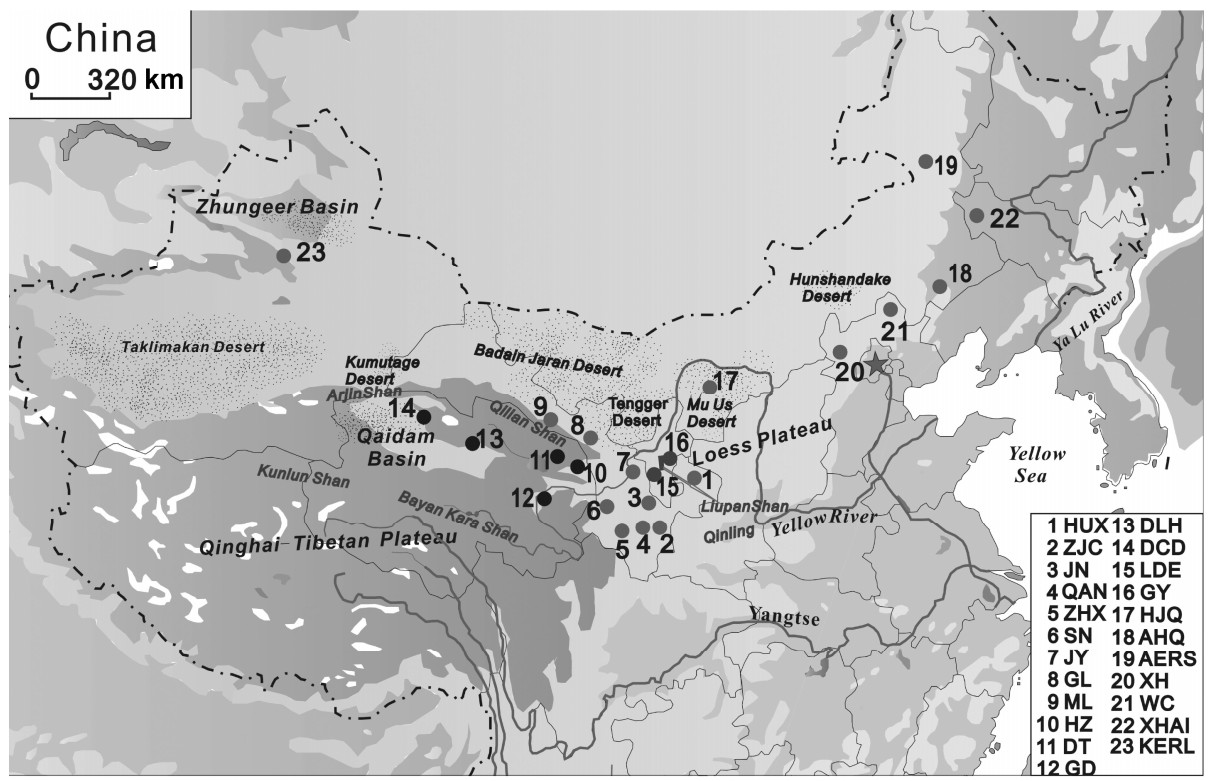
The Daurian Partridge (Perdix dauuricae) is a kind of hunting bird with high economic value. Genetic diversity and structure in the Daurian Partridge were studied by analyzing eight microsatellite loci in 23 populations found throughout the range of the species in China. The objectives were to evaluate the consequences on genetic diversity and differentiation of Daurian Partridge populations and to obtain a profound genetic insight for future management decisions and for effective measures to protect and exploit Daurian Partridges. The results showed that microsatellites were polymorphic in all Daurian Partridge populations, with a high level of genetic diversity over all the loci, especially in the Qaidam Basin populations which have the highest level of diversity. Significant genetic divergence was observed among different groups as well as between populations within the same group; most pairwise FST values were highly significant. Both phylogenetic trees and Bayesian clustering analyses revealed clear differentiation among the 23 populations of the Daurian Partridge, which were classified into two genetically differentiated groups. A bottleneck analysis indicated that Daurian Partridge populations have experienced a recent bottleneck. Our study argues that the Qaidam populations, North China populations, JN population, ZJC population, and Liupan Mountain populations should be paid special attention in order to retain adequate population sizes for maintaining genetic diversity.
The Sulawesi endemic Geomalia (Geomalia heinrichi) is a medium-sized terrestrial bird living in mossy forest at 1700–3500 m a.s.l., mainly 1800–2200 m (Collar, 2005). In some respects it resembles both thrushes (Turdidae) and babblers (Timaliidae/Pellorneidae/Leiotrichidae), and no consensus as to its taxonomic position has been reached to date. The species was discovered by Gerd Heinrich in 1930 in the Latimodjong mountains in central Sulawesi. He made his first observation already on his first day in the field, and initially mistook it for an Island Thrush (Turdus poliocephalus). Throughout the description of his first encounters with the species, he referred to it as a thrush (quoted in Stresemann, 1940). Stresemann (1931) provided a detailed description, pointing out morphological features of which some seemed to indicate that Geomalia was closer to thrushes, and others suggesting that it belonged to the babblers. The early attempts to classify Geomalia by Stresemann(1931, 1940) followed a tradition in which specific characters judged to be more important than others were relied on to determine affinity, rather than synapomorphies. His initial conclusion (Stresemann, 1931) was that he thought that Geomalia was related to the shortwings, particularly Heinrichia calligyna, alternatively best placed in its own monotypic subfamily, Geomaliinae. Later, Stresemann (1940) reconsidered this and placed Geomalia in Timaliinae.
Following Stresemann's initial descriptions and taxonomic conclusions (Stresemann, 1931, 1940), very little primary data have been added. The taxonomic position of this monotypic genus has remained controversial, with opinion swinging between babbler or thrush affinity. Ripley (1952) was the first to disagree with Stresemann (1940), by placing Geomalia in Turdidae, immediately before Zoothera in a linear taxonomy, although without providing any reasons. Subsequent authors have followed either of these two positions. Most major world lists or handbooks have placed Geomalia before Zoothera/Geokichla in Turdidae (Sibley and Monroe, 1990; Dickinson, 2003; Clement, 2000; Clements et al., 2012; Gill and Donsker, 2013). Collar (2005) emphasized the uncertain phylogenetic position of Geomalia by placing it among the first species in Turdinae. Collar(2004, 2005) noted that Geomalia has a distinct juvenile plumage, with spotted underparts, unlike any babbler.
White and Bruce (1986) placed Geomalia in Timaliidae, but mixed up their reference to Stresemann by stating that he placed Geomalia in Timaliidae in 1931, which in fact he did in 1940. They further stated that Stresemann (1931) did not mention a thrush-like juvenile plumage, which is correct in the sense that Stresemann (1931) did not refer to the juvenile plumage as "thrush-like". However, his description, although somewhat vague, stated that the feathers of the breast are edged grey-black in juveniles, not uniformly colored as in the adult (Stresemann, 1931). Holmes and Philipps (1996) and Coates and Bishop (1997) followed White and Bruce (1986) in placing Geomalia in Timaliidae, in the latter case reiterating the incorrect reference to Stresemann (1931).
No cladistic analysis of morphological characters including Geomalia has been undertaken. We here present a hypothesis of the phylogenetic position of Geomaliaheinrichi based on DNA sequences of mitochondrial and nuclear sequences obtained from one of Heinrich's specimens. To the best of our knowledge, this is the first molecular study of the phylogenetic position of Geomalia heinrichi.
Materials and methods
DNA sampling and sequencing
DNA was extracted from feathers and a claw from a juvenile Geomalia heinrichi heinrichi housed in the Museum für Naturkunde der Humboldt-Universität, Berlin, collected in the Latimodjong Mountains, southern Sulawesi on 16 July 1939 by Gerd Heinrich (ZMB 34.160; field number 1018), and from tissue samples of a few additional taxa (Supplementary Table S1).
DNA was extracted using QIA Quick DNEasy Kit (Qiagen, Inc) according to the manufacturer's instructions, but with 30 µL DTT added to the initial incubation step for the extraction from feathers, toepads and claws. For the amplification and sequencing of the fresh samples, we sequenced the mitochondrial cytochrome b (cytb) gene and the entire nuclear myoglobin intron 2 (myo) following the protocols described in Olsson et al.
(2005), and the nuclear ornithine decarboxylase exon 6 (partial), intron 6, exon 7, intron 7 and exon 8 (partial)
(ODC) following the protocols described in Friesen et al. (1999) and Allen and Omland (2003).
We sequenced a few additional taxa, and downloaded some more sequences from GenBank (Supplementary Table S1). Both the cytb gene and the nuclear introns from the museum toepad samples were sequenced in small fragments, using a range of specific primer combinations as described by Irestedt et al. (2006), Dario Zuccon (personal communication) and some primers designed specifically for this study (Supplementary Table S2). All sequences have been submitted to GenBank (Supplementary Table S1).
Phylogenetic analyses
Sequences were aligned using MegAlign 4.03 in the DNAstar package (DNAstar Inc.).
Phylogenies were estimated by Bayesian inference (BI) using MrBayes 3.1.2 (Huelsenbeck and Ronquist, 2001; Ronquist and Huelsenbeck, 2003). All analyses were run under the best-fit models according to the Bayesian Information Criterion (BIC), calculated in jModeltest 0.1.1 (Posada, 2008a, 2008b). The preferred model for the cytb data was the general time-reversible (GTR) model (Lanave et al., 1984; Tavaré, 1986; Rodríguez et al., 1990), assuming rate variation across sites according to a discrete gamma distribution with four rate categories (Γ; Yang, 1994), and an estimated proportion of invariant sites (I; Gu et al., 1995) (GTR + Γ
+ I). For ODC the preferred model was GTR + I, and for myo the HKY (Hasegawa et al., 1985) model + Γ.
The three loci were analysed both separately and concatenated. In the combined BI analysis, the data were divided into locus-specific partitions. In each analysis, four Metropolis-coupled MCMC chains were run for 30 million generations and sampled every 1000 generations. The posterior distributions were summarized as a majority-rule consensus tree. The first 7.5 million generations of each run, well after the chains reached stationarity, were discarded as burn-in.
The concatenated data were also analysed by maximum likelihood bootstrapping (MLBS) and parsimony bootstrapping (MPBS). MLBS (1000 replicates) was conducted with RAxML-HPC2 version 7.3.2 (Stamatakis, 2006; Stamatakis et al., 2008) on the Cipres portal (Miller et al., 2010). The data were partitioned by locus, and as per default GTRCAT was used for the bootstrapping phase, and GTRGAMMA for the final tree inference. The MPBS was performed in PAUP* (Swofford, 2001): heuristic search strategy, 10000 replicates, starting trees obtained by stepwise addition (random addition sequence, 10 replicates), TBR branch swapping.
Myophonus caeruleus, which is part of Muscicapidae, the sister group of Turdidae (Sangster et al., 2010), was used as outgroup in all analyses.
Results
Sequence characteristics
The aligned cytb of 44 taxa comprised 1038 characters, of which 374 (36%) were parsimony informative. For 41 sequences the complete target stretch was available, while 3 included missing data and varied between 976 and 1011 bp in length. We found no unexpected stop codons, indels or distinct double peaks in the chromatograms that would indicate the presence of nuclear pseudogenes (e.g. Sorenson and Quinn, 1998).
The length of the nuclear introns varied depending on multiple indels. The aligned myo sequences of 43 taxa comprised 758 characters, of which 94 (12%) were parsimony informative. For 36 sequences the complete target stretch of 695–713 bp in length was obtained. Nine sequences included missing data. Eight of these varied between 674 and 705 bp in length, one sequence was only 181 bp, and for Ridgwayia pinicola no sequence from this locus was available. Seven indels were shared by two or more taxa, and one of these, a 2 bp deletion, supported the position of Geomalia in the phylogeny (see below; Fig. 1).
Figure
1.
Majority rule (50%) consensus tree based on concatenated nuclear intron 2 of myoglobin and introns 6–7 of ODC and mitochondrial cytochrome b, inferred by Bayesian inference, analysed in three partitions, one per locus. Support values are indicated at the nodes, in the order posterior probability/maximum likelihood bootstrap/parsimony bootstrap; an asterisk represents support 1.00/100%. The shaded rectangle indicates the genus Zoothera. One 2 bp deletion in the myoglobin alignment supporting monophyly of the Zoothera clade including Geomalia heinrichi is indicated by a black bar; a white bar denotes an inferred reversal of this state.
The aligned ODC of 43 taxa comprised 733 characters, of which 104 (14%) were parsimony informative. For 29 sequences the complete target stretch of 690–706 bp in length was available. Fourteen sequences included missing data, the majority varying between 499 and 691 bp in length. The Geomalia sequence was 191 bp, and Ridgwayia pinicola 156 bp. For Zoothera everetti sequence data from this locus was missing.
Single-locus analyses
The trees based on single-locus analyses (hereafter SLAs) varied in resolution. All three SLAs recovered Geomalia as part of the Zoothera clade, with good support (PP 0.92 in the cytb tree; PP 0.97 in the ODC tree and PP 0.99 in the myo tree) (Supplementary Figs. S1–S3).
Concatenated multilocus analyses
The tree based on the concatenated sequences (Fig. 1) was overall congruent with various trees presented by Cibois and Cracraft (2004), Klicka et al. (2005), Nylander et al. (2008), Voelker et al. (2007) and Voelker and Klicka (2008). In agreement with previous studies, the exact positions of some problematic taxa, such as Geokichla sibirica, Ixoreus naevius, Ridgwayia pinicola, Hylocichla mustelinus and Entomodestes leucotis were uncertain to some degree. Geomalia was part of the Zoothera clade with PP 1.00, 95% MLBS and 62% MPBS, although its exact position within the clade was unresolved.
Discussion
The phylogeny is overall relatively well supported and in agreement with previous publications (e.g. Cibois and Cracraft, 2004; Klicka et al., 2005; Nylander et al., 2008; Voelker et al., 2007; Voelker and Klicka, 2008) regarding reference taxa. We are not concerned here with minor differences within clades; all major clades found in earlier works are recovered here, and the position of Geomalia as part of the Zoothera clade is corroborated both by the concatenated data and by all of the SLAs. Within the Zoothera clade, the position of Geomalia is unclear. The present data indicate that it stems from a basal node, suggesting that it may have become isolated on Sulawesi early in the Zoothera radiation. The juvenile plumage of Geomalia shows a dark vertical bar on the rear ear-coverts and a dark malar stripe, and the breast feathers have dark scallops (photos available on www.orientalbirdimages.org), all traits recalling several Zoothera species. It is worth noting that the Borneo endemic Z. everetti resembles Geomalia in having warm orange underparts (albeit with whitish belly), lacking the dark spotting or scalloping typical of the genus Zoothera, and in showing a facial pattern reminiscent of juvenile Geomalia. Juvenile Z. everetti has more heavily scalloped underparts than juvenile Geomalia. It thus appears that the typical Zoothera pattern is retained in juveniles, whereas it has become lost in adults in both these species. The characters that have suggested to some earlier workers that Geomalia is a babbler may be adaptions to its apparently obligate terrestrial life-style. As Geomalia is firmly placed in Zoothera, we propose that it should be assigned to that genus, under the name Zoothera heinrichi.
Acknowledgements
We are grateful to Pascal Eckhoff and Sylke Frahnert at the Museum für Naturkunde der HumboldtUniversität, Berlin for generously granting permission to sample a specimen of Geomalia heinrichi, and Jon Fjeldså for help with the sampling of this specimen and for providing a description of the juvenile plumage and some other comments; and to Jornvall Foundation and the Chinese Academy of Sciences Visiting Professorship for Senior International Scientists (No. 2011T2S04) for other support (both to P.A.).
An B, Zhang LX, Liu NF, Browne S, Ruan LZ, Song S. 2009. Phylogeography of Tibetan snowcock (Tetraogallus tibetanus) in Qinghai-Tibetan Plaetau. Mol Phylogenet Evol, 50: 526–533
Avise JC. 1994. Molecular Markers, Natural History and Evolution. Chapman & Hall, New York
Avise JC, Walker D. 1998. Pleistocene phylogeographic effects on avian populations and the speciation process. Proc R Soc Ser B-Biol, 265: 457–463
Belkhir K, Borsa P, Chikhi L, Raufaste N, Catch F. 2004. GENETIX (version 4.02), Software under WindowsTM for the genetics of the populations Laboratory Genome, Populations, Interactions, CNRS UMR 5000, University of Montpellier Ⅱ, Montpellier, France
Chen Q, Chang C, Liu NF. 1999. Mitochondrial DNA introgression between two parapatric species of Alectoris. Acta Zool Sin, 45(4): 456–463 (in Chinese)
Chen YK, Zhao ZL, Liu NF. 2006. Genetic structure of Przewalski's Rock Partridge (Alectoris magna) population in the Longdong Loess Plateau, China. Biochem Genet, 44: 209–221
Chistiakov DA, Hellemans B, Volckaert FAM. 2006. Microsatellites and their genomic distribution, evolution, function and applications: a review with special reference to fish genetics. Aquaculture, 255: 1–29
Comes HP, Kadereit JW. 1998. The effect of Quaternary climatic changes on plant distribution and evolution. Trends Plant Sci, 3: 432–438
Di Rienzo A, Peterson AC, Garza JC, Valdes AM, Slatkin M, Freimer NB. 1994. Mutational processes of simple-sequence repeat loci in human populations. Proc Natl Acad Sci USA, 91: 3166–3170
El Mousadik A, Petit RJ. 1996. High level of genetic differentiation for allelic richness among populations of the argan tree[Argania spinosa (L. ) Skeels] endemic to Morocco. Theor Appl Genet, 92: 832–839
Excoffier L, Laval G, Schneider S. 2005. ARLEQUIN, version3.0: an integrated software package for population genetics data analysis. Evol Bioinfor Online, 1: 47–50
Frankel OH, Soule ME. 1981. Conservation and Evolution. Cambridge University Press, Cambridge, England
Gaggiotti OE, Excoffier L. 2000. A simple method of removing the effect of a bottleneck and unequal population sizes on pairwise genetic differences. Proc R Soc Lond B, 267: 81–87
Gerloff U, Schlötterer C, Rassmann K, Rambold I, Hohmann G, Fruth B, Tautz D. 1995. Amplification of hypervariable simple sequence repeats (microsatellites) from excremental DNA of wild living bonobos (Pan paniscus). Mol Ecol, 4: 515–518
Goudet J. 2001. FSTAT. A program to estimate and test gene diversities and fixation indices, (Version 2.9.3). Updated from Goudet (1995)
Hamrick JL, Godt MJW. 1989. Allozyme diversity in plant species. In: Brown AHD, Clegg MT, Kahler AL, Weir BS (eds) Plant Population Genetics, Breeding, and Genetic Resources. Sinauer, Sunderland, MA, pp 43–63
Hamrick JL, Godt MJW, Sherman-Broyles SL. 1992. Factors influencing levels of genetic diversity in woody plant species. New Forest, 6: 95–124
Hartl DL, Clark AG. 1997. Principles of Population Genetics. Sinauer, Sunderland, MA
Heuertz M, Hausman JF, Hardy OJ, Vendramin GG, Frascaria-Lacoste N, Vekemans X. 2004. Nuclear microsatellites reveal contrasting patterns of genetic structure between western and southeastern European populations of the common ash (Fraxinus excelsior L. ). Evolution, 58: 976–988
Hewitt GM. 2000. The genetic legacy of the Quaternary ice ages. Nature, 405: 907–913
Hewitt GM. 2004. Genetic consequences of climatic oscillations in the Quaternary. Philos T R Soc Lon B, 359: 183–195
Hou P, Wei M, Zhang LX, Liu NF. 2002. Genetic structure of edge population in Przewalski's rock partridge (Alectoris magna). Acta Zool Sin, 48(3): 333–338 (in Chinese)
Huang ZH, Liu NF. 2007. Phylogeography of rusty-necklaced partridge (Alectoris magna) in northwestern China. Mol Phylogenet Evol, 43: 379–385
Johnsgard PA. 1988. The Quails, Partridges, and Francolins of the World. Oxford University Press, Oxford
Liu NF, Huang ZH, Wen LY. 2004. Subspecies divergence of Przewalski's rock partridge (Alectoris magna) description of a new subspecies. Acta Zootaxon Sin, 29(3): 600–605 (in Chinese)
Liu NF, Wang XT, Luo WY, Chang C. 1988. Avifauna study of the Liupan Mountain in Ningxia. J Lanzhou Univ Nat Sci Ed, 24: 63–75 (in Chinese)
Luikart G, Cornuet JM. 1997. Empirical evaluation of a test for identifying recently bottlenecked populations from allele frequency data. Conserv Biol, 12: 228–237
Madge S, McGowan P. 2002. Pheasants, Partridges and Grouse: A Guide to the Pheasants, Partridges, Quails, Grouse, Guineafowl, Buttonquails and Sandgrouse of the World. Christopher Helm, London
Mantel N. 1967. The detection of disease clustering and a generalized regression approach. Cancer Res, 27: 209–220
Maruyama T, Fuerst PA. 1985. Population bottlenecks and nonequilibrium models in population genetics. Ⅱ. Number of alleles in a small population that was formed by a recent bottleneck. Genetics, 111: 675–689
Michalakis Y, Excoffier L. 1996. A genetic estimation of population subdivision using distance between alleles with special reference to microsatellite loci. Genetics, 142: 1061–1064
Nei M. 1972. Genetic distance between populations. Am Nat, 106: 283–292
Nei M. 1987. Molecular Evolutionary Genetics. Columbia University Press, New York
Nei M, Maruyama T, Chakraborty R. 1975. The bottleneck effect and genetic variability in populations. Evolution, 29: 1–10
Nei M. 1978. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics, 89: 583–590
Prentice HC, Lonn M, Lefkovitch LP, Runyeon H. 1995. Associations between alleles frequencies in Festuca ovina and habitat variation in the alvar grasslands on the Baltic island of Oland. J Ecol, 83: 391–402
Pritchard JK, Stephens M, Donnelly P. 2000. Inference of population structure using multilocus genotype data. Genetics. 155: 945–959
Ramamoorthi J, Thilagam K, Sivaselvam SN, Karthickeyan SMK. 2009. Genetic characterization of Barbari goats using microsatellite markers. J Vet Sci, 10(1): 73–76
Raymond M, Rousset F. 1995a. Genepop version 12: population genetics software for exact tests and ecumenicism. J Hered, 86: 248–249
Raymond M, Rousset F. 1995b. An exact test for population differentiation. Evolution, 49: 1280–1283
Rousset F. 1997. Genetic differentiation and estimation of gene flow from Fstatistics under isolation by distance. Genetics, 145: 1219–1228
Slatkin M. 1987. Gene flow and the geographical structure of natural populations. Science, 236: 787–792
Slatkin M. 1993. Isolation by distance in equilibrium and non-equilibrium populations. Evolution, 47: 264–279
Slatkin M. 1995. A measure of population subdivision based on microsatellite allele frequencies. Genetics, 139: 457–462
Sun HY, Cao ZW, Jiang HL, Yu LG. 1998. Brooding and growth of the younger Perdix dauuricae. Forest Sci Technol, 23(4): 28–30 (in Chinese)
Sun HY, Ge DN, Hong YT, Cao ZW. 1996. Research about domesticating and breeding technology of Daurian Partridge (Perdix dauuricae). Forest Sci Technol, 21(4): 32–35 (in Chinese)
Takezaki N, Nei M. 1996. Genetic distances and reconstruction of phylogenetic trees from microsatellite DNA. Genetics, 144: 389–399
Tautz D. 1989. Hypervariability of simple sequences as a general source for polymorphic DNA markers. Nucleic Acids Res, 17: 6463–6471
Turner PC, McLennan AG, Bates AD, White MRH. 2003. Instant Notes in Molecular Biology, 2nd edn. Science Press, Beijing
Weir BS, Cockerham CC. 1984. Estimating F-statistics for the analysis of population structure. Evolution, 38: 1358–1370
Woodruff DS. 1989. The problems of conserving genes and species. In: Weston D, Pearl M (eds) Conservation for the Twenty First Century. Oxford University Press, New York, pp 76–88
Wright S. 1943. Isolation by distance. Genetics, 28: 114–138
Zhang ZW, Liang W, Sheng G. 1994. Studies on the nest site selection of Daurian partridge. Zool Res, 15(4): 37–43 (in Chinese)
Zhang ZW, Wu YC. 1992. The Daurian Partridge (Perdix dauuricae) in north-central China. Gibier Faune Sauvage, 9: 591–595
Zhang ZW, Liang W. 1997. Breeding ecology of Daurian Partridge (Perdix dauuricae) in Shanxi. Chinese J Zool, 2(2): 23–25 (in Chinese)
Zhao ZJ, Zhang S, Feng KF. 1992. The biology of the Daurian Partridge (Perdix dauuricae suschkini) in northeastern China. Gibier Faune Sauvage, 9: 597–604
Michael A. Weston, Nick Porch, Desley A. Whisson, et al. Do different camera trap lures result in different detection rates of vertebrates because of their attractiveness to invertebrates?. Ecological Management & Restoration, 2024.
DOI:10.1111/emr.12603
2.
Emilia Grzędzicka. Bird Feeder Explorers Are Not Attracted by the Seeds of Invasive Weeds in Winter. Diversity, 2024, 16(2): 81.
DOI:10.3390/d16020081
3.
Christoph Randler, Nadine Kalb. Circadian activity of the fat dormouse Glis glis measured with camera traps at bait stations. Mammal Research, 2021, 66(4): 657.
DOI:10.1007/s13364-021-00583-6


 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds