sellis 2015, (5) LUOMUS 2020, (6) Babushkin et al. 2019, (7) Meyburg et al. 2018
The central part of the Osprey's European range shows a break that is further prolonged by the Baltic Sea (Fig. 1). In the eastern part, species range is continuous, while in the western part, the range is fragmented but the population is increasing thanks to conservation efforts (Dennis 2016). However, between the continuous eastern and growing German population, there are small "islands" in this species' range, located in Poland. Both are in decline. Therefore, conservation actions and studies were recently carried out in order to protect this species in its last Polish populations (Anderwald 2017). We have carried out a GPS telemetry study to follow spatial ecology and mortality rates, but also to reveal if those two populations will show contrasting migratory patterns that might affect the more isolated and faster declining eastern population in particular. Taking into account the migration studies conducted so far (Fig. 1), we predict that the eastern and western populations may show a migratory divide and, therefore, might also be subjected to different selective pressure. Thus, our aim was to check whether migration paths and migration patterns differ between the eastern and western populations as well as between different age and sex classes. Also, we followed mortality rates and reasons to reveal the crucial threats for those declining populations and to check if potentially different migratory behaviour might affect it.
Methods
Study area and bird tagging
The study was carried out in two small, spatially distinct Polish populations, separated by about 235 km (between closest nests). The eastern population forms a small island (only 8–9 pairs in 2020) in the species range, about 200 km from the nearest breeding sites in Lithuania. It is located in the vast Masuria Lakeland area, where Ospreys mostly nest in the interior of large forest complexes. The western population contains about 16 pairs (in 2020) scattered in forests, but some also nesting on electric pylons on farmland. This population is slowly increasing and some exchange of individuals with Ospreys from eastern Germany was observed to occur in both directions.
Tagging was carried out in 2017–2020 and covered both chicks in nests and adults. In the first year, only a single juvenile was equipped with a 30 g "Ornitrack E30" GPS GPRS logger (manufactured by Ornitela Lithuania), while later on 4 individuals were tagged annually in 2018 and 2020 and 10 individuals in 2019, all with 35–40 g "Kite-M" GPS GPRS loggers (manufactured by Ecotone Poland). Well-developed chicks (body mass > 1420 g) were caught in the nests. Adults were caught with large mist nets, provoked by a stuffed white-tailed eagle close to their nests. Devices were mounted on the birds' backs with Teflon ribbon sewn at the sternum with the "Y" method described by Buehler et al. (1995). Data were acquired with a 15-min interval, on average. Sex was identified according to body mass and biometrics (Poole 1982), although in the case of juveniles, they were treated as possible male and possible female as biometry measurements can sometimes overlap between sexes. In the case of adults, direct observations of behaviour at the nesting site and the extent of the breeding patch were found to be reliable criteria in identifying sex.
Analysis
The onset of autumn migration was determined as a long (over 30 km), directional (straight-line) movement in the southern or south-western direction undertaken over at least two consecutive days. A threshold of a daily distance of 30 km was chosen upon a histogram of daily distance covered during the expected time of migration (August–November). Distance up to 30 km formed a visibly more frequent class, while distances over 30 km were represented with a similar frequency range (Additional file 1: Fig. S1). A stopover site was defined as an area where a bird spent more than two consecutive nights in a radius of less than 10 km. The end of autumn migration and beginning of wintering was considered to be the last day of the long, directional movement southwards. Migration direction was measured (in QGIS) as azimuth between location at start of migration and reaching the Mediterranean Sea. At the sea-crossing point, the innate migratory direction might already be distorted by choosing the most convenient site and trying to bypass this barrier, and therefore we measured it at the early migration stage. For mortality statistics, we counted followed individual as perished, if we were able to find its dead corpse. We reported also suspicious losses of data transmission as possible mortality events, but we have not included those cases as mortality events, since tag malfunction is also a common scenario. Total and daily distance covered by the followed individuals were calculated using the moveHMM package (Michelot et al. 2016) in R 4.0.3 and two-point equidistant projection. Straight-line distance was calculated with the same projection in QGIS 3.16. In order to check if sex, age and population had any impact on movement speed, they were used as fixed factors in linear mixed effects models with random effect of individual built into the lme4 package (Bates et al. 2015) to explain the logarithm of daily distance covered during migration. Stopover days were excluded by limiting the dataset to days with movement over 50 km. Models were built in additive pattern. The best supported model was chosen according to the Akaike Information Criterion (AIC). Models with ΔAIC < 2 were averaged in the MuMIn package (Bartoń 2020). In order to check whether migration patterns differed between individuals from the two distinct populations or individuals of different sex and age, departure date, migration duration, migration direction, number of stopover days and distance covered on migration were compared in above-mentioned pairs using the Mann–Whitney test.
Results
Migration routes
Out of 19 Ospreys tagged with GPS tags, we recorded the autumn migration of 12:5 adults (1 for 2 seasons) and 7 juveniles (Table 1). The remaining 7 died before the onset of migration, juveniles mostly due to Goshawk (Accipiter gentilis) predation or fell due to unknown reasons still at the nest or in its vicinity. Two adult males died due to poaching and electrocution at the breeding grounds. Unfortunately, 3 of migrating individuals recorded incomplete tracks, but came far enough to judge their general migration paths (Fig. 2a). The Ospreys showed broad-front migration and crossed the Mediterranean Sea in a vast belt from Gibraltar to the Balkan Peninsula to winter in the central part of Western Africa (Fig. 2a). The only exception was the juvenile that wintered in the Iberian Peninsula (Table 1). Some juveniles travelled long distances over open sea, while adults tended to use less hazardous paths along the eastern shore of the Iberian Peninsula, Corsica and Italian Peninsula (Fig. 2b). Ospreys from the eastern and western populations in Poland exhibited a partial migratory divide, with most of the individuals (86%; incl. all juveniles) from the eastern population heading south (azimuth 205.5 ± 16.8) and crossing the Mediterranean Sea at the middle to reach wintering sites in Ghana, Burkina Faso, Nigeria, Angola. On the contrary, most of the individuals (83%) from the western population headed south-west (azimuth 221.0 ± 20.3) to cross the Mediterranean Sea at its western narrowing and winter in Senegal, Sierra Leone and Spain (Fig. 2c). Differences in azimuth were, however, not significant (U = 12, p = 0.23). At later stages, migration direction was similar between the eastern and western population.
Table
1.
Characteristics of autumn migration of 12 GPS-followed Ospreys from Poland
ID
Year
Sex
Pop.
Age
Tag. date
Depart
Arrival
Stop-over days
Total duration (days)
Dist. (km)
Straight dist. (km)
Wintering destination
Mort.
Complete migration tracks
17988
2017
F
West
juv
06.07
18.09
01.10
0
13
4381
3940
Nigeria
Elec.
LPP11
2020
F
East
juv
03.07
11.09
29.09
0
18
5135
4628
Niger
LPP14
2020
M
West
ad
10.07
10.09
12.10
9
32
6069
5013
Senegal
Elec.
LPP15
2019
M
East
ad
22.06
27.08
11.10
18
45
6855
4921
Burkina Faso
Unkn.
LPP01a
2018
F
West
juv
03.07
01.09
24.09
1
23
4242
3899
Senegal
LPP02
2019
F
East
ad
15.06
14.09
13.10
1
29
6837
5237
Ghana
LPP05
2020
F
East
juv
26.06
24.08
30.10
35
68
9066
6724
Angola
LPP09a
2019
F
West
juv
22.07
21.08
02.11
47
73
3971
2512
Spain
Pred.
LPP09b
2020
F
West
ad
10.07
01.09
23.09
0
22
6468
5504
Sierra Leone
Incomplete migration tracks
LPP1b
2019
M
East
ad
10.08
8.09
4193
3731
Lost in Algeria
?
LPP04
2018
M
West
juv
03.07
31.08
2972
2623
Lost in Morocco
?
LPP10
2019
F
East
juv
23.06
21.08
2229
1430
Lost in Italy
?
LPP02
2020
F
East
ad
2019
16.09
2905
2253
Lost in Tunisia
?
Column "Mort." reports mortality reasons on migration; elec.: electrocution; pred.: predation; unkn.: reason unknown; ?: signal lost, bird status unknown
We observed high mortality in juveniles in their first year, reaching 67% in the case of certain deaths, but possibly even 83% together if unknown cases of suspicious signal losses were added. Some cases even occurred before the onset of migration (Table 2). Five died at fledgling stage: one in the nest of unknown reasons, three were killed by a Goshawk and another one was killed by a wind turbine. Two juveniles died at wintering sites: one was poached in Nigeria, the other died of unknown reasons in Senegal. The last death was confirmed at the start of spring migration in Spain, probably due to Goshawk predation. Another two juveniles stopped transmitting during autumn migration: one in Italy, the other in Morocco, but we have no evidence of their death. Only two juveniles certainly survived the first year and transmitted in the next season.
Table
2.
Mortality numbers of 19 GPS-tagged Ospreys from Poland in 2017–2020
Surprisingly, the mortality of adults was also high and reached 57% (confirmed deaths) already in the first year of tracking. In this case, the reasons were highly anthropogenic in three (out of four) cases: two adults died because of electrocution (one in Senegal and one in Poland) and one was poached (still in Poland). Another individual (from the eastern population) was simply found dead (for unknown reasons) in Burkina Faso. Two other adults from eastern population stopped transmitting in the deserts and were not seen at their breeding grounds in the next year, therefore actual mortality was probably even higher.
Factors affecting autumn migration
Models explaining daily distance covered on migration were poorly distinguishable between the base model with a random effect of individual only, comparing to models involving individual's sex, age and source population (Table 3). Eventually, the averaged model involved all the factors, but all were insignificant and showed very low slope estimates (Table 4). The variance explained by the base model was low (conditional R2 = 0.151) and barely increased when other predictors were added (conditional R2 increased to 0.165–0.167, depending on the model).
Table
3.
Components of linear mixed effects models of daily distance covered by GPS-tagged Ospreys from Poland on autumn migration in 2018–2020
No
Model components
df
logLik
AIC
ΔAIC
Weight
1
Random effect of individual (1|ID)
3
−261.78
529.54
0
0.62
2
Age+(1|ID)
4
−261.71
531.42
1.88
0.13
3
Sex+(1|ID)
4
−261.76
531.52
1.98
0.13
4
Population+(1|ID)
4
−261.76
531.53
1.99
0.12
5
Age+Sex+Population+(1|ID)
6
−261.74
535.28
5.74
–
Akaike Information Criterion (AIC) showed models No. 1–4 were barely distinguishable, therefore were averaged and their contribution to the best supported model is given in the "weight" field
The departure for autumn migration did not differ significantly between individuals of different age or sex, although in the last case, surprisingly, females left breeding grounds later than males (Additional file 1: Fig. S2). Similarly, we found no effect of age or sex on migration distance, duration and number of stopover days (Fig. 3a, c, d; Additional file 1: Table S1). However, when individuals from different source populations were compared, the distance covered on migration was almost significantly higher in the eastern than in the western population (Fig. 3b) although straight-line distance was almost the same (U = 15, p = 0.286). When migration was divided into sections, juveniles covered some surplus kilometres across land in Europe, compared to adults (Fig. 4a). Individuals from the eastern population covered shorter distances over land in Europe, but longer over sea and the Sahara Desert (Fig. 4b). Unfortunately, the sample size was too small to perform reliable statistical comparisons.
Figure
3.
Departure date (a), total distance (b), duration of migration (c) and number of stopover days (d) shown by GPS-tagged Ospreys from eastern (red) and western (blue) Poland on their autumn migration. p value of the Mann–Whitney test between populations is given for close-to-significant differences
Figure
4.
Distance covered on autumn migration by GPS-tagged Ospreys from Poland, depending on migration stage and divided by individual age (a) and source population (b). Yellow boxes represent juveniles, green: adults, red: eastern population, blue: western population
In general, the dynamics of migration was rather even in adults, but showed longer stopovers and more pronounced stopover behaviour in some juveniles (Additional file 1: Fig. S3). Individuals of both age groups were capable of covering a distance of around 800 km in 24 h.
Discussion
Overall, the Ospreys from the two distinct populations differed in their migration routes. With single exceptions, individuals from the eastern and western populations headed south and south-west, respectively. Eastern juveniles, which are supposed to follow their innate migratory mechanism at first migration (Väli et al. 2018), tended to cross the Mediterranean Sea further east than adults, highlighting the difference in direction between eastern and western population. Taking into account that migratory direction is inherited, single exceptions from the main flyways may show that the two populations are not entirely isolated. Osprey males are highly philopatric and breed in the radius of about 30 km from their natal sites (Kinkead 1985). On the contrary, females can show quite long dispersal and may breed hundreds of kilometres from their birth site. In the mentioned population of the Osprey in eastern Poland, alien females (but not males), ringed in Germany and Latvia (> 500 km), were recorded to breed. Thus, gene flow with populations of different migration paths certainly occurs, although with low frequency, as shown for other European populations by Monti et al. (2018b). The authors of the above-mentioned paper reveal that gene flow and migratory behaviour are linked in Europe and the central location of Polish populations might attract dispersing females from three different migratory pathways. However, looking at the rather distinct migratory pathways of Ospreys belonging to the western and central populations studied, gene flow rates are rather low. Preliminary genetic analysis done on Polish Ospreys showed relatively high heterozygosity in the small sample set dominated by the western population. All individuals from this population represented one haplotype (Rutkowski 2019), common for Ospreys in northern and western Europe (Monti et al. 2018b). On the other hand, unique alleles were found in a single individual examined from the eastern population (Rutkowski 2019). This fact may indicate relatively high genetic distance between the two mentioned populations, but needs further confirmation in a greater sample size.
Migration patterns, at least for tested traits, did not differ between age and sex classes. This was quite surprising, since many studies have shown females depart much earlier than males (Bai and Schmidt 2012; Väli and Sellis 2015; Meyburg et al. 2018), adult females and juveniles cover longer distances than adult males (Bai and Schmidt 2012), and males use fewer days for stopovers than females (Alerstam et al. 2006; Meyburg et al. 2018). In terms of the departure date, the difference should be sharp, but not only was this not observed in adult females, but we found them departing later than adult males. This might be an artefact caused by the low sample size, but also it may indicate that foraging conditions were optimal and inter-species competition is low in those small populations. Therefore, instead of moving to other foraging sites, the breeding grounds were suitable for "filling their tanks" before migration. In contrary to other mentioned studies, we did not find age or sex to affect migration dynamics, but also showed that it is variable, especially in the case of juveniles (Additional file 1: Fig. S3).
Individuals from different source populations, although located along similar latitudes, exhibited close-to-significant differences in migration distance (Fig. 3b), with individuals from the eastern population covering, on average, over 2000 km more to reach wintering sites. When migration was divided into stages, we found that the main difference in migration length lies in the most risky parts: crossing the Mediterranean Sea and Sahara Desert (Fig. 4b). This fact may be a factor in differing survival rates and the opposite population trend patterns in the two populations across the migratory divide. Such a pattern was shown for the Common Cuckoo (Cuculus canorus) in Great Britain, which uses two different paths on its way to wintering sites in Africa (Hewson et al. 2016). Those that followed the central Mediterranean passage and later headed through the central part of the Sahara Desert exhibited higher mortality and, in consequence, declined.
We found mortality rates in Polish Ospreys to be high in juveniles, which was expected, but also high in adults. It seems odd that mature and experienced Ospreys in our study do not demonstrate a significant increase in survival. However, other studies support the expected increase in survival with age (i.e. Wahl and Barbraud 2014; Väli et al. 2021). Monti et al. (2014) showed survival of translocated juveniles reaching only 26% in their first autumn/winter season, while in the second calendar year it reached 69%, and later increased in adults to 93%. In France, the survival of individuals reached 49% in their first year and as much as 87% in the second year and in older birds (Wahl and Barbraud 2014). Even higher rates were noted in Swedish Ospreys; 65% of year-old birds survived the first year and 81% thereafter (Ryttman 1994). In Latvia and Estonia, survival of 1-year old birds was reached only 14% for young females and 27% for young males, while in older individuals it reached 64% and 74%, respectively (Väli et al. 2021). In North America, survival in the first year was estimated to be as much as 47%, while in adults to ca. 81% (Henny and Wight 1969). All of these data reported above were based on ringing recoveries and were much higher than the ones reported in our telemetry study. Certainly, GPS devices are more precise in showing where and when mortality exactly occurred and probably less biased than survival estimates from capture-recapture methods. At the same time GPS tags themselves were not found to affect survival of followed individuals (Väli et al. 2021).
In the case of adults, a huge part of mortality was attributed to anthropogenic reasons. We have noted surprisingly small mortality on migration, contrary to what is expected, given the results of Klaassen et al. (2014) on a few migratory birds of prey species, including the Osprey. However, probably also in our case, the mortality on migration is higher and at least some individuals that stopped transmitting on migration, actually perished. Birds' corpses were not found in those cases, which is not surprising especially in the deserts. However, considering the early death of unknown reason of individual just reaching the wintering grounds and two individuals missing while flying through deserts, eastern population seems more susceptible to losses on migration. This corresponds to greater distance they covered, especially at the most hazardous points on the migration route (sea and desert; Fig. 4b). Greater distance covered during migration is associated with a higher risk of mortality in Ospreys, but also in other migratory birds of prey (de Pascalis et al. 2020). The mortality reasons are not necessarily directly linked to longer and more hazardous paths, but may impact the birds' condition by weakening them and making them more susceptible to predation, poaching and starvation, i.e. "disappearing" in the deserts or dying of unknown causes. Certainly, the high mortality of Ospreys from declining Polish population requires further investigation to reveal the most important threats and subsequently counteract them.
Conclusions
Two small and distinct Osprey populations showed a clear but incomplete migratory divide, despite being separated by only a few hundred kilometres, a distance that did not completely limit gene flow through female dispersal. We found high mortality rates in juvenile and adult Ospreys, which were relatively higher while on migration in the case of the eastern population. There may be a link between migration route and mortality rates that led to the steeper decline of the eastern population; however, the sample size of tagged individuals is too low at the moment to make any definite conclusions.
Additional file 1: Figure S1. Histogram of daily distance covered on migration by GPS-tracked Ospreys from Poland in 2017–2020. Figure S2. Differences in departure dates between age and sex classes in GPS-tracked Ospreys from Poland on their autumn migration. Figure S3. Distance covered daily on migration by juvenile (a) and adult (b) Ospreys from Poland followed with GPS loggers in 2017–2020. Table S1. Results of Mann–Whitney test comparing autumn migration traits between individuals of different sex, age and source population (eastern vs. western) among nine Ospreys from Poland followed with GPS telemetry in 2017–2020.
Acknowledgements
We would like to thank Michał Dzwonkowski, Dariusz Kujawa, Cezary Korkosz, Tomasz Kryjan, Stefan Lewandowski, Marcin Sołowiej, Piotr Waszczykowski and Paweł Czechowski for their help in the fieldworks conducted.
Authors' contributions
DA designed the study and obtained funding. PM led data analysis and manuscript writing. DA, MZ, ŁC, SR participated in data analysis and writing. All co-authors were involved in the fieldwork. All authors read and approved the final manuscript.
Availability of data and materials
The dataset supporting the conclusions of this article is stored in the Movebank repository, ID 365184781 and available on request.
Declarations
Ethics approval and consent to participate
Catching and tagging of Ospreys were carried out under the license of General Inspectorate for Environmental Protection—DZP.WG.6401.03.109.2017.dł.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Allport, G.A., Atkinson, P.W., Carvalho, M., Clark, N.A., Green, R.E., 2018. Local site use and frst northbound migration track of non-breeding Steppe Whimbrel Numenius phaeopus alboaxillaris (Lowe 1921). Wader Study 125, 219–227
Arlott, N., 2018. Birds of the Philippines, Sumatra, Java, Bali, Borneo, Sulawesi, the Lesser Sundas and the Moluccas. William Collins, London.
Baker, R.H., 1951. The Avifauna of Micronesia, Its Origin, Evolution, and Distribution. University of Kansas, Lawrence.
BirdLife International, NatureServe, 2015. Bird Species Distribution Maps of the World. BirdLife International and Arlington: NatureServe, Cambridge.
Bishop KD. Shorebirds in New Guinea: their status, conservation and distribution. Stilt. 2006;50:103-134
Brown, D., Crockford, N., Sheldon, R., 2014. Drivers of population change and conservation priorities for the Numeniini populations of the world. UNEP/CMS/COP11/Inf.33. (03 Oct 2018).
Carneiro C, Gunnarsson TG, Alves JA. Faster migration in autumn than in spring: seasonal migration patterns and non-breeding distribution of Icelandic whimbrels Numenius phaeopus islandicus. J Avian Biol. 2019;e01938
Cramp SC. Handbook of the birds of Europe, the Middle East and North Africa. The birds of the Western Palearctic. Vol.3. Waders to gulls. Oxford: Oxford university press; 1983
Crossland AC, Butcher S, Crutchley P, Mugan ND, Kake J. A survey of waders on part of Guadalcanal and the Florida group, Solomon Islands. Stilt. 2016;69-70:48-56
Deignan HG. Checklist of the birds of Thailand. Washington, Smithsonian Institution;1963
Deignan HG. Corrections and additions to the published records of Siamese birds. Auk. 1946;63:243-245
del Hoyo J, Collar NJ. HBW and Birdlife International illustrated checklist of the birds of the world. Vol.1. Non-passerines. Barcelona: Lynx Edicions; 2014
Gill BJ, Bell BD, Chambers GK, Medway DG, Palma RL, Scofield RP, et al. Checklist of the birds of New Zealand, Norfolk and Macquarie Islands, and the Ross Dependency, Antarctica. 4th ed. Wellington: Te Papa Press; 2010
Gill F, Donsker D, Rasmussen P. IOC world bird list (v. 11.1). 2021.
Grantham MJ. A note on the passage of variegatus and phaeopus type whimbrels through Alas Purwo National Park, East Java. Kukila. 2000;11:133-135
Hayman, P., Marchant, J., Prater, T., 1986. Shorebirds: An Identification Guide to the Waders of the World. Croom Helm, London
Iqbal M, Ridwan A. Summering of whimbrel in southern Sumatra, Indonesia. Stilt. 2009;55:20-24
Jeyarajasingam A. A field guide to the birds of Peninsular Malaysia and Singapore. Oxford: Oxford University Press; 2012
Jorgensen AA. Siams Vadefugle. III. Charadriiae (Tringinae). Dansk Orn For Tidsskr. 1949;43:216-237
Kuang FL, Coleman JT, Hassell CJ, Leung KSK, Maglio G, Ke WJ, et al. Seasonal and population differences in migration of whimbrels in the East Asian - Australasian Flyway. Avian Res. 2020;11:24
Lappo EG, Tomkovich PP, Syroechkovski EE. Atlas of breeding waders in the Russian Arctic. Moscow: UF Ofsetnaya Pechat; 2012
Lekagul, B., Round, P. D., 1991. A Guide to the Birds of Thailand. Saha Karn Bhaet, Bangkok
Li D, Davison G, Lisovski S, Battley PF, Ma ZJ, Yang SF, et al. Shorebirds wintering in Southeast Asia demonstrate trans-Himalayan flights. Sci Rep. 2020;10:21232
MacKinnon J, Phillipps K. A field guide to the birds of Borneo, Sumatra, Java, and Bali (The Greater Sunda Islands). Oxford: Oxford university press; 1993
Newton I. The migration ecology of birds. London: Academic Press; 2008
Phillips WWA. A 1975 annotated checklist of the birds of Ceylon (Sri Lanka). Colombo: Wildlife and Nature Protection Society of Ceylon; 1975
Prater AJ, Marchant JH, Vuorinen J. Guide to the identification and ageing of Holarctic waders. Tring: BTO; 1977
Ripley SD. A synopsis of the birds of India and Pakistan: together with those of Nepal, Bhutan, Bangladesh, and Sri Lanka. Bombay: Bombay natural history society; 1982
Robinson HT, Chasen FN. The birds of the Malay Peninsula, Vol. 3: sporting birds, birds of the shore and estuaries. London: Witherby; 1936
Robson C. New Holland field guide to birds of South-East Asia: Thailand, Peninsular Malaysia, Singapore, Vietnam, Cambodia, Laos, Myanmar. London: New Holland; 2005
10.2173/bow.whimbr.01
Stinson DW, Wiles GJ, Reichel JD. Occurrence of migrant shorebirds in the Mariana Islands. J Field Ornitol. 1997;68:42-55
Sweet SK, Asmus A, Rich ME, Wingfield J, Gough L, Boelman NT. NDVI as a predictor of canopy arthropod biomass in the Alaskan arctic tundra. Ecol Appl. 2015;25:779-790
Tan HZ, Ng EYX, Tang Q, Allport GA, Jansen JJFJ, Tomkovich PS, et al. Population genomics of two congeneric Palaearctic shorebirds reveals differential impacts of Quaternary climate oscillations across habitats types. Sci Rep. 2019;9:18172
Tomkovich PS. A new subspecies of the whimbrel (Numenius phaeopus) from central Siberia. Zool Zurnal. 2008;87:1092-1099
Varlamova EV, Solovyev VS. Study of NDVI variations in tundra and taiga areas of Eastern Siberia (Yakutia). Atmos Ocean Opt. 2015;28:64-67
Wells DR. The birds of the Thai-Malay Peninsula, Vol. 1: non-passerines. London: Academic Press; 1999
Alberto Fuentes-López, María Teresa Rebelo, Elena Romera, et al. Genetic diversity of Calliphora vicina (Diptera: Calliphoridae) in the Iberian Peninsula based on cox1, 16S and ITS2 sequences. Biological Journal of the Linnean Society, 2020.
DOI:10.1093/biolinnean/blaa109
2.
Youhua CHEN. Does the diversification rate of endemic birds of mainland China follow abrupt, gradual shifting or constant patterns?. Integrative Zoology, 2017, 12(2): 165.
DOI:10.1111/1749-4877.12205
Table
3.
Components of linear mixed effects models of daily distance covered by GPS-tagged Ospreys from Poland on autumn migration in 2018–2020
No
Model components
df
logLik
AIC
ΔAIC
Weight
1
Random effect of individual (1|ID)
3
−261.78
529.54
0
0.62
2
Age+(1|ID)
4
−261.71
531.42
1.88
0.13
3
Sex+(1|ID)
4
−261.76
531.52
1.98
0.13
4
Population+(1|ID)
4
−261.76
531.53
1.99
0.12
5
Age+Sex+Population+(1|ID)
6
−261.74
535.28
5.74
–
Akaike Information Criterion (AIC) showed models No. 1–4 were barely distinguishable, therefore were averaged and their contribution to the best supported model is given in the "weight" field


 DownLoad:
DownLoad:








 Email Alerts
Email Alerts RSS Feeds
RSS Feeds