Mariya V. SIVAY, Nikita Y. SILKO, Kirill A. SHARSHOV, Aleksander V. PROKUDIN, Laixing LI, Min YANG, Sheng CAO, Aleksander M. SHESTOPALOV. 2011: The role of wild goose (Anser) populations of Russia and the Tibet Plateau in the spread of the avian influenza virus. Avian Research, 2(3): 140-146. DOI: 10.5122/cbirds.2011.0022
Citation:
Mariya V. SIVAY, Nikita Y. SILKO, Kirill A. SHARSHOV, Aleksander V. PROKUDIN, Laixing LI, Min YANG, Sheng CAO, Aleksander M. SHESTOPALOV. 2011: The role of wild goose (Anser) populations of Russia and the Tibet Plateau in the spread of the avian influenza virus. Avian Research, 2(3): 140-146. DOI: 10.5122/cbirds.2011.0022
Mariya V. SIVAY, Nikita Y. SILKO, Kirill A. SHARSHOV, Aleksander V. PROKUDIN, Laixing LI, Min YANG, Sheng CAO, Aleksander M. SHESTOPALOV. 2011: The role of wild goose (Anser) populations of Russia and the Tibet Plateau in the spread of the avian influenza virus. Avian Research, 2(3): 140-146. DOI: 10.5122/cbirds.2011.0022
Citation:
Mariya V. SIVAY, Nikita Y. SILKO, Kirill A. SHARSHOV, Aleksander V. PROKUDIN, Laixing LI, Min YANG, Sheng CAO, Aleksander M. SHESTOPALOV. 2011: The role of wild goose (Anser) populations of Russia and the Tibet Plateau in the spread of the avian influenza virus. Avian Research, 2(3): 140-146. DOI: 10.5122/cbirds.2011.0022
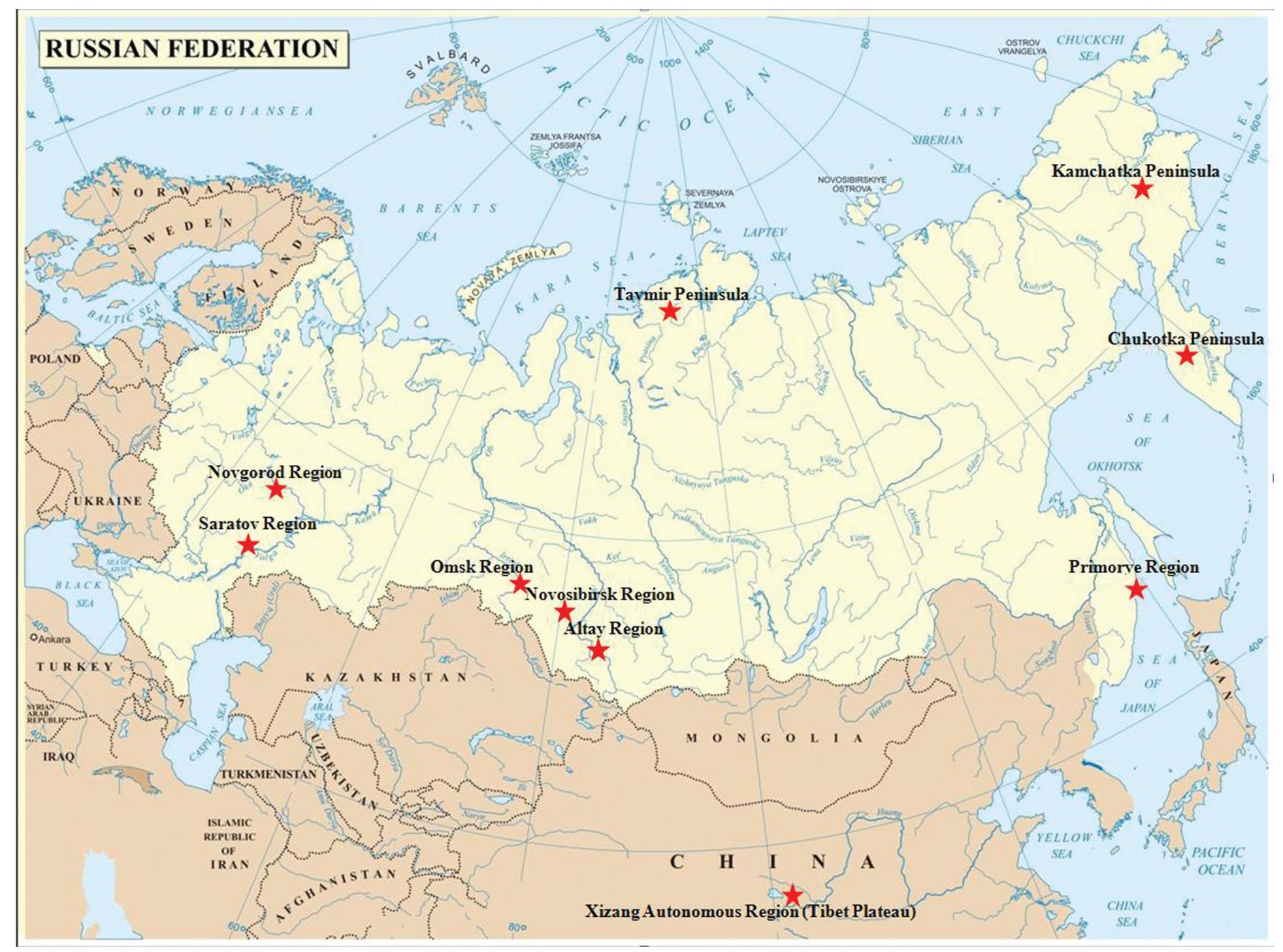
Wild birds of the orders Anseriformes and Charadriiformes represent a natural reservoir of low pathogenic avian influenza (LPAI) viruses (family Orthomyxoviridae). Wild geese (order Anseriformes) relating to waterfowls undertake extensive migration flights reaching thousands of kilometers. Isolation of the avian influenza virus (AIV) from wild geese is quite low or absent. The aims of this study are to monitor the AIV in different wild goose species, nesting on Russian territory and the Tibet Plateau and to analyze the derived data for the purpose of determining the role of these wild bird species in spreading pathogens. In our study 3245 samples from nine wild goose species in nine regions of Russia and on the territory of the Tibet Plateau (the Xizang Autonomous Region) were tested and no AIV were detected. Our study shows the non-essential role of wild geese in the spread of the AIV over long distances and reaches the conclusion that geese are probably not natural reservoirs for the primary viruses. However, further inquiry of AIV in wild goose populations is required. Studies of wild geese and AIV ecology will allow us to obtain more information about pathogen-host relationships and to make arrangements for the maintenance of wild goose populations.
As a species of the genus Lophophorus, the Himalayan Monal (Lophophorus impejanus), also known as the Impeyan Monal, is a national first grade protected species in China. Despite considerable number of surveys conducted for years over its ranges, current knowledge of the Himalayan Monal in China is still poor. Thus, an estimate of its distribution, numbers and habitat was conducted during a two-year investigation from 2008 to 2009 in southern Tibet, especially in Lhozhag, Cona and Yadong counties. In total, 12 sightings of the Himalayan Monal were recorded during this study period. We summarize the domestic results achieved so far about this species.
Appearance and international status
The Himalayan Monal is a relatively large-sized bird within the family Phasianidae. With about 70 cm in length, the weight of males ranges from 1980 to 2380 g and that of females from and 1800 to 2150 g. Adult males possess a long crest, feathered with multi-colored plumage throughout its body, while the females, like other pheasants, are dull in color with their upper parts covered with dark brownish black feathers. Notable features in males are their long crest which is metallic green, changeable reddish copper in the back and sides of the neck and a prominent white back while in flight. Tail feathers of the male are uniformly rufous, darker towards the tips, while the lower tail coverts of the female are white, barred with black and rufous. The female has a prominent white patch in the fore neck and a white strip on the tail. First year males are immature and resemble female chicks, but are larger in the first year and their immaturity is less pronounced (Delacour, 1977).
It is also one of the best recognized pheasant species of the western Himalayas because of the metallic, rainbow-colored plumage, iridescent blue head crest and highly exhibited sexual dimorphism of the male. These characteristics have qualified this mountainous forest bird to the status as national bird of Nepal and the state bird of Uttaranchal and Himalchal Pradesh in India (Ramesh et al., 1999; Ramesh, 2003).
Distribution
The natural range of the the Himalayan Monal spreads from eastern Afghanistan through the Himalayas, including Pakistan, India (the state of Jammu & Kashmir, Himachal Pradesh, Uttaranchal, Sikkim and Arunachal Pradesh), China (the Tibet region), Nepal and Bhutan. There is also a report of its occurrence in Myanmar, based on a specimen taken in 1969 from the Hpungan pass (27°30′N, 96°48′E), along the Indo-Myanmar border (Yin, 1970). Domestically there is evidence showing that this montane bird is resident on the southern slopes of the Himalaya mountains, specifically in the counties Dingjie, Tingri, Nyalam, Kuala, Yadong, Nyingchi, Medog and Zayu (Zheng and Wang, 1998; Li, 2004) and Gongshan County in the northwest of Yunnan Province (He, 2001). Our data suggest that the Himalayan Monal is mainly found in Medog, Zayu, Cona, Lhunze, Lhozhag, Nyalam, Dingjie, Gamba and Yadong counties, on the south slopes of the Himalayas. Its western-most location was confirmed to be in Nyalam County. The bird ranges in elevation from 3800 to 4300 m in the summer and from 3200 to 3500 m in the winter.
Habitat
The Himalayan Monal occupies the upper temperate oak-conifer forests and subalpine oak forests interspersed with open grassy slopes, cliffs and alpine meadows between 2400 and 4500 m elevation, mostly concentrating in a narrow belt between 2700 and 3700 m (Grimmett et al., 1998). They exhibit clear migration in elevation, reaching as low as 2000 m in winter (Ramesh, 2003). Our observations showed that the birds mainly occur in rocky forests where the dominant tree species include Quercus semecarpifolia, Picea spinulosa and Abies spectabilis, characterized by DBH > 20 cm and the oldest tree of the stands was estimated to be more than 500 years old. The bush layer is primarily composed of Salix oritrepha and Rosa tibetica interspersed with steep slopes (45°–50°), cliffs and alpine meadows at an elevation of 3800–4000 m. However, they show tolerance to snow and have been observed to dig through snow for roots, tubers, other plant parts and invertebrates (McGowan, 1994; Kumar, 1997).
Daily behavior around the Kajiu Monastery in Tibet
The Kajiu Monastery, a small Buddhist temple (28°05′38″N, 91°07′34″E) at an elevation of 4000 m is situated on top of a hill in Lakang town, located on the Chinese-Indian border area of Lhozhag County. Our observations suggest that there were 36–37 individual birds, consisting of 8–10 males, 16–20 females and 7–8 sub-adults, inhabiting an area with a radius of of 5 km around the monastery. The birds move up to the monastery along a tortuous path by walking or jumping with slight flapping of the wings when they climb over terraces 1.0–2.0 m high every morning. They spent about two hours to feed on the barrier wall of the temple where the monks provide food to them until they glide swiftly to their natural habitat at the edge of the cliffs. We observed the birds soaring at a distance of more than 500 m, accompanied by hurried and sharp voices when they were interrupted by some disturbances. From 5:00 to 8:00 in the afternoon, they hunted some plants for foods on the meadow nearby cliffs and then hid on the branches of large oak trees at night. The birds are gregarious during the non-breeding season and live in small flocks of the same gender.
Simple summary of breeding ecology
We found a ground nest without any content at an elevation of 3940 m (28°05′35″N, 91°07′21″E), located under an oak tree 8 m tall and 30 cm in DBH on a steep slope near a cliff. The measurement of the nest was 3 cm × 35 cm × 39 cm in depth, width and length. Some feathers and excrements were observed inside the nest.
Other sightings of this bird have been made and some references about nesting of the Himalayan Monal exist. For example, Johnsgard (1986) reported that the clutch size of this species is most commonly four or five eggs in the wild, with three-egg clutches also fairly common and sometime only two eggs are present. The eggs described by Ali and Ripley (1983) were pale yellowish or reddish buff, freckled and spotted with reddish brown. The incubation period, as Howman (1993) reported, is 28 days. Whether the males played an active role in chick rearing could not be established, as reported by Johnsgard (1986), although there are a few suggestions in the literature that males help caring for the young. Still, this is certainly not a typical situation and most authorities contend that he takes no part in the rearing phase. Gaston (1981) described that the Himalayan Monal exhibits a special display flight during the breeding season, consisting of extended gliding, with the tail fanned and the wings held well above the level of the body and with a repeated piping call that varies from fairly soft to strident.
Figure
1.
Two males foraging on the barrier wall of the Kajiu Monastery (photo by Xiaoping YU)
We need to refer to the Buddhist temples and their impact on the conservation of wildlife in the Tibet areas, where a large number of monasteries exist. The size of the Chinese population of the Himalayan Monal was estimated in 2004 to be no more than 1000 individual birds (Li, 2004). The population density of this bird species and the forest cover at the temples are significantly larger than that for other areas (Yu et al., unpublished). The reason for this is that the Buddhist monks and their adherents treat wildlife according their faith and traditional culture. As in most Tibetan temples, the monks of the Kajiu Monastery often provide a variety of foods such as highland barley, tsamb and ghee to wild birds. The monasteries are usually the distribution centers for wild birds with the densest populations. As a result, the population density of the Himalayan Monal near the Kajiu Monastery is 2.03 individual birds per km2, which is much greater than that of Yadong and Cona counties (0.052 birds per km2).
Elements of depletion and conservation
Similar to many other spectacular species, the Himalayan Monal is usually captured by local people for their own purposes. The bird has been decisively incorporated into the traditional folklore of many Himalayan cultures (Delacour, 1977). The use of feathers is deemed significant in rituals and local ceremonies among the people across Asia. For example, men in the Indian state of Himachal Pradesh hunted males for their crests, which adorned their ceremonial hats as a sign of high social status and the defining of property qualification, although this pressure subsided after hunting was banned in Himachal Pradesh in 1982 (Kumar et al., 1997; Ramesh, 2003). In addition, pheasants are aesthetically pleasing to humans and irrefutably a source of food. For instance, chickens provide a stable diet for millions of people in the world and sport hunting is very popular entertainment in Europe and America. Thus, several of these factors have collectively contributed to the removal of wild populations in their native regions. As well, pheasant populations have experienced strong depletion, attributed to loss of their habitats by human encroachment and development in most part of their ranges (McGowan and Gillman, 1997).
Another example is Yadong County in Tibet where the Himalayan Monal and Satyr Tragopan (Tragopan satyra) were illegally hunted by local people using traps set on the paths which the birds must pass on snowy days in the winter. These birds are captured to stuff specimens for commercial purposes.
The present state of knowledge of the Himalayan Monal is sketchy and it is necessary to obtain scientific data to understand their behavior and ecological requirements better. This dearth of information may be partly due to its remoteness and lack of accessibility, dense forests and steep terrain of their habitats making them difficult to observe. Furthermore, poaching and habitat degradation from human activities contribute to the situation. We hope that our survey, designed to gather new information on the distribution and ecology of the Himalayan Monal, will encourage further study and conservation of this magnificent species in Tibet and elsewhere. After more investigations, conservation measures for the Himalayan Monal should be implemented for the protection of this species.
Acknowledgements
We are grateful to the State Forestry Administration to provide financial support to this study. We thank Mr Rong DONG and Mr. Xia LI most sincerely for participating in the fieldwork in a harsh and arduous environment for so long. As well, there are several local people who are not mentioned, but helped in some way or another, which enabled the successful completion of this study. In addition, we are grateful to J.R.B. Miller and G.S. Rawat for their help in providing related literature.
Boyce WM, Sandrock C, Kreuder-Johnson C, Kelly T, Cardona C. 2009. Avian influenza viruses in wild birds: a moving target. Comp Immunol Microbiol Infect Dis, 32: 275–286.
Brown C. 2006. Avian influenza: Virchow's reminder. Am J Pathol, 168: 6–8.
Brown JD, Stallknecht DE. 2008. Wild bird surveillance for the avian influenza virus. Method Mol Biol, 436: 85–97.
Chen H, Smith GJ, Li KS, Wang J, Fan XH, Rayner JM, Vijaykrishna D, Zhang JX, Zhang LJ, Guo CT, Cheung CL, Xu KM, Duan L, Huang K, Qin K, Leung YH, Wu WL, Lu HR, Chen Y, Xia NS, Naipospos TS, Yuen KY, Hassan SS, Bahri S, Nguyen TD, Webster RG, Peiris JS, Guan Y. 2006. Establishment of multiple sublineages of H5N1 influenza virus in Asia: implications for pandemic control. Proc Natl Acad Sci U S A, 103: 2845–2850.
Chen H, Smith GJ, Zhang SY, Qin K, Wang J, Li KS, Webster RG, Peiris JS, Guan Y. 2005. Avian flu: H5N1 virus outbreak in migratory waterfowl. Nature, 436: 191–192.
De Marco MA, Foni E, Campitelli L, Raffini E, Delogu M, Donatelli I. 2003. Long-term monitoring for avian influenza viruses in wild bird species in Italy. Vet Res Commun, 1: 107–114.
Fereidouni SR, Werner O, Starick E, Beer M, Harder TC, Aghakhan M, Modirrousta H, Amini H, Moghaddam MK, Bozorghmehrifard MH, Akhavizadegan MA, Gaidet N, Newman SH, Hammoumi S, Cattoli G, Globig A, Hoffmann B, Sehati ME, Masoodi S, Dodman T, Hagemeijer W, Mousakhani S, Mettenleiter TC. 2010. Avian influenza virus monitoring in wintering waterbirds in Iran, 2003–2007. Virol J, 7: 43.
Germundsson A, Madslien KI, Hjortaas MJ, Handeland K, Jonassen CM. 2010. Prevalence and subtypes of influenza A viruses in wild waterfowl in Norway 2006–2007. Acta Vet Scand, 52: 28.
Gladkova NA, Mikheev AV. 1970. Animal life: Birds. V 5. Prosveshenie, Moskow.
Guan Y, Chen H, Li K, Riley S, Leung G, Webster R, Peiris J, Yuen K. 2007. A model to control the epidemic of H5N1 influenza at the source. BMC Infect Dis, 7: 132.
Ip HS, Flint PL, Franson JC, Dusek RJ, Derksen DV, Gill RE Jr, Ely CR, Pearce JM, Lanctot RB, Matsuoka SM, Irons DB, Fischer JB, Oates RM, Petersen MR, Fondell TF, Rocque DA, Pedersen JC, Rothe TC. 2008. Prevalence of Influenza A viruses in wild migratory birds in Alaska: patterns of variation in detection at a crossroads of intercontinental flyways. Virol J, 5: 71.
Keawcharoen J, van Riel D, van Amerongen G, Bestebroer T, Beyer WE, van Lavieren R, Osterhaus AD, Fouchier RA, Kuiken T. 2008. Wild ducks as long-distance vectors of highly pathogenic avian influenza virus (H5N1). Emerg Infect Dis, 14: 600–607.
Komar N, Olsen B. 2008. Avian influenza virus (H5N1) mortality surveillance. Emerg Infect Dis, 14: 1176–1178.
Kou Z, Li Y, Yin Z, Guo S, Wang M, Gao X, Li P, Tang L, Jiang P, Luo Z, Xin Z, Ding C, He Y, Ren Z, Cui P, Zhao H, Zhang Z, Tang S, Yan B, Lei F, Li T. 2009. The survey of H5N1 flu virus in wild birds in 14 Provinces of China from 2004 to 2007. PLoS One, 4: e6926.
Krauss S, Obert CA, Franks J, Walker D, Jones K, Seiler P, Niles L, Pryor SP, Obenauer JC, Naeve CW, Widjaja L, Webby RJ, Webster RG. 2007. Influenza in migratory birds and evidence of limited intercontinental virus exchange. PLoS Pathog, 3: e167.
Lebarbenchon C, Feare CJ, Renaud F, Thomas F, Gauthier-Clerc M. 2010. Persistence of highly pathogenic avian influenza viruses in natural ecosystems. Emerg Infect Dis, 16: 1057–1062.
Lindh E, Huovilainen A, Ratti O, Ek-Kommonen C, Sironen T, Huhtamo E, Poysa H, Vaheri A, Vapalahti O. 2008. Orthomyxo-, paramyxo- and flavivirus infections in wild waterfowl in Finland. Virol J, 5: 35.
Munster VJ, Fouchier RA. 2009. Avian influenza virus: of virus and bird ecology. Vaccine, 27: 6340–6344.
Neumann G, Chen H, Gao GF, Shu Y, Kawaoka Y. 2010. H5N1 influenza viruses: outbreaks and biological properties. Cell Res, 20: 51–61.
Olsen B, Munster VJ, Wallensten A, Waldenstrom J, Osterhaus AD, Fouchier RA. 2006a. Global patterns of influenza a virus in wild birds. Science, 312: 384–388.
Olsen SJ, Ungchusak K, Birmingham M, Bresee J, Dowell SF, Chunsuttiwat S. 2006b. Surveillance for avian influenza in human beings in Thailand. Lancet Infect Dis, 6: 757–758.
Parmley EJ, Bastien N, Booth TF, Bowes V, Buck PA, Breault A, Caswell D, Daoust P-Y, Davies JC, Elahi SM, Fortin M, Kibenge F, King R, Li Y, North N, Ojkic D, Pasick J, Pryor SP, Robinson J, Rodrigue J, Whitney H, Zimmer P, Leighton FA. 2008. Wild bird influenza survey, Canada, 2005. Emerg Infect Dis, 14(1): 84–87.
Pasick J, Berhane Y, Embury-Hyatt C, Copps J, Kehler H, Handel K, Babiuk S, Hooper-McGrevy K, Li Y, Mai Le Q, Lien Phuong S. 2007. Susceptibility of Canada Geese (Branta canadensis) to highly pathogenic avian influenza virus (H5N1). Emerg Infect Dis, 13: 1821–1827.
Slemons RD, Shieldcastle MC, Heyman LD, Bednarik KE, Senne DA. 1991. Type A influenza viruses in waterfowl in Ohio and implications for domestic turkeys. Avian Dis, 35: 165–173.
Webster RG, Bean WJ, Gorman OT, Chambers TM, Kawaoka Y. 1992. Evolution and ecology of influenza A viruses. Microbiol Rev, 56: 152–179.
WHO. 2002. Manual on animal influenza diagnosis and surveillance. World Health Organization (WHO). China, Harbin.
Winkler WG, Trainer DO, Easterday BC. 1972. Influenza in Canada geese. Bull World Health Organ, 47(4): 507–513.
Zhou JY, Shen HG, Chen HX, Tong GZ, Liao M, Yang HC, Liu JX. 2006. Characterization of a highly pathogenic H5N1 influenza virus derived from bar-headed geese in China. J Gen Virol, 87: 1823–1833.


 DownLoad:
DownLoad:








 Email Alerts
Email Alerts RSS Feeds
RSS Feeds