
Figure
1.
Examples of combinations of experimental host nests used in this study.
| Citation: |
Giampiero Sammuri, Guido Alari Esposito, Giuseppe Anselmi, Francesco Pezzo, Vincenzo Rizzo Pinna, Andrea Sforzi, Flavio Monti. 2025: Moving away from home: Early life movements and space use in juvenile Montagu's Harriers (Circus pygargus) from central Italy. Avian Research, 16(1): 100239. DOI: 10.1016/j.avrs.2025.100239

|
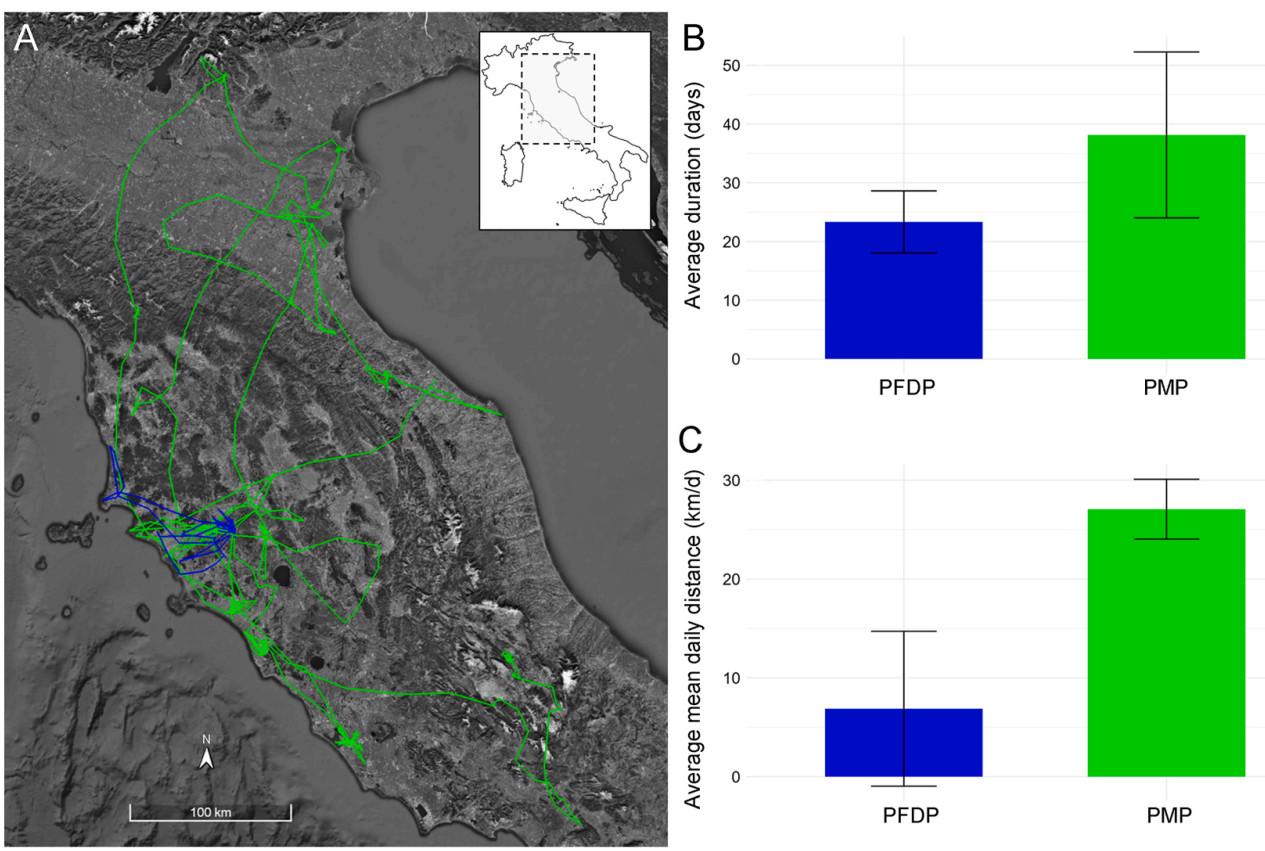
Assessing individual differences and variability in animal movement patterns is essential to improve our understanding of the evolution and ontogeny of migratory strategies. In long-distance migratory species, fledged juveniles often rely on an extremely restricted time span to learn the essential skills for survival and to prepare for migration, possibly the most risky phase of their lives. Collecting detailed information on the dynamics of the movements during the crucial pre-migratory phase is hence essential to understand the solutions developed by migratory species in different environmental contexts. Here, we used high-resolution GPS/GSM transmitters to collect information on the movement ecology of seven juvenile Montagu's Harriers (Circus pygargus) born in central Italy, investigating their early life stages, namely the post-fledging dependence period (PFDP) and the pre-migratory phase (PMP), until autumn migration. After fledging, individuals showed high variability (both in space and time) in home range size, daily distances covered (6.88 ± 11.44 km/day), distance from the nest (1.45 ± 2.8 km) and PFDP length (23.3 ± 5.3 days). Residence time at the natal site significantly decreased, while time interval between revists in the natal area significantly increased, as the PFDP progressed. During the PMP, explored areas and distance from the nest (max value up to 320.8 km) varied among individuals, despite daily distances covered (27 ± 40 km/day) and time allocation between traveling (60.7%) and foraging (39.3%) were similar across individuals. The PMP lasted 38 ± 14 days. Land cover composition of foraging locations was mostly represented by agricultural lands (~78.2%), though habitat use differed among individuals. More than 76% of such locations were outside protected areas. This individual-based tracking study represents a novel approach that improves previous knowledge based on field studies on the early life stages of the Montagu's Harrier. High inter-individual variability in movement patterns, broad-range exploratory movements and foraging locations outside the protected area network make the application of standard conservation measures difficult, raising concerns about the long-term preservation of this vulnerable migratory species in Italy.
Brood parasites, such as the Common Cuckoo (Cuculus canorus, hereafter 'cuckoo'), lay their eggs in the nests of other birds and impose the cost of parental care to the host (Davies, 2000; Feeney et al., 2014; Soler, 2014). Following incubation, cuckoo eggs typically hatch before the host's own eggs and the newly hatched cuckoo evicts the host's eggs and/or nestlings (Davies, 2000). A cuckoo chick's survival prospects are severely diminished if it is forced to cohabit and compete with the host's own chicks (Rutila et al., 2002; Grim et al., 2009). This highlights that accurate timing of egg deposition by the adult cuckoo and early hatching of the cuckoo's egg are important adaptations that underpin successful brood parasitism.
Cuckoo females usually parasitize in the early stage of egg-laying (0–3 eggs) (Moskát and Honza, 2002; Moskát et al., 2009; Honza et al., 2020; Wang et al., 2020a, b). Wyllie (1981) found that the majority of cuckoo eggs were laid during the egg laying period (92%, 83/90), and observed no cases of parasitism after incubation began. Similarly, parasitism of the Reed Warbler (Acrocephalus scirpaccus) shows that no parasitism occurred before or after laying, and all parasitic eggs (N = 45) were laid during the laying stage of the host (Davies and Brooke, 1988). It can be seen from the above studies that cuckoos prefer to parasitize in the early laying stage of the host, which may be beneficial for the hatching of parasitic eggs. Further field observations indicate that cuckoo eggs that were laid before the Great Reed Warbler (Acrocephalus arundinaceus) began to incubate hatch earlier than host eggs. In contrast, one-third of the cuckoo eggs that were laid after the host eggs began to hatch did not have this advantage (Geltsch et al., 2016). In the Great Reed Warbler, the fourth egg is considered as a dividing line such that cuckoo eggs laid prior to it being laid often hatch, while those laid after are less likely to be successful (Geltsch et al., 2016). Therefore, for the cuckoo, being able to sense the number of eggs in the host's nest, or being able to know the host's egg-laying stage (early or late), plays a key role in the success of the parasitism.
Selection should favor brood parasites that identify and lay in host nests that maximize their likelihood of successful parasitism. Indeed, it is well-established that brood parasites choose nests based off a variety of cues, such as nest location (Øien et al., 1996), host behavior (Banks and Martin, 2001), host group size (Feeney et al., 2013), and the degree to which the parasite's eggs resemble those of the host (Yang et al., 2016, 2017; Ma and Liang, 2021). Likewise, the results of numerous field-based monitoring efforts near unanimously report that brood parasites preferentially lay in nests during the early-stages of the host's laying cycle (e.g. Mermoz and Reboreda, 1999; Moskát et al., 2006; Honza et al., 2020) and that eggs laid too early or late tend to be unsuccessful (Langmore et al., 2003). Interestingly, while laying synchrony between the parasite and host is key to successful parasitism, only one recent study (Jelínek et al., 2014) has provided observational data in support of this.
At our research site, cuckoos mainly use the Oriental Reed Warbler (Acrocephalus orientalis) as its primary host. Previous work found that cuckoo females can always parasitize at the appropriate time (1–3 eggs in the nest) (Yang et al., 2016, 2017; Wang et al., 2020a, b, 2021), so we speculate that the cuckoo can evaluate the situation of the target nest, including whether it contains eggs or nestlings, which suggests that they know the stage of the nest (early or late), and finally choose the right time for parasitism. In this study, to verify our speculation, we experimentally investigated whether Common Cuckoos preferentially target Oriental Reed Warbler nests at an earlier nesting stage, as well as whether they preferentially target nests earlier in the egg-laying cycle. As our previous work indicates that female cuckoos preferentially target hosts early in their egg-laying cycle (see Wang et al., 2020a, b), we set comparison of nests with chicks compared to nests with eggs and hypothesized that cuckoos preferentially target nests that contain eggs rather than chicks, and that they also target nests with fewer eggs than more eggs as this suggests that they are earlier in the egg-laying cycle.
This study was performed in Zhalong National Nature Reserve (46°48′–47°31′ N, 123°51′–124°37′ E) located in Heilongjiang Province, Northeast China. Field experiments were carried out during the breeding season (June to August) between 2017–2020. At our study site, habitats are primarily reed swamps, and to our knowledge, the Common Cuckoo only parasitizes the Oriental Reed Warbler (clutch size: 4–6 eggs) (Wang et al., 2020a; b, 2021). Our previous work indicates that parasitism rates are high, ranging from 34.3% to 65.5% per year (Yang et al., 2016, 2017).
We systematically searched for warbler nests in the reed beds by monitoring the activities of adult birds during the breeding season. Nests were monitored daily to ascertain the date that the first egg was laid, clutch size, parasitism status and nest fate. Once a warbler laid the first egg in their nest, two additional artificial nests were set up approximately 1 m away from the host nest and assigned to one of three experimental groups: 1) two and four model eggs placed in the experimental nests (Fig. 1; group 1; N = 55); 2) four model eggs and two model nestlings (Fig. 1; group 2; N = 30); and 3) two model eggs and two model nestlings (Fig. 1; group 3; N = 23). Experimental nests were old warbler nests that we collected in the field during the previous year after breeding had finished, and we combined them using twine (as per Wang et al., 2020a). During experiments these nests were placed nearby host nests (Fig. 1) and slightly higher than the host's nest, as cuckoos preferentially target more visible nests (Patten et al., 2011). We camouflaged the camera with reed leaves and wrapped it in plastic to avoid rain damage. Model eggs (length = 24.18 ± 0.50 mm; width = 21.56 ± 0.63 mm; mass = 7.64 ± 0.13 g) and chicks (length = 20.93 ± 0.42 mm; width = 16.01 ± 0.35 mm; mass = 5.86 ± 0.17 g) were made of a white synthetic clay (Fig. 1). Experimental nests were monitored daily for six days to record whether a parasitism event occurred. All nests were also video recorded from dawn until dusk each day during the experiment. We used mini digital cameras (Uniscom-T71, 70 mm × 26 mm × 12 mm; Mymahdi Technology Co. Ltd., Shenzhen, China) to capture cuckoo behaviour at host nests. To achieve longer battery life, the camcorders were equipped with external power supply (20,000 mAh, Romoss-Sense 6; Romoss Technology Co. Ltd., Shenzhen, China).
During experiments, all parasitism events occurred during the host's laying period, we only observed parasitism of the experimental nests, and no hosts abandoned their nests during the experiments. Three example video recordings showing cuckoo laying behavior in group 1 (Appendix Video S1), group 2 (Appendix Video S2), and group 3 (Appendix Video S3), are provided as supplementary files.
Supplementary video related to this article can be found at https://doi.org/10.1016/j.avrs.2022.100042
Binomial tests (two-tailed test) were used to test for the choice of nests by cuckoo females. For all experimental nests, if one parasitism event at a particular nest was recorded, the cuckoo egg was removed, and the original experimental status was re-established to ensure that all combinations of nests were parasitized with similar probability. Difference was considered to be significant at p < 0.05, highly significant at p < 0.01, and not significant at p > 0.05. Statistical analysis was conducted using IBM SPSS Version 25.0.
For group 1, 55 pairs of nests were set up, 32 of which were parasitized (58%). Cuckoos were significantly more likely to parasitize the nests containing 2 eggs (25 of 32, 78%) compared to 4 eggs (7 of 32, 22%) (Binomial test, p = 0.002; Fig. 2; group 1). For group 2, 30 nests were set up, 14 of which were parasitized (47%). We found that cuckoos were not more likely to parasitize nests containing 4 eggs (10 of 14, 71%) compared to 2 nestlings (4 of 14, 29%) (Binomial test, p = 0.180; Fig. 2; group 2). For group 3, 23 nests were set up, 12 of which were parasitized (52%). Cuckoos were significantly more likely to parasitize the nests containing 2 eggs (10 of 12, 83%) compared to 2 nestlings (2 of 12, 17%) (Binomial test, p = 0.039; Fig. 2; group 3).
Our results show that Common Cuckoo females preferentially parasitized nests with incomplete clutches of eggs rather than nestlings or complete clutches. This result suggests that cuckoos are able to evaluate which nests are more suitable for parasitism based on nesting stage and the number of eggs within the host nest. While observational data near unanimously indicate that brood parasites target nests that maximize incubation synchrony between their own and host's eggs (e.g., Moskát et al., 2006; Honza et al., 2020), to our knowledge these data comprise the first experimental evidence that cuckoos show preferences for nests at an earlier nesting stage and earlier in the laying cycle.
Videos of our experiments provide direct insights into the decision-making process that cuckoos face when choosing a nest to lay in. For instance, when posed with a choice between nests containing either two or four eggs, we found that one cuckoo initially landed on the rim of the nest with four eggs and after approximately 10 s of inspecting the nest contents, moved across to the nest with two eggs before laying (Appendix Video S1). In another instance, the cuckoo initially approached the nest containing two nestlings, but moved across to the nest with two eggs before laying (Appendix Video S3). Although specifically selected, these videos appear to provide direct evidence of the cuckoos assessing nest contents prior to laying their egg, preferring nests with eggs over chicks and nests with fewer eggs compared to more eggs in a manner that would maximize their likelihood of synchronizing the incubation of the own egg with that of their host. For the model nestling experimental treatment, the results were a little not satisfying our expectations in group 2, which may be caused by fewer samples in the fieldwork.
Overall, our study provides experimental results supporting the widely held presumption that cuckoos, like other brood parasites, strategically choose nests to maximize synchrony with their hosts, and highlights the key role that information and strategic decisions play in underpinning successful brood parasitism.
WL and LW designed the study, LW, GH and YZ carried out field experiments, LW and WEF wrote the draft manuscript, and WL involved in discussion and improved the manuscript. All authors read and approved the final manuscript.
The experiments comply with the current laws of China. Fieldwork was performed with permission (No. ZL-GZNU-2019-06) from Zhalong National Nature Reserve, Heilongjiang, China. Experimental procedures were in accordance with the Animal Research Ethics Committee of Guizhou Normal University (No. GZNUECEE-2022-001).
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
We are grateful to Wenfeng Wang and Jianhua Ma for their assistance with fieldwork. We also thank the Zhalong National Nature Reserve for their support and permission to carry out this study. This work was supported by the National Natural Science Foundation of China (Nos. 31960105 to LW, and 31970427 to WL) and the specific research fund of the Innovation Platform for Academicians of Hainan Province. LW was funded by the Guizhou Natural Science Foundation (No. ZK[2022]-316).
|
Allen, A.M., Singh, N.J., 2016. Linking movement ecology with wildlife management and conservation. Front. Ecol. Evol. 3, 155.
|
|
Arroyo, B.E., Bretagnolle, V., 2000. Evaluating the long-term effectiveness of conservation practices in Montagu's Harrier Circus pygargus. In: Chancellor, R.D., Meyburg, B.-U. (Eds.), Raptors at Risk. Pica Press, Bodmin, Cornwall, pp. 403-408.
|
|
Barbraud, C., Kato, A., Trehou, G., Dugué, H., Musseau, R., 2024. Factors affecting time activity budgets of breeding White Storks Ciconia ciconia assessed using bio-logging. ARDEOLA 71, 321-335.
|
|
Bartoń, K., 2020. MuMIn: multi-model inference. R package version.
|
|
Berthold, P., 1996. Control of Bird Migration. Chapman and Hall, London, United Kingdom.
|
|
Bildstein, K.L., 2006. Migrating Raptors of the World: Their Ecology and Conservation. Cornell University Press, Ithaca, NY, U.S.A.
|
|
Brichetti, P., Fracasso, G., 2018. The Birds of Italy: Anatidae-Alcidae; Edizioni Belvedere, Milano, Italy.
|
|
Cramp, S., Simmons, K.E.L., 1980. Handbook of the Birds of Europe, the Middle East and North Africa. The Birds of the Western Palearctic, Vol. 2. Oxford University Press, Oxford.
|
|
Egea-Casas, O., López-López, P., Álvarez, E., Cortone, G., Galán, M., Iglesias-Lebrija, J.J., et al., 2023. Similar post-fledging behavior observed in wild and reintroduced juvenile Bonelli's Eagle in two Mediterranean islands. Ornithol. Appl. 125, duad037.
|
|
Fox, J., Weisberg, S., 2019. An R Companion to Applied Regression, third ed. Sage, Thousand Oaks, CA.
|
|
Garrido, J.R., Numa, C., Barrios, V., Qninba, A., Riad, A., Haitham, O., et al., 2021. The Conservation Status and Distribution of the Breeding Birds of Prey of North Africa. IUCN, Gland, Switzerland. xvi + 102pp.
|
|
Giovacchini, P., 2003. L'avifauna del Sito di Importanza Comunitaria "Monte Labbro e Alta Valle dell'Albegna" (Grosseto, Toscana meridionale). Atti Soc. Tosc. Sci. Nat. Res. B 110, 71-82.
|
|
Jahn, A.E., Bejarano, V., Cueto, V.R., Di Giacomo, A.S., Fontana, C.S., 2017. Movement ecology research to advance conservation of South America's grassland migratory birds. Perspect. Ecol. Conserv. 15, 209-215.
|
|
Klaassen, R.H.G., Schlaich, A.E., Both, C., Bouten, W., Koks, B., 2019. Individual variation in home range size reflects different space use strategies in a central place foraging raptor bird. In: Schlaich, A.E. (Ed.), Migrants in Double Jeopardy. Doctoral Thesis. University of Groningen, The Netherlands.
|
|
Limiñana, R., Soutullo, A., Urios, V., Surroca, M., 2006. Vegetation height selection in Montagu's Harriers Circus pygargus breeding in a natural habitat. Ardea 94, 280-284.
|
|
Meyer, K.S., Spaar, R., Bruderer, B., 2003. Sea crossing behaviour of falcons and harriers at the southern Mediterranean coast of Spain. Avian Sci. 3, 153-162.
|
|
Newton, I., 2010. Bird Migration. Collins, London.
|
|
Pandolfi, M., Rossana, P., D'Astore, P., 1990. Analysis of breeding behaviour in Montagu's Harrier Circus pygargus in a site of CentraI Italy. Avocetta 14, 97-102.
|
|
Panuccio, M., Agostini, N., Mellone, U., 2005. Autumn migration strategies of honey buzzards, black kites, marsh and Montagu's harriers over land and over water in the central Mediterranean. Avocetta 29, 27-32.
|
|
Ravasini, M., 2022. Albanella Minore Cyrcus pygargus. In: Lardelli, R., Bogliani, G., Brichetti, P., Caprio, E., Celada, C., Conca, G., et al. (Eds.), Atlante Degli Uccelli Nidificanti in Italia. Edizioni Belvedere, Latina, pp. 286-287.
|
|
Rondinini, C., Battistoni, A., Teofili, C., 2022. Lista Rossa IUCN dei vertebrati italiani 2022. Comitato Italiano IUCN e Ministero dell'Ambiente e della Sicurezza Energetica, Roma, Italy.
|
|
Schlaich, A., 2019. Migrants in Double Jeopardy: Ecology of Montagu’s Harriers on Breeding and Wintering Grounds. University of Groningen, NL. Doctoral thesis.
|
|
Schlaich, A.E., Klaassen, R.H., 2021. Montagu’s harrier Circus pygargus. In: Panuccio, M., Mellone, U., Agostini, N. (Eds.), Migration Strategies of Birds of Prey in Western Palearctic. CRC Press, pp. 152–159.
|
|
Schlaich, A.E., Bretagnolle, V., Both, C., Koks, B.J., Klaassen, R.H., 2023. On the wintering ecology of Montagu's Harriers in West Africa: itinerancy in relation to varying annual environmental conditions. Ardea 111, 321-342.
|
|
Serratosa, J., Oppel, S., Rotics, S., Santangeli, A., Butchart, S.H., Cano-Alonso, L.S., et al., 2024. Tracking data highlight the importance of human-induced mortality for large migratory birds at a flyway scale. Biol. Conserv. 293, 110525.
|
|
Todd, L.D., Poulin, R.G., Brigham, R.M., Bayne, E.M., Wellicome, T.I., 2007. Pre-migratory movements by juvenile Burrowing Owls in a patchy landscape. Avian Conserv. Ecol. 2, 4.
|
|
Trierweiler, C., 2010. Travels to Feed and Food to Breed: the Annual Cycle of a Migratory Raptor, Montagu’s Harrier, in a Modern World. Doctoral Thesis. University of Groningen. University of Groningen, the Netherlands.
|
|
Warnock, N., 2010. Stopping vs. staging: the difference between a hop and a jump. J. Avian Biol. 41, 621-626.
|
|
Westrip, J.R.S., Burfield, I.J., Allen, D.J., Numa, C., 2022. The Conservation Status of Breeding Raptors in the Mediterranean. IUCN, Málaga, Spain.
|
|
Zuur, A.F., Ieno, E.N., 2016. A protocol for conducting and presenting results of regression-type analyses. Methods Ecol. Evol. 7, 636-645.
|
|
Züst, Z., Mukhin, A., Taylor, P.D., Schmaljohann, H., 2023. Pre-migratory flights in migrant songbirds: the ecological and evolutionary importance of understudied exploratory movements. Movement Ecol. 11, 78.
|
| Kai Gao, Xuemei Yang, Xi Huang, Wenhong Deng. 2024: Integrating host biological and ecological variables to predict probability of haemosporidian infection in raptors. Avian Research, 15(1): 100185. DOI: 10.1016/j.avrs.2024.100185 | |
| Jorge García-Macía, Ernesto Álvarez, Manuel Galán, Juan José Iglesias-Lebrija, Marc Gálvez, Gerard Plana, Núria Vallverdú, Vicente Urios. 2023: Home range variability and philopatry in Cinereous vultures (Aegypius monachus) breeding in Iberia. Avian Research, 14(1): 100134. DOI: 10.1016/j.avrs.2023.100134 | |
| Xueqin Deng, Qingshan Zhao, Junjian Zhang, Andrea Kölzsch, Diana Solovyeva, Inga Bysykatova-Harmey, Zhenggang Xu, Helmut Kruckenberg, Lei Cao, Anthony David Fox. 2021: Contrasting habitat use and conservation status of Chinese-wintering and other Eurasian Greater White-fronted Goose (Anser albifrons) populations. Avian Research, 12(1): 71. DOI: 10.1186/s40657-021-00306-0 | |
| Charlotte E. Hacker, Brandon D. Hoenig, Liji Wu, Wei Cong, Wei Cong, Jingjing Yu, Yunchuan Dai, Ye Li, Jia Li, Yadong Xue, Yu Zhang, Yunrui Ji, Hanning Cao, Diqiang Li, Yuguang Zhang, Jan E. Janecka. 2021: Use of DNA metabarcoding of bird pellets in understanding raptor diet on the Qinghai-Tibetan Plateau of China. Avian Research, 12(1): 42. DOI: 10.1186/s40657-021-00276-3 | |
| Adrian Orihuela-Torres, Juan M. Pérez-García, Zebensui Morales-Reyes, Lara Naves-Alegre, José A. Sánchez-Zapata, Esther Sebastián-González. 2021: Avian-power line interactions in the Gobi Desert of Mongolia: are mitigation actions effective?. Avian Research, 12(1): 41. DOI: 10.1186/s40657-021-00277-2 | |
| Nadejda Djorgova, Dimitar Ragyov, Valko Biserkov, Jordan Biserkov, Boris P. Nikolov. 2021: Habitat preferences of diurnal raptors in relation to human access to their breeding territories in the Balkan Mountain Range, Bulgaria. Avian Research, 12(1): 29. DOI: 10.1186/s40657-021-00265-6 | |
| Roberto Sánchez, Javier Sánchez, Javier Oria, Francisco Guil. 2020: Do supplemental perches influence electrocution risk for diurnal raptors?. Avian Research, 11(1): 20. DOI: 10.1186/s40657-020-00206-9 | |
| Satish Pande, Reuven Yosef, Federico Morelli, Rajkumar Pawar, Ram Mone. 2018: Diet and habitat affinities in six raptor species in India. Avian Research, 9(1): 36. DOI: 10.1186/s40657-018-0129-2 | |
| Zhiping Huo, Junfeng Guo, Xia Li, Xiaoping Yu. 2014: Post-fledging dispersal and habitat use of a reintroduced population of the Crested Ibis (Nipponia nippon). Avian Research, 5(1): 7. DOI: 10.1186/s40657-014-0007-5 | |
| Cheng WEN, Dong HAN. 2013: Raptor migration monitoring in the spring of 2009 at Baiwangshan, Beijing. Avian Research, 4(4): 319-327. DOI: 10.5122/cbirds.2013.0031 |
Figures(6) / Tables(3)





 DownLoad:
DownLoad:

 Email Alerts
Email Alerts RSS Feeds
RSS Feeds