Animals must strike a balance between anti-predation behavior and other essential behaviors, such as foraging. Within the same species, strategies may vary on individuals' risk-taking preferences, and in this process the environment is a determinant, in addition to predator regime. The Crested Ibis (Nipponia nippon) exhibits such tendency. This is an endangered species, once inhabiting exclusively in China's Qinling Mountain. This used to be the sole remaining wild population. However, over recent decades, this population has expanded. A portion has relocated to breed in the lower plain area, which is characterized by elevated level of human activities and landscape complexity. We used flight initiation distance (FID) as an indicator of the ibises' risk-taking preference, particularly their response to human proximity. Additionally, we examined the environmental factors influencing their foraging site selection, including altitude, terrain openness, human activity intensity and human construction. Our findings revealed a significantly shorter FID among individuals relocating to plain habitats, indicating a higher tolerance of human proximity. The results showed that FID decreased with distance to the nearest human settlement. Another finding is that FID was independent of instant human activity intensity and environmental factors (altitude and terrain openness). These different may arise from various combinations of human activity, predation risk, and food abundance within the two habitats. These results provide insights into the insitu conservation of the threatened species within the context of global urbanization.
Prey species constantly need to weigh the benefits of a specific habitat against predation risk there. The necessity is even higher when they move to a novel environment. Studies have revealed that birds adopt different strategies when adapting to different habitats (Yeh et al., 2007; Valcarcel and Fernández-Juricic, 2009; Luther and Baptista, 2010). On top of this, environmental changes caused by human activities mean that birds constantly need to adapt to new conditions (Shochat et al., 2006; Sih et al., 2011). One of such changes is the ever-expanding urban areas. While some species struggle to cope with human encounters and face extinction, others are adapting well to the updated proximity and even successfully invading urban habitats (Tomialojc, 1982; Stork, 2010; Carrete and Tella, 2011; Gil and Brumm, 2014). Møller (2013), based on published data, examined 13 categories of life history and ecological factors, including habitat generalism, body size, and diet, which might impact animals’ adaptation to the new urban environments, and found that the response to predators was the most predominant among the 13 considered categories.
Wild animals are subject to human activities in general, to varying degrees (Frid and Dill, 2002; Møller et al., 2007). While hunting is lethal, others, like eco-tourism and agriculture activities, may not kill, but may still lead to decline in wild animal population (e.g. Donald et al., 2001). Possible explanations include: a) physiological pressure from perceived danger (e.g., human approach; Cresswell, 2008); b) reduced foraging efficiency when being interrupted; and c) increased energy expenditure during sudden flight from humans (Tatner and Bryant, 1986). Such effects can negatively affect both the physical condition and the reproduction of wild species (Tatner and Bryant, 1986; Zanette et al., 2011; Tarjuelo et al., 2015). Therefore, human activities might be threat to wild animals which invade man-made areas (Frid and Dill, 2002; Tablado and Jenni, 2015).
The Crested Ibis (Nipponia nippon) is a medium-size wading bird that has been classified as an “Endangered” species by IUCN (2023). In the mid-20th century, their population sharply declined, to the point where they were not observed at all in Russia, China and the Korean Peninsula, which, in addition to Japan, were their only distribution regions from 1960s to 1970s (Ding, 2004). When the last five individuals in Japan were placed in captivity, this species was evaluated as extinct in wild. However, a small relict population was rediscovered in the mountain area of Yangxian, Shaanxi Province in Central China in May 1981. Since its rediscovery, population soared hundreds of times from 7 to over 4500 individuals in 2020, thanks to significant conservation efforts at both national and provincial levels (Liu, 1981; Xu et al., 2023). The increase, interestingly, was accompanied by distribution variation. Mountain areas with relatively low human density were once the primary breeding habitat for the bird. Yet since 1997, some individuals have migrated to lower plain areas and started breeding there (Ding, 2004). This migrant numbers have been continuously growing, reaching approximately two-thirds of the wild population (66.67%, nest = 224; Xu et al., unpublished data) breeding in plain areas by 2021. Plain areas consequently have become crucial foraging and breeding grounds for the Crested Ibis. Nevertheless, here humans have more presence compared with in mountain habitats (Li et al., 2002; Hu et al., 2016). Moreover, as local economy expands, the impact of human activities on avian species also grows (Li et al., 2002; Ding, 2010; Ye et al., 2022). Birds’ responses to human presence could significantly shape their adaption to the choice of habitats (Huang et al., 2022). Understanding their behavioral changes becomes crucial in addressing the challenges faced by the ibis and similar species in human-altered landscapes.
Fight initiation distance (FID) is the distance at which an animal initiates flight when approached by a potential predator. FID has been previously used as a metric of anti-predation behavior in ibises (Blumstein, 2003; Bateman and Fleming, 2014). Generally, the FID of avian species in habitats with higher human population densities, such as cities, was shorter than that of their conspecifics in low human-density areas (Bautista et al., 2004; Webb and Blumstein, 2005; Weston et al., 2012). The variation in tolerance was often considered to be because of behavioral habituation (Gil and Brumm, 2014). Local adaptation to urban habitats could also explain this difference, indicating that certain genotypes have increased in frequency or become favored among individuals in urban environments (Evans et al., 2010; van Dongen et al., 2015). This phenomenon is expected to lead to genetic differentiation among populations (Carrete et al., 2016). Differential selection among behavioral syndromes could explain the variation in recruitment to urban habitats, resulting in the sorting of phenotypic under different levels of interference pressures between urban and rural environments (Schell, 2018). So far, the reasons for this difference remain controversial and challenging to examine due to the necessity for long-term monitoring of population dynamics, often involving the marking of individuals.
The maximum distance between the original (mountain) and new (plain) habitats was 19 km. On top of that, the general winter-feeding habitats of the breeding ibis can be 25 km away from each other (15–45 km; Ding, 2004, 2010). Such wide ranges of ~20 km demonstrated ibis's high mobility. Thanks to such distance, there could be meaningful behavioral contrasts between populations in different habitats, which is good for investigation. Moreover, behaviors of individuals that venture into plain habitats will yield insight into suitable reintroduction sites and understanding the future dynamics of the wild population. Based on this information, we can make more informed decisions and implement targeted conservation strategies to safeguard the endangered species and its habitats.
Our main study objectives were to compare anti-predation behavior of Crested Ibis in their original habitats (mountain) with that in newly colonized habitats (plain), as well as to identify factors behind selecting specific foraging sites, which included altitude, terrain openness, and human activity. To realize these goals, the study covered an extensive area of 1662 km2, encompassing the primary distribution range. Although ibis has remarkable ability to move between different habitats, only part of the population expands to the plain area while others remain in mountainous habitats. Therefore, we inferred that the tolerance to human activity might influence the ibis’ ability to breed in the plain area. Consequently, we hypothesized that the ibises expanding to the plain area have a higher tolerance to human. Accordingly, we predicted that these ibises would display a shorter FID and forage in closer proximity to humans. By exploring these behavioral aspects, we aimed to better understand the adaptability of Crested Ibis in human-influenced environments and to make more effective conservation strategies.
2.
Materials and methods
2.1
Study area
This study was carried out in Yangxian County, located in southern Shaanxi, Central China (33°05′ N–33°45′ N, 107°25′ E−107°82′ E). The Shaanxi Hanzhong Crested Ibis National Nature Reserve, also the primary distribution area of wild Crested Ibis population, locates in this region. Stretching into eastern part of Hanzhong Basin, Yangxian has a warm and temperate climate zone with humid monsoons. Around this region there are both mountainous and plain terrain. In particular, the middle and lower part of the Yangxian mountains is the original distribution areas of Crested Ibis, including forests, small farmlands (mainly paddy fields) and villages. Since 1997, the Crested Ibis has expanded its habitat to include the lower plains of Yangxian. Compared with mountain environment, plain is characterized by fewer tall vegetations, more human-made constructions, and extensive tracts of uninterrupted farmland (also mainly paddy fields) (Song et al., 2018). Now ibises breed and forage in both habitats (Huang et al., 2022).
2.2
Data collection
The study was conducted in both the original mountain and plain areas during both the breeding (May to June) and non-breeding (September to October) season of 2015. All nestlings of the Crested Ibis have been banded with a unique combination of color bands (1987–2000) or numbered color bands (2000–2022). Thus, individuals can be identified using monocular or binoculars observation. To minimize bias, all approaches were conducted by the same researcher (Y.Z). The same line transects, also employed in the Crested Ibis population census, were used to avoid testing the same individuals repeatedly. If the birds were foraging in a flock, the nearest individual in the direction of approach was selected and recorded for its FID. Starting distance, the distance between an observer and an individual at the moment of approach, was also recorded.
Variables related to human activity intensity and the characteristics of the site were recorded. These included long-term human activity intensity (distance to the nearest road and distance to the nearest settlement), and instant intensity (number of people within a 50-m radius). To exclude the influence of landscape characteristics on ibis’ flight behavior, we only collected data on terrain openness during non-breeding season. Terrain openness consists of two factors: valley width, which is the distance between the bottom of two hills horizontally, and terrain openness, which is elevation angle of two mountain ridges from the site (if applicable). Given the close proximity between foraging sites and nocturnal roosts, as documented by Ding (2004) (2.82 ± 1.49 km in the non-breeding season and 0.56 ± 0.68 km in the breeding season), it is inferred that the features observed at foraging sites are also characteristic of the home range population.
2.3
Statistical analyses
All analyses were performed in R version 4.1.2 (R Core Team, 2021). The repeatability of individual FID was evaluated using the library ‘rptR’ (Nakagawa and Schielzeth, 2010). The difference in ibis flock sizes and the long-term and instant human activity intensities between two habitats (mountain area and plain area) were processed using logistic regression. The differences of altitude and the distance to the nearest road between original and plain habitats were analyzed by t-test, as these two factors both had significant correlation with the distance to the closest settlement (distance to the nearest road: r = 0.253, n = 176, p < 0.001; altitude: r = −0.167, n = 176, p = 0.026) and flock size (distance to the nearest road: r = 0.392, n = 160, p < 0.001; altitude: r = −0.157, n = 160, p = 0.048).
A generalized linear mixed model (GLMM) implicated in the library ‘lme4’ (Bates et al., 2013) was used to explore the influence on FID (the explanatory variables: habitat, flock size, altitude, long-term and instant human activity intensity, and their interaction terms with habitat; and the random factor: season i.e., breeding and non-breeding season). The terrain openness was showed as mean ± SE, as this data was only available in mountain areas. The effect of terrain openness (terrain openness and valley width) on FID was studied using a linear model with season as random effects.
Considering the ecological significance of SD and to avoid the statistics biases, the buffer distance (SD minus FID) was also used respectively as a covariate in models, to test whether buffer distance influences FID.
3.
Results
In total, 192 adult Crested Ibises were studied, and their FIDs were recorded. The average FID was 25.55 ± 8.79 m (mean ± SE), ranging from 9.10 m to 54.60 m. Among recorded individuals, 16 adults were approached twice during the same season in the same type of habitat. The repeatability of FID was 0.55 (SE = 0.18, p < 0.001), indicating consistency in this behavior. There was no significant difference on FID between marked individuals and non-marked individuals (t = −0.28, p = 0.78).
Comparison of the foraging sites between the plain and the original habitats rendered several differences (Table 1). Those in the original habitat had higher altitude, were less open in terms of terrain openness, and experienced low intensity of human activities.
Table
1.
Comparison of foraging site selection between individuals in the plain habitat and the mountain (original) habitat.
Mountain (original) habitat
Plain habitat
z/t
P
Flock size a
2.24 ± 0.27
2.68 ± 0.31
−1.64
0.101
Distance to the nearest settlement a
168.30 ± 21.08
95.49 ± 9.36
2.66
0.008**
Instant human activity intensity a
1.06 ± 0.11
2.05 ± 0.25
−3.27
0.001**
Altitude b (m)
617.43 ± 17.56
557.73 ± 23.77
−2.02
0.045*
Distance to the nearest road b
87.60 ± 9.49
69.80 ± 8.93
−1.37
0.174
Terrain openness c (°)
117.32 ± 6.60
–
–
–
Valley width c (m)
357.97 ± 64.90
–
–
–
*means p < 0.05, **means p < 0.01. a Difference between original and expending habitat was tested by fitting a Logistic Regression. b Difference between original and expending habitat was tested by t-test. c These factors were only measured in original habitat (mountainous area).
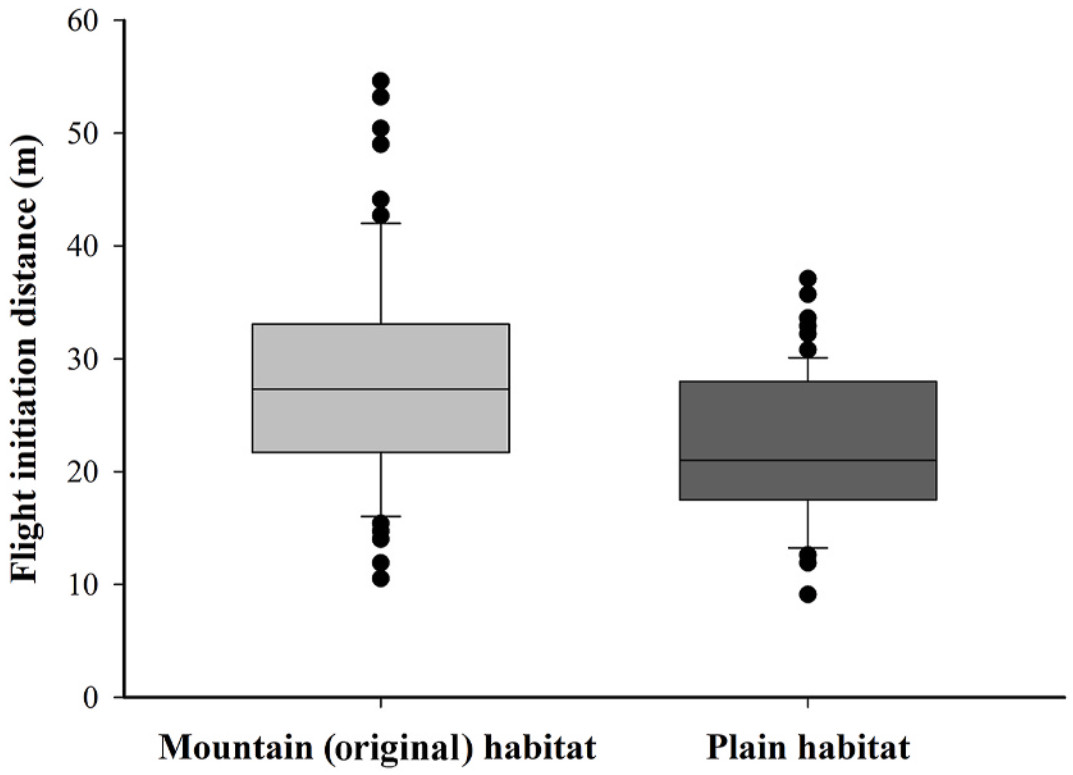
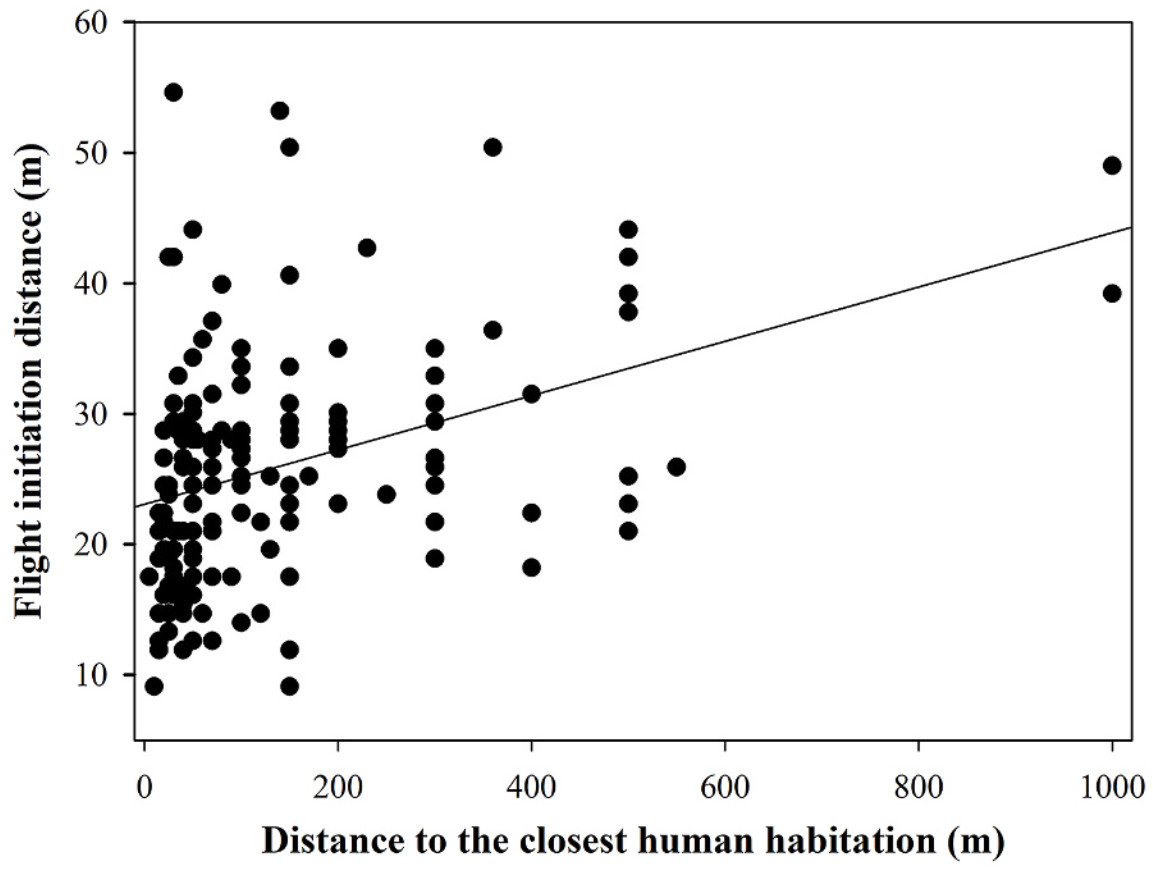
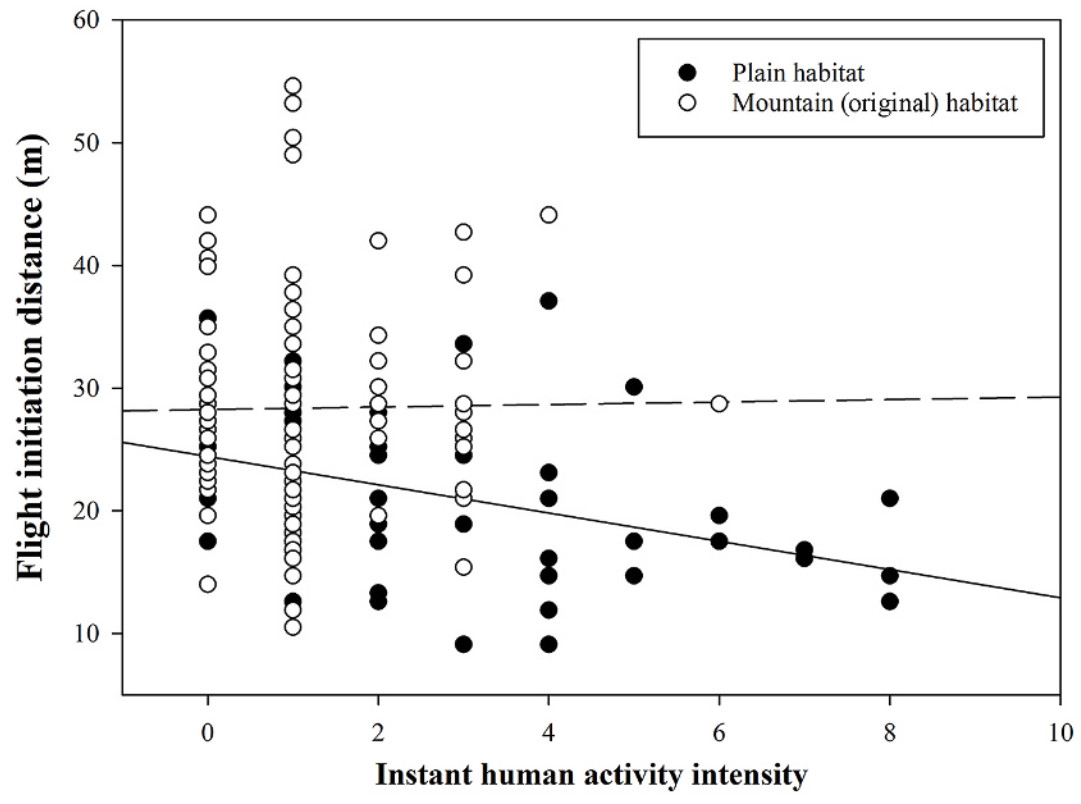
In terms of FID, the FID of ibises foraging in plain area was significantly shorter compared to those in the original area (Fig. 1; Tables 2 and 3). Furthermore, the FID was negatively related with the distance to the nearest settlement (Fig. 2; Tables 2 and 3), while terrain openness (valley width and terrain openness) and group size were not found to significantly impact FID (Tables 2 and 3). These results were consistent for both starting distance and buffer distance as a covariance (Tables 2 and 3). Comparing the models with different covariates, the starting distance has a significant effect on FID (Table 2), while the buffer distance (which is calculated as the starting distance minus the FID) did not (Table 3). Moreover, the interaction between instant human activity intensity and habitat was found to be significant when starting distance (SD) was used as a covariate (Table 2; Fig. 3), whereas this significance was not observed in models employing buffer distance (BD) as the covariate (Table 3).
Figure
1.
The flight initiation distance (m) of Crested Ibis in original (N = 98, grey) and plain (N = 78, black) habitats.
Table
2.
Models of the relationship between mean FID of Crested Ibis and environment variables with starting distance.
Estimate ± SE
χ2/t
P
(A)
(Intercept)
15.39 ± 4.00
Starting distance
0.24 ± 0.04
44.45
<0.001***
Habitats
−0.27 ± 1.57
7.49
0.006**
Flock size
−0.14 ± 0.20
0.45
0.504
Distance to the nearest road
0.001 ± 0.004
0.006
0.937
Distance to the nearest settlement
0.011 ± 0.004
7.91
0.005**
Altitude
−0.005 ± 0.003
2.92
0.087
Instant human activity intensity
3.48 ± 1.46
3.49
0.062
Habitats × Instant human activity intensity
−2.40 ± 0.82
8.64
0.003**
(B)
(Intercept)
17.84 ± 8.49
3.68
0.001**
Starting distance
0.17 ± 0.07
2.44
0.021*
Terrain openness
0.01 ± 0.08
0.14
0.892
Valley width
0.001 ± 0.017
0.09
0.932
Terrain openness × Valley width
−0.00 ± 0.00
−0.11
0.917
The model for both habitat (A) had the statistics r2 = 0.428. The statistics were represented by Chi-square. The model for FID and terrain openness (B) only used the data from the mountain (original) habitat and it had the statistics r2 = 0.230. The statistics were presented as t-values. The starting distance was as a covariance and season was a random variable in both models.
Table
3.
Models of the relationship between mean FID of Crested Ibis and environment variables with buffer distance.
Estimate ± SE
χ2/t
P
(A)
(Intercept)
32.44 ± 4.17
Buffer distance
−0.02 ± 0.05
0.25
0.616
Habitats
−3.25 ± 1.77
14.95
<0.001***
Flock size
0.05 ± 0.24
0.05
0.829
Distance to the nearest road
0.006 ± 0.008
0.49
0.485
Distance to the nearest settlement
0.015 ± 0.004
11.92
<0.001***
Altitude
−0.005 ± 0.003
2.55
0.110
Instant Human activity intensity
2.04 ± 1.67
2.16
0.142
Habitats × Instant human activity intensity
−1.54 ± 0.94
2.69
0.101
(B)
(Intercept)
30.37 ± 8.25
3.68
0.001**
Buffer distance
0.01 ± 0.09
0.14
0.889
Terrain openness
−0.04 ± 0.09
−0.48
0.638
Valley width
0.02 ± 0.02
0.79
0.439
Terrain openness × Valley width
0.00 ± 0.00
−0.30
0.764
The model for both habitat (A) had the statistics r2 = 0.284. The statistics were represented by Chi-square. The model for FID and terrain openness (B) only used the data from the mountain (original) habitat and it had the statistics r2 = 0.080. The statistics were presented as t-values. The buffer distance was a covariate and season was a random variable in both models.
Figure
3.
The flight initiation distance (m) of Crested Ibis in relation to instant human activity intensity (number of humans). The lines are the linear regression lines (Plain area: solid line; Mountain area: dash line).
The Crested Ibis that relocated to the plain area exhibits a relatively shorter FID compared to that of those in the original habitat. The foraging sites selected by the plain groups are closer to human settlements (Song et al., 2018). They displayed a greater tolerance towards humans than those in the original habitat and tended to choose foraging sites where humans are more frequent presence. Based on these facts, it seems that variation in tolerance of human approach may influence the distribution patterns, particularly when animals invade urban habitats (Martin and Réale, 2008; Carrete and Tella, 2010; van Dongen et al., 2015; Worrell et al., 2016). As there is no obvious territorial behavior (Ding, 2010), Crested Ibis can choose and move between original and plain habitats. The foraging site of ibis can be very far apart. On condition of this, we propose that the difference in FIDs between ibises of the two habitats is more likely the result of phenotypic sorting under different pressures, rather than habituation.
The plain group had relatively shorter FIDs. This difference may be due to variations in predation risk between the two habitats because FID is not only an indicator of an individual's tolerance to human activity, but also its risk-taking preference against potential predators (Blumstein, 2006). Predators of Crested Ibis include raptors, snakes and mustelids (Ding, 2004). Snakes and mustelids mainly prey on eggs and nestlings, while raptors target juveniles and adults. Resident birds predators such as Chinese Goshawk (Accipiter soloensis), Northern Goshawk (Accipiter gentilis), Common Buzzard (Buteo buteo) and Common Kestrel (Falco tinnunculus) are common raptors in this region (Ding et al., 2001). The original habitat, being a mountainous area, presents potentially higher predation risk because there are more predators. By contrast, the human-disturbed habitat is almost a refuge for many prey species, since most raptors are not able to access urbanized area (Ibáñez-Álamo and Soler, 2010; Møller, 2012). There is negative correlation between predation risk and FID across both species and individuals (Møller, 2008, 2014). Our study generally is consistent with the pattern that individuals in the original habitat are likely adopting the “safety” strategy while ibises in the plain habitat exhibit the opposite pattern. This difference of FID may also come from food availability. Large area of paddy fields in the plain habitat (t = −10.015, p < 0.0001; Song et al., 2018) provide abundant food resources for Crested Ibis. Ibis may be more reluctant to leave resourceful foraging grounds than relatively barren foraging grounds. This will also make the FID of ibis in the plains shorter. We propose that it is influenced by individuals' diverse escape patterns, which in turn are made based on weighing human activity intensity, predation risk and food resources.
The repeatability of traits, including behaviors, is important to explain the phenotypic correlation. On average, animal behavior repeatability has been reported to be 0.37 (Bell et al., 2009). However, few studies mentioned the repeatability of the FID (e.g., Seltmann et al., 2012), as its challenging to target marked individual or verify individual identity. Some studies have relied on territoriality of reproductive individuals to avoid pseudo replication (e.g., Jiang and Møller, 2017). Fortunately, in the case of ibis, since 2000, the Crested Ibis National Nature Reserve has implemented a system of numbered colored rings to mark all the ibis chicks breeding in the wild. This enabled us to confirm individual identity. The repeatability of FID in our study (r = 0.554) was much higher than the average reported in other behavior study (Bell et al., 2009), indicating that FID is a reliable measurement and a suitable monitoring index for Crested Ibis.
We found a positive relationship between FID and the distance to the nearest settlement. Long-term human activity intensity (settlement) is a stronger predictor of FID than the instant (number of humans). Neither do environmental factors, such as terrain openness and altitude, have a significant effect on FID. These results suggest that escape behavior, as reflected by FID, may not be as flexible as we previously assumed (Carere and van Oers, 2004; Wolf et al., 2007). As this study covered only FID and environmental factors within foraging sites, there is a need to assess the ibises’ entire home range for a comprehensive understanding of other factors influencing their behavior.
One study relevant to ours is Blumstein's (2010) demonstration that FID can be influenced by starting distance (SD) in various taxa. He went on suggesting using SD as an important ecological index due to the increasing energy cost of monitoring (Blumstein, 2010; Engelhardt and Weladji, 2011; Cooper and Blumstein, 2014). However, other researchers have argued that the SD and FID may cause collinearity issues (Møller, 2012) and that SD lacks biologically significance, being merely an artifact (Cooper, 2008; Dyer Dumont et al., 2012). To avoid ambiguity, we used both SD and buffer distance (BD) to represent the influence of approaching. Our results revealed that SD was significantly related with FID, while BD was not. The number of humans around foraging sites varied influence on FID across habitats only when considering SD. Moreover, the relationship between other factors and FID remained unchanged. Although it is challenging to verify whether SD directly affected FID this way, we propose that this novel method may provide a new perspective on reevaluating the significant relationship between SD and FID. The introduction of innovative statistical and/or measuring method could provide further insight into the intricate relationship of these two factors.
In conclusion, our results demonstrate that FID serves as a relatively stable indicator of risk-taking behavior and tolerance to human activity in the Crested Ibis. We found that individuals in the plain habitat have shorter FIDs, indicating a higher level of tolerance to human presence. Additionally, we found a negative correlation between FID and long-term human activity intensity, suggesting that increased human activity influences the ibis’ flight response. The portion relocated to urban areas is more likely a result of phenotypic sorting under different pressures, with potential contributions from learning mechanisms, rather than habituation. These findings carry significant implications for in-situ conservation efforts of threatened species. They also improve our understanding of animal dispersal mechanisms within the context of global urbanization. If FID is regarded as a behavioral trait, conservationists can develop targeted strategies to safeguard and promote the well-being of the Crested Ibis population amid the dynamic changes in its habitat selection. This includes habitat-specific conservation measures tailored to the distinct needs.
Ethics statement
In this study, we did not capture or band birds, but only estimated flight initiation distance (FID) based on observations of long-distance behavior. All our study procedures were approved by Shaanxi Hanzhong Crested Ibis National Nature Reserve.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We really appreciate the support of Yi Liu, Jiaqi Yan and Hongxing Su from Shaanxi Hanzhong Crested Ibis National Nature Reserve for their assistance during the fieldwork. We are grateful to Dr. Yuanxing Ye who kindly gave constructive comments on the manuscript.
Bateman, P.W., Fleming, P.A., 2014. Switching to plan B: changes in the escape tactics of two grasshopper species (Acrididae: Orthoptera) in response to repeated predatory approaches. Behav. Ecol. Sociobiol. 68, 457–465.
Bates, D., Maechler, M., Bolker, B., Walker, S., Christensen, R.H.B., Singmann, H., 2013. lme4: Linear Mixed-Effects Models Using Eigen and S4 (Version 1.0-5). R package. .
Bautista, L.M., García, J.T., Calmaestra, R.G., Palacín, C., Martín, C.A., Morales, M.B., et al., 2004. Effect of weekend road traffic on the use of space by raptors. Conserv. Biol. 18, 726–732.
Bell, A.M., Hankison, S.J., Laskowski, K.L., 2009. The repeatability of behaviour: a meta-analysis. Anim. Behav. 77, 771–783.
Blumstein, D.T., 2003. Flight-initiation distance in birds is dependent on intruder starting distance. J. Wildl. Manag. 67, 852–857.
Blumstein, D.T., 2006. Developing an evolutionary ecology of fear: how life history and natural history traits affect disturbance tolerance in birds. Anim. Behav. 71, 389–399.
Blumstein, D.T., 2010. Flush early and avoid the rush: a general rule of antipredator behavior? Behav. Ecol. 21, 440–442.
Carere, C., van Oers, K., 2004. Shy and bold great tits (Parus major): body temperature and breath rate in response to handling stress. Physiol. Behav. 82, 905–912.
Carrete, M., Martínez-Padilla, J., Rodríguez-Martínez, S., Rebolo-Ifrán, N., Palma, A., Tella, J.L., 2016. Heritability of fear of humans in urban and rural populations of a bird species. Sci. Rep. 6, 31060.
Carrete, M., Tella, J.L., 2010. Individual consistency in flight initiation distances in burrowing owls: a new hypothesis on disturbance-induced habitat selection. Biol. Lett. 6, 167–170.
Carrete, M., Tella, J.L., 2011. Inter-individual variability in fear of humans and relative brain size of the species are related to contemporary urban invasion in birds. PLoS One 6, e18859.
Cooper, W.E., Blumstein, D.T., 2014. Novel effects of monitoring predators on costs of fleeing and not fleeing explain flushing early in economic escape theory. Behav. Ecol. 25, 44–52.
Cooper Jr., W.E., 2008. Strong artifactual effect of starting distance on flight initiation distance in the actively foraging Lizard Aspidoscelis exsanguis. Herpetologica 64, 200–206.
Cresswell, W., 2008. Non-lethal effects of predation in birds. Ibis 150, 3–17.
Ding, C., 2004. Research on the Crested Ibis. Shanghai Scientific and Technological Educational Publishing House, Shanghai.
Ding, C., 2010. Crested ibis. Chinese Birds 1, 156–162.
Ding, C., Liu, D., Yu, X., Chu, G., 2001. The birds in the distribution area of the Crested Ibis (Nipponia nippon). Chin. J. Zool. 37, 48–55.
Donald, P.F., Green, R.E., Heath, M.F., 2001. Agricultural intensification and the collapse of Europe’s farmland bird populations. Proc. R. Soc. A B 268, 25–29.
Dyer Dumont, F., Pasquaretta, C., Réale, D., Bogliani, G., Hardenberg, A., 2012. Flight initiation distance and starting distance: biological effect or mathematical artefact? Ethology 118, 1051–1062.
Engelhardt, S.C., Weladji, R.B., 2011. Effects of levels of human exposure on flight initiation distance and distance to refuge in foraging eastern gray squirrels (Sciurus carolinensis). Can. J. Zool. 89, 823–830.
Evans, J., Boudreau, K., Hyman, J., 2010. Behavioural syndromes in urban and rural populations of song sparrows. Ethology 116, 588–595.
Frid, A., Dill, L.M., 2002. Human-caused disturbance stimuli as a form of predation risk. Conserv. Ecol. 6, 11.
Gil, D., Brumm, H., 2014. Avian Urban Ecology: Behavioural and Physiological Adaptations. Oxford University Press, Oxford.
Hu, C., Song, X., Ding, C., Ye, Y., Qing, B., Wang, C., 2016. The size of winter-flooded paddy fields no longer limits the foraging habitat use of the endangered Crested Ibis (Nipponia nippon) in winter. Zool. Sci. 33, 345–351.
Huang, Y., Ye, Y., Zhang, Y., Barras, A.G., Wang, C., Qing, B., et al., 2022. Tall trees drive the nest-site selection of wild Crested Ibis Nipponia nippon. Bird. Conserv. Int. 32, 486–497.
Ibáñez-Álamo, J.D., Soler, M., 2010. Investigator activities reduce nest predation in blackbirds Turdus merula. J. Avian Biol. 41, 208–212.
IUCN, 2023. The IUCN Red List of Threatened Species. Version 2022-2. .
Jiang, Y., Møller, A.P., 2017. Escape from predators and genetic variance in birds. J. Evol. Biol. 30, 2059–2067.
Li, X., Ma, Z., Ding, C., Zhai, T., Li, D., 2002. Relationship between the distribution of crested ibis and local farmers. Acta Zool. Sin. 48, 725–732.
Liu, Y., 1981. Rediscovery of the crested ibis in qin mountain. Acta Zool. Sin. 27, 273.
Luther, D., Baptista, L., 2010. Urban noise and the cultural evolution of bird songs. Proc. R. Soc. B 277, 469–473.
Martin, J., Réale, D., 2008. Animal temperament and human disturbance: implications for the response of wildlife to tourism. Behav. Process. 77, 66–72.
Møller, A.P., 2008. Flight distance of urban birds, predation, and selection for urban life. Behav. Ecol. Sociobiol. 63, 63–75.
Møller, A.P., 2012. Urban areas as refuges from predators and flight distance of prey. Behav. Ecol. 23, 1030–1035.
Møller, A.P., 2013. Behavioral and ecological predictors of urbanization. In: Gil, D., Brumm, H. (Eds.), Avian Urban Ecology. Oxford University Press, Oxford, pp. 54–68.
Møller, A.P., 2014. Life history, predation and flight initiation distance in a migratory bird. J. Evol. Biol. 27, 1105–1113.
Nakagawa, S., Schielzeth, H., 2010. Repeatability for Gaussian and non-Gaussian data: a practical guide for biologists. Biol. Rev. 85, 935–956.
R Core Team, 2021. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. .
Schell, C.J., 2018. Urban evolutionary ecology and the potential benefits of implementing genomics. J. Hered. 109, 138–151.
Seltmann, M.W., Öst, M., Jaatinen, K., Atkinson, S., Mashburn, K., Hollmén, T., 2012. Stress responsiveness, age and body condition interactively affect flight initiation distance in breeding female eiders. Anim. Behav. 84, 889–896.
Shochat, E., Warren, P.S., Faeth, S.H., McIntyre, N.E., Hope, D., 2006. From patterns to emerging processes in mechanistic urban ecology. Trends Ecol. Evol. 21, 186–191.
Sih, A., Ferrari, M.C., Harris, D.J., 2011. Evolution and behavioural responses to human-induced rapid environmental change. Evol. Appl. 4, 367–387.
Song, Z., Zou, Y., Hu, C., Ye, Y., Wang, C., Qing, B., et al., 2018. Silver spoon effects of hatching order in an asynchronous hatching bird. Behav. Ecol. 30, 509–517.
Tablado, Z., Jenni, L., 2015. Determinants of uncertainty in wildlife responses to human disturbance. Biol. Rev. 92, 216–233.
Tarjuelo, R., Barja, I., Morales, M.B., Traba, J., Benítez-López, A., Casas, F., et al., 2015. Effects of human activity on physiological and behavioral responses of an endangered steppe bird. Behav. Ecol. 26, 828–838.
Tatner, P., Bryant, D.M., 1986. Flight cost of a small passerine measured using doubly labeled water: implications for energetics studies. Auk 103, 169–180.
Tomialojc, L., 1982. Synurbanization of Birds and Prey–Predators Relations. Animals in Urban Environments. Ossolineum, Wroclaw, Poland, pp. 131–137.
Valcarcel, A., Fernández-Juricic, E., 2009. Antipredator strategies of house finches: are urban habitats safe spots from predators even when humans are around? Behav. Ecol. Sociobiol. 63, 673–685.
van Dongen, W.F.D., Robinson, R.W., Weston, M.A., Mulder, R.A., Guay, P.-J., 2015. Variation at the DRD4 locus is associated with wariness and local site selection in urban black swans. BMC Evol. Biol. 15, 253.
Webb, N.V., Blumstein, D.T., 2005. Variation in human disturbance differentially affects predation risk assessment in western gulls. Condor 107, 178–181.
Weston, M., McLeod, E.M., Blumstein, D., Guay, P.-J., 2012. A review of flight-initiation distances and their application to managing disturbance to Australian birds. Emu 112, 269–286.
Wolf, M., van Doorn, G.S., Leimar, O., Weissing, F.J., 2007. Life-history trade-offs favour the evolution of animal personalities. Nature 447, 581–584.
Worrell, T., Admiraal, R., Bateman, P., Fleming, P., 2016. Are tourism and conservation compatible for ‘island tame’species? Anim. Conserv. 20, 155–163.
Xu, X., Ye, Y., Briggs, E., Wang, C., Qing, B., Song, Z., et al., 2023. Why do parents produce small broods of offspring that have lower body mass, survival, and lifetime reproductive success? A case study in a long-lived bird. Behav. Ecol. Sociobiol. 77, 30.
Ye, Y., Santoro, S., Song, Z., Hu, C., Zhang, Z., Qing, B., et al., 2022. Dispersal patterns of the endangered Crested Ibis suggest high breeding densities drive natal dispersal. Ornithol. Appl. 125, duac042.
Yeh, P.J., Hauber, M.E., Price, T.D., 2007. Alternative nesting behaviours following colonisation of a novel environment by a passerine bird. Oikos 116, 1473–1480.
Zanette, L.Y., White, A.F., Allen, M.C., Clinchy, M., 2011. Perceived predation risk reduces the number of offspring songbirds produce per year. Science 334, 1398–1401.
Table
1.
Comparison of foraging site selection between individuals in the plain habitat and the mountain (original) habitat.
Mountain (original) habitat
Plain habitat
z/t
P
Flock size a
2.24 ± 0.27
2.68 ± 0.31
−1.64
0.101
Distance to the nearest settlement a
168.30 ± 21.08
95.49 ± 9.36
2.66
0.008**
Instant human activity intensity a
1.06 ± 0.11
2.05 ± 0.25
−3.27
0.001**
Altitude b (m)
617.43 ± 17.56
557.73 ± 23.77
−2.02
0.045*
Distance to the nearest road b
87.60 ± 9.49
69.80 ± 8.93
−1.37
0.174
Terrain openness c (°)
117.32 ± 6.60
–
–
–
Valley width c (m)
357.97 ± 64.90
–
–
–
*means p < 0.05, **means p < 0.01. a Difference between original and expending habitat was tested by fitting a Logistic Regression. b Difference between original and expending habitat was tested by t-test. c These factors were only measured in original habitat (mountainous area).
Table
2.
Models of the relationship between mean FID of Crested Ibis and environment variables with starting distance.
Estimate ± SE
χ2/t
P
(A)
(Intercept)
15.39 ± 4.00
Starting distance
0.24 ± 0.04
44.45
<0.001***
Habitats
−0.27 ± 1.57
7.49
0.006**
Flock size
−0.14 ± 0.20
0.45
0.504
Distance to the nearest road
0.001 ± 0.004
0.006
0.937
Distance to the nearest settlement
0.011 ± 0.004
7.91
0.005**
Altitude
−0.005 ± 0.003
2.92
0.087
Instant human activity intensity
3.48 ± 1.46
3.49
0.062
Habitats × Instant human activity intensity
−2.40 ± 0.82
8.64
0.003**
(B)
(Intercept)
17.84 ± 8.49
3.68
0.001**
Starting distance
0.17 ± 0.07
2.44
0.021*
Terrain openness
0.01 ± 0.08
0.14
0.892
Valley width
0.001 ± 0.017
0.09
0.932
Terrain openness × Valley width
−0.00 ± 0.00
−0.11
0.917
The model for both habitat (A) had the statistics r2 = 0.428. The statistics were represented by Chi-square. The model for FID and terrain openness (B) only used the data from the mountain (original) habitat and it had the statistics r2 = 0.230. The statistics were presented as t-values. The starting distance was as a covariance and season was a random variable in both models.
Table
3.
Models of the relationship between mean FID of Crested Ibis and environment variables with buffer distance.
Estimate ± SE
χ2/t
P
(A)
(Intercept)
32.44 ± 4.17
Buffer distance
−0.02 ± 0.05
0.25
0.616
Habitats
−3.25 ± 1.77
14.95
<0.001***
Flock size
0.05 ± 0.24
0.05
0.829
Distance to the nearest road
0.006 ± 0.008
0.49
0.485
Distance to the nearest settlement
0.015 ± 0.004
11.92
<0.001***
Altitude
−0.005 ± 0.003
2.55
0.110
Instant Human activity intensity
2.04 ± 1.67
2.16
0.142
Habitats × Instant human activity intensity
−1.54 ± 0.94
2.69
0.101
(B)
(Intercept)
30.37 ± 8.25
3.68
0.001**
Buffer distance
0.01 ± 0.09
0.14
0.889
Terrain openness
−0.04 ± 0.09
−0.48
0.638
Valley width
0.02 ± 0.02
0.79
0.439
Terrain openness × Valley width
0.00 ± 0.00
−0.30
0.764
The model for both habitat (A) had the statistics r2 = 0.284. The statistics were represented by Chi-square. The model for FID and terrain openness (B) only used the data from the mountain (original) habitat and it had the statistics r2 = 0.080. The statistics were presented as t-values. The buffer distance was a covariate and season was a random variable in both models.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds