Ministry of Education Key Laboratory for Ecology of Tropical Islands, Key Laboratory of Tropical Animal and Plant Ecology of Hainan Province, College of Life Sciences, Hainan Normal University, Haikou, 571158, China
Funds:
the National Natural Science Foundation of China31970427
the National Natural Science Foundation of China32270526
Nests are important structures for birds to raise their offspring and for signaling. Many birds incorporate feathers into the nest, since feathers were traditionally thought to serve the function of insulation. Hypotheses in recent years have considered that some birds place feathers in conspicuous locations in the nest for decoration to trigger a fear response in the competitors. In this study, we investigated whether decorative feathers could deter nest usurpation by Crested Mynas (Acridotheres cristatellus) by manipulating nest box contents. The results revealed that Crested Mynas preferred black feathers to white feathers as decorations and occupied nest boxes decorated with black feathers significantly less than those decorated with white feathers, suggesting that black decorative feathers in the nest could be more effective in preventing nest usurpation by Crested Mynas and that white decorative feathers may have other functions. The black feathers in the nest are prominently placed at the edge of the nest to convey the message that "this nest is occupied" or "the owner of this nest has been preyed upon" to visitor Crested Mynas, thus effectively preventing them from usurping the nest at a later stage.
Among several materials used by birds to create their nests, the use of feathers has received particular attention in evolutionary ecology studies (Collias and Collias, 1984; Hansell, 2000; Deeming and Reynolds, 2015). Feathers were traditionally thought to have a thermoregulatory function because many finches deposit small, soft feathers in the nest cup, in direct contact with the eggs (Møller, 1984; Lombardo et al., 1995; Pinowski et al., 2006; Dawson et al., 2011; Windsor et al., 2013). Some nest lining feathers can be used to cultivate antimicrobial producing bacteria, which protect birds from pathogenic infection (Peralta-Sánchez et al., 2010, 2011, 2014; Ruiz-Castellano et al., 2019), with white feathers being better culture media than pigmented feathers (Peralta-Sánchez et al., 2010, 2014). However, some species place flight or contour feathers at the edge of the nest, which are not intended to provide insulation but may serve a decorative purpose (García-Navas et al., 2015; Rubalcaba et al., 2017; Järvinen and Brommer, 2020a, 2020b). Feathers are a limited resource for wild birds, especially woodland nesters because they usually come from dead or killed birds (Hansell, 1995). Therefore, feather aggregations in a nest may indicate that nesters are in good health or are highly inclined to devote their time and energy to breeding, thus engaging in nest decorative behavior involving the use of feathers for sexual signaling (Polo and Veiga, 2006; Sanz and García-Navas, 2011; Mainwaring et al., 2016). There are other functions of feathers in nest decoration, where Slagsvold and Wiebe (2021b) proposed that birds trigger fear responses in competitors by decorating nests with feathers, thereby reducing the risk of nest usurpation. The authors suggest that naive prospecting birds may perceive the nest feathers as the result of a predation event and that nest owners decorate their nests with bright white feathers visible from the cavity to deter other species from entering. However, the recently proposed hypothesis that feathers prevent nest usurpation still requires verification across more bird species.
As far as we known, the behavior of birds using feathers for nest decoration has received more attention. However, few studies have considered the differences in the color of feathers within the nest (but see Slagsvold and Wiebe, 2021b), despite the evidence that birds differ in their preferences for specific colors of feathers (Ruiz-Castellano et al., 2018). Therefore, we hypothesize that different colored feathers in the nest may play different functions that need further exploration.
The Crested Myna (Acridotheres cristatellus) is a secondary cavity-nesting bird (Ding et al., 2019) that can breed in nest boxes (Liu and Liang, 2021a). Previous studies have found that Crested Mynas use snake sloughs to decorate nests (Liu et al., 2021), and sloughs effectively reduces the risk of nest predation (Liu and Liang, 2021b). Crested Mynas also decorate their nests with feathers (usually black or white; Fig. 1), which are prominently displayed on the edge of the nest. However, the function of decorative feathers in Crested Myna nests has not been studied. Therefore, we conducted an experimental study from March to June 2022 using a nest manipulation design. First, we estimated the probability of Crested Mynas' nests containing white or black feathers during the nesting and incubating periods. Secondly, we explored the effect of different nest feather colors (black and white) on nest usurpation by Crested Mynas. We predicted a significant difference in the proportion of Crested Myna nests that included black or white feathers as lining material. However, since white and black feathers may have different functions, we explored Crested Myna's preference for nests containing black or white feathers.
Figure
1.
Crested Mynas added black feathers to the nest during nesting and egg incubation to decorate the nest.
This study was conducted in 2022 in Dingcheng Town, Ding'an (19°37′ N, 110°19′ E), Hainan, south China. This area has a tropical monsoon maritime climate, with a mild weather and abundant heat and rainfall, and an average annual temperature of 24.2 ℃ (Liu et al., 2023). Natural tree cavities are a scarce resource, as old or decaying trees are often cut down by local farmers in a timely manner; thus, the availability of artificial nest boxes can effectively mitigate the limitation of nest site availability for local cavity-nesting birds (Liu et al., 2023). Crested Mynas are the main species using nest boxes to breed in the study area, along with a small number of Common Mynas (Acridotheres tristis), Japanese Tits (Parus minor) and Oriental Magpie-robins (Copsychus saularis). During the breeding season, Mynas decorate their nests with feathers of different colors (mostly black and white), and nests of the Japanese Tits and Oriental Magpie-bobins are often usurped by the dominant Crested Mynas.
The Crested Myna is widely distributed in China south of the Qinling-Huaihe River as well as in India and Vietnam (Craig and Feare, 1998; Ding et al., 2019; Zheng, 2023). It has been introduced to other parts of China (Liu and Chen, 2021) as well as to the Philippines, Canada, and other countries due to cage trade (Ding et al., 2019). The Crested Myna is usually black, with white spots on the wings at the base of the flight feathers and white horizontal spots on the undertail coverts (Ding et al., 2019; Liu and Chen, 2021). The body length is 23–28 cm and the body weight is 78–150 g (Ding et al., 2019). The subspecies A. c. brevipennis is a resident of Hainan Island (Zheng, 2023), with the breeding period lasting from mid-March to mid-August, where the birds fly frequently over the nest sites (Ding et al., 2019; Liu and Liang, 2021a). Breeding occurs at two peak laying times, in late April and early June (Liu and Liang, 2021a). The male and female parents construct the nest together, adding feathers, snake sloughs, green leaves, and plasticine to the interior part of the nest (Liu and Liang, 2021a).
2.2
Nest manipulation design
In this study, field data were collected during the first peak breeding season (March–June) of Crested Mynas. In late February or early March, vertical nest boxes (the inner diameter of the nest box: length 15 cm, width 15 cm, depth 30 cm, the diameter of the entrance: 6 cm; also see Liu and Liang, 2021a; n = 140) were fixed using a wire to a pole that was 3 m above the ground. Nest boxes with an entrance orientated between 180° and 360° (0° represents the north) to reduce the damage caused by rainfall and typhoons (Liu and Liang, 2021a). The distance between adjacent nest boxes within the group was 30–50 m to ensure that the habitats of nest boxes within the group were similar. In order to explore whether Crested Mynas have a preference for decorative feather color (black or white), we observed the nest photographs of Crested Mynas taken in 2022 during the early nesting and incubating periods (Fig. 1), and we analyzed the rate of Crested Myna nests that included black or white feathers as lining material. In order to test whether the decorative feathers inside the nest had a protective function against nest usurpation, we conducted 20 groups of nest box manipulation experiments, the contents of the nest boxes were treated sequentially: exp.1–exp.4 (Fig. 2). For exp.1 (white feathers treatment; n = 20), the nest box was lined with a flat layer of moss, with three long white feathers on top of the moss. For exp.2 (white paper treatment; n = 20), the nest box was lined with a flat layer of moss, with three long white paper strips placed on top of the moss. For exp.3 (black feathers treatment; n = 20), the nest box was lined with a flat layer of moss, with three long black feathers placed on top of the moss. For exp.4 (control treatment, n = 20), the nest box was lined with a flat layer of moss only. The feather length added to the nest box was 10–15 cm, purchased online (Taobao Inc., Hangzhou, China). The nest boxes were inspected at least once a week, and changes in the nest box contents were recorded.
Figure
2.
Nest box manipulation experiments. For exp.1, three long white feathers placed on top of the moss; for exp.2, three long white paper strips placed on top of the moss; for exp.3, three long black feathers placed on top of the moss; for exp.4, only moss.
When at least one Crested Myna egg was present in the nest box, the Crested Myna was considered to have occupied the nest box for breeding (Liu and Liang, 2021a). Chi-square tests were used to analyze the rate of Crested Myna nests that included black or white feathers as lining material. Chi-square tests were used to analyze the differences in the occupancy rates of nest boxes among exp.1–exp.4. All tests were two-tailed, with a significance level of P < 0.05. Data were presented in the form of mean ± standard deviation (Mean ± SD). IBM SPSS 22.0 software (IBM Corp., Armonk, NY, USA) was used for all data analyses.
3.
Results
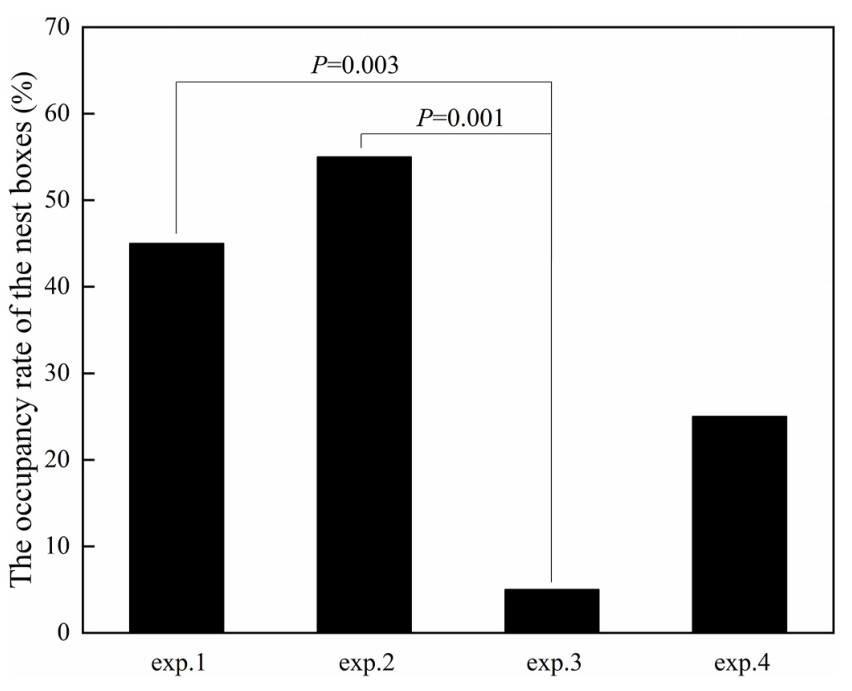
When reviewing the pictures of nests during nesting and egg incubation, long black feathers were prominently placed at the edge of the Crested Myna nest (Fig. 1). The percentage of Crested Myna nests that included black feathers (93.9%, n = 115) was significantly higher than the percentage of white feathers added to the nest (63.5%, n = 115) (Chi-square tests, χ2 = 31.768, df = 1, P < 0.001). There was a significant difference in occupancy rates of nest boxes among exp.1–exp.4 (Chi-square tests, χ2 = 13.447, df = 3, P = 0.004; Table 1). The occupancy rate was lower for exp.3 (5%) than for exp.1 (45%; Chi-square tests, χ2 = 8.533, df = 1, P = 0.003), exp.2 (55%; Chi-square tests, χ2 = 11.905, df = 1, P = 0.001) and exp.4 (25%; Chi-square tests, χ2 = 3.137, df = 1, P = 0.077). And the occupancy rate was lower for exp.4 than for exp.1 (Chi-square tests, χ2 = 1.758, df = 1, P = 0.158) and exp.2 (Chi-square tests, χ2 = 3.750, df = 1, P = 0.053) (Fig. 3).
Table
1.
Chi-square tests to analyze the differences in occupancy rates of Crested Mynas of nest boxes with different contents.
We found that adding black feathers to the nest was a common behavior, and that long black feathers in the nest are prominently placed at the edge of the nest by the Crested Mynas, which could convey the message that "this nest is occupied" or "the owner of this nest has been preyed upon" to visitor Crested Mynas. The occupancy rate of nest boxes in black feathers treatment was significantly lower than that of nest boxes in white feathers and white paper treatments, indicating that black long feathers could prevent nest usurpation by Crested Mynas more effectively than white long feathers, and the white decorative feathers may have other functions.
Collecting long feathers as decorative nesting materials for birds may be costly (Mainwaring et al., 2016), as they usually come from dead or killed birds (Hansell, 1995); therefore, the aggregation of nest decorations signal that nest owners are potentially high quality and have high levels of resources (Veiga and Polo, 2005; Sanz and García-Navas, 2011; Sergio et al., 2011). It has been suggested that Eurasian Blue Tits (Cyanistes caeruleus) add feathers to their nests to signal high quality to other blue tits (an extended phenotype, Sanz and García-Navas, 2011; Järvinen and Brommer, 2020b; Weduwen et al., 2021). We suggest that the addition of feathers to nests of Crested Mynas also serves as a signal of high quality to prospectors, alerting them that "this nest has a high-quality owner". The feathers used by birds for nest decoration are usually bright colored and vivid (Veiga and Polo, 2005; Sanz and García-Navas, 2011; Ruiz-Castellano et al., 2018; Slagsvold and Wiebe, 2021b), so that they are easily visible to prospecting birds from the cavity entrance. However, we found that Crested Mynas prefer to decorate their nests with long black feathers, and they placed long black feathers inside the nest at the edge of the nest. The placement of feathers in the nest was similar to that of Rock Sparrows (Petronia petronia), resulting in maximal visibility (García-Navas et al., 2015). We suggested long black feathers may be more representative of the body quality of the Crested Myna than white feathers, possibly because black long feathers are more difficult to collect than white long feathers in the study area. Our study area was located in an agroecosystem, and villagers in the study area raise free-ranging white domestic geese or ducks. White feathers of domestic geese or ducks were seen in their activity areas from time to time, while black long feathers animals were almost absent. As long black feathers were rarely seen in the study area, the collection of long black feathers increased flight costs to the birds; therefore, the collection of more black long feathers in the nest may indicate better body quality.
Farmers in southwest China often insert poultry feathers in conspicuous places to prevent birds from entering their fields to steal crops (Fig. 4). In nature, some birds use feathers as decorative materials and insert them in conspicuous places on their nests. So, can the decorative feathers in the nest prevent other prospecting birds from entering the nest? Slagsvold and Wiebe (2021b) found that Pied Flycatchers (Ficedula hypoleuca), European Blue Tits (Cyanistes caeruleus) and Tree Swallows (Tachycineta bicolor), all showed hesitation to entering nest boxes with white feathers and boxes with black feathers. The authors suggested that feathers in the nest can prevent other prospecting birds from entering the nest. The hesitancy of the three bird species can be attributed to the fact that naive prospecting birds perceive feathers in nests as being the product of predation events, so that they fear entering the nest box to explore. Similarly, our results found that adding long black feathers to the nest effectively reduced invasion by Crested Mynas, as long black feathers could convey the message that "this nest is occupied" or "the owner of this nest has been preyed upon" to visitor Crested Mynas.
Figure
4.
Farmers use feathers to prevent birds from entering their fields (Photo source: Nanguo Morning Post client).
Slagsvold and Wiebe (2021b) found that the hesitation time for the nest boxes with black feathers was significantly shorter than that for the nest boxes with white feathers, and the authors explained that nest owners decorate their nests with bright white feathers, which made it easier to see the "predation" event from the cavity, to effectively deter small prospecting birds from entering. However, in the present study we found the occupancy rate of Crested Mynas on white feathers treatment was significantly higher than black feathers treatment, which may be due to: 1) the nest prospectors think that "this nest has been occupied by other individuals". Long black feathers added to the tit nests, probably to confuse nest prospectors because Crested Mynas are black in color and can blend in with the collected long black feathers. Coupled with the low light intensity inside the cavity, nest prospectors cannot easily detect whether the nest owner is inside the nest, so they do not dare to easily enter the nest. 2) the nest prospectors think that "this nest owner has been preyed upon". The black feathers on the surface of the nest material can deceive the nest prospectors into thinking that "this nest owner has been preyed upon", and they are unable to confirm whether the predator is in the nest, so they do not dare to easily enter the nest. Therefore, Crested Mynas decorate their nests with long black feathers, probably to trigger a fear response in their competitors to reduce the risk of nest usurpation.
Slagsvold and Wiebe (2021b) found that white feathers can effectively prevent nest usurpation; however, our study found that Crested Mynas occupied white feathers treatment more than only moss treatment. In other words, the white feathers in the nests did not prevent the nest from being occupied by Crested Mynas, which might be related to the brightness of the bottom of the nest box. Previous studies have shown that light intensity decreases with increasing depth of the cavity, where the visibility of the bottom of the nest decreases and color vision of birds becomes impaired (Wesołowski and Maziarz, 2012). Slagsvold and Wiebe (2021a) also illustrated that the visibility in deep nest boxes (24 cm) was lower than that in shallow boxes, and Pied Flycatchers were more hesitant to enter the cavity. The depth of the nest boxes we used was 30 cm, making the visibility of the bottom of the nest decrease; thus, prospecting competitors would be afraid to enter the bottom of the nest box. However, the white color of the bottom illuminates the bottom of the nest cavity, making it easy for prospectors to identify whether the nest owner or predator was present from the entrance to the hole. In addition, white feathers in the nest have other advantages, such as protecting birds from pathogenic infection (Peralta-Sánchez et al., 2010, 2011, 2014; Ruiz-Castellano et al., 2019), with white feathers being better culture media than pigmented feathers (Peralta-Sánchez et al., 2010, 2014). Therefore, nests containing white feathers may be more attractive, and mynas are more likely to occupy the white feathers treatment. Future studies should investigate whether the deterrent effect of feathers as nest decorations is also effective against nest parasitoids and nest predators.
5.
Conclusion
To sum up, we found that nest decoration by Crested Mynas using feathers was a common behavior, with black feathers being preferred to white feathers. Slagsvold and Wiebe (2021b) suggested that birds use fear of feathers (black and white color) to protect the nest from usurpation. However, we found that long black feathers used as nest decorations were effective in reducing nest usurpation by Crested Mynas. Notably, white feathers in the nest did not reduce nest usurpation by Crested Mynas, which may be related to the brightness of the base of the nest. However, few studies have explored the effect of different feather colors on nest encroachment prevention (but see Slagsvold and Wiebe, 2021b). Although it has been shown that birds differ in their preference for different colored feathers (Ruiz-Castellano et al., 2018), we suggest that the fear of feathers hypothesis should be tested in other species, with a focus on the differences in feather color.
Data availability
The datasets used in this study are provided as supplementary material (Appendix Table S1).
Ethics statement
The experiments comply with the current laws of China, where they were performed. Experimental procedures were in agreement with the Animal Research Ethics Committee of Hainan Provincial Education Centre for Ecology and Environment, Hainan Normal University (No. HNECEE-2014-005). We minimized disturbance to nesting mynas by performing nest boxes checks quickly (typically at the nest for <3 min per check).
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We are grateful to Fangfang Zhang, Xuan Zhang, Hanlin Yan, Yuhan Zhang and Cheng Chen for their help with the experiments and data collection.
Borgia, G., Gore, M.A., 1986. Feather stealing in the satin bowerbird (Ptilonorhynchus violaceus): male competition and the quality of display. Anim. Behav. 34, 727-738.
Collias, N.E., Collias, E.C., 1984. Nest Building and Bird Behavior. Princeton University Press, Princeton.
Craig, A., Feare, C., 1998. Starlings and Mynas. Black Publishers Ltd., London.
Dawson, R.D., O'Brien, E.L., Mlynowski, T.J., 2011. The price of insulation: costs and benefits of feather delivery to nests for male tree swallows Tachycineta bicolor. J. Avian Biol. 42, 93-102.
Deeming, D.C., Reynolds, S.J., 2015. Nests, Eggs, and Incubation: New Ideas about Avian Reproduction. Oxford University Press, New York.
Ding, P., Zhang, Z., Liang, W., Li, X., 2019. The Forest Birds of China. Hunan Science & Technology Press, Changsha.
Endler, J.A., Endler, L.C., Doerr, N.R., 2010. Great bowerbirds create theaters with forced perspective when seen by their audience. Curr. Biol. 20, 1679-1684.
García-Navas, V., Valera, F., Griggio, M., 2015. Nest decorations: an 'extended' female badge of status? Anim. Behav. 99, 95-107.
Hansell, M.H., 1995. The demand for feathers as building material by woodland nesting birds. Hous. Theor. Soc. 42, 240-245.
Hansell, M.H., 2000. Bird Nests and Construction Behavior. Cambridge University Press, Cambridge.
Järvinen, P., Brommer, J.E., 2020a. Nest ornaments and feather composition form an extended phenotype syndrome in a wild bird. Behav. Ecol. Sociobiol. 74, 1-8.
Järvinen, P., Brommer, J.E., 2020b. Lining the nest with more feathers increases offspring recruitment probability: selection on an extended phenotype in the blue tit. Ecol. Evol. 10, 13327-13333.
Liu, J., Liang, W., 2021a. The breeding ecology of the crested myna Acridotheres cristatellus on tropical hainan Island. Ornithol. Sci. 20, 83-92.
Liu, J., Liang, W., 2021b. Snake slough in birds' nests acts as a nest predator deterrent. Ethol. Ecol. Evol. 33, 591.
Liu, J., Liu, J., Liang, W., 2021. Snake slough in nests of Crested Mynas: effect on breeding success and nestling growth. Eur. Zool. J. 88, 616-621.
Liu, J., Zhang, F., Liu, Y., Liang, W., 2023. Egg recognition and nestling discrimination in the Crested Myna (Acridotheres cristatellus): size matters. Avian Res. 14, 100111.
Liu, Y., Chen, S., 2021. The CNG Field Guide to the Birds of China. Hunan Science & Technology Press, Changsha.
Lombardo, M.P., Bosman, R.M., Faro, C.A., Houtteman, S.G., Kluisza, T.S., 1995. Effect of feathers as nest insulation on incubation behavior and reproductive performance of tree swallows (Tachycineta bicolor). Auk 112, 973-981.
Mainwaring, M.C., Hartley, I.R., Lambrechts, M.M., Deeming, D.C., 2014. The design and function of birds' nests. Ecol. Evol. 4, 3909-3928.
Mainwaring, M.C., Hartley, I.R., 2013. The energetic costs of nest building in birds. Avian Biol. Res. 6, 12-17.
Mainwaring, M.C., Wolfenden, A., Read, J.E., Robson, J.M.A., Tomlinson, C.J., Hartley, I. R., 2016. Feathering the nest: the effects of feather supplementation to blue tit. Avian Biol. Res. 9, 89-95.
Møller, A.P., 1984. On the use of feathers in birds' nests: predictions and tests. Ornis Scand. 15, 38-42.
Moreno, J., 2012. Avian nests and nest-building as signals. Avian Biol. Res. 5, 238-251.
Penteriani, V., Delgado, M.M., 2008. Owls may use faeces and prey feathers to signal current reproduction. PLoS One 3, e3014.
Peralta-Sánchez, J.M., Møller, A.P., Soler, J.J., 2011. Colour composition of nest lining feathers affects hatching success of barn swallows, Hirundo rustica (Passeriformes: hirundinidae). Biol. J. Linn. Soc. 102, 67-74.
Peralta-Sánchez, J.M., Møller, A.P., Martín-Platero, A.M., Soler, J.J., 2010. Number and colour composition of nest lining feathers predict eggshell bacterial community in barn swallow nests: an experimental study. Funct. Ecol. 24, 426-433.
Peralta-Sánchez, J.M., Soler, J.J., Martín-Platero, A.M., Knight, R., Martínez-Bueno, M., Møller, A.P., 2014. Eggshell bacterial load is related to antimicrobial properties of feathers lining barn swallow nests. Microb. Ecol. 67, 480-487.
Pinowski, J., Haman, A., Jerzak, L., Pinowska, B., Barkowska, M., Grodzki, A., Haman, K., 2006. The thermal properties of some nests of the Eurasian tree sparrow Passer montanus. J. Therm. Biol. 31, 573-581.
Polo, V., Veiga, J.P., 2006. Nest ornamentation by female spotless starlings in response to a male display: an experimental study. J. Anim. Ecol. 75, 942-947.
Rubalcaba, J.G., Fuentes, D., Veiga, J.P., Polo, V., 2017. Nest decoration as social signals by males and females: greenery and feathers in starling colonies. Behav. Ecol. 28, 1369-1375.
Ruiz-Castellano, C., Ruiz-Rodríguez, M., Tomás, G., Soler, J.J., 2019. Antimicrobial activity of nest-lining feathers is enhanced by breeding activity in avian nests. FEMS Microbiol. Ecol. 95, fiz05.
Ruiz-Castellano, C., Tomás, G., Ruiz-Rodríguez, M., Soler, J.J., 2018. Nest material preferences by spotless starlings. Behav. Ecol. 29, 137-144.
Sanz, J.J., García-Navas, V., 2011. Nest ornamentation in blue tits: is feather carrying ability a male status signal? Behav. Ecol. 22: 240-247.
Sergio, F., Blas, J., Blanco, G., Tanferna, A., López, L., Lemus, J.A., Hiraldo, F., 2011. Raptor nest decorations are a reliable threat against conspecifics. Science 331: 327-330.
Slagsvold, T., Wiebe, K.L., 2021a. Egg covering in cavity nesting birds may prevent nest usurpation by other species. Behav. Ecol. Sociobiol. 75, 116.
Slagsvold, T., Wiebe, K.L., 2021b. Nest decoration: birds exploit a fear of feathers to guard their nest from usurpation. R. Soc. Open Sci. 8, 211579.
Soler, J.J., Møller, A.P., Soler, M., 1998. Nest building, sexual selection and parental investment. Evol. Ecol. 12, 427-441.
Veiga, J.P., Polo, V., 2005. Feathers at nests are potential female signals in the spotless starling. Biol. Lett. 1, 334-337.
Weduwen, Dd, Keogan, K., Samplonius, J.M., Phillimore, A.B., Shutt, J.D., 2021. The correlates of intraspecific variation in nest height and nest building duration in the Eurasian blue tit Cyanistes caeruleus. J. Avian Biol. 52, e02528.
Wesołowski, T., Maziarz, M., 2012. Dark tree cavities-a challenge for hole-nesting birds? J. Avian Biol. 43, 454-460.
Windsor, R.L., Fegely, J.L., Ardia, D.R., 2013. The effects of nest size and insulation on thermal properties of tree swallow nests. J. Avian Biol. 44, 305-310.
Zheng, G., 2023. A Checklist on the Classification and Distribution of the Birds of China, fourth ed. China Science Publishing & Media Ltd., Beijing.


 DownLoad:
DownLoad:







 Email Alerts
Email Alerts RSS Feeds
RSS Feeds