Oscar Humberto Marín-Gómez, Michelle García-Arroyo, Camilo E. Sánchez-Sarria, J. Roberto Sosa-López, Diego Santiago-Alarcon, Ian MacGregor-Fors. 2020: Nightlife in the city: drivers of the occurrence and vocal activity of a tropical owl. Avian Research, 11(1): 9. DOI: 10.1186/s40657-020-00197-7
Citation:
Oscar Humberto Marín-Gómez, Michelle García-Arroyo, Camilo E. Sánchez-Sarria, J. Roberto Sosa-López, Diego Santiago-Alarcon, Ian MacGregor-Fors. 2020: Nightlife in the city: drivers of the occurrence and vocal activity of a tropical owl. Avian Research, 11(1): 9. DOI: 10.1186/s40657-020-00197-7
Oscar Humberto Marín-Gómez, Michelle García-Arroyo, Camilo E. Sánchez-Sarria, J. Roberto Sosa-López, Diego Santiago-Alarcon, Ian MacGregor-Fors. 2020: Nightlife in the city: drivers of the occurrence and vocal activity of a tropical owl. Avian Research, 11(1): 9. DOI: 10.1186/s40657-020-00197-7
Citation:
Oscar Humberto Marín-Gómez, Michelle García-Arroyo, Camilo E. Sánchez-Sarria, J. Roberto Sosa-López, Diego Santiago-Alarcon, Ian MacGregor-Fors. 2020: Nightlife in the city: drivers of the occurrence and vocal activity of a tropical owl. Avian Research, 11(1): 9. DOI: 10.1186/s40657-020-00197-7
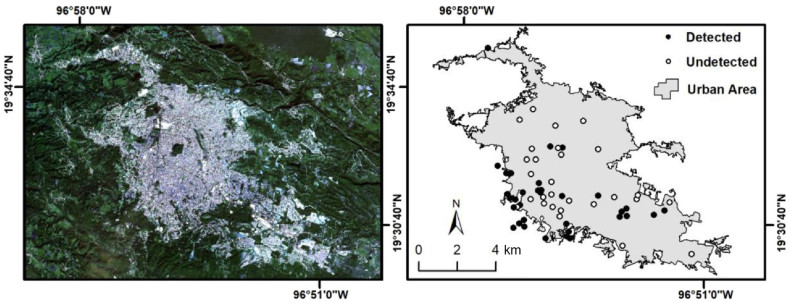
Cities differ from non-urban environments by the intensity, scale, and extent of anthropogenic pressures, which can drive the occurrence, physiology, and behavior of the organisms thriving in these settings. Traits as green cover often predict the occurrence patterns of bird species in urban areas. Yet, anthropogenic noise and artificial light at night (ALAN) could also limit the presence and disrupt the behavior of birds. However, there is still a dearth of knowledge about the influence of urbanization through noise and light pollution on nocturnal bird species ecology. In this study, we assessed the role of green cover, noise, and light pollution on the occurrence and vocal activity of the Mottled Owl (Ciccaba virgata) in the city of Xalapa (Mexico).
Methods
We obtained soundscape recordings in 61 independent sites scattered across the city of Xalapa using autonomous recording units. We performed a semi-automated acoustic analysis of the recordings, corroborating all Mottled Owl vocalizations. We calculated two measures of anthropogenic noise at each study site: daily noise (during 24 h) and masking noise (mean noise amplitude at night per site that could mask the owl's vocalizations). We further performed generalized linear models to relate green cover, ALAN, daily noise, and masking noise in relation to the owl's occurrence (i.e., detected, undetected). We also ran linear models to assess relationships among the beginning and ending of vocal activity with ALAN, and with the anthropogenic and masking noise levels at the moment of which vocalizations were emitted. Finally, we explored variations of the vocal activity of the Mottled Owl measured as vocalization rate across time.
Results
The presence of Mottled Owls increased with the size of green cover and decreased with increases in both artificial light at night and noise levels. At the temporal scale, green cover was positively related with the ending of the owl's vocal activity, while daily noise and ALAN levels were not related to the timing and vocal output (i.e., number of vocalizations). Furthermore, the Mottled Owl showed a marked peak of vocal activity before dawn than after dusk. Although anthropogenic noise levels varied significantly across the assessed time, we did not find an association between high vocal output during time periods with lower noise levels.
Conclusions
Spatially, green cover area was positively related with the presence of the Mottled Owl in Xalapa, while high noise and light pollution were related to its absence. At a temporal scale, daily noise and ALAN levels were not related with the timing and vocal output. This suggests that instead of environmental factors, behavioral contexts such as territoriality and mate interactions could drive the vocal activity of the Mottled Owl. Further studies need to incorporate a wider seasonal scale in order to explore the variation of different vocalizations of this species in relation to environmental and biological factors.
Our current understanding is that the Helmeted Guineafowl flock is basically a group of birds that remain together for at least one breeding season and that members follow a dominant male during foraging in the non-breeding season (dry season, May to September in the Krugersdorp Game Reserve). One function of this behavior is to maximize the discovery of underground food resources that occur in patches (Van Niekerk, 1979, 1980, 2002; Crowe and Elbin, 1987). Ecological constraints are lifted as rain falls (October in the Krugersdorp Game Reserve) and the members engage in sequential mate selection activities. Monogamous pairs form and males protect their females and chicks (Crowe and Elbin, 1987). According to Elbin (1979) the prime reproductive system is the family unit (mother, father and offspring) as revealed by studies conducted on free-ranging Domestic Guineafowl in Pennsylvania (USA). In short, behavioral studies have concentrated on seasonal events and the reproductive system of Helmeted Guineafowl (Skead, 1962; Elbin, 1979), while this paper focuses on the behavioral patterns of individuals and how they contribute to the maintenance of the flock. More specifically the purposes of this paper are: 1) to describe the cyclic re-grouping of members within the flock to maintain cohesion and flock stability and 2) to describe patterns of co-operation within the flock that have not been the focus of previous studies.
Study area and methods
This study was conducted in the 1400-ha Krugersdorp Game Reserve (KGR) (26°05′11″S, 27°42′35″E), in Gauteng Province, South Africa. From March to July 1981, seven flocks (8–15 members) in a designated area were trapped using walk-in funnel traps. A total of 52 Helmeted Guineafowl were fitted with metal rings and numbered patagial tags, representing at least 40% of all members in each flock (Van Niekerk, 1983, 2009). Adults were sexed through cloacal examination (Siopes and Wilson, 1973). These flocks were visited weekly from April 1982 to March 1983 and then monthly from April 1983 to February 1984. With the aid of a mounted telescope all marked guineafowl were noted on standard maps while the author traversed a fixed route during late afternoons, totaling ~150 hours of observations.
Periods of 45–60 min were spent in close proximity to one flock that was selected randomly for intensive study during each visit from April 1982 to March 1983, totaling about 60 hours of observations (Van Niekerk, 2009). In late afternoons the study flock was mainly located in a small area (400 m2), which made it possible to place permanent beacons that would allow for the estimation of distances between members. The association time between individuals and all interactive behavioral patterns (such as pecking) were recorded. Individuals were regarded as associated when they foraged close to one another (30–100 cm apart with no other guineafowl between them) for 60 s or more. Social spread (m) is the direct distance between the two furthest members through the flock (excluding breeding pairs). Where information was lacking, facts were gleaned from a similar unpublished study of a flock of 10 Helmeted Guineafowl that visited a private garden near Constantia (33°59′45″S, 18°26′06″E) in Cape Town. This study (January 1992 to May 1994) was done by Ereka van der Merwe, who distinguished individuals on the basis of casque size and shape (Prinsloo et al., 2005). Females and males were distinguished on the basis of fights (only males engage in fights) and calls (only females emit a "buckwheat" call). Agonistic displays refer to chases, pecks, humpbacked postures and head-bobbing by high ranking males in the direction of a low ranking member or incoming conspecific intruder.
Results
Cyclic events
During the breeding season (October–March) the seven flocks remained in small separate breeding territories (0.5–1 ha), but joined neighboring flocks during the non-breeding season (April–September) to form larger flocks (~30 individuals) that foraged across a larger area (71 ha). Juveniles from the different flocks were pooled from April onwards, which was a period when flocks moved further away from cover as their offspring grew from chicks to the juvenile stage (Siegfried, 1966). This was necessary since food resources had been depleted (Van Niekerk, 2002; Van Niekerk, unpublished). Despite the apparent joining of neighboring flocks, the marked flocks returned to their respective roosts during the late afternoon. In fact, the colored patagial tags revealed that the separate flocks remained in their respective subgroups even while they were with the larger congregated flock.
Flock size and structure
The mean monthly size of all flocks in the study was 7.75 (SD = 3.02, n = 33, range = 7–14) (Van Niekerk, 1985). The study flock basically consisted of three adult males (A, B and C), three adult females (D, E and F), four to six offspring and two non-permanent females that sometimes visited the flock for two or three days at a time. At least two other flocks had two or more adult males and females, but during the 1981/82 breeding season not one marked flock succeeded in raising more than one brood of chicks.
Social ranking and spatial position of members in the study flock
Male A was the highest ranking male because 1) he was at the top of the pecking order (Table 1), 2) he initiated foraging direction and the rest of the flock followed him, 3) he was the first to court and breed in two consecutive breeding seasons and 4) he was the member that initiated action to chase intruding guineafowl from other flocks that had moved into the breeding territory (Van Niekerk, 2002; Van Niekerk, unpublished). During the non-breeding season (April–October), the other two males remained close to him, with females and offspring further behind. The non-breeding male often ran towards male A and performed a submissive display in front of male A (Van Niekerk, 1985).
Table
1.
Number of agonistic interactions of members of the study flock arranged in a sociogram
Females usually remained between the adult males and their offspring. No rank order was observed among females. The non-breeding female was often observed in a tree above the flock from where she emitted a "buckwheat" call, which is a typical female call (Maier, 1982).
Close-knit unit
The study flock remained a permanent group and members were often found close together throughout the study period. The social spread of the flock was 25–40 m during the period before the chicks arrived and 5–10 m just after their arrival (Fig. 1). In fact, there was a significant negative correlation between social spread and mean flock size (r = –0.79, p < 0.01). The shorter distance during the brooding phase (January–February) was a direct consequence of clustering around the chicks (1–10 days old). The chicks moved along with the flock, often directly beneath the adults and made body contact with various adults that emitted a soft "swe swe swe" contact call. When approached by humans (n = 10), the Black-backed Jackal (Canis mesomelas) (n = 6) and the Savanna Baboon (Papio ursinus) (n = 8), some adult members scattered in different directions while the entire flock produced an inferno of panic calls to distract the intruders (Maier, 1982). The chicks also scattered and hid in nearby grass. As the chicks reached juvenile age, they formed a separate subgroup right behind the flock, about 1–3 m away. This distance increased as they reached sub-adulthood during the non-breeding season (April–September). This was the time when a troop of baboons was seen killing one sub-adult and probably more, as the number of sub-adults dwindled from six to three during the first year. The 1981/82 offspring remained with the flock for at least 18 months.
Figure
1.
Social spread correlated with mean group sizes of the study flock based on four observations per month
The study flock did not dissolve when male A and female D paired off to breed nearby for a period of ~28 days during December 1981, who remained only about 20 m from the area where the rest of the flock usually foraged. In fact, the remainder of the flock continued to forage in a territory of ~0.5 ha around the nest. Male A was not regularly observed in the flock during incubation, but did arrive from time to time without his female, who was incubating their eggs. During the 1981/82 breeding season male B and female E paired off to produce chicks after A and D returned to the flock with their chicks in January 1982. However, this second brood perished within one month.
A comparison of the agonistic displays by the two high ranking males (A and B) towards members or intruders and low ranking members (such as humpbacked displays and chases) demonstrates that B was less agonistic than A in the presence of A. Once A paired off to breed and moved further away from the flock, B showed more agonistic displays and assumed the role of dominant male. The rest of the flock clustered around him during the absence of A. The position of A was restored once he returned to the flock in March, when both males were present together again (Fig. 2).
Figure
2.
Alternate frequencies of agonistic displays by the two highest ranking males in the study flock based on 45 hours of observations, totaling 62 displays
The study flock remained together during at least two breeding seasons up to March 1983. The chicks of the second breeding season joined the offspring from the previous year in a subgroup. Female D disappeared during March 1982, but was replaced by a new female during the following breeding season. The Constantia group showed a similar degree of stability over a two-year period.
Associations within the study flock
Table 2 demonstrates that adult males clustered together just prior to and during the first part of the breeding season. In fact, when intruding conspecifics approached these males stood together side by side and moved together in the direction of the imposters. These bonds became less prevalent during the period from the arrival of the chicks to the non-breeding season (Table 3), when the high ranking males spent more time with the chicks (Table 4). Breeding females (D and E) spent more time with their own males than with the non-breeding male (respectively χ2 = 656.7, df = 2, p < 0.01 and χ2 = 1201.1, df = 2, p < 0.01; Table 3). This is expected in a monogamous mating system. However, the second ranking male (male B) spent more time with his female than the highest ranking male (male A) did during flocking (Table 3). The non-breeding male spent most of his time with the higher ranking males (A and B) and to a less extent with the breeding females (D and E) (χ2 = 502.47, df = 3, p < 0.01), but did not engage in any courtship behavior (Table 3). Similarly, the non-breeding female (F) spent most of her time with the higher ranking males and also did not perform any courtship behavior. Her association with the breeding females (D and E) was virtually non-existent (Tables 3) (χ2 = 631.6, df = 3, p < 0.01).
Table
2.
Association patterns of adult members of the study flock in % and time (min) from September 1982 to January 1983
Table
3.
Association patterns of adult members of the study flock in % and time (min) from February 1983 to May 1983 while observed in the flock (pairs were absent from time to time)
Table 4 shows the time flock members spent with male A's chicks. The mother (D) was the least caring (Table 4). The father (A) and the second ranking male (B) spent the most time with the chicks. The non-breeding male and female spent more time with the chicks than the breeding females (χ2 = 205.8, df = 5, p < 0.01). In the Constantia flock, an adult male member, which was not the father, often spread his wings over the chicks to shield them from intruders such as dogs and humans. In the same flock, the off-spring of the breeding female from the previous year helped with incubation when the mother got up to feed.
Leadership succession
Male A remained the highest ranking male for at least two breeding seasons and produced chicks during both. He was never challenged (involved in physical battles) by a male within the flock. However, in the Constantia flock, a new male and female joined the original flock from the nearby Kirstenbosch Botanical Gardens. When they joined, a bloody physical battle that lasted for days ensued between the new male and the original highest ranking male. Blood oozed from their bare facial parts as they pecked at each other's wattles. The new male did not succeed and remained on the periphery of the flock during the next 10 months. During the start of the next breeding season, the new male launched a renewed attack on the highest ranking male and succeeded. The old male and his female retired to the side of the flock while the new male started initiating foraging direction with the rest of the flock following.
Discussion and conclusions
The study flock was not a number of Helmeted Guineafowl that associated randomly. The members remained around the dominant male and followed him as he initiated foraging. Although a pecking order could be established among males, the frequency of agonistic actions was very low, which suggests that agonistic interactions are limited to ensure cohesion. The function of the pecking order was not to dispel adult members from the flock but simply to maintain a rank hierarchy. The second highest ranking male took center stage with the rest of the flock associated around him in order to maintain the cohesive nature of the flock when the dominant male and his female left to breed. This alternate responsibility was important to prevent eviction from the territory or a take-over from neighboring dominant males. The two high ranking males foraged away from the females during the non-breeding season, but still in the same flock, in order to repel conspecific intruders, but during the breeding season they spent more time with their females (Table 3). In other words, it was a monogamous mating system. No polygamy or promiscuity was recorded; that is, the two high ranking males did not flirt with each other's females or flirt with females that joined from neighboring flocks. Prolonged squabbles for adult females could lead to flock instability. There was no evidence of sequential mate selection. Flirting by paired males was recorded by Elbin et al. (1986) elsewhere.
On arrival of parents and chicks, the highest ranking male spent more time with his chicks than the mother, who spent relatively little time with her chicks (Table 4). This is ascribed to the females foraging actively to restore bodily condition after breeding (Crowe and Elbin, 1987). During this time, the adult non-breeding male and female spent more time with the chicks of the dominant male than their own mother did (Table 4). Since the history of the flock was not known, these helpers could be offspring from a previous year since they were at least two years old when observations started. In the Constantia study, it was demonstrated that offspring from a previous year helped their mother with incubation.
The small social spread during the brooding stage illustrates that most members form a close-knit unit to protect the chicks. It could be argued that formation of a cluster is a response to an urge to associate closely when predators close in, because chicks are easy prey (Fig. 1). However, the fact that all adults 1) helped to distract predators, 2) emitted soft "swe swe swe" contact calls and 3) associated closely with the chicks, illustrates that co-operative nurturing indeed took place.
Unlike free ranging Domestic Guineafowl, where the family is the primary reproductive and brooding unit (Elbin, 1979), this study shows that wild Helmeted Guineafowl base their survival on co-operation in a flock with a number of adult Helmeted Guineafowl. Some adults did not pair off to breed, whereas breeding appears to be the prerogative of the highest ranking male, followed by the second highest ranking male. However, intrinsic/extrinsic triggers could determine how many guineafowl pairs form from a flock (Van Niekerk, unpublished). Extrinsic factors could be predator pressure and rainfall, while intrinsic factors could include how many offspring remained in the flock from the previous year (cf. Crowe, 1978b).
Among Crested Dendroperdix sephaena and Coqui Peliperdix coqui Francolins, the mother and father produce a brood and when their offspring reach sub-adulthood (7–8 months old) they are repelled from the covey (Van Niekerk, 2001; Van Niekerk and Van Ginkel, 2003). Among Red-necked Pternistis afer and Swainson's P. swainsonii Spurfowls, the father occupies a territory and adult females that enter are courted and inseminated. The mother raises her chicks without the assistance of the father, but the chicks are also repelled when they are about seven to eight months old (Van Niekerk et al., 2009; Van Niekerk, unpublished). In comparison, Helmeted Guineafowl live in flocks and their offspring remain in the flock for at least 18 months. Pairs break away from the flock to breed and return to the flock with their chicks. The non-breeding flock members assist in raising the chicks.
Although Helmeted Guineafowl live in flocks, the breeding pairs actually fare less well than Coqui or Crested Francolin pairs in terms of reproductive efficiency. Efficiency is the mean number of juveniles from a brood expressed as a percentage of the mean clutch size. The mean clutch sizes are 12 for Helmeted Guineafowl, 5.8 for Swainson's Spurfowl, 4.8 for Crested Francolin and 5.0 for Coqui Francolin (Hockey et al., 2005). Reproductive efficiency is 38.2% for Helmeted Guineafowl, 36.6% for Swainson's Spurfowl, 41.8% for Crested Francolin and 44.0% for Coqui Francolin: χ2 = 45.5, df = 3, p < 0.01 (Van Niekerk, unpublished).
Clearly, if the flock is compared to the family system of the francolins, it is not necessarily advantageous for a higher juvenile output. In fact, flocking as a protective unit strategy becomes less convincing if one considers that in the case of the francolins only two adults look after their offspring, while in the case of spurfowls only the mother raises the chicks. However, unlike the francolins and spurfowls, Helmeted Guineafowl move widely in open country where the vigilance potential of more members minimizes surprise attacks and so enhances chick survival (Treisman, 1975). Secondly, flocking is an effective strategy to repulse conspecific intruders from encroaching on their established territory. This last point became especially evident when the non-breeding members remained in the territory and protected it during the absence of the highest ranking male.
To summarize, cooperation within the flock was revealed by: 1) males that joined to repel intruders, 2) the second highest ranking male taking center stage in the flock while the highest ranking male was absent, 3) shared brooding functions among adult members and 4) predator evasion, especially when chicks were nurtured jointly.
Aronson MFJ, La Sorte FA, Nilon CH, Katti M, Goddard MA, Lepczyk CA, et al. A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc R Soc B. 2014;281:20133330.
Canário F, Leitão AH, Tomé R. Predation attempts by Short-eared and Long-eared Owls on migrating songbirds attracted to artificial lights. J Raptor Res. 2012;46:232-4.
Chace JF, Walsh JJ. Urban effects on native avifauna: a review. Landsc Urban Plan. 2006;74:46-69.
Clarke JA. Moonlight's influence on predator/prey interactions between Short-eared Owls (Asio tlammeus) and deermice (Peromyscus maniculatus). Behav Ecol Sociobiol. 1983;13:205-9.
Crawley MJ. The R Book. Chichester: Wiley; 2012.
Da Silva A, Valcu M, Kempenaers B. Light pollution alters the phenology of dawn and dusk singing in common European songbirds. P Roy Soc B. 2015;370:20140126.
Debrot AO. Nocturnal foraging by artificial light in three Caribbean bird species. J Caribb Ornithol. 2014;27:40-1.
Dice LR. Minimum intensities of illumination under which owls can find dead prey by sight. Amer Naturalist. 1945;79:385-416.
Dominoni DM, Greif S, Nemeth E, Brumm H. Airport noise predicts song timing of European birds. Ecol Evol. 2016;6:6151-9.
Dykstra C, Simon M, Daniel F, Hays J. Habitats of suburban barred Owls (Strix varia) and red-shouldered hawks (Buteo lineatus) in Southwestern Ohio. J Raptor Res. 2012;46:190-200.
Enríquez PL. Los búhos neotropicales: Diversidad y conservación. México: El Colegio de la Frontera Sur; 2015.
Falfán I, MacGregor-Fors I. Woody neotropical streetscapes: a case study of tree and shrub species richness and composition in Xalapa. Madera bosques. 2016;22:95-110.
Falfán I, Muñoz-Robles CA, Bonilla-Moheno M, MacGregor-Fors I. Can you really see 'green'? Assessing physical and self-reported measurements of urban greenery. Urban For Urban Gree. 2018;36:13-21.
Fischer JD, Schneider SC, Ahlers AA, Miller JR. Categorizing wildlife responses to urbanization and conservation implications of terminology. Conserv Biol. 2015;29:1246-8.
Francis CD, Ortega CP, Cruz A. Noise pollution changes avian communities and species interactions. Curr Biol. 2009;19:1415-9.
Fröhlich A, Ciach M. Noise shapes the distribution pattern of an acoustic predator. Curr Zool. 2017;64:575-83.
Fröhlich A, Ciach M. Noise pollution and decreased size of wooded areas reduces the probability of occurrence of Tawny Owl Strix aluco. Ibis. 2018;160:634-46.
Fröhlich A, Ciach M. Nocturnal noise and habitat homogeneity limit species richness of owls in an urban environment. Environ Sci Pollut R. 2019;26:1-8.
Fuller RA, Warren PH, Gaston KJ. Daytime noise predicts nocturnal singing in urban robins. Biol Lett. 2007;3:368-70.
Galeotti P. Territorial behaviour and habitat selection in an urban population of the tawny owl Strix aluco L. Boll Zool. 1990;57:59-66.
Gaston KJ, Bennie J. Demographic effects of artificial nighttime lighting on animal populations. Environ Rev. 2014;22:323-30.
Gaston KJ, Bennie J, Davies TW, Hopkins J. The ecological impacts of nighttime light pollution: a mechanistic appraisal. Biol Rev. 2013;88:912-27.
Gerhardt RP, Gerhardt DM, Flatten CJ, González NB. The food habits of sympatric Ciccaba Owls in Northern Guatemala. J Field Ornithol. 1994;65:258-64.
González-Oreja JA. Relationships of area and noise with the distribution and abundance of songbirds in urban greenspaces. Landsc Urban Plan. 2017;158:177-84.
Gorenzel WP, Salmon TP. Characteristics of American Crow urban roosts in California. J Wildl Manag. 1995;59:638-45.
Grimm NB, Faeth SH, Golubiewski NE, Redman CL, Wu J, Bai X, et al. Global change and the ecology of cities. Science. 2008;319:756-60.
Gryz J, Krauze-Gryz D. Changes in the tawny owl Strix aluco diet along an urbanisation gradient. Biologia. 2019;74:279-85.
Hardouin LA, Robert D, Bretagnolle V. A dusk chorus effect in a nocturnal bird: support for mate and rival assessment functions. Behav Ecol Sociobiol. 2008;62:1909.
Himsworth CG, Jardine CM, Parsons KL, Feng AYT, Patrick DM. The characteristics of wild rat (Rattus spp.) populations from an inner-city neighborhood with a focus on factors critical to the understanding of rat-associated zoonoses. PLoS ONE. 2014;9:e91654.
Hindmarch S, Elliott JE. A specialist in the city: the diet of barn owls along a rural to urban gradient. Urban Ecosyst. 2015;18:477-88.
Hölker F, Wolter C, Perkin EK, Tockner K. Light pollution as a biodiversity threat. Trends Ecol Evol. 2010;25:681-2.
Holt DW, Berkley R, Deppe C, Enríquez-Rocha P, Petersen JL, Rangel-Salazar JL, et al. Mottled Owl (Ciccaba virgata). In: del Hoyo J, Elliott A, Sargatal J, Christie DA, de Juana E, editors. Handbook of the Birds of the World Alive. Barcelona: Lynx Edicions; 2019.
Howell S, Webb S. A guide to the birds of Mexico and northern Central America. Oxford: Oxford University Press; 1995.
INEGI. Prontuario de la información geográfica municipal de los Estados Unidos Mexicanos. Xalapa, Veracruz de Ignacio de la Llave. Clave geoestadística. 2009:30087.
Isaac B, White J, Ierodiaconou D, Cooke R. Response of a cryptic apex predator to a complete urban to forest gradient. Wildlife Res. 2013;40:427-36.
Isaksson C, Rodewald AD, Gil D. Behavioral and ecological consequences of urban life in birds. Front Ecol Evol. 2018;6:50.
Kettel EF, Gentle LK, Quinn JL, Yarnell RW. The breeding performance of raptors in urban landscapes: a review and meta-analysis. J Ornithol. 2018;159:1-18.
Knight E, Hannah K, Foley G, Scott C, Brigham R, Bayne E. Recommendations for acoustic recognizer performance assessment with application to five common automated signal recognition programs. Avian Conserv Ecol. 2017;12:14.
Kotler BP, Brown JS, Hasson O. Factors affecting gerbil foraging behavior and rates of owl predation. Ecology. 1991;72:2249-60.
Liu Z, He C, Zhou Y, Wu J. How much of the world's land has been urbanized, really? A hierarchical framework for avoiding confusion. Landsc Ecol. 2014;29:763-71.
Lloyd H. Population densities of some nocturnal raptor species (Strigidae) in southeastern Peru. J Field Ornithol. 2013;74:376-80.
Longcore T, Rich C. Ecological light pollution. Front Ecol Environ. 2004;2:191-8.
Lövy M, Riegert J. Home range and land use of urban Long-eared Owls. Condor. 2013;115:551-7.
Luther D, Gentry K. Sources of background noise and their influence on vertebrate acoustic communication. Behaviour. 2013;150:1045-68.
Manzanares Mena L, Macías Garcia C. Songbird community structure changes with noise in an urban reserve. J Urban Ecol. 2018;4:1-8.
Marín-Gómez OH, MacGregor-Fors I. How early do birds start chirping? Dawn chorus onset and peak times in a Neotropical city. Ardeola. 2019;66:327-41.
Marín-Gómez OH, Toro Y, López-García MM, Garzón-Zuluaga JI, Santa-Aristizabal DM. First records of the Spectacled Owl (Pulsatrix perspicillata) in urban areas, with notes on reproduction. North-West J Zool. 2017;13:368-71.
Marín-Gómez OH, Santiago-Alarcon D, Dátillo W, MacGregor-Fors I. Where has the city choir gone? Loss of the temporal structure of bird dawn choruses in urban areas. Landsc Urban Plan. 2020;194:103665.
Maxwell SL, Fuller RA, Brooks TM, Watson JEM. Biodiversity: the ravages of guns, nets and bulldozers. Nature. 2016;536:143-5.
Menq W, Anjos L. Habitat selection by owls in a seasonal semi-deciduous forest in southern Brazil. Braz J Biol. 2015;75:143-9.
Merchant ND, Fristrup KM, Johnson MP, Tyack PL, Witt MJ, Blondel P, et al. Measuring acoustic habitats. Methods Ecol Evol. 2015;6:257-65.
Mori E, Bertolino S. Feeding ecology of long-eared owls in winter: an urban perspective. Bird Study. 2015;62:257-61.
Mori E, Menchetti M, Ferretti F. Seasonal and environmental influences on the calling behaviour of Eurasian Scops Owls. Bird Study. 2014;61:277-81.
Odom KJ, Mennill DJ. Vocal duets in a nonpasserine: an examination of territory defence and neighbour-stranger discrimination in a neighbourhood of barred owls. Behaviour. 2010;147:619-39.
Patón D, Romero F, Cuenca J, Escudero JC. Tolerance to noise in 91 bird species from 27 urban gardens of Iberian Peninsula. Landsc Urban Plan. 2012;104:1-8.
Penteriani V, Delgado M. The dusk chorus from an owl perspective: eagle owls vocalize when their white throat badge contrasts most. PLoS ONE. 2009;4:e4960.
Penteriani V, Delgado MD, Stigliano M, Campioni L, Sánchez M. Owl dusk chorus is related to the quality of individuals and nest-sites. Ibis. 2014;156:892-5.
Poppleton M. Urban raptors: owl and hawk adaptation to urban centers. JUST. 2016;4:49-60.
Priyadarshani N, Marsland S, Castro I. Automated birdsong recognition in complex acoustic environments: a review. J Avian Biol. 2018;49:e01447.
R Core Team. R: A language and environment for statistical computing[Internet]. Vienna, Austria: R Foundation for Statistical Computing; 2018. .
Ralph CJ, Geupel GR, Pyle P, Martin TE, DeSante DF. Handbook of field methods for monitoring landbirds. Albany: U.S.D.A., Forest Service, Pacific Southwest Research Station. Gen. Tech. Rep. PSW-GTR-144; 1996.
Ranazzi L, Manganaro A, Ranazzi R, Salvati L. Woodland cover and Tawny Owl Strix aluco density in a Mediterranean urban area. Biota. 2000;1:27-34.
Rebolo-Ifrán N, Tella JL, Carrete M. Urban conservation hotspots: predation release allows the grassland-specialist burrowing owl to perform better in the city. Sci Rep. 2017;7:3527.
Restrepo-Cardona JS, Betancur López A, Cano Castaño N. Abundancia y nuevos registros de búhos simpátricos en Manizales y Villamaría (Caldas, Colombia). Bol Cient Mus His Nat. 2015;19:220-9.
Rivera-Rivera E, Enríquez PL, Flamenco-Sandoval A, Rangel-Salazar JL. Ocupación y abundancia de aves rapaces nocturnas (Strigidae) en la Reserva de la Biosfera Selva El Ocote, Chiapas. México. Rev Mex Biodiv. 2012;83:742-52.
Rullman S, Marzluff JM. Raptor presence along an urban-wildland gradient: influences of prey abundance and land cover. J Raptor Res. 2014;48:257-72.
Santiago-Alarcon D, Delgado VC. Warning! Urban threats for birds in Latin America. In: MacGregor-Fors I, Escobar-Ibáñez JF, editors. Avian ecology in Latin American Cityscapes. Cham: Springer; 2017. p. 125-42.
Saufi S, Ravindran S, Hamid NH, Abidin CMRZ, Ahmad H, Ahmad AH, et al. Diet composition of introduced Barn Owls (Tyto alba javanica) in urban area in comparison with agriculture settings. J Urban Ecol. 2020;6:1-8.
Scobie CA, Bayne EM, Wellicome TI. Influence of human footprint and sensory disturbances on night-time space use of an owl. Endanger Species Res. 2016;31:75-87.
Senzaki M, Yamaura Y, Francis CD, Nakamura F. Traffic noise reduces foraging efficiency in wild owls. Sci Rep. 2016;6:30602.
Serieys LE, Bishop J, Okes N, Broadfield J, Winterton DJ, Poppenga RH, et al. Widespread anticoagulant poison exposure in predators in a rapidly growing South African city. Sci Total Environ. 2019;666:581-90.
Seto KC, Fragkias M, Güneralp B, Reilly MK. A Meta-analysis of global urban land expansion. PLoS ONE. 2011;6:e23777.
Ševčík R, Riegert J, Šindelář J, Zárybnická M. Vocal activity of the Central European Boreal Owl population in relation to varying environmental conditions. Ornis Fennica. 2019;96:1-12.
Shonfield J, Bayne EM. The effect of industrial noise on owl occupancy in the boreal forest at multiple spatial scales. Avian Conserv Ecol. 2017;12:13.
Shonfield J, Heemskerk S, Bayne EM. Utility of automated species recognition for acoustic monitoring of Owls. J Raptor Res. 2018;52:42-55.
Sierro J, Schloesing E, Pavón I, Gil D. European blackbirds exposed to aircraft noise advance their chorus, modify their song and spend more time singing. Front Ecol Evol. 2017;5:68.
Slabbekoorn H. Songs of the city: noise-dependent spectral plasticity in the acoustic phenotype of urban birds. Anim Behav. 2013;85:1089-99.
Sol D, Lapiedra O, González-Lagos C. Behavioural adjustments for a life in the city. Anim Behav. 2013;85:1101-12.
Vázquez-Pérez JR, Enríquez PL. Factores temporales y ambientales asociados a los llamados de los búhos en la Reserva Selva El Ocote, Chiapas, México. Hornero. 2016;31:83-8.
Weaving MJ, White JG, Isaac B, Cooke R. The distribution of three nocturnal bird species across a suburban-forest gradient. Emu. 2011;111:52-8.
Wildlife Acoustics. . Accessed 20 May 2019.
Williams-Linera G. Vegetación de bordes de un bosque nublado en el Parque Ecológico Clavijero, Xalapa, Veracruz, México. Rev Biol Trop. 1993;41:443-53.
Zuur AF, Leno EN, Elphick CS. A protocol for data exploration to avoid common statistical problems. Methods Ecol Evol. 2010;1:3-14.
Table
3.
Association patterns of adult members of the study flock in % and time (min) from February 1983 to May 1983 while observed in the flock (pairs were absent from time to time)

 DownLoad:
DownLoad:







 Email Alerts
Email Alerts RSS Feeds
RSS Feeds