Per ALSTRÖM, Urban OLSSON, Fumin LEI. 2013: A review of the recent advances in the systematics of the avian superfamily Sylvioidea. Avian Research, 4(2): 99-131. DOI: 10.5122/cbirds.2013.0016
Citation:
Per ALSTRÖM, Urban OLSSON, Fumin LEI. 2013: A review of the recent advances in the systematics of the avian superfamily Sylvioidea. Avian Research, 4(2): 99-131. DOI: 10.5122/cbirds.2013.0016
Per ALSTRÖM, Urban OLSSON, Fumin LEI. 2013: A review of the recent advances in the systematics of the avian superfamily Sylvioidea. Avian Research, 4(2): 99-131. DOI: 10.5122/cbirds.2013.0016
Citation:
Per ALSTRÖM, Urban OLSSON, Fumin LEI. 2013: A review of the recent advances in the systematics of the avian superfamily Sylvioidea. Avian Research, 4(2): 99-131. DOI: 10.5122/cbirds.2013.0016
The systematics of the avian superfamily Sylvioidea are reviewed, focusing on studies of relationships among families and within genera, more superficially on taxonomic studies at the species level. For the families Bernieridae and Phylloscopidae, new analyses based on already published sequence data are presented. Our understanding of relationships has been vastly improved in recent years due to a large number of molecular studies. However, the relationships among the different families remain largely obscured, probably mainly as a result of rapid divergence of the different primary lineages (families). Also, species level taxonomy has been much improved in recent years due to a large number of studies applying molecular markers and/or vocalizations and other life-history data. It seems likely that the number of species will continue to increase, as new groups are being studied with modern integrative methods.
Figure
1.
Phylogeny of superfamily Sylvioidea based on concatenated sequences of one mitochondrial (cytochrome b) and six nuclear loci (FGB, GAPDH, LDHB, MB, ODC1, RAG1; in total ~6.3 kbp), analyzed by Bayesian inference (partitioned by locus). Support values are given in the order posterior probability (PP) / maximum likelihood (ML) bootstrap; ** indicates posterior probability 1.00 or ML bootstrap 100%; – indicates no ML bootstrap support for this node, but clade recovered in ML search for best topology. Homoplasy-free insertions (+) and deletions (–) are indicated. From Fregin et al. (2012), two long branches have been shortened (broken). The family names are those recommended by Fregin et al. (2012); a few of the generic names have been changed to reflect more recent taxonomic recommendations (e.g. Alström et al., 2011b; Moyle et al., 2012).
The superfamily Sylvioidea has received much attention in recent years, with three major studies of interfamilial relationships (Alström et al., 2006; Johansson et al., 2008a; Fregin et al., 2012) and multiple studies focusing on different families within it (e.g. Cibois et al., 1999, 2001; Helbig and Seibold, 1999; Sheldon et al., 2005; Moyle and Marks, 2006; Nguembock et al., 2007; Fregin et al., 2009; Gelang et al., 2009; Alström et al., 2011a, b, 2013; Moyle et al., 2012; Olsson et al., 2013a). The first comprehensive study of the whole superfamily (Alström et al., 2006) was based on one nuclear and one mitochondrial sequence. That study identified 10 well-supported major clades, which were proposed to be recognized at family level (Acrocephalidae, Aegithalidae, Alaudidae, Cettiidae, Cisticolidae, Hirundinidae, Megaluridae, Phylloscopidae, Pycnonotidae and Timaliidae), as well as three lineages comprising one to two species each that were not recognized nomenclaturally (Donacobius, Melocichla + Sylvietta and Panurus). Johansson et al. (2008a) recognized also Bernieridae, of which Alström et al (2006) had no representatives, and referred to the Melocichla + Sylvietta clade as the "Sphenoeacus group" (comprising Macrosphenus,
Sphenoeacus, Bradypterus victorini and two species of Sylvietta). Both these groupings were previously identified by Beresford et al. (2005), the former as "Malagasy endemic 'warblers'" and the latter as the "Sphenoeacus group" (containing also Achaetops, in addition to the previously mentioned genera).
The finding that the type genus of Sylviidae, Sylvia, was nested within the large Timaliidae group (as previously discovered by Fjeldså et al., 1999; Cibois, 2003a, b) caused a taxonomic problem, as the older name Sylviidae has priority over Timaliidae (ICZN, 1999), and it would be inconvenient both to discard the name Timaliidae and to use the name Sylviidae in a completely new context. To solve this difficulty, it was suggested that Sylviidae be suppressed, and Timaliidae used for the group including mostly traditional Timaliidae species (Cibois, 2003a, b; Alström et al., 2006). However, Gelang et al. (2009), again based on DNA sequence data, resurrected Sylviidae, but restricted it to a clade containing mostly traditional Timaliidae species, and this arrangement has been followed by subsequent authors (Fregin et al., 2012; Moyle et al., 2012; Gill and Donsker, 2013).
The latest and most comprehensive analysis of Sylvioidea (Fregin et al., 2012) proposed new family names for the genera Scotocerca and Erythrocercus, the Scotocercidae (a monotypic family) and the Erythrocercidae (with three species), respectively, and formally proposed the name Macrosphenidae for the "Sphenoeacus group". They also tentatively recommended the use of Hyliidae for Hylia and Pholidornis, and argued against inclusion of Paridae, Remizidae and Stenostiridae in Sylvioidea. In total, their classification comprised 23 families in Sylvioidea (Fig. 1), containing over 1200 species in more than 220 genera (Gill and Donsker, 2013).
Sylvioidea is essentially an Old World radiation. Most of the families are either absent or represented by just a few species in the New World. A striking exception concerns the family Hirundinidae, which is well represented on all continents (excluding Antarctica), likely the result of the extraordinary flight capability of its members.
The present review focuses on studies of relationships among the families within Sylvioidea, as well as on relationships within genera, but only cursorily deals with taxonomic studies at the species level. For the families Bernieridae and Phylloscopidae, new analyses based on previously published sequence data are presented. Recent advances in the species-level taxonomy of Indian subcontinent birds have been reviewed by Rasmussen (2012), which see for additional information.
Relationships among families
In the study by Alström et al. (2006), based on two loci, one mitochondrial and one nuclear, most of the deep internal nodes were unresolved. However, three of the relationships among the 10 clades they proposed to be recognized at the family level had high posterior probability (0.95–1.00): the Megaluridae + Donacobius + Acrocephalidae clade, the Phylloscopidae + Aegithalidae + Cettiidae + Hirundinidae + Pycnonotidae clade, and the sister relationship between Alaudidae + Panurus and the other Sylvioidea. However, none of these clades obtained >50% parsimony bootstrap support, so they were not considered reliable. Johansson et al.
(2008a), utilizing three nuclear introns (including the one used by Alström et al., 2006), found strong support for a Megaluridae + Donacobius + Bernieridae clade, as well as for the sister relationships between these and Acrocephalidae. They also found strong support for the position of the Alaudidae + Panurus clade and Nicator outside of the other Sylvioidea, but no other strongly supported relationships among families.
The analysis of six nuclear and one mitochondrial loci by Fregin et al. (2012; Fig. 1) corroborated the Locustellidae (formerly Megaluridae; see below) + Donacobiidae (treated at family rank; see below) + Bernieridae clade, but found no statistical support for a close relationship between these and Acrocephalidae. They also found the Alaudidae + Panuridae clade and Nicatoridae (all treated at family level; see below) in a polytomy with the rest of Sylvioidea (the Sylvioidea clade excluding Alaudidae, Panuridae and Nicatoridae receiving 1.00 posterior probability and 74% maximum likelihood bootstrap support). Moreover, the Cettiidae + Scotocercidae clade and the Cettiidae + Scotocercidae + Erythrocercidae + Aegithalidae + Phylloscopidae clade were strongly supported both in the Bayesian and maximum likelihood analyses. The Cettiidae + Scotocercidae grouping had previously been shown to be supported by morphological characters (Alström et al., 2011c). However, other relationships among families were generally unsupported, or strongly supported by Bayesian inference but with poor or moderate maximum likelihood bootstrap support. Most of the deep nodes with poor or conflicting support were very short, suggesting rapid divergence resulting in difficulties in reconstruction of relationships (e.g. Lewis et al., 2005; Rokas and Carroll, 2006).
With respect to the relationships among the five primary "babbler" clades, using partly different loci, Gelang et al. (2009), Fregin et al. (2012) and Moyle et al. (2012) recovered the same topology, i.e. ((((Leiotrichidae/-inae, Pellorneidae/-inae), Timaliidae/-inae), Zosteropidae/-inae), Sylviidae/-inae) (cf. Fig. 1). The latter study was the only one that obtained unanimously strong support for these relationships, whereas the overall weakest support was found in the study with the largest number of unlinked loci (Fregin et al., 2012). The comparatively poor support (especially for the Leiotrichidae/-inae + Pellorneidae/-inae clade) in the Fregin et al. (2012) study might have been the result of the small number of species in each clade compared to the other studies.
Relationships within families
Alaudidae
The Alaudidae (larks) are found on all continents, although by far the largest number of species are found in Africa, and only one species each is found in the New World and Australasia (de Juana et al., 2004). This is one of the least studied of the families within Sylvioidea. The circumscription of this family has not been disputed, and it seems unlikely that its inclusiveness will change. However, the relationships within and among the different genera are uncertain, and the number of genera and their composition have fluctuated dramatically over the years (e.g. Roberts, 1940; Meinertzhagen, 1951; Vaurie, 1951; Macdonald, 1952a, b, 1953; Verheyen, 1958; Peters, 1960a; Clancey, 1966, 1980; Harrison, 1966; Maclean, 1969; Wolters, 1979; Dean et al., 1992; Dickinson, 2003; Pätzold, 2003; de Juana et al., 2004). Certain genera, notably Mirafra, have acted as "dumping grounds", whereas several monotypic genera (e.g. Pseudalaemon, Lullula, Ramphocoris) with uncertain affinities have been erected.
Until recently, only one phylogeny of the Alaudidae, based on mitochondrial sequences from a small number of mostly African species, has been published (Tieleman et al., 2003). However, a phylogeny including 81 of the 97 species and representatives of all 21 recognized genera, based on two mitochondrial and three nuclear loci has just appeared (Alström et al., 2013). The latter has revealed many unpredicted relationships, including some non-monophyletic genera (Calandrella, Mirafra,
Melanocorypha, Spizocorys), and proposed a revised generic classification. For example, the genus Calandrella is shown to be separated into two non-sister clades, with C. brachydactyla, C. cinerea and C. acutirostris being closely related to Eremophila, whereas the other Calandrella species (e.g. C. rufescens and C. cheleensis) are more closely related to a clade containing the two monotypic genera Eremalauda and Chersophilus; the close affinity between Eremalauda and Chersophilus was also totally unexpected. Alström et al. (2013) proposed that the name Alaudala be used for the second of these traditional Calandrella clades. Another surprising result from Alström et al.'s (2013) multilocus analysis was that Melanocorypha leucoptera and M. mongolica are not closely related, hence it was proposed that the former be moved to the genus Alauda.
Lark taxonomy based on morphology has received much attention in Africa (Meinertzhagen, 1951; Winterbottom, 1957; Lawson, 1961; Clancey, 1989; Ryan and Bloomer, 1997; García et al., 2008) and Eurasia (Meinertzhagen, 1951; Vaurie, 1951, 1954; Dickinson and Dekker, 2001). Recent studies have revealed considerable hidden diversity in some taxa, such as Mirafra assamica, which was proposed to be split into four species based mainly on vocal, behavioral and morphological data (Alström, 1998; corroborated by molecular study by Alström et al., 2013); the Calendulauda (previously Certhilauda) albescens complex, which was suggested to consist of three instead of two species based on mitochondrial, morphological and vocal data (Ryan et al., 1998); the Certhilauda curvirostris complex, which was recommended to be treated as five instead of one species based on mitochondrial and morphological evidence (Ryan and Bloomer, 1999); and Galerida cristata, which was shown to consist of two partly sympatric and largely reproductively isolated species in Morocco, G. cristata sensu stricto and G. macrorhyncha (Guillaumet et al., 2005, 2006, 2008). In an analysis of mitochondrial data, Alström et al. (2013) found several unexpectedly deep divergences between taxa presently treated as conspecific (e.g. within Ammomanes cinctura, Ammomanes deserti, Calandrella brachydactyla, Eremophila alpestris), but also some shallow splits between currently recognized species (e.g. Certhilauda brevirostris–C. semitorquata–C. curvirostris [cf. Ryan and Bloomer, 1999];
Calendulauda barlowi–C. erythrochlamys [cf. Ryan et al., 1998]; Mirafra cantillans–M. javanica). Moreover, Zink et al. (2008) found a deep divergence in mitochondrial DNA between eastern and western populations of Alauda arvensis. It seems likely that the total number of recognized lark species is underestimated, and we predict that future studies based on molecular and/or vocal data will show that many of the currently recognized subspecies warrant recognition as separate species.
The realization that this exclusively African family exists as a distinct evolutionary lineage has gradually grown out of molecular evidence, which piece by piece brought together a morphologically diverse assemblage of species as a monophyletic group. Sefc et al. (2003) found that Sylvietta, which was placed in Acrocephalidae by Sibley and Monroe (1990), was not closely related to any other warbler taxa in their analysis of mitochondrial markers. Beresford et al. (2005) included Achaetops, Bradypterus victorini, Macrosphenus, Sphenoeacus and Sylvietta in their analysis of nuclear RAG1 and RAG2 sequences and identified a clade consisting of these species as the "Sphenoeacus group". The position of Bradypterus victorini, removed from other species in the genus, prompted a reversion to its older generic name Cryptillas. Fuchs et al. (2006) confirmed the close association of Sylvietta, Macrosphenus and Sphenoeacus based on one nuclear and one mitochondrial marker, and found that Melocichla was also a member of the "Sphenoeacus group"; Alström et al. (2006) also recovered a clade including Sylvietta and Melocichla. Johansson et al. (2008a) corroborated the existence of the "Sphenoeacus group" in their analysis of three nuclear loci by finding a clade consisting of Cryptillas
(Bradypterus) victorini, Macrosphenus, Sphenoeacus and Sylvietta, but they did not consider Achaetops or Melocichla. Fregin et al. (2012) used one mitochondrial and six nuclear markers and again found support for a clade consisting of Cryptillas victorini, Macrosphenus, Melocichla, Sphenoeacus and Sylvietta, but did not include Achaetops in their analysis. Fregin et al. (2012) erected the family Macrosphenidae for this clade, although it lacks known diagnostic morphological features. In total, 18 species in 6 genera are presently placed in this family (Gill and Donsker, 2013).
Nguembock et al. (2007, 2012), who analyzed 15 and 17 cisticolid genera, respectively, using one to three mitochondrial and one nuclear markers, and Olsson et al. (2013a; Fig. 2), who analyzed representatives of all genera believed to belong in Cisticolidae using two mitochondrial and two nuclear markers, are the most complete studies to date. These authors identified four main clades: (1) the speciose genus Apalis and several smaller (including monotypic) genera, all with exclusively African distributions; (2) the large genus Cisticola and several smaller (including monotypic) genera, of which all except two species of Cisticola are restricted to Africa; (3) the fairly large genera Prinia (Africa, Asia) and Orthotomus (Asia) and the two monotypic African genera Heliolais and Urorhipis; and (4) the genus Neomixis containing three species, all restricted to Madagascar. Olsson et al. (2013a) classified these clades as Eremomelinae, Cisticolinae, Priniinae and Neomixinae, respectively. A fifth lineage, containing the Philippine Micromacronus sordidus, which was shown by Oliveros et al. (2012) to be part of Cisticolidae, was in an incertae sedis position within Cisticolidae in the study by Olsson et al. (2013a).
Figure
2.
Family Cisticolidae based on concatenated nuclear MB and ODC and mitochondrial cytochrome b and ND2, based on Olsson et al. (2013). Posterior probabilities (above) and maximum likelihood bootstrap (below) values are indicated at the nodes; an asterisk represents posterior probability 1.00 or maximum likelihood bootstrap 100%. Changes of generic names are indicated in red (Heliolais erythroptera and Urorhipis rufifrons moved to genus Prinia).
Studies using molecular markers have found several of the cisticolid genera to be non-monophyletic, resulting in taxonomic revisions. The genus Apalis was shown by Nguembock et al. (2007) to be non-monophyletic, as Apalis ruwenzorii and A. pulchra were more closely related to Artisornis metopias than to other Apalis species, leading to erection of the genus Oreolais for the two former species (Nguembock et al., 2008). Moreover, the two African taxa currently placed in Artisornis have previously been placed in Orthotomus (e.g. Hall and Moreau, 1970; Watson et al., 1986), but this has been shown to be incorrect as A. metopias is not closely related to Orthotomus (Nguembock et al., 2007, 2008, 2012; Fregin et al., 2012; Olsson et al., 2013a). Recently,
Orthotomus cuculatus1 was moved from Orthotomus to Phyllergates, as it has been shown to be part of Cettiidae (Alström et al., 2006, 2011b; Fuchs et al., 2006; Nguembock et al., 2007, 2012). Olsson et al. (2013a) synonymized Urorhipis and Heliolais with Prinia. They also showed that Prinia burnesii is a babbler (Pellorneidae sensu Fregin et al., 2012), and reinstated the generic name Laticilla for this species.
1 Note that the frequently used spelling cucullatus is apparently incorrect (Gill and Donsker, 2013).
Sheldon et al. (2012) reconstructed the phylogeny of all Orthotomus species based on mitochondrial and nuclear DNA. They revealed e.g. a deep split and nonmonophyletic relationships within O. castaneiceps, and showed that several taxa previously considered subspecies of O. atrogularis were highly divergent.
Prinia fluviatilis was recognized as a species due to its distinctive song (Chappuis, 1974), and P. superciliaris was split from P. atrogularis on the basis of morphological and vocal evidence (Rasmussen and Anderton, 2005, 2012). Several studies underway suggest that the genus Prinia contains considerably more species than presently recognized, and this seems likely to be the case also in the speciose genus Cisticola.
Locustellidae
The family Locustellidae (grassbirds, bush warblers, grasshopper warblers and allies) is widely distributed across Africa, Eurasia and Australasia, with a total of 57 species in 9 genera (Gill and Donsker, 2013). It was first recognized by Sibley and Monroe (1990), partly based on DNA-DNA hybridization work by Sibley and Ahlquist (1990), as the subfamily Megalurinae, containing the genera Megalurus, Cincloramphus, Eremiornis,
Amphilais, Megalurulus, Buettikoferella, Chaetornis,
Graminicola and Schoenicola. Based on DNA sequence data from a broad selection of sylvioid genera, Alström et al. (2006) and Johansson et al. (2008a) recognized Megaluridae at the family level, comprising the genera Megalurus, Bradypterus, Locustella and Dromaeocercus. Hence, their circumscription included three of the genera placed in Acrocephalinae by Sibley and Monroe (1990). Moreover, molecular studies also revealed that the monotypic genus Graminicola is a babbler (Alström et al., 2006; Gelang et al., 2009; Olsson et al., 2013a), and that the aberrant Bradypterus victorini is not related to this clade (Beresford et al., 2005; now placed in Cryptillas – see under Macrosphenidae above). Alström et al. (2011a) noted that Locustellidae has priority over Megaluridae (as stated by Bock, 1994: 152).
The first comprehensive study of Locustellidae was published by Alström et al. (2011a; Fig. 3). It was based on a dataset comprising one mitochondrial and four nuclear loci for most of the species believed to belong in this family. The phylogeny strongly disagreed with earlier classifications at the generic level. All of the genera with more than one representative were found to be non-monophyletic: Bradypterus was separated into an Asian and an African clade, with Locustella and Megalurus pryeri nested within the former, and the monotypic Malagasy Dromaeocercus within the latter; only two of the five Megalurus species formed a clade that did not include other genera, and both Cincloramphus and Eremiornis were nested in one of the Megalurus clades. The non-monophyly of Bradypterus and Locustella had previously been found based on mitochondrial ND2 sequences of all Locustella, two Asian and three African Bradypterus and two Megalurus (M. pryeri and M. gramineus) (Drovetski et al., 2004). Moreover, the affinity of M. pryeri to Locustella had previously been suggested based on morphology (Morioka and Shigeta, 1993). Alström et al. (2011a) proposed a revised classification recognizing four instead of seven genera (Fig. 3). They acknowledged the non-monophyly of one of these genera (Megalurus), but stressed that the classification was tentative and took account of the phylogenetic uncertainty (i.e. conflict between their results and a study by Beresford et al., 2005 using another locus for five Locustellidae species, as well as morphology and vocalizations).
Figure
3.
Family Locustellidae based on concatenated nuclear ODC, MB, LDHB and GAPDH and mitochondrial cytochrome b sequences, based on Alström et al. (2011a). Species names follow revised taxonomy proposed by Alström et al. (2011c), but colors indicate genera according to Dickinson (2003) (red – Bradypterus; purple – Megalurus; dark blue – Dromaeocercus; pale blue –
Cincloramphus; orange – Eremiornis). # indicates that only cytochrome b was available for analysis. Posterior probabilities and maximum likelihood and parsimony bootstrap values are indicated at the nodes, in this order; an asterisk represents posterior probability 1.00 or bootstrap 100%. B. baboecala SA and B. baboecala Nig refer to samples from South Africa (transvaalensis/tongensis) and Nigeria (centralis), respectively. 1 Node affected differently by different types of analyses. 2 Non-monophyletic Megalurus tentatively recognized due to conflicting results between present and another study based on different loci and DNA versus morphology.
A recent study by Oliveros et al. (2012) unexpectedly found that the monotypic Sulawesi genus Malia was nested in Locustellidae and that the Philippine Robsonius was sister to Locustellidae. Five putative Locustellidae genera remain to be studied genetically: Amphilais
(monotypic, Madagascar), Megalurulus (four species, Melanesia), Buettikoferella (monotypic, Timor), Chaetornis (monotypic, Indian Subcontinent) and Elaphrornis (monotypic, Sri Lanka) (Gill and Donsker, 2013).
The species in the traditional genera Locustella and Bradypterus are renowned for being difficult to identify by morphological characters, and there has been much taxonomic confusion over the years (see Bairlein et al., 2006; Kennerley and Pearson, 2010). One new species,
Locustella (Bradypterus) alishanensis, was described as recently as 2000 from Taiwan, where it had been known to exist for many years but with confused taxonomy (Rasmussen et al., 2000). Dickinson et al. (2000) revised the poorly known Bradypterus seebohmi complex based on morphology and vocalizations, and tentatively recommended recognition of four species: B. seebohmi sensu stricto (s.s.), B. timorensis, B. montis and B. mandelli
(placed in Locustella by Alström et al., 2011). Alström et al. (2011a) analyzed mitochondrial cytochrome b from different populations of the latter two, and suggested that they were so similar that their status as separate species needs to be re-evaluated. Locustella timorensis
(previously Bradypterus timorensis), until recently only known from two specimens collected in 1932 on Timor (Dickinson et al., 2000), was recently rediscovered, along with a putative undescribed taxon from nearby Alor (Trainor et al., 2012; Verbelen and Trainor, 2012). Another undescribed taxon in the same complex has recently been found on Taliabu island, Indonesia (Rheindt, 2010). The L. thoracica (previously B. thoracicus) complex was revised by Alström et al. (2008) based on multiple independent datasets, and they proposed that three instead of one species be recognized. Locustella amnicola has usually been treated as a subspecies of L. fasciolata (e.g. Dickinson, 2003; Kennerley and Pearson, 2010), but has also been given species status (Watson et al., 1986 [with a note by Ernst Mayr stating that "The status of this species is in doubt."]; Stepanyan, 1990). Both Drovetski et al. (2004) and Alström et al. (2011a) showed that these two are markedly different genetically, supporting treatment as separate species. Alström et al. (2011a) also found deep cytochrome b divergences among some other taxa currently treated as conspecific, and suggested that species status might be warranted if corroborated by independent data: B. baboecala tongensis + B. b. transvaalensis vs. B. b. centralis + B. b. elgonensis; B. lopezi mariae + B. l. usambarae vs. B. l. ufipae; and M. palustris toklao vs. M. p. forbesi. Conversely, Alström et al. (2011a) and an earlier study by Drovetski et al.
(2004), found slight divergences between L. pleskei and L. ochotensis, and noted that their status as separate species needs to be studied further.
Bernieridae
The family name Bernieridae was proposed by Cibois et al. (2010) for a clade of Malagasy "warblers". These were previously classified in the traditional families Timaliidae, Sylviidae and Pycnonotidae, but were shown by mitochondrial DNA to form an endemic Malagasy radiation (Cibois et al., 1999, 2001; Fjeldså et al., 1999). Cibois et al. (2010) recognized 10 species in seven genera: Bernieria madagascariensis, Xanthomixis zosterops, X. cinereiceps, X. apperti, X. tenebrosa, Oxylabes madagascariensis, Thamnornis chloropetoides, Crossleyia xanthophrys, Hartertula flavoviridis and Cryptosylvicola randrianasoloi. Dickinson (2003) and Gill and Donsker (2013) included also Randia pseudozosterops in Bernieridae, although this species has not yet been included in any phylogenetic study. The first four species listed above have often been placed in the bulbul (greenbul) genus Phyllastrephus, and H. flavoviridis has been placed in the "warbler" genus Neomixis (e.g. Delacour, 1946).
The study by Fregin et al. (2012) is the most complete analysis of this family thus far with respect to number of loci, although the study by Alström et al.
(2011), based on four nuclear and one mitochondrial loci, investigated one additional monotypic genus (Cryptosylvicola), and the mitochondrial study by Cibois et al. (2001) included two additional species in the genus Xanthomixis. The tree in Fig. 4, which is based on all previously published sequence data, is well resolved and well supported, except for some uncertain relationships within a clade containing Oxylabes + Bernieria,
Hartertula, Cryptosylvicola and Thamnornis.
Figure
4.
Family Bernieridae based on concatenated nuclear ODC, MB, LDHB, GAPDH and RAG1 and mitochondrial cytochrome b sequences (only cytochrome b for X. cinereiceps; no LDHB, GAPDH for X. zosterops; and no RAG1 for C. randrianasoloi), based on Cibois(1999, 2001), Alström et al. (2011c) and Fregin et al. (2012), reanalyzed by Bayesian inference in six partitions (by locus). See Supplementary Appendix for details of samples and analyses. Posterior probabilities and maximum likelihood bootstrap values are indicated at the nodes, in this order; an asterisk represents posterior probability 1.00 or bootstrap 100%.
The family Donacobiidae was proposed by Aleixo and Pacheco (2006). It contains the single species Donacobius atricapilla, which occurs in Central and South America. This taxon has a chequered history: it was placed in the family Mimidae by e.g. Mayr and Greenway (1960), and later in the subfamily Troglodytinae by Sibley and Ahlquist (1990) and Sibley and Monroe (1990). However, Barker et al. (2004) found evidence for an association with Zosterops and Prinia based on one mitochondrial and one nuclear marker, and Alström et al. (2006) confirmed that it was firmly nested within Sylvioidea also using one mitochondrial and one nuclear marker (different ones from those used in Barker et al., 2004). Johansson et al. (2008a) found strong support for the position of Donacobius in a clade with Locustellidae and Bernieridae based on three nuclear loci, and this was corroborated by Fregin et al.
(2012) based on seven molecular markers. However, the relationships among these three families are uncertain, probably due to a short time between their separations, as indicated by the short internode between Donacobius and Bernieridae. From a biogeographical point of view, the distribution of Donacobius is remarkable, as all of its closest relatives, Locustellidae and Bernieridae, occur in the Old World.
Fregin et al. (2009) revised the family Acrocephalidae based on one mitochondrial and three nuclear loci for 32 out of the 38 species of Acrocephalus (mitochondrial data only for nine species), all eight Hippolais, all three Chloropeta and one Nesillas (Fig. 5). They recovered all Acrocephalus except A. aedon in one clade. The latter was sister to a clade with C. natalensis and C. similis as sisters to H. pallida and H. opaca, with H. caligata and H. rama as sisters to these four, and the generic name Iduna was erected for these seven species, with the suggested option of placing I. aedon in the monotypic genus Phragamaticola; the latter was adopted by Kennerley and Pearson (2010). The third species of Chloropeta, C. gracilirostris, was in an effectively unresolved position, and the monotypic genus Calamonastides was reinstated for it. The other four Hippolais (H. icterina,
H. polyglotta, H. olivetorum and H. languida) formed a clade with two sister pairs, and the generic name Hippolais was suggested to be restricted to these four species.
Figure
5.
Family Acrocephalidae based on concatenated nuclear ODC, MB, LDHB and mitochondrial cytochrome b sequences, based on Fregin et al. (2009). # indicates that only cytochrome b was available for analysis. Posterior probabilities and maximum likelihood and parsimony bootstrap values are indicated at the nodes, in this order; an asterisk represents posterior probability 1.00 or bootstrap 100%. Colors represent taxonomy according to Dickinson (2003): black – Acrocephalus; blue – Hippolais; red – Chloropeta; purple –
Nesillas.
The Acrocephalus warblers occurring on different islands in eastern Polynesia were previously considered a single widespread species, A.caffer (Holyoak and Thibault, 1984; Dickinson, 2003), whereas some authors suggested splitting this species into three, mainly because of wide geographical separation: A. caffer s.s. on the Society Islands, A. atyphus on Tuamotu, and A. mendanae in the Marquesas (Pratt et al., 1987; BirdLife International, 2000; Cibois et al., 2011a, b). Using mitochondrial sequence data, Cibois et al. (2007) showed that A. mendanae formed two independent lineages, which they proposed be treated as specifically distinct:
A. percernis (closely related to A. atyphus) and A. mendanae s.s. (sister to A. aequinoctialis). In a later study, Cibois et al. (2008) analyzed mitochondrial DNA from extinct Acrocephalus taxa from different Society Islands, and concluded that A. caffer s.l. was more appropriately treated as one extant species, A. caffer s.s. (Tahiti), and two extinct ones, A. longirostris (Mo'orea) and A. musae
(Raiatea, Huahine). A comprehensive study of Pacific Acrocephalus warblers (Cibois et al., 2011a) revealed complex colonization histories for several taxa, and revealed that A. luscinius is seprated into three distantly related clades, calling for a taxonomic revision.
Until recently, Acrocephalus orinus was only known from a single specimen collected in winter in India in 1867, and its validity as a species was controversial. A series of studies, and the surprise finding of a live individual in Thailand in March 2006, gradually led to the discovery of a breeding population at high elevation in north-eastern Afghanistan and adjacent Tajikistan, and has been found during the breeding season, and almost certainly also breeds, in Kyrgyzstan, eastern Uzbekistan and south-eastern Kazakhstan (Bensch and Pearson, 2002; Round et al., 2007; Pearson et al., 2008; Svensson et al., 2008, 2010; Timmins et al., 2010; Ayé et al., 2010; Koblik et al., 2011).
The four Palearctic Iduna species (previously Hippolais) have long been treated as two species, H. pallida and H. caligata (e.g. Watson et al., 1986), although the former was suggested to be split into H. pallida s.s. and H. opaca based on morphological, vocal and behavioral characteristics (Svensson, 2001; Parkin et al., 2004), and the latter has often been split into H. caligata s.s. and H. rama on the same grounds, as well as sympatric breeding distributions (e.g. Stepanyan, 1990; Svensson, 2001; Parkin et al., 2004). Helbig and Seibold (1999) found deep divergences within both H. pallida s.l. and H. caligata s.l. in mitochondrial DNA, later confirmed by Ottosson et al. (2005) in an extensive study that also included morphological data, and by Fregin et al. (2009) using mitochondrial and nuclear loci (Fig. 5), further supporting the splits based on non-molecular data.
Hirundinidae
The globally distributed family Hirundinidae (swallows and martins) form a morphologically and ecologically rather homogeneous taxon, constrained by their aerial foraging behavior (Turner, 2004). In total, 88 species are recognized (Gill and Donsker, 2013). Mayr and Bond (1943) analyzed the swallows and martins based on morphology and nest-building characteristics, and judged that they could be divided into ten natural groups. Phedina was judged to be "possibly related to Hirundo". White-backed Swallow (Cheramoeca), Greyrumped Swallow (Pseudhirundo) and the African sawwings (Psalidoprogne) were judged to be "so clear-cut and isolated that they require no further comment". The Barn and Cliff Swallow group consisted of Delichon,
Hirundo and Petrochelidon. The genus Riparia was judged "a primitive genus without close relatives", while the crag martins (Ptyonoprogne); the rough-wing swallow group (Alopochelidon, Neochelidon and Stelgidopteryx); the Atticora group (Atticora, Notiochelidon, Orochelidon and Pygochelidon); the tree swallows (Tachycineta, then split also into Callichelidon, Lamprochelidon,
Iridoprocne) and the purple martin group (Progne) each qualified as a distinct group.
Sheldon and Winkler (1993) performed the first molecular study of the family, using the DNA-DNA hybridization to estimate the phylogeny of a limited number of species, and later used the phylogeny to superimpose nest construction data. Their results largely corroborated the groups defined by Mayr and Bond (1943). Winkler and Sheldon (1993) reconstructed the nestexcavating Psalidoprogne as sister to all other swallows, which were divided into two monophyletic groups, the mud-nesting Hirundo group, and a clade containing both nest excavating and cavity-nesting species. The ancestral mode of nest-building in this clade appeared to be nest-excavating, as cavity nesting is reconstructed as a recent apomorphy, which is only found in two lineages out of three in a polytomy containing the nestexcavating Riparia riparia, the cavity-nesting New World endemics (Atticora, Haplochelidon, Neochelidon,
Progne, Pygochelidon and Stelgidopteryx) and the cavitynesting New World tree swallows (Tachycineta). The nest-excavating Cheramoeca, Phedina, Pseudhirundo and Riparia cincta all trace their ancestry to nodes basal to the cavity-nesting species.
The most comprehensive molecular study of swallow phylogeny (Sheldon et al., 2005) identified the two river martins Pseudochelidon as sisters to all other swallows, albeit based on mitochondrial sequence data only. The combined mitochondrial and nuclear data available for a wide range of other swallows reconstructed two large clades comprising the remaining species, which Sheldon et al. (2005) termed "core martins, mud nesters and basal relicts". The core martin clade was dominated by Progne, Riparia and Tachycineta, and also included the less speciose Alopochelidon, Atticora, Haplochelidon, Neochelidon, Notiochelidon, Phedina, Pygochelidon and Stelgidopteryx. The mud-nesters contained Cecropsis, Delichon, Hirundo, Petrochelidon and Ptyonoprogne. The basal relicts were placed as sisters to the mudnesters and consisted of the groups judged by Mayr and Bond (1943) to be highly distinct and isolated from the others: White-backed Swallow (Cheramoeca), Greyrumped Swallow (Pseudhirundo) and the African sawwings (Psalidoprogne). Although Sheldon et al. (2005) were unable to determine the exact branching sequence of the "core martins, mud nesters and basal relicts", they concluded that excavation of nests is the ancestral condition in swallows, and that the mud-nesters constitute a monophyletic radiation. Similarly, the core martins constituted a monophyletic group, within which two clades are cavity-nesters. The position of these two clades was unresolved in relation to the nest-excavating sand martins, so it was not possible to determine whether cavity-nesting arose once or twice.
Beside these major analyses, several studies focusing on particular genera have further clarified the phylogeny of swallows. Babin (2005) analyzed the geographic variation and speciation in the rough-winged swallows (Stelgidopteryx), shedding light on a complex picture of ancestral polymorphism not yet sorted among lineages. Dor et al. (2010) analyzed the Hirundo clade, arriving at results that are entirely congruent with Sheldon et al. (2005) but with the addition of H. megaensis and H. nigrorufa, which were not investigated in the latter analysis. Dor et al. (2010) and Zink et al. (2006) also undertook phylogeographical studies of H. rustica. Moyle et al. (2008) estimated a phylogeny of the New World martins (Progne), and discovered remarkably intricate and close relationships, polyphyly and possible hybrid introgression, and came to the conclusion that a population genetic perspective would probably be required to clarify the taxonomy, particularly among Amazonian populations.
Whittingham et al. (2002), Cerasale et al. (2012) and Dor et al. (2012) studied the phylogeny of Tachycineta using different approaches. Whittingham et al. (2002) used concatenated mitochondrial loci, Cerasale et al. (2012) used complete mitochondrial genomes, while Dor et al. (2012) used 16 nuclear introns to compare concatenation to a coalescent-based species tree inference method. All these approaches yielded different topologies, particularly concerning the positions of the North American T. bicolor, Peruvian T. stolzmanni, Hispaniolan T. euchrysea and Bahamian endemic T. cyaneoviridis.Dor et al. (2012) argued that the method of extracting information from the variability in coalescence times between independent gene genealogies outperforms other methods of analysis, thus advocating their species tree as the currently most reliable hypothesis. Similarly, Kirchman et al. (2000) analyzed relationships among different populations of the Petrochelidon fulva complex, and confirmed previous suggestions that the South American populations are distinct (P. rufocollaris).
A phylogeographical study of Riparia riparia and R. diluta, which were previously considered a single species (e.g. Peters, 1960b), was undertaken by Pavlova et al. (2008). It corroborated fieldwork suggesting that these are specifically distinct (Gavrilov and Savchenko, 1991; Goroshko, 1993), and suggested that they diverged sometime between the late Pliocene and middle Pleistocene. It also suggested a Pleistocene split between samples from central Siberia and Mongolia. However, no molecular studies have been carried out yet on the southern, mainly Chinese, populations, which have usually been treated as conspecific with R. diluta.
Pycnonotidae
The Pycnonotidae comprises approximately 150 species (Gill and Donsker, 2013) distributed across Africa and southern Asia. The first major molecular study of bulbuls (27 species; Pasquet et al., 2001), using mitochondrial sequence data, focused on the genus Criniger, but their taxon sampling allowed general inferences about the family as a whole. Their results found a basal division into an African and a mainly Asian clade, and revealed that African and Asian clades of Criniger were not closely related. Warren et al. (2005) showed that the bulbuls on Madagascar and on the islands of the Indian Ocean (Hypsipetes) originated from Asian ancestors. Moyle and Marks (2006) followed up by using also nuclear genes and additional taxa (in total 57 species). Their phylogeny provided further detail and corroborated the two major clades, of which only the Asian clade included taxa also occurring in Africa, namely representatives of the genus Pycnonotus. In addition to the two main clades, Moyle and Marks (2006) found the monotypic African genus Calyptocichla to be sister to the Asian clade, but with weak support. A study by Johansson et al. (2007a) further clarified the phylogeny of the Afrotropical clade, among other things placing Calyptocichla in this clade, and proposing a revised taxonomy in which the genus Atimastillas was split from Chlorocichla, and the polyphyletic Andropadus was divided into four genera, Andropadus, Arizelocichla, Eurillas and Stelgidillas. Oliveros and Moyle (2010) studied the origin of the Philippine bulbuls, adding substantial resolution to the Asian clade. They found a number of instances of high genetic divergence within species, e.g. three deeply divergent lineages within Ixos philippinus (confirmed for two of these by Silva-Iturriza et al., 2010), and non-monophyly of the genus Ixos. Their preferred taxonomic solution was to include the genus Microscelis and Philippine members of Ixos in Hypsipetes, a genus to which they have previously been allocated (e.g. Rand and Deignan, 1960).
The phylogenetic position of Neolestes torquatus, which was originally described as an aberrant malaconotid by Cabanis (1875), has been studied genetically and morphologically, and has been shown to be a bulbul, belonging in the Afrotropical clade (Dowsett et al., 1999; Moyle and Marks, 2006; Johansson et al., 2007a; Oliveros and Moyle, 2010; Zuccon and Ericson, 2010). Johansson et al. (2007a) and Zuccon and Ericson (2010) found good support for Calyptocichla in a sister group to the Afrotropical clade, in a clade that also contained Andropadus importunus and A. gracilirostris. Today, all generally recognized genera in Pycnonotidae have been included in molecular phylogenies, but much work remains to be done at the intra- and interspecific levels, as illustrated by Fuchs et al. (2011), who found significant biometric differences between lowland and montane populations of Phyllastrepus debilis in Tanzania.
Five primary clades have been identified in the babbler group, three of which are made up of traditional babblers, one of a mixture of babblers (mainly Yuhina) and white-eyes (latter traditionally Zosteropidae), and one containing babblers and Sylvia warblers (Cibois, 2003a; Alström et al., 2006; Gelang et al., 2009; Fregin et al., 2012; Moyle et al., 2012). The three most recent of these papers all recovered the same five primary clades, but differed in their taxonomic recommendations. Gelang et al. (2009) separated Sylviidae and Timaliidae at the family level, and treated Leiothrichinae, Pellorneinae, Timaliinae and Zosteropinae as subfamilies within Timaliidae. Fregin et al. (2012) recognized all five clades at the family level: Leiothrichidae, Pellorneidae, Timaliidae, Sylviidae and Zosteropidae. Moyle et al. (2012) took a third position in recognizing Sylviidae, Timaliidae and Zosteropidae at the family level, with Leiothrichinae, Pellorneinae and Timaliinae as subfamilies in Timaliidae. We here follow Fregin et al. (2012) and Gill and Donsker (2013) and treat the five primary babbler clades at the rank of family, as we consider that to be in better agreement with the treatment of the other primary clades in Sylvioidea than the treatments of Gelang et al. (2009) and Moyle et al. (2012).
Timaliidae
The Timaliidae clade is restricted to (southern) Asia, and comprises the genera Dumetia, Macronus (see below), Pomatorhinus (see below), Rhopocichla, Spelaeornis, Sphenocichla (see below), Stachyris (see below), Timalia and Xiphirhynchus (see below) (Cibois, 2003; Gelang et al., 2009; Moyle et al., 2012), in total c. 55 species (Gill and Donsker, 2013). Macronus, Pomatorhinus and Stachyris, as traditionally circumscribed (e.g. Deignan, 1964b; Dickinson, 2003), have all been found to be non-monophyletic, and Moyle et al. (2012) suggested a number of rearrangements: resurrection of Mixornis (for M. gularis, M. flavicollis and M. kelleyi; two latter unstudied by molecular methods); restricting Macronus to M. striaticeps and M. ptilosus; reinstatement of Cyanoderma for several species placed in Stachyris or Stachyridopsis (e.g. C. chrysaeum, C. erythropterum, C. ruficeps and C. rufifrons); restricting Stachyris to a group containing e.g. S. nigriceps, S. striolata, the recently described S. nonggangensis (Zhou and Jiang, 2008; not studied by molecular methods) and Sphenocichla; and splitting Pomatorhinus into Pomatorhinus s.s. (comprising most species, including P. [previously Xiphirhynchus] superciliaris) and Megapomatorhinus (new genus name, containing M. hypoleucos, M. erythrocnemis, M. erythrogenys and M. swinhoei).
The relationships within Pomatorhinus s.l. have recently been examined by Dong et al. (2010a) using two mitochondrial and four nuclear markers for a relatively small number of taxa and by Reddy and Moyle (2011) based on the same two mitochondrial markers for a larger number of taxa. Both studies, as well as Moyle et al. (2012), inferred Stachyris s.s. nested within Pomatorhinus s.l., although there was conflict between the studies regarding the position of the Stachyris s.s. clade in relation to the primary Pomatorhinus s.l. clades. Moreover, all these, as well as Cibois (2003a) and Gelang et al. (2009), recovered Xiphirhynchus (see above) deep inside the Pomatorhinus s.l. clade. Reddy and Moyle (2011) found multiple deep divergences within traditional species of Pomatorhinus s.l. as well as nonmonophyletic species, and also identified 27 lineages with diagnosable plumage differences (i.e. phylogenetic species sensu Cracraft, 1989). Previously, Collar (2006) and Collar and Robson (2007) suggested elevation of several taxa treated as subspecies to the rank of species based on morphology and vocalizations.
The genera Actinodura, Babax (see below), Cutia, Garrulax (see below), Heterophasia, Leiothrix, Liocichla, Minla, Turdoides, Kupeornis (see below) and Phyllanthus (see below) have been recovered as members of the Leiothrichidae clade (Cibois, 2003; Gelang et al., 2009; Moyle et al., 2012), in total c. 130 species (Gill and Donsker, 2013). They are widely distributed across (southern) Asia, although the Turdoides clade (including Kupeornis and Phyllanthus; see below) has radiated extensively in Africa.
The genera Garrulax, Actinodura, Minla, Heterophasia and Turdoides, as generally circumscribed (e.g. Deignan, 1964b; Dickinson, 2003), have been found to be non-monophyletic (Cibois, 2003; Gelang et al., 2009; Luo et al., 2009; Dong et al., 2010b; Moyle et al., 2012), and based on these results Moyle et al. (2012) suggested a taxonomic revision of this clade. They proposed that Garrulax be divided into Garrulax, Grammatoptila (monotypic: G. striata), Ianthocincla and Trochalopteron, and Babax synonymized with Ianthocincla. Moreover, they proposed that Kupeornis and Phyllanthus be subsumed in Turdoides, Heterophasia annectans moved to Minla, and Minla cyanouroptera and M. strigula transferred to Actinodura. However, it should be noted that no molecular study has included the type species of Garrulax, G. rufifrons, so the circumscription of this genus should be considered preliminary. The reinstatement of Trochalopteron was earlier proposed by Luo et al. (2009) based on molecular data as well as the observation that all species in that clade have speckled eggs, unlike any other traditional Garrulax (though eggs have not been studied for all species). Earlier, Rasmussen and Anderton (2005) and Collar and Robson (2007) proposed reclassifications based on morphological features, and some of these agree with the molecular results, whereas others do not, and some have not yet been tested by molecular markers; Rasmussen and Anderton (2012) followed various genetic analyses in further revising Garrulax and other laughingthrush genera.
The taxonomy of Garrulax canorus has been assessed by molecular markers by Li et al. (2006, 2009), who proposed that the Taiwanese taxon should be treated as specifically distinct, G. taewanus (placed in genus Leucodioptron in latter study). The same conclusion was previously reached by Collar (2006) based on morphology. Several other taxa were also elevated from the rank of subspecies to species by Rasmussen and Anderton(2005, 2012), Collar (2006) and Collar and Robson (2007), mainly based on morphology and vocalizations. A new species, Liocichla bugunorum, was described as recently as 2006 (Athreya, 2006).
Pellorneidae
The Pellorneidae clade comprises species in the genera Gampsorhynchus, Graminicola, Illadopsis, Jabouilleia (see below), Kenopia, Malacocincla, Malacopteron, Napothera, Pellorneum, Ptilocichla, Ptyrticus (see below), Rimator (see below), Schoeniparus (formerly Alcippe; see below), Trichastoma (see below) and Turdinus (Cibois, 2003; Gelang et al., 2009; Moyle et al., 2012), in total c. 70 species (Gill and Donsker, 2013). In addition, Laticilla (previously Prinia) burnesii was found by Olsson et al. (2013a) to belong in this clade. With the exception of the African Illadopsis, this family occurs only in Asia.
The genera Illadopsis, Napothera, Malacocincla and Pellorneum, as traditionally defined (e.g. Deignan, 1964b; Dickinson, 2003), have been inferred to be nonmonophyletic (Cibois, 2003; Moyle et al., 2012), and in the taxonomic revision suggested by Moyle et al. (2012) several taxa were synonymized: Ptyrticus with Illadopsis; Trichastoma with Pellorneum; and Jabouilleia and Rimator with Napothera. These authors also moved some Napothera and some Malacocincla to Turdinus, while the remaining Malacocincla were placed in Pellorneum. However, the name Turdinus seems to be misapplied. The type species of this genus, T. macrodactylus, was not included in the analysis by Moyle et al. (2012), but in the study by Gelang et al. (2009), it was sister to Graminicola, with strong support, while Malacocincla abbotti, which was also placed in Turdinus by Moyle et al. (2012), was in a different clade (with high posterior probability but low maximum likelihood bootstrap support). The revised classification by Collar and Robson (2007), which was based mainly on morphological traits, agrees in some respects but differs in others from the treatment by Moyle et al. (2012).
A new species, Jabouilleia naungmungensis, was recently described from Myanmar (Rappole et al., 2005), although this was considered a subspecies of J. (=Rimator) danjouei by Collar and Robson (2007) and Collar (2011). Graminicola bengalensis was recently suggested to be split into two species, G. bengalensis s.s. (Indian subcontinent) and G. striatus (southern China, northern South-East Asia) (Leader et al., 2010).
Sylviidae sensu stricto
The circumscription of Sylviidae today is radically different from the traditional usage of this name (e.g. Watson et al., 1986; Sibley and Monroe, 1990; Dickinson, 2003; Bairlein et al., 2006; see introduction). The Sylviidae s.s. clade includes the genera Chamaea, Chrysomma, Conostoma (see below), Fulvetta (see below), Horizorhinus (see below), Lioparus (see below), Lioptilus, Moupinia (sometimes synonymized with Chrysomma), Myzornis, Paradoxornis (see below), Parisoma (see below), Parophasma, Pseudoalcippe (see below), Rhopophilus and Sylvia (Barhoum and Burns, 2002; Cibois, 2003; Gelang et al., 2009; Moyle et al., 2012), in total c. 70 species (Gill and Donsker, 2013). This family is geographically more widespread than the previous babbler clades. Although the majority occurs in southern Asia, Sylvia (including Horizorhinus, Parisoma and Pseudoalcippe; see below) is also distributed in northern Asia, Europe and Africa, Parophasma and Lioptilus are restricted to Africa, and Chamaea is restricted to western North America, as the only babbler representative in the New World.
The genus Paradoxornis has been shown to be nonmonophyletic, with at least Conostoma nested within Paradoxornis (Yeung et al., 2011). In a comprehensive analysis of the parrotbills, Yeung et al. (2011) identified three major clades in both mitochondrial and nuclear gene trees, further supported by body size and plumage coloration. The monotypic genus Conostoma was confirmed to belong in one of these clades. However, while trees inferred by combined nuclear sequences supported monophyly of the parrotbills, mitochondrial DNA suggested that one of the parrotbill clades was more closely related to Chamaea, Fulvetta and Lioparus (two latter treated in genus Alcippe) than to the other parrotbills; however, the latter topology received no maximum likelihood bootstrap support (but high posterior probability). Penhallurick and Robson (2009) proposed a reclassification of the parrotbills into eight genera (including a new genus name, Sinosuthora, containing S. alphonsiana, S. brunnea, S. conspicillata, S. przewalskii, S. webbiana and S. zappeyi), based on size, plumage and vocalizations, backed up by a published study of mitochondrial DNA (Yeung et al., 2006; see correction of one of these generic names in Penhallurick, 2010). Previously, King and Robson (2008) had suggested splitting Paradoxornis ruficeps into two species, P. ruficeps s.s. and P. bakeri.
Shirihai et al. (2001) and Voelker and Light (2011) inferred the relationships of the genus Sylvia based on mitochondrial DNA, and found that the African species previously placed in the genus Parisoma were nested within the essentially Palearctic Sylvia. The latter authors, and previously Voelker et al. (2009), also found that Horizorhinus dohrni and Pseudoalcippe abyssinica formed a sister clade to Sylvia borin, and placed both in the genus Sylvia; P. abyssinica had previously been found to be closely related to Sylvia (S. melanocephala) by Cibois (2003a). This was later corroborated by Moyle et al. (2012; using P. atriceps), who also found Lioptilus nigricapillus to be part of this clade. Several groups of Sylvia warblers have been the subject of taxonomic studies based on multiple character sets, usually including DNA, which have resulted in the recognition of a larger number of species, e.g. the splitting of S. crassirostris from S. hortensis (Shirihai et al., 2001), S. balearica from S. sarda (Shirihai et al., 2001), and S. subalpina (synonym S. moltonii) from S. cantillans (Brambilla et al., 2008a, b, c). The long-standing uncertainty over species limits in the S. curruca complex (e.g. Watson et al., 1986; Cramp, 1992; Martens and Steil, 1997; Shirihai et al., 2001; Loskot, 2001, 2005; Bairlein et al., 2006) was recently investigated using mitochondrial DNA (Olsson et al., 2013b), but although six long-separated clades representing the taxa althaea, blythi, curruca, halimodendri, margelanica and minula were recovered, the gene tree partly disagreed with morphological evidence, and it was concluded that independent data were required to resolve the issue.
Zosteropidae
The following genera have been found to belong in the Zosteropidae clade: Chlorocharis, Cleptornis, Heleia, Lophozosterops, Oculocincta, Rukia, Speirops, Sterrhoptilus, Yuhina, Woodfordia, Zosterops and Zosterornis (latter placed in Stachyris in earlier studies) (Cibois et al., 2002; Cibois, 2003a; Zhang et al., 2007; Gelang et al., 2009; Moyle et al., 2009, 2012). Most of these have been represented by single species in these analyses, but those that have been represented by two or more species (Lophozosterops, Yuhina, Zosterops and Zosterornis) have been found to be non-monophyletic. For example, Yuhina is separated into at least three (Moyle et al., 2012) or four (Zhang et al., 2007; Moyle et al., 2009) monophyletic groups, with Y. diademata being sister to all Zosteropidae, and one of these Yuhina clades is sister to the rest of the Zosteropidae. The main Zosterops clade, which also includes at least Chlorocharis, Rukia, Speirops and Woodfordia, has been termed a "great speciator" by Moyle et al. (2009), as it shows evidence of an exceptionally rapid speciation rate and expansion over a vast area covering subsaharan Africa, much of southern Asia and Australasia. The monotypic genus Hypocryptadius was previously considered related to Zosterops, but was recently shown to be a forest-adapted sparrow (Fjeldså et al., 2010). A thorough taxonomic revision of the Zosteropidae is obviously greatly needed.
The genus Alcippe provides one of the most extreme examples of the difference between past taxonomic judgments based on morphology (e.g. Deignan, 1964b; Dickinson, 2003) and recent molecular phylogenies. In a study based on mitochondrial DNA by Pasquet et al. (2006) the genus was found to consist of four different unrelated clades, the positions of which have gradually been clarified in multilocus analyses by Gelang et al. (2009) and Moyle et al. (2012). One clade, Schoeniparus (containing S. cinereus, S. castaneceps, S. rufogularis, S. brunneus and S. dubius), was found to be nested in Pellorneidae; two clades, Fulvetta (including e.g. F. cinereiceps, F. ruficapilla and F. vinipectus) and Lioparus (monotypic: L. chrysotis), were nested in Sylviidae; whereas the position of the Alcippe s.s. clade (comprising e.g. A. poioicephala and A. morrisonia) is somewhat uncertain. The latter clade was recovered by Gelang et al. (2009) as sister to Pellorneidae with good support, whereas Moyle et al. (2012) recovered it as sister to Leiothrichidae, also with good support. The latter authors remarked that the position of this clade is mainly weakly supported or even contradicted by single loci. More research is needed to establish where the Alcippe s.s. clade belongs in the babbler phylogeny.
Phylogeographical studies of the Alcippe morrisonia complex by Zou et al. (2007) and Song et al. (2009) revealed unexpectedly deep divergences between different geographical areas (oldest split dated to 11.6 million years ago by Song et al., 2009). Moreover, Zou et al. (2007) found that one of these clades (A. m. fratercula from Yunnan, China) is more closely related to A. peracensis annamensis (Vietnam) than to other taxa treated as A. morrisonia. Based on these results, Gill and Donsker (2013) split A. morrisonia into four species: A. morrisonia s.s. (Taiwan), A. davidi (Shaanxi, Gansu, Sichuan, Hunan, Guangxi Provinces in China, northern Vietnam), A. fratercula (Yunnan Province in China to central Myanmar and northern Thailand) and A. hueti (Fujian, Guangdong and Hainan Provinces, China). Collar (2006) and Collar and Robson (2007) revised the taxonomy of several species of Fulvetta and Alcippe s.s. based mainly on morphological characters; several of these have a contorted taxonomic history.
Phylloscopidae
The family Phylloscopidae, as presently circumscribed, comprising the genera Phylloscopus and Seicercus, was proposed by Alström et al. (2006). Sibley and Ahlquist (1990) and Sibley and Monroe (1990) placed these genera in the subfamily Acrocephalinae together with many other genera, while Dickinson (2003) put them in the subfamily Phylloscopinae, together with Tickellia, Abroscopus, Eremomela, Sylvietta and Graueria. Neither of the classifications by Sibley and Ahlquist (1990), Sibley and Monroe (1990) or Dickinson (2003) are consistent with phylogenetic studies (Beresford et al., 2005; Alström et al., 2006; Johansson et al., 2007b; Fregin et al., 2012; Olsson et al., 2013a), although Graueria has not yet been studied phylogenetically. The most comprehensive phylogeny of Phylloscopidae to date (Johansson et al., 2007b) was based on two mitochondrial and one nuclear loci for 55 of the 68 species recognized at that time (Dickinson, 2003). Other studies based on smaller, but partly different datasets have been published (Richman and Price, 1992; Martens et al., 2004, 2008; Olsson et al., 2004, 2005; Päckert et al., 2004, 2009). Figure 6 shows a tree for all of the species in these studies based on the same sequences, but reanalyzed here. As has already been remarked in some of the previous studies, the tree shows that Seicercus is nested within Phylloscopus, rendering the latter genus non-monophyletic. The tree also suggests that Seicercus is non-monophyletic, although this has poor statistical support. As has also been suggested by previous authors, the taxonomic implications are that either Seicercus be synonymized with Phylloscopus or the complex be split into more than two genera. We await a more complete taxonomic study that is in preparation.
Figure
6.
Family Phylloscopidae based on concatenated nuclear MB and mitochondrial cytochrome b and 12S sequence data, based on Olsson et al. (2004, 2005), Alström et al. (2006, 2010, 2011d), Saitoh et al. (2010) and Johansson et al. (2007), reanalyzed by Bayesian inference in three partitions (by locus). See Supplementary Appendix for details of samples and analyses. Posterior probabilities and maximum likelihood bootstrap (MLBS) values are indicated at the nodes, in this order; an asterisk represents posterior probability 1.00 or bootstrap 100%. 1P. calciatilis + P. cantator 85% MLBS; 2P. borealis + P. examinandus 60% MLBS; 3S. castaniceps + S. grammiceps MLB 87%.
Alström et al. (2006) found that, based on sequence data from two loci, one mitochondrial and one nuclear, that two species of Cettia, one species each of Urosphena, Tesia, Abroscopus and Tickellia, and Orthotomus cuculatus formed a clade, well separated from a diverse number of other passerines. They proposed the family name Cettiidae for this group. This clade (limited to one species each of Cettia, Abroscopus and Tickellia) was corroborated by Johansson et al. (2008a) based on three nuclear loci. Irestedt et al. (2011) concluded, based on four loci, that Hemitesia is also part of this clade. Two of these studies (Alström et al., 2006; Irestedt et al., 2011) indicated that the genus Cettia is non-monophyletic. Alström et al. (2011a) revised the family based on one mitochondrial and three nuclear loci (Fig. 7). They confirmed the previous findings, including the strong non-monophyly of the genus Cettia, as this genus is scattered across the entire family tree. Based on these results, they proposed a revised classification in which Cettia is divided into Cettia s.s. and Horornis and one species (C. pallidipes) transferred to Urosphena (where it was sometimes placed previously); Oligura (Tesia) castaneocoronata moved to Cettia; Hemitesia synonymized with Urosphena; and Phyllergates reinstated as the generic name for O. cuculatus (Fig. 7). Previously, King (1989) proposed a new classification of some Tesia and Urosphena based on morphology, vocalizations and behavior, which is partly in agreement with the molecular study of Alström et al. (2011a).
Figure
7.
Family Cettiidae based on concatenated nuclear ODC, MB and GAPDH and mitochondrial cytochrome b sequence data (only cytochrome b for the three species indicated with *, and for most of the samples of different subspecies), based on Alström et al. (2011b). Species names follow revised taxonomy proposed by Alström et al. (2011b). Names in black were previously placed in Cettia. # indicates node with poor support. Illustrations by Ren Hathway (Phyllergates), Brian Small (U. neumanni) and Jan Wilczur (Abroscopus, Tickellia) from Bairlein et al. (2006), and by Brian Small (others) from Kennerley and Pearson (2010).
Two of the species, Horornis carolinae and H. haddeni, were recently described (Rozendaal, 1987; LeCroy and Barker, 2006; respectively). Moreover, the taxonomy of several taxa has been much debated. Horornis diphone has variously been treated as a single species, or split into H. diphone s.s. and H. canturians, generally without explanation (cf. Delacour, 1942–1943; King and Dickinson, 1975; Morony et al., 1975; Watson et al., 1986; Sibley and Monroe, 1990; Inskipp et al., 1996; Baker, 1997; Dickinson, 2003; Bairlein et al., 2006; Kennerley and Pearson, 2010). According to mitochondrial DNA, the Japanese H. diphone cantans and central Chinese H. d. canturians were recently found to form a clade separate from the northern H. d. borealis (cf. Fig. 7). This is inconsistent with morphology, as the two latter are similar in plumage and structure, while the first one is more divergent. The authors concluded that a more comprehensive study was needed before any conclusions could be drawn. Furthermore, Horornis seebohmi has often been treated as a subspecies of H. diphone s.l. (e.g. Delacour, 1943; Watson et al., 1986; Baker, 1997), although some authors considered H. seebohmi to be a distinct species, based on unpublished differences in song and lack of the pronounced sexual size dimorphism of H. diphone/H. canturians (King and Dickinson, 1975; Inskipp et al., 1996; Dickinson, 2003). The specific status of H. seebohmi has recently been supported based on vocalizations and mitochondrial DNA (Hamao et al., 2008) as well as multilocus data (Alström et al., 2011a).
Horornis acanthizoides has recently been split into H. acanthizoides s.s. and H. brunnescens based on a study of morphology, vocalizations and mitochondrial DNA (Alström et al., 2007). Olsson et al. (2006) concluded, based on congruence of gene trees from one mitochondrial and one nuclear loci, that H. vulcanius is nested within H. flavolivaceus, and that some of the subspecies of H. flavolivaceus be moved to H. vulcanius. This was, however, contradicted in the study by Alström et al. (2011a; Fig. 7), which comprised a larger number of loci and samples (including all of the samples from Olsson et al., 2006). The latter study inferred deep divergences between two main H. flavolivaceus clades, with the taxon intricatus represented in both (Fig. 7). The authors concluded that more data, including unsampled subspecies, would be needed to resolve this issue. Deep divergences have also been found between different populations of Horornis fortipes (Alström et al., 2011a; cf. Fig. 7), although contradictory results from different analyses prevented taxonomic conclusions from being drawn. The same study also found that western and eastern populations of Cettia cetti were separated into rather divergent clades (cf. Fig. 7), calling for additional studies.
Scotocercidae
The family Scotocercidae was recently proposed, comprising the single species Scotocerca inquieta (Fregin et al., 2012). This was based on molecular (Alström et al., 2011c; Fregin et al., 2012) and morphological data (Alström et al., 2011c). The earlier of these studies also suggested that western and eastern populations might be different at the species level, and called for further studies.
Erythrocercidae
The Afrotropical genus Erythrocercus has been reconstructed in an unresolved position in relation to Cettidae and Aegithalidae (Alström et al., 2011c; Fregin et al., 2012). The three species in the genus (Dickinson, 2003; Gill and Donsker, 2013) are morphologically and ecologically divergent from both Cettidae and Aegithalidae, and Fregin et al. (2012) proposed the family name Erythrocercidae based on molecular, morphological and ecological evidence.
Aegithalidae
The family Aegithalidae, bushtits (or long-tailed tits), is one of the smallest families within Sylvioidea, comprising only 13 species (Gill and Donsker, 2013). Nine of these are Eurasian Aegithalos, two are Asian Leptopoecile, one is the Javan endemic Psaltria, and one is the North and Central American Psaltriparus. Psaltria does not appear to have been investigated in any molecular phylogeny, but three multilocus studies have shown Aegithalos and Psaltriparus to be sisters, with Leptopoecile sister to these (Johansson et al., 2008a; Päckert et al., 2010; Fregin et al., 2012). Dai et al. (2010) analyzed mitochondrial DNA for four Chinese Aegithalos species, while the relationships among all Aegithalidae species except Psaltria were studied by a combination of mitochondrial and nuclear markers by Päckert et al. (2010). The latter study obtained a well-resolved and strongly supported tree, except for a surprising lack of differentiation among four of the species recognized by Gill and Donsker (2013): A. fuliginosus, A. iouschistos, A. bonvaloti and A. sharpei. Additionally, they found deep splits among different populations of especially A. concinnus. Dai et al. (2011, 2013) confirmed the existence of deeps splits within A. concinnus, and evaluated their causes. In a study of mitochondrial DNA of A. caudatus sampled across Russia, Zink et al. (2008) found no strong geographical structure.
Hyliidae
Few taxa can show a record of taxonomic volatility as extreme as that of the Afrotropical Pholidornis, which has been placed in Dicaeidae, Estrildidae, Hyliidae, Meliphagidae, Nectariniidae, Remizidae and Sylviidae (Sefc et al., 2003 and references therein). The uncertainty regarding the Afrotropical Hylia has been less extreme, but opinion has been divided. The most favored positions have been to treat it as a sunbird (Sclater, 1930; Bannerman, 1948; Brosset and Erard, 1986) or a warbler (Chapin, 1953; Sibley and Monroe, 1990; Dowsett and Dowsett-Lemaire, 1993; Keith, 1997), but e.g. Meliphagidae (Beecher, 1953) and Paridae (Keith, 1997) have also been suggested. There is morphological evidence for a relationship between the two genera in the form of a brush-tipped tongue and a long hyoid with flattened epibranchial horns, characters that also suggest a relationship with Nectarinidae (Bannerman and Bates, 1924). These similarities prompted Bates (1930) to erect the family Hyliidae, but this was not widely accepted by subsequent authors, most of whom did not consider Pholidornis and Hylia as members of the same family. The phylogenetic position of both Hylia and Pholidornis continued to be contentious up to the first molecular analysis by Sefc et al. (2003), who showed that they were closely related and belong in Sylvioidea, later corroborated for Hylia by Beresford et al. (2005), Fuchs et al. (2006), Irestedt e al. (2011) and Fregin et al. (2012). Fregin et al. (2012) tentatively proposed that the family Hyliidae should be resurrected for Hylia and Pholidornis.
Marks (2010) revealed the existence of four (five) markedly divergent haplotype groups within Hylia prasina, indicating that further taxonomic studies are warranted.
Pnoepygidae
The Asian Pnoepyga wren babblers were classified as babblers (Timaliidae s.l.) until Gelang et al. (2009) found that they were not closely related to any of the babbler clades, and erected the monotypic family Pnoepygidae. The family contains only four species (Gill and Donsker, 2013). The taxonomy of the Taiwanese endemic P. formosana has been debated, and it has variously been treated as conspecific with P. pusilla (Deignan, 1964b; Cheng, 1987) or with P. albiventer (Dickinson, 2003; Liu et al., 2010). Collar (2006) suggested, based on morphological evidence, that the Taiwanese taxon should be considered a distinct species, and the same conclusion was reached by Päckert et al. (2012) based on DNA sequence data and analyses of songs. Päckert et al. (2012) also found both molecular and acoustic evidence of a divergence within P. albiventer. They proposed that the Chinese taxon should be treated as a species, P. mutica, but refrained from suggesting any taxonomic action for a genetically equally divergent Myanmar population within the P. albiventer complex, for which they lacked acoustic data.
The relationships within the superfamily Sylvioidea have been enormously clarified in recent years thanks to a large number of molecular studies. These analyses have confirmed numerous conclusions drawn from earlier morphological studies, but they have also suggested multiple cases of previously unexpected relationships. Some of the surprising findings revealed by molecular methods have been supported by more careful reevaluation of morphological data (earlier morphologybased classifications have generally been based on superficial morphological similarity rather than thorough analyses). However, many of the results from molecular studies need to be corroborated by independent data. Most molecular phylogenies are based on a small number of loci (sometimes only mitochondrial), and even in multilocus analyses, strongly supported relationships frequently receive their support from a small fraction of the analyzed loci, with the majority of the loci neither supporting nor contradicting these relationships. As genomic data are becoming cheaper and more easily obtainable, many of the recent discoveries will be reevaluated by considerably larger datasets in the not too distant future. One of the challenges will be to resolve the relationships among the different families within Sylvioidea, which are still largely unresolved. However, if, as seems likely to be the case based on the presently available data, the lack of resolution results mainly from rapid divergence of the primary lineages (families), these relationships might remain obscure.
The total number of species within Sylvioidea has increased much in recent years, mainly due to studies applying molecular markers and/or vocalizations and other life-history data, and in many groups multiple cryptic species have been detected. It seems likely that the number of species is going to continue to increase, as new groups are being studied with modern integrative methods (using a combination of morphological, vocal, genetic, behavioural and other data). In particular, some polytypic species of larks, "warblers", bulbuls, and "babblers" are likely to comprise considerably more species than presently recognized. As good taxonomic assessments are a fundamental basis for many evolutionary studies, as well as for conservation purposes, continued efforts to revise the species-level taxonomy within Sylvioidea are much needed.
Acknowledgements
We are grateful to Nigel Collar, Silke Fregin, Margaret Koopman/Niven Library at the Percy FitzPatrick Institute of African Ornithology, Peter Kennerley, Lars Larsson, David Pearson, He Peng, Pamela Rasmussen, Craig Robson and an anonymous reviewer for comments on the manuscript or other assistance; and the Chinese Academy of Sciences Visiting Professorship for Senior International Scientists (No. 2011T2S04) to P.A., and NSFC (No. 31010103901) to F.L. and P.A.
Aleixo A, Pacheco JF. 2006. A family name for the monotypic oscine passerine genus Donacobius. Rev Brasil Ornitol, 14: 172–173.
Alström P, Barnes KN, Olsson U, Barker FK, Bloomer P, Khan AA, Qureshi MA, Guillaumet A, Crochet P-A, Ryan PG. 2013. Multilocus phylogeny of the avian family Alaudidae (larks) reveals complex morphological evolution, non-monophyletic genera and hidden species diversity. Mol Phylogenet Evol. doi: .
Alström P, Davidson P, Duckworth JW, Eames JC, Le TT, Nguyen C, Olsson U, Robson C, Timmins RJ. 2010. Description of a new species of Phylloscopus warbler from Vietnam and Laos. Ibis, 152: 145–168.
Alström P, Fregin S, Norman JA, Ericson PGP, Christidis L, Olsson U. 2011a. Multilocus analysis of a taxonomically densely sampled dataset reveal extensive non- monophyly in the avian family Locustellidae. Mol Phylogenet Evol, 38: 381–397.
Alström P, Höhna S, Gelang M, Ericson PGP, Olsson U. 2011b. Non-monophyly and intricate morphological evolution within the avian family Cettiidae revealed by multilocus analysis of a taxonomically densely sampled dataset. BMC Evol Biol, 11: 352.
Alström P, Fjeldså J, Fregin S, Olsson U. 2011c. Gross morphology betrays phylogeny: the Scrub Warbler Scotocerca inquieta is not a cisticolid. Ibis, 153: 87–97.
Alström P, Saitoh T, Williams D, Nishiumi I, Shigeta Y, Ueda K, Irestedt M, Björklund M, Olsson U. 2011d. The Arctic Warbler Phylloscopus borealis – three anciently separated cryptic species revealed. Ibis, 153: 395–410.
Alström P, Olsson U, Colston P. 1997. Re-evaluation of the taxonomic status of Phylloscopus proregulus kansuensis Meise. Bull Br Ornithol Club, 117: 177–193.
Alström P, Olsson U, Colston PR. 1990. Description of a possible new species of leaf warbler of the genus Phylloscopus from China. Bull Br Ornithol Club, 110: 43–47.
Alström P, Olsson U, Colston PR. 1992. A new species of Phylloscopus warbler from central China. Ibis, 134: 329–334.
Alström P, Olsson U, Rasmussen PC, Yao C-T, Ericson PGP, Sundberg P. 2007. Morphological, vocal and genetic divergence in the Cettia acanthizoides complex (Aves: Cettiidae). Zool J Linn Soc, 149: 437–452.
Alström P, Olsson U, Round PD. 1991. The taxonomic status of Acrocephalus agricola tangorum. Forktail, 6: 3–13.
Alström P, Olsson U. 1990. Taxonomy of the Phylloscopus proregulus complex. Bull Br Ornithol Club, 110: 38–43.
Alström P, Olsson U. 1992. Taxonomic status of Phylloscopus affinis and P. subaffinis. Bull Br Ornithol Club, 112: 111–126.
Alström P, Olsson U. 1995. A new species of Phylloscopus warbler from Sichuan Province, China. Ibis, 137: 459–468.
Alström P, Olsson U. 1999. The Golden-spectacled Warbler: a complex of sibling species, including a previously undescribed species. Ibis, 141: 545–568.
Alström P, Rasmussen PC, Olsson U, Sundberg P. 2008. Species delimitation based on multiple criteria: the Spotted Bush Warbler Bradypterus thoracicus complex (Aves, Megaluridae). Zool J Linn Soc, 154: 291–307.
Alström P. 1998. Taxonomy of the Mirafra assamica complex. Forktail, 13: 97–107.
Alström P. Ericson PGP, Olsson U, Sundberg P. 2006. Phylogeny and classification of the avian superfamily Sylvioidea. Mol Phylogenet Evol, 38: 381–397.
Athreya R. 2006. A new species of Liocichla (Aves: Timaliidae) from Eaglenest Wildlife Sanctuary, Arunachal Pradesh, India. Indian Birds, 2: 82–94.
Ayé R, Hertwig ST, Schweizer M. 2010. Discovery of a breeding area of the enigmatic large-billed reed warbler Acrocephalus orinus. J Avian Biol, 41: 452–459.
Babin MJ. 2005. Geographic variation in rough-winged swallows (Aves: Hirundinidae: Stelgidopteryx. M.S. thesis. Louisiana State University and Agricultural and Mechanical College, Louisiana, USA.
Bairlein F, Alström P, Aymí R, Clement P, Dyrcz A, Gargallo G, Hawkins F, Madge S, Pearson D, Svensson L. 2006. Family Sylviidae (Warblers). In: del Hoyo J, Elliott A, Christie DA (eds) Handbook of the Birds of the World. Volume 12. Lynx Edicions, Barcelona, pp 492–709.
Baker K. 1997. Warblers of Europe, Asia and North Africa. A & C Black, London.
Bannerman DA, Bates GL. 1924. On the birds collected in Northwestern and Northern Cameroon and parts of northern Nigeria – Part Ⅱ. Systematic Notes. Ibis, 66: 200–276.
Bannerman DA. 1948. The Birds of Tropical West Africa. Vol. 6. Crown Agents, London.
Barhoum DN, Burns KJ. 2002. Phylogenetic relationships of the Wrentit based on mitochondrial cytochrome b sequences. Condor, 104: 740–749.
Barker FK, Barrowclough GF, Groth JG. 2002. A phylogenetic hypothesis for passerine birds: taxonomic and biogeographic implications of an analysis of nuclear DNA sequence data. Proc R Soc Lond B, 269: 295–308.
Barker FK, Cibois A, Schikler P, Feinstein J, Cracraft J. 2004. Phylogeny and diversification of the largest avian radiation. Proc Natl Acad Sci USA, 101: 11040–11045.
Bates GL. 1930. Handbook of the Birds of West Africa. Bale, Sons and Danielson, London.
Beecher WJ. 1953. A phylogeny of the oscines. Auk, 70: 270–333.
Bensch S, Helbig AJ, Salomon M, Seibold I. 2002. Amplified fragment length polymorphism analysis identifies hybrids between two subspecies of warblers. Mol Ecol, 11: 473–481.
Bensch S, Pearson D. 2002. The Large-billed Reed Warbler Acrocephalus orinus revisited. Ibis, 144: 259–267.
Beresford P, Barker FK, Ryan PG, Crowe TM. 2005. African endemics span the tree of songbirds (Passeri): molecular systematics of several evolutionary 'enigmas'. Proc Roy Soc Lond B, 272: 849–858.
BirdLife International. 2000. Threatened Birds of the World. Lynx Edicions and BirdLife International, Barcelona and Cambridge, UK.
Bock WJ. 1994. History and nomenclature of avian family-group names. Bull Amer Mus Nat Hist, 222: 1–283.
Brambilla M, Vitulano S, Spina F, Baccetti N, Gargallo G, Fabbri E, Guidali F, Randi E. 2008a. A molecular phylogeny of the Sylvia cantillans complex: cryptic species within the Mediterranean basin. Mol Phylogenet Evol, 48: 461–472.
Brambilla M, Janni O, Guidali F, Sorace A. 2008b. Song perception among incipient species as a mechanism for reproductive isolation. J Evol Biol, 21: 651–657.
Brambilla M, Quaglierini A, Reginato F, Vitulano S, Guidali F. 2008c. Syntopic taxa in the Sylvia cantillans species complex. Acta Ornithol, 43: 217–220.
Brosset A, Erard C. 1986. Les Oiseaux des Régions Forestiêres du Gabon, Vol. 1. Société National de Protection de la Nature, Paris
Cabanis J. 1875. [Neue Arten]. J Ornithol, 23: 234–238.
Cerasale DJ, Dor R, Winkler DW, Lovette IJ. 2012. Phylogeny of the Tachycineta genus of new world swallows: insights from complete mitochondrial genomes. Mol Phylogenet Evol, 63: 64–71.
Chapin JP. 1953. The birds of the Belgian Congo. Part Ⅲ. Bull AMNH, 75A: 1–826.
Chappuis C. 1974. Illustration sonore de problèmes bioacoustiques posés par les oiseaux de la zone éthiopienne. Alauda, 42: 197–222.
Cheng T-H. 1987. A Synopsis to the Avifauna of China. Parey Scientific Publ., Hamburg, Germany.
Chikuni K, Minaka N, Ikenaga H. 1996. Molecular phylogeny of some Passeriformes, based on cytochrome b sequences. J Yamashina Inst Orn, 28: 1–8.
Cibois A, Beadell JS, Graves GR, Pasquet E, Slikas B, Sonsthagen SA, Thibault J-C, Fleischer RC. 2011a. Charting the course of reed-warblers across the Pacific islands. J Biogeogr, 38: 1963–1975.
Cibois A, Cracraft J. 2004. Assessing the passerine "Tapestry": phylogenetic relationships of the Muscicapoidea inferred from nuclear DNA sequences. Mol Phylogenet Evol, 32: 264–273.
Cibois A, Kalyakin MV, Han LX, Pasquet E. 2002. Molecular phylogenetics of babblers (Timaliidae): reevaluation of the genera Yuhina and Stachyris. J Avian Biol, 33: 380–390
Cibois A, Normand D, Gregory SMS, Pasquet E. 2010. Bernieridae (Aves: Passeriformes): a family-group name for the Malagasy sylvioid radiation. Zootaxa, 2554: 65–68.
Cibois A, Pasquet E, Schulenberg TS. 1999. Molecular systematics of the Malagasy babblers (Passeriformes: Timaliidae) and warblers (Passeriformes: Sylviidae), based on cytochrome b and 16S rRNA sequences. Mol Phylogenet Evol, 13: 581–595.
Cibois A, Slikas B, Schulenberg TS, Pasquet E. 2001. An endemic radiation of Malagasy songbirds is revealed by mitochondrial DNA sequence data. Evolution, 55: 1198–1206.
Cibois A, Thibault J-C, Pasquet E. 2007. Uniform phenotype conceals double colonization by reed-warblers of a remote Pacific archipelago. J Biogeogr, 34: 1155–1166.
Cibois A, Thibault J-C, Pasquet E. 2008. Systematics of the extinct reed warblers Acrocephalus of the Society Islands of eastern Polynesia. Ibis, 150: 365–376.
Cibois A, Thibault J-C, Raust P, Pasquet E. 2011b. Systematics of the reed-warblers (Acrocephalus atyphus) of the Tuamotu archipelago, Eastern Polynesia. Emu, 111: 139–147.
Cibois A. 2003a. Mitochondrial DNA phylogeny of babblers (Timaliidae). Auk, 120: 35–54.
Cibois A. 2003b. Sylvia is a babbler: taxonomic implications for the families Sylviidae and Timaliidae. Bull Br Ornithol Club, 123: 257–261.
Clancey PA. 1966. Subspeciation in southern African populations of Sabota Lark Mirafra sabota Smith. Ostrich, 37: 207–213.
Clancey PA. 1989. Taxonomic and distributional findings on some birds from Namibia. Cimbebasia, 11: 111–133.
Collar NJ, Robson C. 2007. Family Timaliidae (Babblers). In: del Hoyo J, Elliott A, Christie DA (eds) Handbook of the Birds of the World. Vol. 12. Picathartes to Tits and Chickadees. Barcelona: Lynx Edicions, pp 70–291.
Collar NJ. 2006. A partial revision of the Asian babblers (Timaliidae). Forktail, 22: 85–112.
Collar NJ. 2011. Taxonomic notes on some Asian babblers (Timaliidae). Forktail, 27: 100–102.
Cracraft J, Barker FK, Braun M, Harshman J, Dyke GJ, Feinstein J, Stanley S, Cibois A, Schikler P, Beresford P, García-Moreno J, Sorenson MP, Yuri T, Mindell DP. 2004. Phylogenetic relationships among modern birds (Neornithes). Toward an avian tree of life. In: Cracraft J, Donoghue MJ (eds), Assembling the Tree of Life. Oxford University Press, Oxford.
Cracraft J. 1989. Speciation and its ontology: the empirical consequences of alternative species concepts for understanding patterns and processes of differentiation. In: Otte D, Endler JA (eds) Speciation and Its Consequences. Sinauer, Sunderland, Mass, pp 28–59.
Cramp S. 1992. The Birds of the Western Palearctic, Vol. 7. Oxford University Press, Oxford.
Dai C, Chen K, Zhang R, Yang X, Yin Z, Tian H, Zhang Z, Hu Y, Lei F. 2010. Molecular phylogenetic analysis among species of Paridae, Remizidae and Aegithalos based on mtDNA sequences of COI and cyt b. Chinese Birds, 1: 112–123.
Dai C, Wang W, Lei F. 2013. Multilocus phylogeography (mitochondrial, autosomal and Z-chromosomal loci) and genetic consequences of long-distance male dispersal in Black-throated tits (Aegithalos concinnus). Heredity, 110: 457–465.
Dai C, Zhao N, Wang W, Lin C, Gao B, Yang X, Zhang Z, Lei F. 2011. Profound climatic effects on two East Asian BlackThroated Tits (Aves: Aegithalidae), revealed by ecological niche models and phylogeographic analysis. PLoS ONE, 6(12): e29329.
de Juana E, Suárez F, Ryan P, Alström P, Donald P. 2004. Family Alaudidae (Larks). In: del Hoyo J, Elliott A, Christie DA (eds) Handbook of the Birds of the World, Vol. 9. Lynx Edicions, Barcelona, pp 496–601.
Dean WRJ, Fry CH, Keith S, Lack P. 1992. Family Alaudidae: Larks. In: Keith S, Urban EK, Fry CH (eds) The Birds of Africa, Vol. 4. Academic Press, London, pp 13–124.
Degnan SM, Moritz C. 1992. Phylogeography of mitochondrial DNA in two species of white-eyes in Australia. Auk, 109: 800–811.
Deignan H. 1964a. Subfamily Panurinae, parrotbills. In: Mayr E, Paynter RA (eds) Check-list of Birds of the World, Vol. X. Museum of Comparative Zoology, Cambridge, Mass, pp 430–442.
Deignan H. 1964b. Subfamily Timaliinae, babblers. In: Mayr E, Paynter RA (eds). Check-list of Birds of the World, Vol. X. Museum of Comparative Zoology, Cambridge, Mass, pp 240–427.
del Hoyo J, Elliott A, Christie DA. 2006. Handbook of the Birds of the World. Old World Flycatchers to Old World Warblers, Vol. 11. Lynx Edicions, Barcelona.
Delacour J. 1942–1943. The bush-warblers of the genera Cetti and Bradypterus, with notes on allied genera and species. Ibis, 6: 509–519.
Delacour J. 1943. A revision of the genera and species of the family Pycnonotidae (bulbuls). Zoologica, 28: 17–28.
Delacour J. 1946. Les timaliinés. L'Oiseau, 16: 7–36.
Dickinson E. 2003. The Howard and Moore Complete Checklist of the Birds of the World. Third ed. Helm, London.
Dickinson EC, Dekker RWRJ. 2001. Systematic notes on Asian birds. 11. A preliminary review of the Alaudidae. Zool Verh Leiden, 335: 61–84.
Dickinson EC, Rasmussen PC, Round PD, Rozendaal FG. 2000. Systematic notes on Asian birds. 1. A review of the russet bush-warbler Bradypterus seebohmi (Ogilvie-Grant, 1895). Zool Verh Leiden, 331: 12–64.
Dong F, Li S-H, Yang X-J. 2010a. Molecular systematics and diversification of the Asian scimitar babblers (Timaliidae, Aves) based on mitochondrial and nuclear DNA sequences. Mol Phylogenet Evol, 57: 1268–1275.
Dong F, Wu F, Liu LM, Yang XJ. 2010b. Molecular phylogeny of the barwings (Aves: Timaliidae: Actinodura), a paraphyletic group, and its taxonomic implications. Zool Stud, 49: 703–709.
Dor R, Carling MD, Lovette IJ, Sheldon FH, Winkler DW. 2012. Species trees for the tree swallows (Genus Tachycineta): An alternative phylogenetic hypothesis to the mitochondrial gene tree. Mol Phylogenet Evol, 65: 317–322.
Dor R, Safran RJ, Sheldon FH, Winkler DW, Lovette IJ. 2010. Phylogeny of the genus Hirundo and the Barn Swallow subspecies complex. Mol Phylogenet Evol, 56: 409–418.
Dowsett RJ, Dowsett-Lemaire F. 1993. A contribution to the distribution and taxonomy of Afrotropical and Malagasy birds. Tauraco Research Reports 5. Tauraco Press, Liège.
Dowsett RJ, Olson SL, Roy MS, Dowsett-Lemaire F. 1999. Systematic status of the Black-collared Bulbul Neolestes torquatus. Ibis, 141: 22–28.
Dowsett-Lemaire F, Dowsett RJ. 1987. European and African Reed Warblers, Acrocephalus scirpaceus and A. baeticatus: vocal and other evidence for a single species. Bull Br Ornithol Club, 107: 74–85.
Drovetski SV, Zink RM, Fadeev IV, Nesterov EV, Koblik EA, Red'kin YA, Rohwer S. 2004. Mitochondrial phylogeny of Locustella and related genera. J Avian Biol, 35: 105–110.
Eck S. 1994. Die geographisch-morphologische Vikarianz der grossen palaearktischen Rohrsänger (Aves: Passeriformes: Sylviidae: Acrocephalus[arundinaceus]). Zool Abh Staatl Mus Tierkd Dresden, 48: 161–168.
Ericson PGP, Johansson US. 2003. Phylogeny of Passerida (Aves: Passeriformes) based on nuclear and mitochondrial sequence data. Mol Phylogenet Evol, 29: 126–138.
Fjeldså J, Goodman SM, Schulenberg TS, Slikas B. 1999. Molecular evidence for relationships of Malagasy birds. In: Adams NJ, Slotow RH (eds) Proceedings of the 22nd International Ornithological Congress, Durban. BirdLife South Africa, Johannesburg, pp 3084–3094.
Fjeldså J, Irestedt M, Ericson PGP, Zuccon D. 2010. The Cinnamon Ibon Hypocryptadius cinnamomeus is a forest canopy sparrow. Ibis, 152: 747–760.
Fregin S, Haase M, Olsson U, Alström P. 2009. Multi-locus phylogeny of the family Acrocephalidae (Aves: Passeriformes) –The traditional taxonomy overthrown. Mol Phylogenet Evol, 52: 866–878.
Fregin S, Haase M, Olsson U, Alström P. 2012. New insights into family relationships within the avian superfamily Sylvioidea (Passeriformes) based on seven molecular markers. BMC Evol Biol, 12: 157.
Fry CH, Williamson K, Ferguson-Lees IJ. 1974. A new subspecies of Acrocephalus baeticatus from Lake Chad and a taxonomic reappraisal of Acrocephalus dumetorum. Ibis, 116: 340–346.
Fuchs J, Fjeldså J, Bowie RCK, Voelker G, Pasquet E. 2006. The African warbler genus Hyliota as a lost lineage in the oscine songbird tree: molecular support for an African origin of the Passerida. Mol Phylogenet Evol, 39: 186–197.
Fuchs J, Fjeldså J, Bowie RCK. 2011. Diversification across an altitudinal gradient in the Tiny Greenbul (Phyllastrephus debilis) from the Eastern Arc Mountains of Africa. BMC Evol Biol, 11: 117.
García JT, Suárez F, Garza V, Calero-Riestra M, Hernández J, Pérez-Tris J. 2008. Genetic and phenotypic variation among geographically isolated populations of the globally threatened Dupont's lark Chersophilus duponti. Mol Phylogenet Evol, 46: 237–251.
Gavrilov EI, Savchenko AP. 1991. On species validity of the Pale Sand Martin (Riparia diluta Sharpe et Wyatt, 1893). Byul Mosk O-va Isp Prir Otd Biol, 96: 34–44. (in Russian)
Gelang M, Cibois A, Pasquet E, Olsson U, Alström P, Ericson PGP. 2009. Phylogeny of babblers (Aves, Passeriformes): major lineages, family limits and classification. Zool Scr, 32: 279–296.
Gill F, Donsker D. 2013. IOC World Bird List (version 3.3). . Accessed 5 March 2013.
Goroshko OA. 1993. Taxonomic status of the pale (sand?) martin Riparia (riparia?) diluta (Sharpe & Wyatt, 1893). Russ Ornit Zhurnal, 2: 303–323. (in Russian)
Guillaumet A, Crochet PA, Godelle B. 2005. Phenotypic variation in Galerida larks in Morocco: the role of history and natural selection. Mol Ecol, 14: 3809–3821.
Guillaumet A, Crochet PA, Pons J-M. 2008. Climate-driven diversification in two widespread Galerida larks. BMC Evol Biol, 8: 32.
Guillaumet A, Pons JM, Godelle B, Crochet PA. 2006. History of the Crested Lark in the Mediterranean region as revealed by mtDNA sequences and morphology. Mol Phylogenet Evol, 39: 645–656.
Haffer J. 1991. Familie Sylviidae-Zweigsänger (Grasmücken und Verwandte). In: von Blotzheim GUN, Bauer KM (eds) Handbuch der Vögel Mitteleuropas, Vol. 12. Aula-Verlag, Wiesbaden, pp 11–21 and 208–216.
Hall BP, Moreau RE. 1970. An Atlas of Speciation in African Passerine Birds. Trustees of the British Museum (Natural History), London.
Hamao S, Veluz MJS, Saitoh T, Nishiumi I. 2008. Phylogenetic relationship and song differences between closely related bush warblers (Cettia seebohmi and C. diphone). Wilson J Ornithol, 120: 268–276.
Hanotte O, Knox AG, Prigogine A. 1987. The taxonomic status of Nicator: Evidence from feather protein electrophoresis. Biochem Syst Ecol, 15: 629–634.
Harrison CJO. 1966. The validity of some genera of larks (Alaudidae). Ibis, 108: 573–583.
Hartert E. 1909. Die Vögel der Paläarktischen Fauna. Verlag von R. Friedländer und Sohn, Berlin.
Helbig AJ, Martens J, Seibold I, Henning F, Schottler B, Wink M. 1996. Phylogeny and species limits in the Palaearctic chiffchaff Phylloscopus collybita complex: mitochondrial genetic differentiation and bioacoustic evidence. Ibis, 138: 650–666.
Helbig AJ, Salomon M, Bensch S, Seibold I. 2001. Male-biased gene flow across an avian hybrid zone: evidence from mitochondrial and microsatellite DNA. J Evol Biol, 14: 277–287.
Helbig AJ, Seibold I, Martens J, Wink M. 1995. Genetic differentiation and phylogenetic relationships of Bonelli's Warbler Phylloscopus bonelli and Green Warbler Phylloscopus nitidus. J Avian Biol, 26: 139–153.
Helbig AJ, Seibold I. 1999. Molecular phylogeny of Palearctic–African Acrocephalus and Hippolais warblers (Aves: Sylviidae). Mol Phylogenet Evol, 11: 246–260.
Holyoak DT, Thibault J-C. 1984. Contribution a l'eìtude des oiseaux de Polyneìsie orientale. Meìm. MNHN, Paris, 127: 1–209.
Inskipp T, Lindsey N, Duckworth W. 1996. An Annotated Checklist of the Birds of the Oriental Region. Oriental Bird Club, Sandy, Bedfordshire
International Commission on Zoological Nomenclature (ICZN). 1999. International Code of Zoological Nomenclature. Fourth Edition. The International Trust for Zoological Nomenclature, Padova.
Irestedt M, Gelang M, Olsson U, Ericson PGP, Sangster G, Alström P. 2011. Neumann's Warbler Hemitesia neumanni (Sylvioidea): the sole African member of a Palaeotropic Miocene radiation. Ibis, 153: 78–86.
Irwin DE, Alström P, Olsson U, Benowitz-Fredericks ZM. 2001a. Cryptic species in the genus Phylloscopus (Old World leaf warblers). Ibis, 143: 233–247.
Irwin DE, Bensch S, Irwin JH, Price TD. 2005. Speciation by distance in a ring species. Science, 307: 414–416.
Irwin DE, Bensch S, Price TD. 2001b. Speciation in a ring. Nature, 409: 333–337.
Irwin DE, Thimgan MP, Irwin JH. 2008. Call divergence is correlated with geographic and genetic distance in greenish warblers (Phylloscopus trochiloides): a strong role for stochasticity in signal evolution? J Evol Biol, 21: 435–448.
Irwin DE. 2000. Song variation in an avian ring species. Evolution, 54: 998–1010.
Johansson US, Fjeldså J, Lokugalappatti LGS, Bowie RCK. 2007a. A nuclear DNA phylogeny and proposed taxonomic revision of African greenbuls (Aves, Passeriformes, Pycnonotidae). Zool Scr, 36: 417–427.
Johansson US, Alström P, Olsson U, Ericson PGP, Sundberg P, Price TD. 2007b. Build-up of the Himalayan avifauna through immigration: a biogeographical analysis of the Phylloscopus and Seicercus warblers. Evolution, 61: 324–333.
Johansson US, Fjeldså J, Bowie RCK. 2008a. Phylogenetic relationships within Passerida (Aves: Passeriformes). Mol Phylogenet Evol, 48: 858–876.
Johansson US, Bowie RCK, Hackett SJ, Schulenberg TS. 2008b. The phylogenetic affinities of Crossley's babbler (Mystacornis crossleyi): adding a new niche to the vanga radiation of Madagascar. Biol Lett, 4: 677–680.
Jones AW, Kennedy RS. 2008. Evolution in a tropical archipelago: comparative phylogeography of Philippine fauna and flora reveals complex patterns of colonization and diversification. Biol J Linn Soc, 95: 620–639.
Jønsson KA, Fjeldså J, Ericson PGP, Irestedt M. 2007. Systematic placement of an enigmatic Southeast Asian taxon Eupetes macrocerus and implications for the biogeography of a main songbird radiation, the Passerida. Biol Lett, 3: 323–326.
Jønsson KA, Fjeldså J. 2006. A phylogenetic supertree of oscine passerine birds (Aves: Passeri). Zool Scr, 35: 149–186.
Keith S. 1997. Genus Hylia Cassin, Hylia prasina (Cassin), Green Hylia. In: Urban EK, Fry CH, Keith S (eds) The Birds of Africa, Vol. 5. Academic Press, London, pp 428–430.
Kennerley P, Pearson D. 2010. Reed and Bush Warblers. Christopher Helm, London.
King B, Dickinson EC. 1975. A Field Guide to the Birds of SouthEast Asia. Collins, London.
King B, Robson C. 2008. The taxonomic status of the three subspecies of Greater Rufous-headed Parrotbill Paradoxornis ruficeps. Forktail, 24: 120–122.
King B. 1989. The avian genera Tesia and Urosphena. Bull Br Ornithol Club, 109: 162–166.
Kirchman JJ, Whittingham LA, Sheldon FH. 2000. Relationships among Cave Swallow Populations (Petrochelidon fulva) determined by comparisons of microsatellite and cytochrome b data. Mol Phylogenet Evol, 14: 107–121.
Knox AG, Collinson M, Helbig AJ, Parkin DT, Sangster G. 2002. Taxonomic recommendations for British birds. Ibis, 144: 707–710.
Koblik EA, Red'kin YA, Meer MS, Derelle R, Golenkina SA, Kondrashov FA, Arkhipov V. 2011. Acrocephalus orinus: A case of mistaken identity. PLoS ONE, 6(4): e17716.
Lawson WJ. 1961. The races of the Karoo Lark, Certhilauda albescens (Lafresnaye). Ostrich, 32: 64–74.
Leader PJ, Carey GJ, Olsson U, Baral HS, Alström P. 2010. The taxonomic status of Rufous-rumped Grassbird Graminicola bengalensis, with comments on its distribution and status. Forktail, 26: 121–126.
LeCroy M, Barker FK. 2006. A new species of bush-warbler from Bougainville Island and a monophyletic origin for Southwest Pacific Cettia. Am Mus Novit, 3511: 1–20.
Leisler B, Heidrich P, Schulze-Hagen K, Wink M. 1997. Taxonomy and phylogeny of reed warblers (genus Acrocephalus) based on mtDNA sequences and morphology. J Ornithol, 138: 469–496.
Lewis PO, Holder MT, Holsinger KE. 2005. Polytomies and Bayesian phylogenetic inference. Syst Biol, 54: 241–253.
Li SH, Li JW, Han LX, Yao CT, Shi HT, Lei FM, Yen CW. 2006. Species delimitation in the Hwamei Garrulax canorus. Ibis, 148: 698–706.
Li SH, Yeung CKL, Feinstein J, Han LX, Le MH, Wang CX, Ding P. 2009. Sailing through the Late Pleistocene: unusual historical demography of an East Asian endemic, the Chinese Hwamei (Leucodioptron canorum canorum), during the last glacial Period. Mol Ecol, 18: 622–633.
Liu X, Ding Z, Fang W, Lin W, Cai M, Yan C. 2010. The Avifauna of Taiwan. Taiwan Forestry Bureau, Taipei. (in Chinese)
Loskot VM. 2001. Taxonomic revision of the Hume's Whitethroat Sylvia althaea Hume, 1878. Avian Ecol Behav, 6: 41–42.
Loskot VM. 2005. Morphological variation and taxonomic revision of five south-eastern subspecies of Lesser Whitethroat Sylvia curruca (L. ) (Aves: Sylviidae). Zool Mededelingen Leiden, 79: 157–165.
Luo X, Qu YH, Han LX, Li SH, Lei FM. 2009. A phylogenetic analysis of laughingthrushes (Timaliidae: Garrulax) and allies based on mitochondrial and nuclear DNA sequences. Zool Scr, 38: 9–22.
Macdonald JD. 1952a. Notes on the Long-bill Lark Certhilauda curvirostris. Ibis, 94: 122–127.
Macdonald JD. 1952b. Notes on the taxonomy of the clapper larks of South Africa. Ibis, 94: 629–635.
Macdonald JD. 1953. Taxonomy of the Karoo and Red-back Larks of western South Africa. Bull Br Mus Nat Hist Zool, 1: 319–350.
Mackworth-Praed CW, Grant CHB. 1941. A survey of the African species placed in the genus Bradypterus. Ibis, 83: 441–455.
Maclean GL. 1969. South African Lark genera. Cimbebasia A, 1: 79–94.
Marks BD. 2010. Are lowland rainforests really evolutionary museums? Phylogeography of the green hylia (Hylia prasina) in the Afrotropics. Mol Phylogen Evol, 55: 178–184.
Martens J, Eck S, Päckert M, Sun Y-H. 1999. The Golden-spectacled Warbler Seicercus burkii – a species swarm (Aves: Passeriformes: Sylviidae), part 1. Zool Abhandl Mus Dresden, 50: 281–327.
Martens J, Steil B. 1997. Reviergesänge und Speziesdifferenzierung in der Klappergrasmücken-Gruppe Sylvia[curruca]. J Ornithol, 138: 1–23.
Martens J, Sun Y-H, Päckert M. 2008. Intraspecific differentiation of Sino-Himalayan bush-dwelling Phylloscopus leaf warblers, with description of two new taxa. Vert Zool, 58: 233–265.
Martens J, Tietze DT, Eck S, Veith M. 2004. Radiation and species limits in the Asian Pallas's warbler complex (Phylloscopus proregulus s. l. ). J Ornithol, 145: 206–222.
Martens J. 1988. Phylloscopus borealoides Portenko — ein verkannter Laubsänger der Ost-Paläarktis. J Ornithol, 129: 343–351.
Martens J. 2010. A preliminary review of the leaf warbler genera Phylloscopus and Seicercus. Br Orn Club Occas Publs, 5: 41–116.
Mayr E, Amadon D. 1951. A classification of recent birds. Amer Mus Novitat, 1496: 1–42.
Mayr E, Bond J. 1943. Notes on the generic classification of the swallows Hirundinidae. Ibis, 85: 334–341.
Mayr E, Greenway Jr JC. 1960. Check-list of Birds of the World, Vol. IX. Museum of Comparative Zoology, Cambridge, MA.
Mayr E. 1948. Geographic variation in the Reed-Warbler. Emu, 47: 205–210.
Meinertzhagen R. 1951. A review of the Alaudidae. Proc Zool Soc Lond, 121: 81–132.
Milá B, Warren BH, Heeb P, Thébaud C. 2010. The geographic scale of diversification on islands: genetic and morphological divergence at a very small spatial scale in the Mascarene grey white-eye (Aves: Zosterops borbonicus). BMC Evol Biol, 10: 158.
Morioka H, Shigeta Y. 1993. Generic allocation of the Japanese Marsh Warbler Megalurus pryeri (Aves: Sylviidae). Bull Nat Sci Mus Tokyo A, 19: 37–43. (in Japanese)
Morony JJ, Bock WJ, Farrand J. 1975. Reference List of the Birds of the World. American Museum of Natural History, New York.
Moyle RG, Andersen MJ, Oliveros CH, Steinheimer F, Reddy S. 2012. Phylogeny and biogeography of the core babblers (Aves: Timaliidae). Syst Biol, 61: 631–651
Moyle RG, Filardi CE, Smith CE, Diamond JM. 2009. Explosive Pleistocene diversification and hemispheric expansion of a "great speciator". Proc Natl Acad Sci USA, 106: 1863–1868.
Moyle RG, Marks BD. 2006. Phylogenetic relationships of the bulbuls (Aves: Pycnonotidae) based on mitochondrial and nuclear DNA sequence data. Mol Phylogenet Evol, 40: 687–695.
Moyle RG, Slikas B, Whittingham LA, Winkler DW, Sheldon FH. 2008. DNA sequence assessment of phylogenetic relationships among New World martins (Hirundinidae: Progne). Wilson J Ornithol, 120: 683–691.
Nguembock B, Cruaud C, Denys C. 2012. A large evaluation of passerine cisticolids (Aves: Passeriformes): more about their phylogeny and diversification. Open Ornithol J, 5: 42–56.
Nguembock B, Fjeldså J, Couloux A, Cruaud C, Pasquet E. 2008. Polyphyly of the genus Apalis and a new generic name for the species pulchra and ruwenzorii. Ibis, 150: 756–765.
Nguembock B, Fjeldså J, Tillier A, Pasquet E. 2007. A phylogeny for the Cisticolidae (Aves: Passeriformes) based on nuclear and mitochondrial DNA sequence data, and a re-interpretation of a unique nest-building specialization. Mol Phylogenet Evol, 42: 272–286.
Oatley G, Bowie RCK, Crowe TM. 2011. The use of subspecies in the systematics of southern African white-eyes: historical entities or eco-geographic variants. J Zool, 284: 21–30.
Oatley G, Voelker G, Crowe TM, Bowie RC. 2012. A multi-locus phylogeny reveals a complex pattern of diversification related to climate and habitat heterogeneity in southern African white-eyes. Mol Phylogenet Evol, 64: 633–644.
Oliveros CH, Moyle RG. 2010. Origin and diversification of Philippine bulbuls. Mol Phylogenet Evol, 54: 822–832.
Oliveros CH, Reddy S, Moyle RG. 2012. The phylogenetic position of some Philippine "babblers" spans the muscicapoid and sylvioid bird radiations. Mol Phylogenet Evol, 65: 799–804.
Olsson U, Alström P, Colston PR. 1993. A new species of Phylloscopus warbler from Hainan Island, China. Ibis, 135: 2–7.
Olsson U, Alström P, Ericson PGP, Sundberg P. 2005. Nonmonophyletic taxa and cryptic species – evidence from a molecular phylogeny of leaf-warblers (Phylloscopus, Aves). Mol Phylogenet Evol, 36: 261–276.
Olsson U, Alström P, Gelang M, Ericson PGP, Sundberg P. 2006. Phylogeography of Indonesian and Sino-Himalayan region bush warblers (Cettia, Aves). Mol Phylogenet Evol, 41: 556–565.
Olsson U, Alström P, Sundberg P. 2004. Non-monophyly of the avian genus Seicercus (Aves: Sylviidae) revealed by mitochondrial DNA. Zool Scr, 33: 501–510.
Olsson U, Irestedt M, Sangster G, Ericson PGP, Alström P. 2013a. Systematic revision of the avian family Cisticolidae based on a multi-locus phylogeny of all genera. Mol Phylogenet Evol, 66: 790–799.
Olsson U, Leader PJ, Carey GJ, Khan AA, Svensson L, Alström P. 2013b. New insights into the intricate taxonomy and phylogeny of the Sylvia curruca complex. Mol Phylogenet Evol, 67: 72–85.
Ottosson U, Bensch S, Svensson L, Waldenström J. 2005. Differentiation and phylogeny of the olivaceous warbler Hippolais pallida species complex. J Ornithol, 146: 127–136.
Päckert M, Blume C, Sun Y-H, Liang W, Martens J. 2009. Acoustic differentiation reflects mitochondrial lineages in Blyth's leaf warbler and white-tailed leaf warbler complexes (Aves: Phylloscopus reguloides, Phylloscopus davisoni). Biol J Linn Soc, 96: 584–600.
Päckert M, Martens J, Liang W, Hsu Y-L, Sun Y-H. 2012. Molecular genetic and bioacoustic differentiation of Pnoepyga WrenBabblers. J Ornithol, 154: 329–337.
Päckert M, Martens J, Sun Y-H, Veith M. 2004. The radiation of the Seicercus burkii complex and its congeners (Aves: Sylviidae): molecular genetics and bioacoustics. Org Div Evol, 4: 341–364.
Päckert M, Martens J, Sun Y-H. 2010. Phylogeny of long-tailed tits and allies inferred from mitochondrial and nuclear markers (Aves: Passeriformes, Aegithalidae). Mol Phylogenet Evol, 55: 952–967.
Parkin DT, Collinson M, Helbig AJ, Knox AG, Sangster G, Svensson L. 2004. Species limits in Acrocephalus and Hippolais warblers from the Western Palaearctic. Brit Birds, 97: 276–299.
Pasquet E, Bourdon E, Kalyakin MV, Cibois A. 2006. The fulvettas (Alcippe, Timaliidae, Aves): a polyphyletic group. Zool Scr, 35: 559–566.
Pasquet E, Han L-X, Khobket O, Cibois A. 2001. Towards a molecular systematics of the genus Criniger, and a preliminary phylogeny of the bulbuls (Aves, Passeriformes, Pycnonotidae). Zoosystema, 23: 857–863.
Pätzold R. 2003. Kompendium der Lerchen Alaudidae. Jan Schimkat Medienpublizistik, Dresden.
Pavlova A, Zink RM, Drovetski SV, Rohwer S. 2008. Pleistocene evolution of closely related sand martins Riparia riparia and R. diluta. Mol Phylogenet Evol, 48: 61–73.
Pearson DJ, Backhurst GC. 1988. Characters and taxonomic position of Basra Reed Warbler. Brit Birds, 81: 171–178.
Pearson DJ, Kennerley PR, Bensch S. 2008. A second museum specimen of large-billed reed warbler Acrocephalus orinus. Bull Br Ornithol Club, 128: 136–137.
Penhallurick J. 2010. A correction to Penhallurick & Robson (2009). Forktail, 26: 147–148.
Penhallurick J, Robson C. 2009. The generic taxonomy of parrotbills (Aves: Timaliidae). Forktail, 25: 137–141.
Peters JL. 1960a. Family Alaudidae. In: Mayr E, Greenway JC (eds) Check-list of Birds of the World, Vol. IX. Museum of Comparative Zoology, Cambridge, Mass, pp 3–80.
Peters JL. 1960b. Family Hirundinidae. In: Mayr E, Greenway JC (eds) Check-list of Birds of the World, Vol. IX. Museum of Comparative Zoology, Cambridge, Mass, pp 80–129.
Pratt HD, Bruner PL, Berrett DG. 1987. The Birds of Hawaii and the Tropical Pacific. Princeton University Press, Princeton, NJ.
Rand A L, Deignan HG. 1960. Family Pycnonotidae. In: Mayr E, Greenway JC (eds) Check-List of Birds of the World, Vol. IX. Museum of Comparative Zoology, Cambridge, pp 221–300.
Rappole JH, Renner SC, Nay Myo Shwe, Sweet PR. 2005. A new species of scimitar-babbler (Timaliidae: Jabouilleia) from the sub-Himalayan region of Myanmar. Auk, 122: 1064–1069.
Rasmussen PC, Anderton JC. 2005. Birds of South Asia: the Ripley Guide, First Ed. Lynx Edicions, Barcelona.
Rasmussen PC, Anderton JC. 2012. Birds of South Asia: the Ripley Guide, Second Ed. Lynx Edicions, Barcelona.
Rasmussen PC, Round PD, Dickinson EC, Lambert FG. 2000. A new bush-warbler (Sylviidae, Bradypterus) from Taiwan. Auk, 117: 279–289.
Rasmussen PC. 2012. Then and now: new developments in Indian systematic ornithology. J Bombay Nat Hist Soc, 109: 3–16.
Reddy S, Cracraft J. 2007. Old World Shrike-babblers (Pteruthius) belong with the New World Vireos. Vireonidae. Mol Phylogenet Evol, 44: 1352–1357.
Reddy S, Moyle RG. 2011. Systematics of the scimitar babblers (Pomatorhinus: Timaliidae): phylogeny, biogeography, and species-limits of four species complexes. Biol J Linn Soc, 102: 846–869.
Reeves AB, Drovetski SV, Fadeev V. 2008. Mitochondrial DNA data imply a stepping-stone colonizaiton of Beringia by arctic warbler Phylloscopus borealis. J Avian Biol, 39: 567–575.
Reichenow A. 1902–1903. Die Vögel Afrikas. Neudamm (J. Neumann).
Rheindt FE. 2006. Splits galore: the revolution in Asian leaf warbler systematics. Bird Asia, 5: 25–39.
Rheindt FE. 2010. New biogeographic records for the avifauna of Taliabu (Sula Islands, Indonesia), with preliminary documentation of two previously undiscovered taxa. Bull Br Ornithol Club, 130: 33–51.
Richman AD, Price T. 1992. Evolution of ecological differences in the Old World Leaf Warblers. Nature, 355: 817–821.
Roberts A. 1940. The Birds of South Africa. HF & G. Witherby, London.
Rokas A, Carroll SB. 2006. Bushes in the tree of life. PLoS Biol, 4: e352.
Round PD, Hansson B, Pearson DJ, Kennerley PR, Bensch S. 2007. Lost and found: the enigmatic large-billed reed warbler Acrocephalus orinus rediscovered after 139 years. J Avian Biol, 38: 133–138.
Rozendaal F. 1987. Description of a new species of bush warbler of the genus Cettia Bonaparte, 1834 (Aves: Sylviidae) from Yamdena, Tanimbar Islands, Indonesia. Zool Mededel, 61: 177–202.
Ryan PG, Bloomer P. 1997. Geographic variation in Red Lark Certhilauda burra plumage, morphology, song and mitochondrial DNA haplotypes. Ostrich, 68: 31–36.
Ryan PG, Bloomer P. 1999. The Long-billed Lark complex: a species mosaic in south-western Africa. Auk, 116: 194–208.
Ryan PG, Dean WRJ, Madge SC, Pearson DJ. 2006. Family Cisticolidae. In: del Hoyo J, Elliott A, Christie DA (eds). Handbook of the Birds of the World. Vol 11. Old World Flycatchers to Old World Warblers. Lynx Edicions, Barcelona, pp 378–491
Ryan PG, Hood I, Bloomer P, Komen J, Crowe TM. 1998. Barlow's Lark: a new species in the Karoo Lark Certhilauda albescens complex of southwest Africa. Ibis, 140: 605–619.
Saitoh T, Alström P, Nishiumi I, Shigeta Y, Williams D, Olsson U, Ueda K. 2010. Old divergences in a boreal bird supports longterm survival through the Ice Ages. BMC Evol Biol, 10: 35.
Saitoh T, Nishiumi I, Alström P, Olsson U, Ueda K. 2006. Deep phylogeographical divergences among far eastern populations of the widespread Arctic Warbler. J Ornithol, 147: 242.
Saitoh T, Shigeta Y, Ueda K. 2008. Morphological differences among populations of the Arctic Warbler with some intraspecific taxonomic notes. Ornithol Sci, 7: 135–142.
Salomon M, Bried J, Helbig AJ, Riofrio J. 1997. Morphometric differentiation between male Common Chiffchaffs, Phylloscopus (c. ) collybita Vieillot, 1817, and Iberian Chiffchaffs, P. (c. ) brehmii Homeyer, 1871, in a secondary contact zone (Aves). Zoologischer Anzeiger, 236: 25–36.
Salomon M, Voisin JF, Bried J. 2003. On the taxonomic status and denomination of the Iberian Chiffchaffs. Ibis, 145: 87–97.
Salomonsen F. 1929. Bemerkungen über die Gruppe Acrocephalus arundinaceus L. J Ornithol, 77: 267–281.
Schönwetter M. 1979. Handbuch der Oologie. Band Ⅱ. Akademie-Verlag, Berlin.
Sclater WL. 1930. Systema Avium Aethiopicarum. British Ornithologists' Union, London
Sefc KM, Payne RB, Sorenson MD. 2003. Phylogenetic relationships of African sunbird-like warblers: Moho (Hypergerus atriceps), Green Hylia (Hylia prasina) and Tit-hylia (Pholidornis rushiae). Ostrich, 74: 8–17.
Sheldon FH, Gill FB. 1996. A reconsideration of songbird phylogeny, with emphasis on the evolution of titmice and their sylvioid relatives. Syst Biol, 45: 473–495.
Sheldon FH, Oliveros CH, Taylor SS, McKay B, Lim HC, Rahman MA, Mays H, Moyle RG. 2012. Molecular phylogeny and insular biogeography of the lowland tailorbirds of Southeast Asia (Cisticolidae: Orthotomus). Mol Phylogenet Evol, 65: 54–63.
Sheldon FH, Whittingham LA, Moyle RG, Slikas B, Winkler DW. 2005. Phylogeny of swallows (Aves: Hirundinidae) estimated from nuclear and mitochondrial DNA sequences. Mol Phylogenet Evol, 35: 254–270.
Shirihai H, Gargallo G, Helbig AJ. 2001. Sylvia warblers. Identification, taxonomy and phylogeny of the genus Sylvia. Christopher Helm, London.
Sibley CG, Ahlquist JE. 1990. Phylogeny and ClassiWcation of Birds. A Study in Molecular Evolution. Yale University Press, New Haven and London.
Sibley CG, Monroe Jr BL. 1990. Distribution and Taxonomy of Birds of the World. Yale University Press, New Haven.
Sibley CG, Monroe Jr BL. 1993. A Supplement to Distribution and Taxonomy of Birds of the World. Yale University Press, New Haven.
Silva-Iturriza A, Ketmaier V, Tiedemann R. 2010. Mitochondrial DNA suggests multiple colonizations of central Philippine islands (Boracay, Negros) by the sedentary Philippine bulbul Hypsipetes philippinus guimarasensis (Aves). J Zool Syst Evol Res, 48: 269–278.
Song G, Qu Y, Yin Z, Li S, Liu N, Lei F. 2009. Phylogeography of the Alcippe morrisonia (Aves: Timaliidae): long population history beyond late Pleistocene glaciations. BMC Evol Biol, 9: 143.
Spellman GM, Cibois A, Moyle RG, Winker K, Barker FK. 2008. Clarifying the systematics of an enigmatic avian lineage: What is a Bombycillid? Mol Phylogenet Evol, 49: 1036–1040.
Spicer GS, Dunipace L. 2004. Molecular phylogeny of songbirds (Passeriformes) inferred from mitochondrial 16S ribosomal RNA gene sequences. Mol Phylogenet Evol, 30: 325–335.
Stresemann E, Arnold J. 1949. Speciation in the group of Great Reed-Warblers. J Bombay Nat Hist Soc, 48: 428–443.
Svensson L, Prys-Jones R, Rasmussen PC, Olsson U. 2008. Discovery of ten new specimens of large billed reed warbler Acrocephalus orinus, and new insights into its distributional range. J Avian Biol, 39: 605–610.
Svensson L, Prys-Jones R, Rasmussen PC, Olsson U. 2010. The identification and distribution of the enigmatic Large-billed Reed Warbler Acrocephalus orinus. Ibis, 152: 323–334.
Svensson L. 2001. Identification of western and eastern olivaceous, booted and Sykes's warblers. Bird World, 14: 192–219.
Tieleman BI, Williams JB, Bloomer P. 2003. Adaptation of metabolism and evaporative water loss along an aridity gradient. Proc Roy Soc Lond B, 270: 207–214.
Timmins RJ, Ostrowski S, Mostafawi N, Noori H, Rajabi AM, Svensson L, Olsson U, Poole CM. 2010. New information on the Large-billed Reed Warbler Acrocephalus orinus, including its song and breeding habitat in north-eastern Afghanistan. Forktail, 26: 9–23.
Trainor CR, Verbelen F, Hoste S. 2012. Rediscovery of the Timor Bush-warbler Locustella (Bradypterus) timorensis on Alor and Timor, Wallacea: clarifying taxonomic affinities, defining habitat and survey recommendations. Bird Conserv Int, 22: 354–369.
Treplin S, Siegert R, Bleidorn C, Shokellu Thompson H, Fotso R, Tiedemann R. 2008. Molecular phylogeny of songbirds (Aves: Passeriformes) and the relative utility of common nuclear marker loci. Cladistics, 24: 328–349.
Turner A. 2004. Family Hirundinidae (swallows and martins). In: Del Hoyo J, Elliott A, Christie D (eds) The Birds of the World, Vol. 9, Lynx Edicions, Barcelona, Spain, pp 602–685
Vaurie C. 1951. A study of Asiatic larks. Notes from the Walter Koelz collections number 11. Bull AMNH, 97: 435–526.
Vaurie C. 1954. Systematic Notes on Palearctic birds. No. 7. Alaudidae and Motacillidae (genus Anthus). Amer Mus Novit, 1672: 1–13.
Vaurie C. 1959. The Birds of the Palearctic Fauna. Passeriformes. Witherby, London.
Verbelen F, Trainor CR. 2012. Rediscovery of the Timor Bush Warbler Locustella (Bradypterus) timorensis on Alor and Timor, Wallacea, Indonesia. Bird Asia, 17: 47–48.
Verheyen R. 1958. Contribution a l'anatomie de base et la systematique des Alaudidae (Passeriformes). Alauda, 26: 1–25.
Voelker G, Light JE. 2011. Palaeoclimatic events, dispersal and migratory losses along the Afro-European axis as drivers of biogeographic distribution in Sylvia warblers. BMC Evol Biol, 11: 163.
Voelker G, Melo M, Bowie RCK. 2009. A Gulf of Guinea island endemic is a member of a Mediterranean-centered bird genus. Ibis, 151: 580–583.
Voelker G, Spellman GM. 2003. Nuclear and mitochondrial DNA evidence of polyphyly in the avian superfamily Muscicapoidea. Mol Phylogenet Evol, 30: 386–394.
Voous KH. 1977. List of Recent Holarctic Bird Species, Second Ed. British Ornithologists' Union, London.
Warren BH, Bermingham E, Prys-Jones RP, Thebaud C. 2005. Tracking island colonization history and phenotypic shifts in Indian Ocean bulbuls (Hypsipetes: Pycnonotidae). Biol J Linn Soc, 85: 271–287.
Warren BH, Bermingham E, Prys-Jones RP, Thébaud C. 2006. Immigration, species radiation and extinction in a highly diverse songbird lineage: white-eyes on Indian Ocean islands. Mol Ecol, 15: 3769–3786.
Watson GE, Traylor MA, Jr Mayr E. 1986. In: Mayr E, Cottrell GW (eds) Check-list of Birds of the World. Vol. 11. Museum of Comparative Zoology, Cambridge, Massachusetts.
Whittingham LA, Slikas B, Winkler DW, Sheldon FH. 2002. Phylogeny of the tree swallow genus, Tachycineta (Aves: Hirundinidae), by Bayesian analysis of mitochondrial DNA sequences. Mol Phylogenet Evol, 22: 430–441.
Williamson K. 1968. Identification for Ringers. The genera Cettia, Locustella, Acrocephalus and Hippolais, Third Ed. British Trust for Ornithology, Tring.
Winkler DW, Sheldon FH. 1993. The evolution of nest construction in swallows (Hirundinidae): a molecular phylogenetic perspective. Proc Natl Acad Sci USA, 90: 5705–5707.
Winterbottom JM. 1957. The problem of status of the Dusky Bush Lark Mirafra nigricans. Ostrich, 28: 240–241.
Wolters HE. 1979. Die Vogelarten der Erde. Paul Parey, Hamburg and Berlin.
Wolters HE. 1982. Die Vogelarten der Erde. 7. Lieferung Paul Parey, Hamburg and Berlin.
Yeung CKL, Lei F, Yang X, Han L, Lin M, Li SH. 2006. Molecular phylogeny of parrotbills. Paper presented at the 24th International Ornithological Congress of 2006, Hamburg, Germany. Abstract J Orn, 147(Suppl. 1): 87–88.
Yeung CKL, Lin R-C, Lei F, Robson C, Hung LM, Liang W, Zhou F, Han L, Li S-H, Yang X. 2011. Beyond a morphological paradox: complicated phylogenetic relationships of the parrotbills (Paradoxornithidae, Aves). Mol Phylogenet Evol, 61: 192–202.
Zahavi A. 1957. The breeding birds of the Huleh swamp and lake (northern Israel). Ibis, 99: 606.
Zhang S, Yang L, Yang X, Yang J. 2007. Molecular phylogeny of the yuhinas (Sylviidae: Yuhina): a paraphyletic group of babblers including Zosterops and Philippine Stachyris. J Ornithol, 148: 417–426
Zhou F, Jiang AW. 2008. A new species of babbler (Timaliidae: Stachyris) from the Sino-Vietnamese border region of China. Auk, 125: 420–424.
Zink RM, Pavlova A, Drovetski S, Rohwer S. 2008. Mitochondrial phylogeographies of five widespread Eurasian bird species. J Ornithol, 149: 399–413.
Zink RM, Pavlova A, Rohwer S, Drovetski SV. 2006. Barn swallows before barns: population histories and intercontinental colonization. Proc R Soc London B, 273: 1245–1252.
Zou F, Lim HC, Marks BD, Moyle RG, Sheldon FH. 2007. Molecular phylogenetic analysis of the Grey-cheeked Fulvetta (Alcippe morrisonia) of China and Indochina: A case of remarkable genetic divergence in a "species". Mol Phylogenet Evol, 44: 165–174.
Zuccon D, Ericson PGP. 2010. The phylogenetic position of the Black-collared Bulbul Neolestes torquatus. Ibis, 152(2): 386–392.

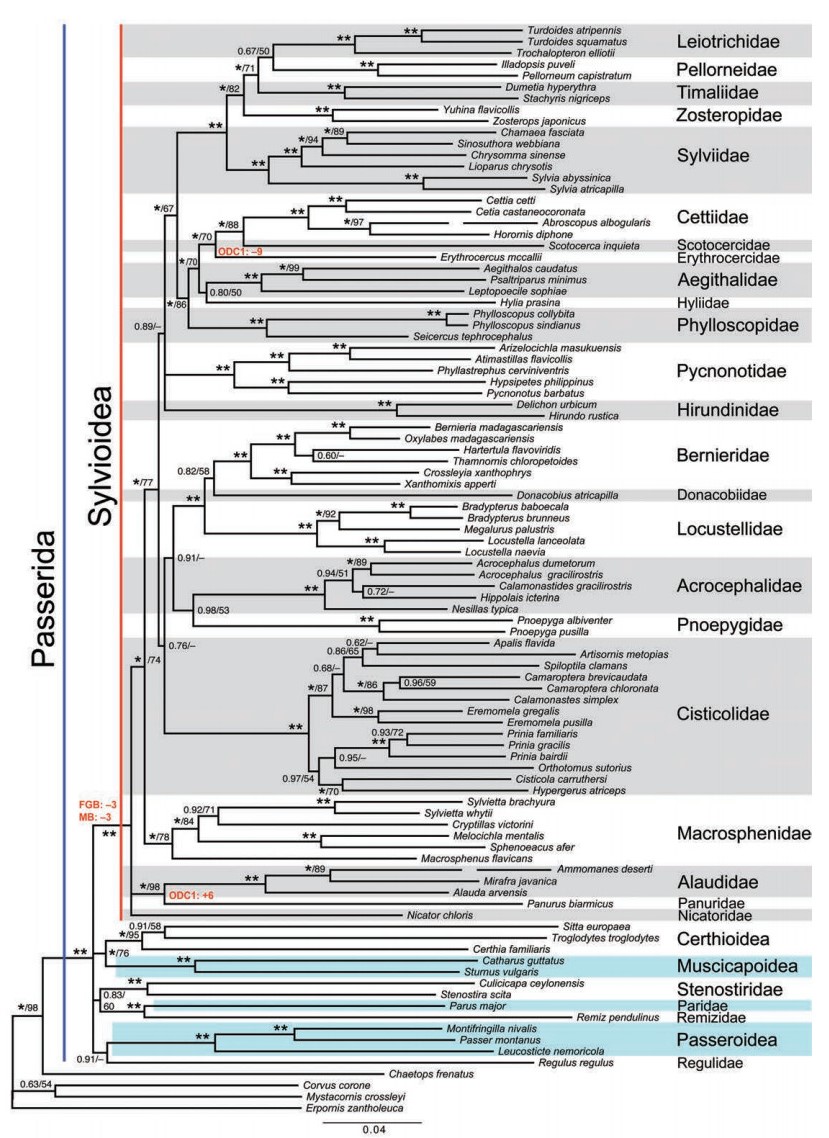
 DownLoad:
DownLoad:
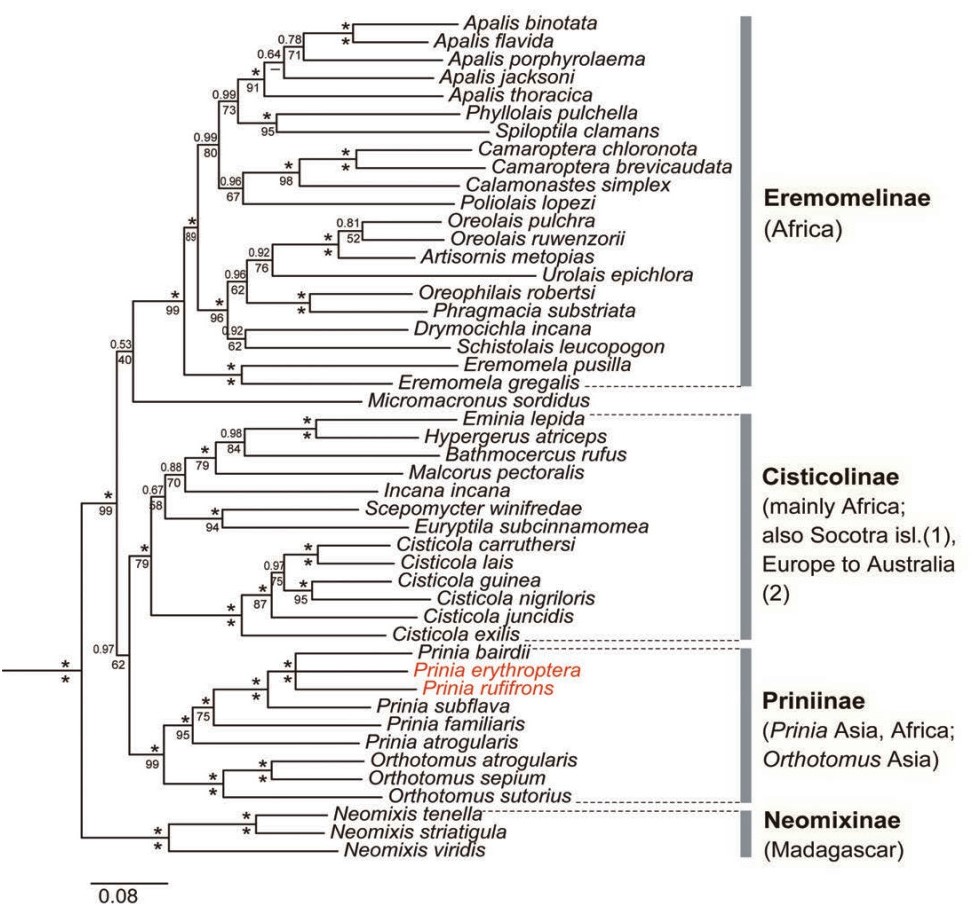
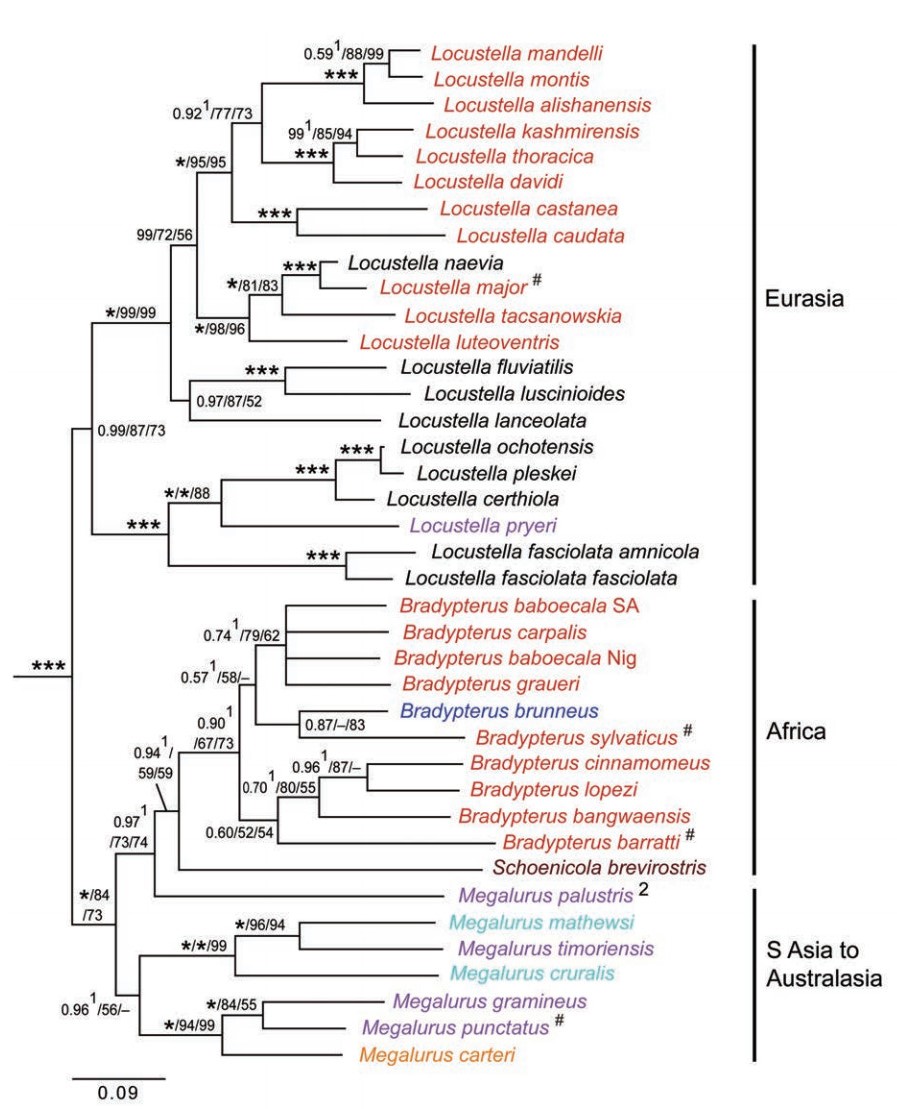
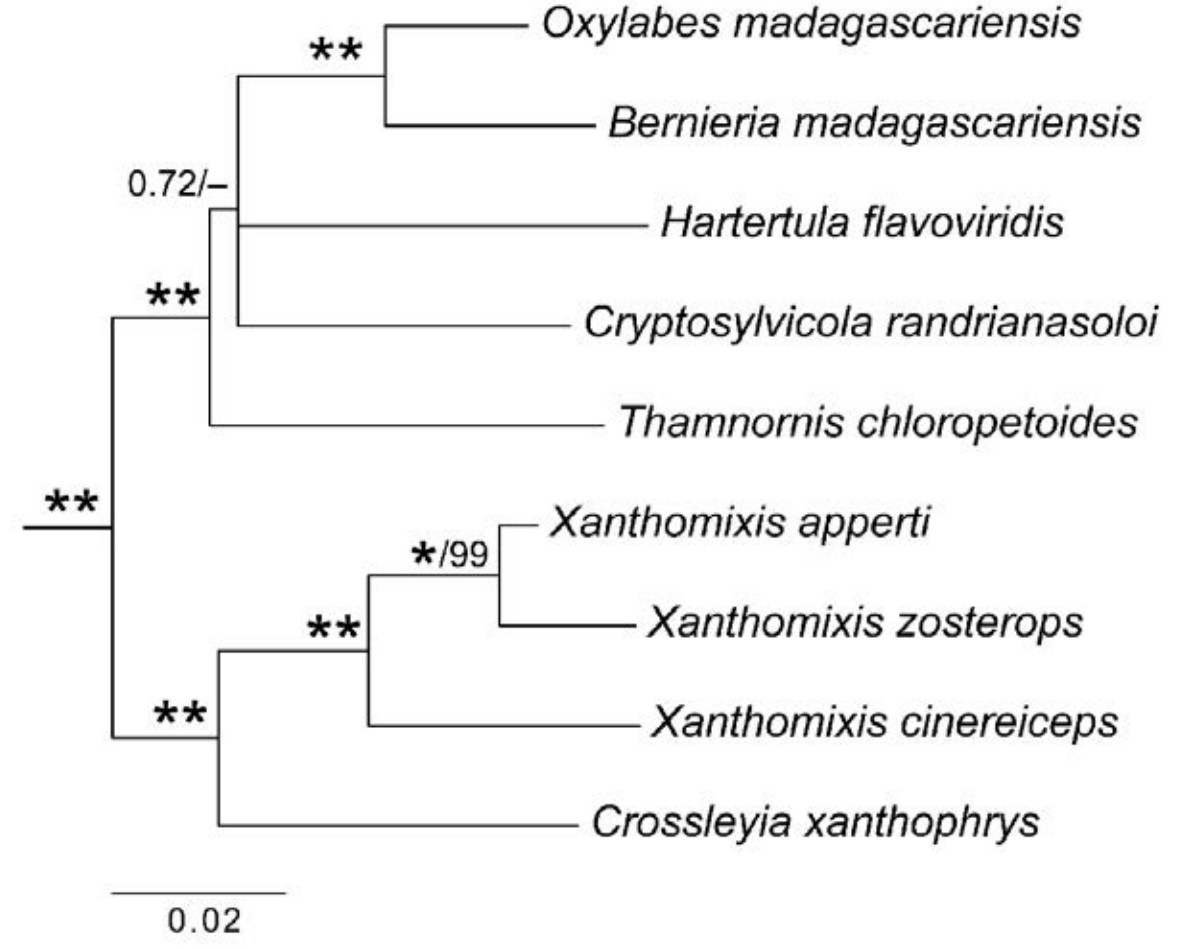
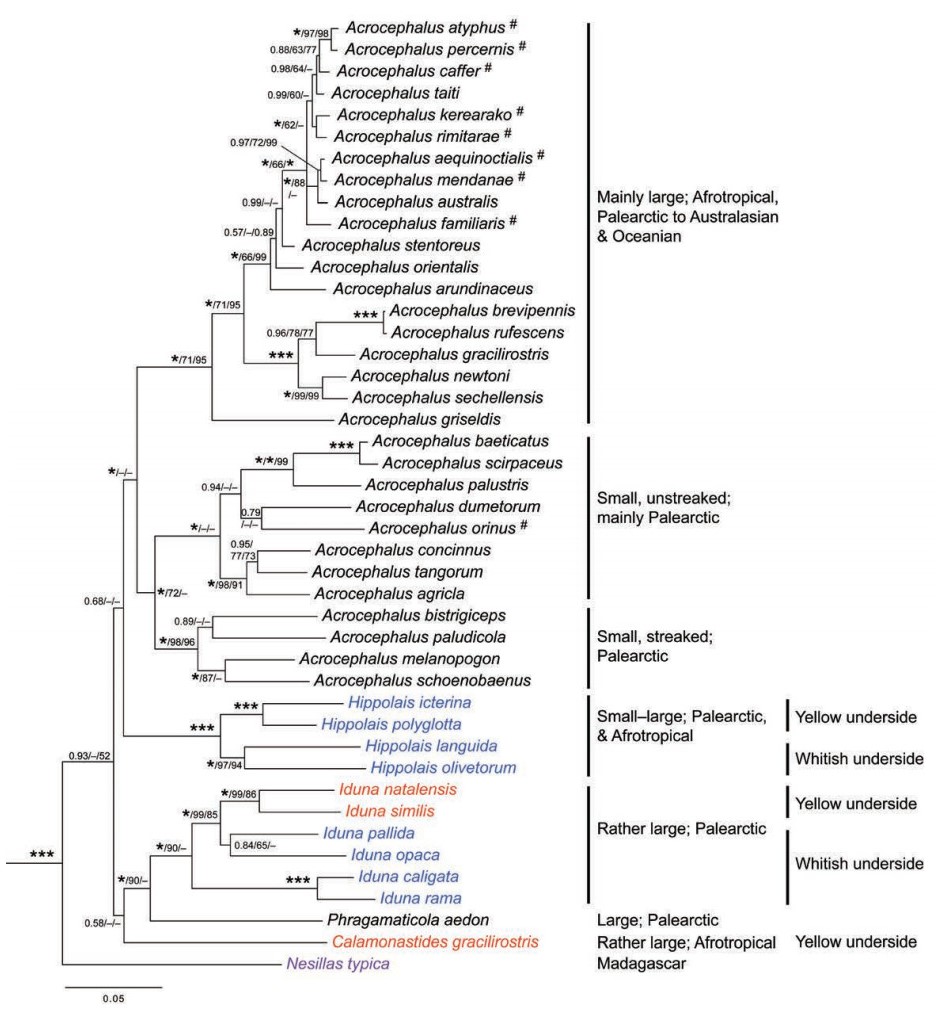
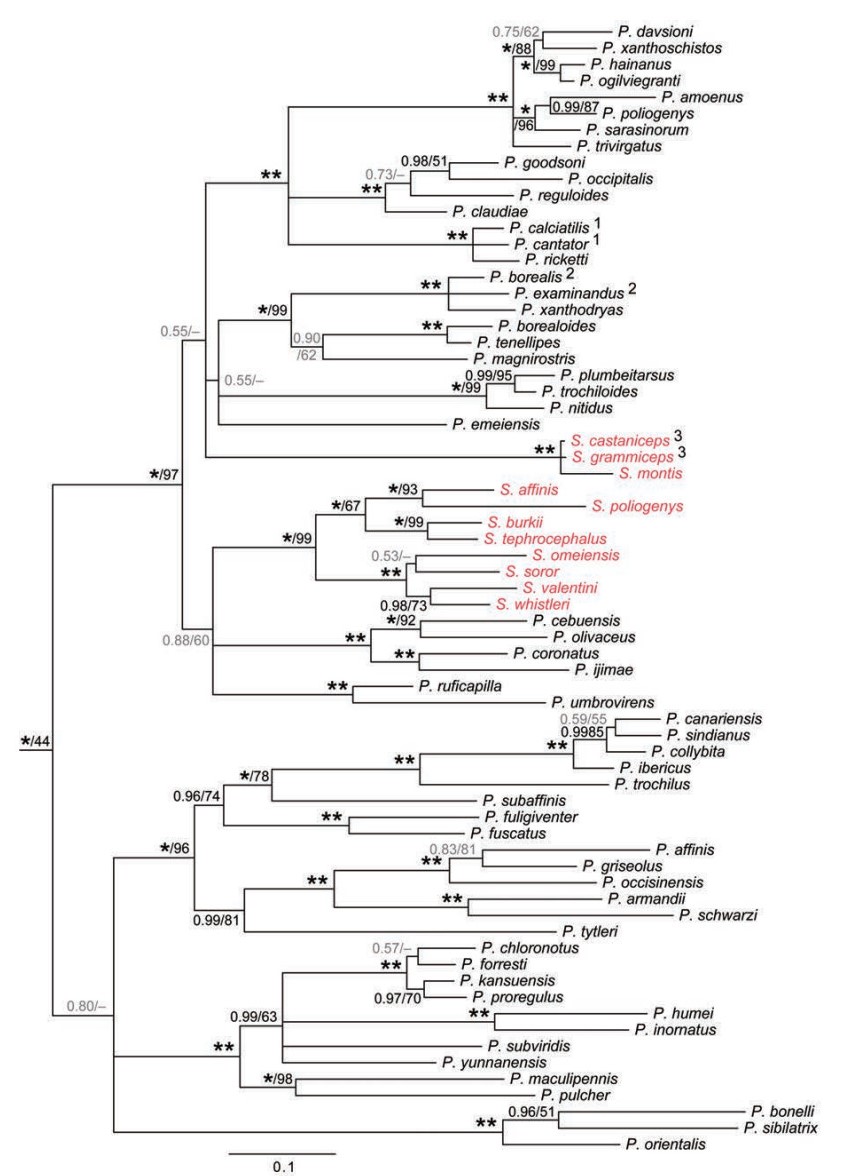
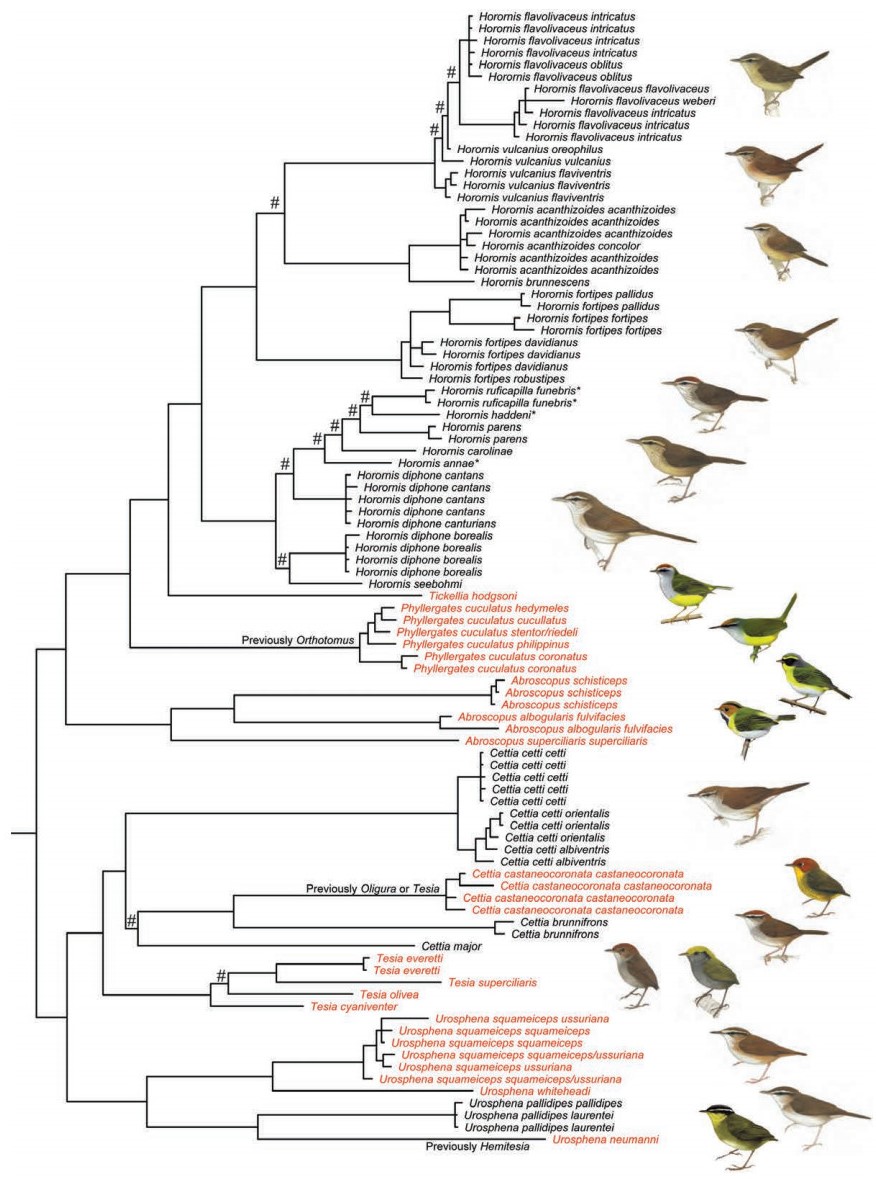





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds