Gang Feng, Xiongwei Huang, Lingfeng Mao, Na Wang, Xueting Yang, Yanping Wang. 2020: More endemic birds occur in regions with stable climate, more plant species and high altitudinal range in China. Avian Research, 11(1): 17. DOI: 10.1186/s40657-020-00203-y
Citation:
Gang Feng, Xiongwei Huang, Lingfeng Mao, Na Wang, Xueting Yang, Yanping Wang. 2020: More endemic birds occur in regions with stable climate, more plant species and high altitudinal range in China. Avian Research, 11(1): 17. DOI: 10.1186/s40657-020-00203-y
Gang Feng, Xiongwei Huang, Lingfeng Mao, Na Wang, Xueting Yang, Yanping Wang. 2020: More endemic birds occur in regions with stable climate, more plant species and high altitudinal range in China. Avian Research, 11(1): 17. DOI: 10.1186/s40657-020-00203-y
Citation:
Gang Feng, Xiongwei Huang, Lingfeng Mao, Na Wang, Xueting Yang, Yanping Wang. 2020: More endemic birds occur in regions with stable climate, more plant species and high altitudinal range in China. Avian Research, 11(1): 17. DOI: 10.1186/s40657-020-00203-y
Ministry of Education Key Laboratory of Ecology and Resource Use of the Mongolian Plateau & Inner Mongolia Key Laboratory of Grassland Ecology, School of Ecology and Environment, Inner Mongolia University, Hohhot 010021, China
2.
Key Laboratory of Animal Ecology and Conservation Biology, Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, China
3.
Co-Innovation Center for Sustainable Forestry in Southern China, College of Biology and the Environment, Nanjing Forestry University, Nanjing 210037, China
4.
Jiangsu Key Laboratory for Biodiversity and Biotechnology, College of Life Sciences, Nanjing Normal University, Nanjing 210023, China
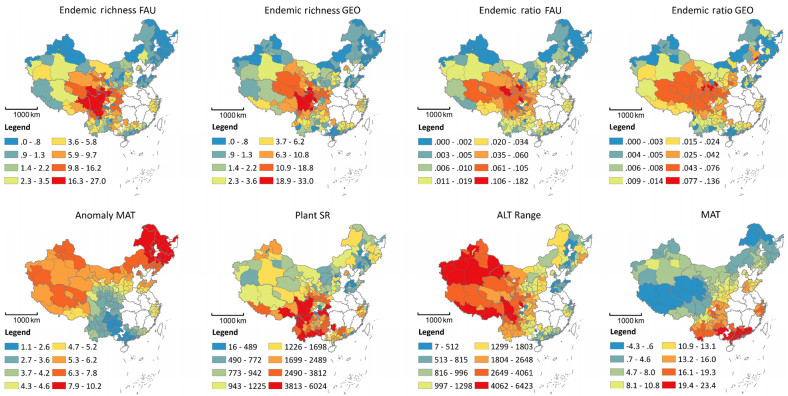
Geographical distribution of endemic species and its multiple scale drivers are an important topic in biodiversity conservation, because these species are especially vulnerable to climate change and habitat degradation, and therefore have high conservation priority. Here, for the first time, we simultaneously linked glacial-interglacial climate change, contemporary climate, plant species richness and altitudinal range with endemic bird distribution in China.
Methods
Ordinary least squares models and simultaneous autoregressive models were used to assess the associations between endemic bird richness, endemic bird ratio and each explanatory variable. Structural Equation Models were also performed to test the direct and indirect effects of these variables on endemic bird richness and endemic bird ratio.
Results
Higher endemic bird richness and endemic bird ratio occurred in regions with stable glacial-interglacial temperature, i.e., southwest China. Plant species richness and altitudinal range were also positively correlated with endemic bird richness and endemic bird ratio. Glacial-interglacial temperature change, contemporary precipitation, plant species richness and altitudinal range were all included in the best combination of variables for endemic bird richness. Importantly, glacial-interglacial temperature change had strong direct effects on both endemic bird richness and endemic bird ratio, while plant species richness only had a direct effect on endemic bird richness.
Conclusions
Our results indicate that endemic birds in China concentrate in southwest regions where there are stable glacial-interglacial temperature, more plant species and larger altitudinal range. Notably, while stable climate has strong direct effects on both endemic bird richness and endemic bird ratio, plant species may affect endemic bird richness through its effect on overall bird species richness. Importantly, the recent anthropogenic activities have also significantly intensified in this region, which would pose huge challenge for biodiversity conservation in China.
Some bird species are known to dismantle materials from old nests for re-nesting (Sedgwick and Knopf, 1988; Kershner et al., 2001) as well partaking in kleptoparasitism of nesting material from active nests (Jones et al., 2007; Slager et al., 2012). The Hair-crested Drongo (Dicrurus hottentottus) tends to reuse materials from failed nests to re-nest, but also exhibits a non-typical behavior by dismantling their own nest after the young have fledged (Li et al., 2009). Li et al. (2009) proposed that nest dismantling was an adaptive behavior to increase fitness by reducing the risk of predation, reducing competition for nesting sites, or both. Such adaptations are complex and sometimes could be explained by other hypotheses.
Parasitic and non-parasitic invertebrates can take refuge in the nests of birds (Woodroffe, 1953). Some of these invertebrates are already present in the vegetation used for nesting material, thus using the vegetation as their refugia and food source; however some will use the birds as their host (Merino and Potti, 1995; Brown et al., 2001) and are introduced into the nest via nesting birds. Often ectoparasites play a negative role on the avian species they utilize (Merino and Potti, 1995; Brown et al., 2001). Birds may avoid inhabiting parasite infested areas, and can develop ways to either eliminate or avoid recruiting parasites (Hart, 1990). The role of parasites has even been responsible for other adaptations such as preening (Marshall, 1981; Clayton, 1991; Cotgreave and Clayton, 1994). In the case of the Haircrested Drongo, the question can be made if this unique nest dismantlement behavior is an adaptation to control parasite populations, which may otherwise have a negative effect in the fitness of the species. Parasite control may be needed since drongos often nest in the same tree and even on the same branch used from previous nesting periods (Li et al., 2009).
To our knowledge, there has never been any formal macro or microscopic evaluations of Hair-crested Drongo nests. Our objectives in this study were to document the presence of parasitic and non-parasitic invertebrates within the nests of Hair-crested Drongos using funneling techniques in a field setting. This study was designed to further the ongoing study of drongo's nest dismantling behavior. Though this study was not designed to test any formal hypotheses, our goal was to provide the basic ground work for future hypothesis testing that could possibly help explain the nest dismantling behavior.
Methodology
Study species and site
This study was conducted at Dongzhai National Nature Reserve (31.95°N, 114.25°E), which is located in the Dabieshan Mountains of Henan Province of central China (Li et al., 2009). The reserve is located in the transitional area of subtropical and temperate zones. The study was conducted at Baiyun Station, which is located within the Reserve. Dongzhai National Nature Reserve is known for its high avian diversity (over 300 species) and was established initially as a bird reserve (Song and Qu, 1996). The Hair-crested Drongo is a common bird throughout much of southeast Asia and is known to breed in central and northern China (Zheng, 2011). The Hair-crested Drongo is a migratory species arriving at the study site to in April, with the breeding season lasting till early August (Li et al., 2009).
Invertebrate/parasite sampling
Invertebrates, including parasites were sampled from collected nests of the Hair-crested Drongos. Nests were removed by cutting their main support branch after the nests became inactive and prior to the nest being dismantled by adults. Upon collection, macroscopic evaluations of ten nests were conducted to determine if any invertebrates were visible to the naked eye. Following macroscopic evaluation, the eight of the collected nests were bagged to make sure there was no outside contamination. Nests were first sampled using the Berlese funneling technique (Berlese, 1905; Nolan, 1955; Brown et al., 2001), with the sampling methodology being modified after two collections because of contamination issues. Originally, nests were put directly under a lamp, with a funnel placed directly underneath the nest to direct invertebrates into a sterile vial containing an alcohol solution (70% ethyl alcohol, 30% H2O) for at least 24 hours, with nests being tapped to dislodge possible invertebrates from the nest and into the funnel. The modified sampling technique entailed taking the bagged nest and securing the funnel and vial through a hole in the bottom of the bag by tape to prevent any escape or entry of invertebrates. These modified funnels were then placed outside in an open area receiving direct sunlight for approximately 36 hours, and were set up so that the majority of this time was during daylight hours. Samples were not collected during rain events or in untypical cloud cover. Vials were labeled for identification and taken back to Beijing Normal University laboratories for evaluation. Because our goal was not to quantify the amount of parasites/invertebrates present, we only documented the presence/absence during macro and microscopic evaluations.
Results
A total of 10 nests were observed macroscopically, and 8 of these were sampled microscopically (2 Berlese, and 6 modified funneling technique). It was immediately evident that the light used in the standard Berlese funneling technique acted as a visual lure (Neethirajan et al., 2007), thus attracted several flying insects and contaminated the first two nest microscopic evaluations. Using the modified funneling technique reduced the contamination by other insects, and using the energy of the sun proved to be a more effective and efficient method under field conditions. Due to high diversity of insects and difficulty identifying each species, collected invertebrates were identified down to the taxonomic class, order, or family level. Macroscopic observation showed that lice (order: Phthiraptera) were present in the majority of the nests, and an unidentified larvae and chrysalis were observed. Microscopic evaluations showed that parasites and other invertebrates were present within the nests (Table 1). Lice, adult and larvae rove beetles (family: Staphylinidae), springtails (class: Collembola), and of ticks (family: Ixodidae) were observed. One nest yielded rove beetles, another separatenest had springtails, and 2 nests contained ticks. Lice were present in all samples.
Table
1.
Presence and frequency of parasitic and non parasitic invertebrates in six nests of Hair-crested Drongos at Dongzhai National Nature Reserve, Henan, China
Nest number
Lice Phthirapteraa
Springtails Collembolaa
Rove Beetles Staphylinidaea
Ticks Ixodidaea
1
√
√
√
2
√
3
√
√
4
√
5
√
6
√
√
Frequency (%)
100
17
17
33
a Taxonomic descriptions represents the closest class, order, or family species that were able to be identified.
Though our research objectives were not to test methodology efficiency, much was learned about the methodology to conduct such research and could benefit similar studies in the future. In this case, modifications had to be made in order to achieve research objectives. The modified funneling technique used in this study could prove to be useful in areas where laboratories are not available, or in areas where local surroundings cannot be controlled. Our modified funneling technique shows that such research can be done using limited resources coupled with natural resources (i.e., sunlight) to produce results. Though the use of sunlight for funneling techniques is not a new concept (Bondy, 1940), this is the first time such an application has been used for investigating invertebrate and avian nest relationships.
We found that several parasitic and non-parasitic invertebrates do inhabit the nests of Hair-crested Drongos. Rove beetles belong to the family Staphylinidae, which is one of the largest and most diverse beetle families in the world and are known to inhabit the nests of many different taxa (see Klimaszewski et al., 1996). Rove beetles have been found to play different roles in avian nests depending on the beetle species (Majka et al., 2006). Certain rove beetles will feed on the vegetative material found in nests while some will feed on other invertebrates, including parasites, which in turn could be beneficial to bird species by reducing parasitic loads (Majka et al., 2006). The proportion of rove beetles in the nests of Hair-crested Drongos found in this study (17%) is comparable to the proportion found in the nests of Great Tits (Parus major) (13%) (Heeb et al., 2000). Because of the high diversity found in Staphylindae coupled with the lack of this type of research in this region, the possibility of the rove beetle species detected in this study being new unidentified species is likely, or could exhibit new geographic extensions of certain species. Majka et al. (2006) found 14 different beetle species in owl nests in Nova Scotia, Canada, many of which were new records demonstrating geographic range extensions, and some were first time recordings from bird nests. Majka et al. (2006) also found that 8 of the 14 species beetles found belonged to the family Staphylinidae.
Springtails were another invertebrate detected within the nests of Hair-crested Drongos. Pung et al. (2000) found that springtails were found in 5% of Red-cockaded Woodpecker (Picoides borealis) nests. Though our results show a higher frequency of springtails (17%), this could be attributed to our low sample size, geographic variation, or could further demonstrate the variability of arthropod community structure within avian nests. Furthermore, Pung et al. (2000) found a lower frequency of springtails but detected 11 other types of arthropods, including six different types of mites, thus having a higher diversity of arthropods than what was found in our study.
The presence of parasites, such as lice and ticks, has been well documented to have host relationships with avian species, however most researches focus on the presence of such species that use the actual bird species as a host rather than the in the nest. It is important to understand what effect these invertebrates have on certain aspects of avian biology such as the nest site selection, adult survivorship, and overall fitness of a species. Lice have shown to reduce the survival in Feral Pigeons (Colomba livia) by increasing the energy needed for thermoregulation (Clayton et al., 1999). Parasites have played a role in the adaptation of preening behavior of birds, in which preening serves as a way to control harmful ectoparasites (Marshall, 1981; Clayton, 1991; Cotgreave and Clayton, 1994). The presence of parasites can cause the avoidance of nesting sites (Chapman, 1973; Brown and Brown, 1986; Loye and Carroll, 1991) and can even cause nest and nestling abandonment (Duffy, 1983; Calyton and Moore, 1997). The frequency of lice on live hosts or in nests is species specific (both avian and louse). Rozsa et al. (1996) found that five species of lice were found on both Hooded Crows (Corvus corone cornix) and Rooks (C. frugilegus), however the frequency was different between species with 53% of Hooded Crows and 92% of Rooks being infested. Furthermore, Rozsa et al. (1996) found Rooks to have higher lice richness, diversity, and loads, in which they contribute some of these differences to be associated with the community structure of these species with rooks being colonial and Hooded Crows being more solitary. Pung et al. (2000) only found louse in 2% of Red-cockaded Woodpecker nests. All of these findings discussed indicate the variability and complexity of invertebrate-avian host relationships.
Several studies have shown that territory may be more of a factor in nest site selection rather than decreased nest quality (see Loye and Carroll, 1998). In the case of the Hair-crested Drongo, the main supporting hypothesis as to explain the nest-dismantling behavior is that this species dismantles its nest to reduce competition for breeding sites and to increase species fitness. Having ectoparasitic present in the nests of Haircrested Drongos certainly supports the plausibility that other behaviors, such as nest-dismantling could have been adapted to control such parasites. Even though, we do not propose the nest dismantling behavior is strictly driven by the presence of parasitic and non-parasitic invertebrates, we do suggest it may be a surrogate factor, and could warrant further investigation. Further investigations into whether or not parasites in the nests have any effects on nest site selection, rate of nest dismantlement, and fitness of this species would further benefit what we know about the Hair-crested Drongo.
Acknowledgements
We appreciate the support by Dongzhai National Nature Reserve for conducting this study. Funding and support was provided by National Science Foundation East Asia Pacific Summer Institute (EAPSI), Chinese Ministry of Science and Technology, and China Science and Technology Exchange Center. Further funding and support was provided by Alabama A & M University and Beijing Normal University. Thanks to Luis BIANCUCCI, Matthew LEROW, Peng ZHANG, Chang GAO, Ji LUO, Yang LIU, and Langyu GU, for their assistance in and out of the field.
Bivand R, Altman M, Anselin L, Assunção R, Berke O, Bernat A, et al. spdep: Spatial dependence: weighting schemes, statistics and models. R package version 0.5-92. 2015. .
Chen C, Zeng D, Zhao Y, Wu Y, Xu J, Wang Y. Correlates of extinction risk in Chinese endemic birds. Avian Res. 2019;10:8.
Davies TJ, Buckley LB, Grenyer R, Gittleman JL. The influence of past and present climate on the biogeography of modern mammal diversity. Philos T Roy Soc B. 2011;366:2526–35.
Dynesius M, Jansson R. Evolutionary consequences of changes in species' geographical distributions driven by milankovitch climate oscillations. P Natl Acad Sci USA. 2000;97:9115–20.
Eiserhardt WL, Borchsenius F, Plum CM, Ordonez A, Svenning J-C. Climate-driven extinctions shape the phylogenetic structure of temperate tree floras. Ecol Lett. 2015;18:263–72.
Essl F, Rabitsch W, Dullinger S, Moser D, Milasowszky N. How well do we know species richness in a well-known continent? Temporal patterns of endemic and widespread species descriptions in the European fauna. Glob Ecol Biogeogr. 2013a;22:29–39.
Essl F, Moser D, Dirnböck T, Dullinger S, Milasowszky N, Winter M, et al. Native, alien, endemic, threatened, and extinct species diversity in European countries. Biol Conserv. 2013b;164:90–7.
Feng G, Mao L, Sandel B, Swenson NG, Svenning J-C. High plant endemism in China is partially linked to reduced glacial-interglacial climate change. J Biogeogr. 2016;43:145–54.
Feng G, Mao L, Benito BM, Swenson NG, Svenning J-C. Historical anthropogenic footprints in the distribution of threatened plants in China. Biol Conserv. 2017;210:3–8.
Feng G, Ma Z, Sandel B, Mao L, Normand S, Ordonez A, et al. Species and phylogenetic endemism in angiosperm trees across the Northern Hemisphere are jointly shaped by modern climate and glacial–interglacial climate change. Glob Ecol Biogeogr. 2019;28:1393–402.
Fjeldså J, Lovett JC. Geographical patterns of old and young species in African forest biota: the significance of specific montane areas as evolutionary centres. Biodivers Conserv. 1997b;6:325–46.
Gao W. Studies on Birds and Their Ecology in Northeast China. Beijing: Science Press; 2006 (in Chinese).
Goldewijk KK, Beusen A, van Drecht G, de Vos M. The HYDE 3.1 spatially explicit database of human-induced global land-use change over the past 12, 000 years. Glob Ecol Biogeogr. 2011;20:73–86.
Hasumi H, Emori S. K-1 coupled gcm (miroc) description. Tokyo: Center for Climate System Research, University of Tokyo; 2004.
Hawkins BA, DeVries PJ. Tropical niche conservatism and the species richness gradient of North American butterflies. J Biogeogr. 2009;36:1698–711.
He F, Li S, Zhang X, Ge Q, Dai J. Comparisons of cropland area from multiple datasets over the past 300 years in the traditional cultivated region of China. J Geogr Sci. 2013;23:978–90.
He F, Li S, Zhang X. A spatially explicit reconstruction of forest cover in China over 1700-2000. Glob Planet Change. 2015;131:73–81.
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A. Very high resolution interpolated climate surfaces for global land areas. Int J Climatol. 2005;25:1965–78.
Jetz W, Rahbek C, Colwell RK. The coincidence of rarity and richness and the potential signature of history in centres of endemism. Ecol Lett. 2004;7:1180–91.
Lei F, Qu Y, Lu J, Liu Y, Yin Z. Conservation on diversity and distribution patterns of endemic birds in China. Biodivers Conserv. 2003;12:239–54.
Lei F, Qu Y, Song G, Alström A, Fjeldså J. The potential drivers in forming avian diversity hotspots in the East Himalaya Mountains of Southwest China. Integr Zool. 2015;10:171–81.
Liang C, Feng G, Si X, Mao L, Yang G, Svenning J-C, et al. Bird species richness is associated with phylogenetic relatedness, plant species richness, and altitudinal range in Inner Mongolia. Ecol Evol. 2018;8:53–8.
López-Pujol J, Zhang F, Ge S. Plant biodiversity in China: richly varied, endangered, and in need of conservation. Biodivers Conserv. 2006;15:3983–4026.
Lu L, Mao L, Yang T, Ye J, Liu B, Li HL, et al. Evolutionary history of the angiosperm flora of China. Nature. 2018;554:234–8.
Maas B, Putra DD, Waltert M, Clough Y, Tscharntke T, Schulze CH. Six years of habitat modification in a tropical rainforest margin of Indonesia do not affect bird diversity but endemic forest species. Biol Conserv. 2009;142:2665–71.
Myers N, Mittermeier RA, Mittermeier CG, Da Fonseca GAB, Kent J. Biodiversity hotspots for conservation priorities. Nature. 2000;403:853–8.
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, Minchin PR, et al. vegan: Community ecology package. 2015. .
Orme CDL, Davies RG, Burgess M, Eigenbrod F, Pickup N, Olson VA, et al. Global hotspots of species richness are not congruent with endemism or threat. Nature. 2005;36:1016–9.
Otto-Bliesner BL, Brady EC, Clauzet G, Tomas R, Levis S, Kothavala Z. Last glacial maximum and Holocene climate in CCSM3. J Clim. 2006;19:2526–44.
R Core Team. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; 2016.
Rosseel Y. lavaan: an R package for structural equation modeling. J Stat Softw. 2012;48:1–36.
Sandel B, Arge L, Dalsgaard B, Davies RG, Gaston KJ, Sutherland WJ, et al. The influence of late quaternary climate-change velocity on species endemism. Science. 2011;334:660–4.
Scharlemann JPW, Green RE, Balmford A. Land-use trends in Endemic Bird Areas: global expansion of agriculture in areas of high conservation value. Glob Chang Biol. 2004;10:2046–51.
Stein A, Gerstner K, Kreft H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol Lett. 2014;17:866–80.
Hothorn T, Bretz F, Westfall P. Package "multcomp". 2013. .
Wiens JJ, Donoghue MJ. Historical biogeography, ecology and species richness. Trends Ecol Evol. 2004;19:639–44.
Wu Y, DuBay SG, Colwell RK, Ran J, Lei F. Mobile hotspots and refugia of avian diversity in the mountains of south-west China under past and contemporary global climate change. J Biogeogr. 2017;44:615–26.
Yang L. The Avifauna of Yunnan China. Non-Passeriformes, vol. I. Kunming: Yunnan Science and Technology Press; 1995 (in Chinese).
Yang L, Yang X. The Avifauna of Yunnan China, vol. II. Passeriformes. Kunming: Yunnan Science and Technology Press; 2004 (in Chinese).
Young KA. Habitat diversity and species diversity: testing the competition hypothesis with juvenile salmonids. Oikos. 2001;95:87–93.
Zhang J, Kissling WD, He F. Local forest structure, climate and human disturbance determine regional distribution of boreal bird species richness in Alberta. Canada. J Biogeogr. 2013;40:1131–42.
Zheng GM. A checklist on the classification and distribution of the birds of China. Beijing: Science Press; 2017 (in Chinese).
Gang Feng, Ying-Jie Xiong, Hua-Yu Wei, et al. Endemic medicinal plant distribution correlated with stable climate, precipitation, and cultural diversity. Plant Diversity, 2023, 45(4): 479.
DOI:10.1016/j.pld.2022.09.007
2.
Xiongwei Huang, Congtian Lin, Liqiang Ji, et al. Species inventories from different data sources “shaping” slightly different avifauna diversity patterns. Frontiers in Ecology and Evolution, 2023, 11
DOI:10.3389/fevo.2023.1121422
3.
Handy Erwin Pier Leimena, Achmad Sjarmidi, Tati Suryati Syamsudin. IDENTIFICATION OF THE MOLUCCAN MEGAPODE (Eulipoa wallacei) NATURAL HABITAT IN HARUKU ISLAND, INDONESIA AND ITS VEGETATION COMPOSITION. BIOTROPIA, 2023, 30(2): 158.
DOI:10.11598/btb.2023.30.2.1778
4.
Lorenzo Peruzzi, David Dolci, Alessandro Chiarucci. Potential climatic and elevational range shifts in the Italian narrow endemic Bellevalia webbiana (Asparagaceae) under climate change scenarios. Nature Conservation, 2022, 50: 145.
DOI:10.3897/natureconservation.50.91265
5.
Flávio Mariano Machado Mota, Neander Marcel Heming, José Carlos Morante-Filho, et al. Amount of bird suitable areas under climate change is modulated by morphological, ecological and geographical traits. Frontiers in Ecology and Evolution, 2022, 10
DOI:10.3389/fevo.2022.987204
6.
Hongyan Yao, Pengcheng Wang, Nan Wang, et al. Functional and phylogenetic structures of pheasants in China. Avian Research, 2022, 13: 100041.
DOI:10.1016/j.avrs.2022.100041
7.
Berenika Mioduszewska, Alice M I Auersperg, Mark O’Hara. Treasure islands: foraging ecology and the emergence of tool use in wild Goffin’s cockatoos. Current Opinion in Behavioral Sciences, 2022, 45: 101118.
DOI:10.1016/j.cobeha.2022.101118
8.
Brittany T. Trew, Ilya M. D. Maclean, Brian McGill. Vulnerability of global biodiversity hotspots to climate change. Global Ecology and Biogeography, 2021, 30(4): 768.
DOI:10.1111/geb.13272
9.
Şerban Procheş, Syd Ramdhani, Alice C. Hughes, et al. Southeast Asia as One of World’s Primary Sources of Biotic Recolonization Following Anthropocene Extinctions. Frontiers in Ecology and Evolution, 2021, 9
DOI:10.3389/fevo.2021.634711
Table
1.
Presence and frequency of parasitic and non parasitic invertebrates in six nests of Hair-crested Drongos at Dongzhai National Nature Reserve, Henan, China
Nest number
Lice Phthirapteraa
Springtails Collembolaa
Rove Beetles Staphylinidaea
Ticks Ixodidaea
1
√
√
√
2
√
3
√
√
4
√
5
√
6
√
√
Frequency (%)
100
17
17
33
a Taxonomic descriptions represents the closest class, order, or family species that were able to be identified.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds