Chao Yu, Xuying Lu, Deli Sun, Mengnan Chu, Xueyun Li, Qun Li. 2024: River width and depth as key factors of diurnal activity energy expenditure allocation for wintering Spot-billed Ducks in the Xin’an River Basin. Avian Research, 15(1): 100159. DOI: 10.1016/j.avrs.2024.100159
Citation:
Chao Yu, Xuying Lu, Deli Sun, Mengnan Chu, Xueyun Li, Qun Li. 2024: River width and depth as key factors of diurnal activity energy expenditure allocation for wintering Spot-billed Ducks in the Xin’an River Basin. Avian Research, 15(1): 100159. DOI: 10.1016/j.avrs.2024.100159
Chao Yu, Xuying Lu, Deli Sun, Mengnan Chu, Xueyun Li, Qun Li. 2024: River width and depth as key factors of diurnal activity energy expenditure allocation for wintering Spot-billed Ducks in the Xin’an River Basin. Avian Research, 15(1): 100159. DOI: 10.1016/j.avrs.2024.100159
Citation:
Chao Yu, Xuying Lu, Deli Sun, Mengnan Chu, Xueyun Li, Qun Li. 2024: River width and depth as key factors of diurnal activity energy expenditure allocation for wintering Spot-billed Ducks in the Xin’an River Basin. Avian Research, 15(1): 100159. DOI: 10.1016/j.avrs.2024.100159
Rivers are important habitats for wintering waterbirds. However, they are easily influenced by natural and human activities. An important approach for waterbirds to adapt to habitats is adjusting the activity time and energy expenditure allocation of diurnal behavior. The compensatory foraging hypothesis predicts that increased energy expenditure leads to longer foraging time, which in turn increases food intake and helps maintain a constant energy balance. However, it is unclear whether human-disturbed habitats result in increased energy expenditure related to safety or foraging. In this study, the scan sample method was used to observe the diurnal behavior of the wintering Spot-billed Duck (Anas poecilorhyncha) in two rivers in the Xin'an River Basin from October 2021 to March 2022. The allocation of time and energy expenditure for activity in both normal and disturbed environments was calculated. The results showed that foraging accounted for the highest percentage of time and energy expenditure. Additionally, foraging decreased in the disturbed environment than that in the normal environment. Resting behavior showed the opposite trend, while other behaviors were similar in both environments. The total diurnal energy expenditure of ducks in the disturbed environment was greater than that in the normal environment, with decreased foraging and resting time percentage and increased behaviors related to immediate safety (swimming and alert) and comfort. These results oppose the compensatory foraging hypothesis in favor of increased security. The optimal diurnal energy expenditure model included river width and water depth, which had a positive relationship; an increase in either of these two factors resulted in an increase in energy expenditure. This study provides a better understanding of energy allocation strategies underlying the superficial time allocation of wintering waterbirds according to environmental conditions. Exploring these changes can help understand the maximum fitness of wintering waterbirds in response to nature and human influences.
A study has confirmed that birds possess the ability to adapt their activity patterns and energy allocations in response to energy demands (Pelletier et al., 2008). The compensatory foraging hypothesis predicts that when energy expenditure increases, waterbirds increase their food intake and the time allocation of foraging behaviors to ensure a constant energy balance (Montgomery and Baumgardt, 1965; Peace and Wang, 2019). Studies have found that waterbirds increase foraging time (Yu et al., 2020) to compensate for feed reduction (Yu et al., 2019b) by reducing the time spent on behaviors that are unrelated to energy intake, such as alerting, fighting, and resting (Li et al., 2015). However, safety is an important consideration in examining the energy expenditure of waterbirds in disturbed environments. Striking a delicate balance, these birds must invest additional energy to ensure their security, while the impact of their foraging activities may only become evident over a prolong period of time (Ito, 2019). For example, when human disturbance increases, more vigilance time is invested (Yu et al., 2019b) and the allocation of time for searching for food is reduced compared to a safe environment, thereby affecting the costs of vigilance and foraging (Gyimesi et al., 2012). It is widely recognized that behavioral coefficients and energy expenditure vary across different types of behaviors (Nolet et al., 2001; Engelhardt et al., 2014). Waterbirds exhibit various behavioral adjustments influenced by habitat factors, resulting in fluctuations in their time allocations. Additionally, changes in behavior time allocation can also impact energy expenditure. Consequently, daily energy expenditure exhibits significant variation. Waterbirds alter their behavior activities, time allocation, and energy expenditure in response to worsening environmental conditions, aiming to increase energy intake and minimize total diurnal energy expenditures. Therefore, the strategy, time, and energy allocation of behaviors of animals choosing to change habitats is important to understand.
The rivers in the Xin'an River Basin are an important wintering waterbird habitat in the East Asian–Australasian Flyway. The rivers have narrow channels, and their hydrology, especially river depth, width, and velocity, rapidly changes. In addition, the rivers are long and pass through different natural and human environments, with anthropogenic disturbances potentially affecting the habitat chosen by wintering birds. Accordingly, waterbirds adjust their behavior and energy costs to maximize their fitness. During the winter season, Spot-billed Ducks are commonly sighted in the Xin'an River Basin. They exhibit a range of behaviors including food foraging, responding to human disturbances, and resting on the water and rocks in the rivers. Understanding the effect of the wintering strategy of waterbirds on behavioral time and energy expenditure budgets is important for understanding the future effects of environmental changes to their habitats.
In this study, the time allocation of different behaviors of Spot-billed Ducks in a changing environment was observed and recorded, and the energy expenditure budget and total value of each behavior in different environments during the day were calculated. Simultaneously, we conducted an analysis of the diurnal energy expenditure characteristics of wintering Spot-billed Ducks in rivers. Furthermore, we identified the environmental factors that influence the allocation strategies of diurnal energy expenditure. We hypothesized that as total energy expenditure increases, so does foraging-related behaviors (according to the compensatory foraging hypothesis); however, energy expenditure that relates to ensuring security is also likely to increase. Therefore, we sought to understand how Spot-billed Ducks adjust the energy expenditure of different behaviors in the case of worsened environmental conditions, such as increased water depth, velocity, and disturbance, as well as the extent to which environmental factors affect energy expenditure.
2.
Materials and methods
2.1
Study sites
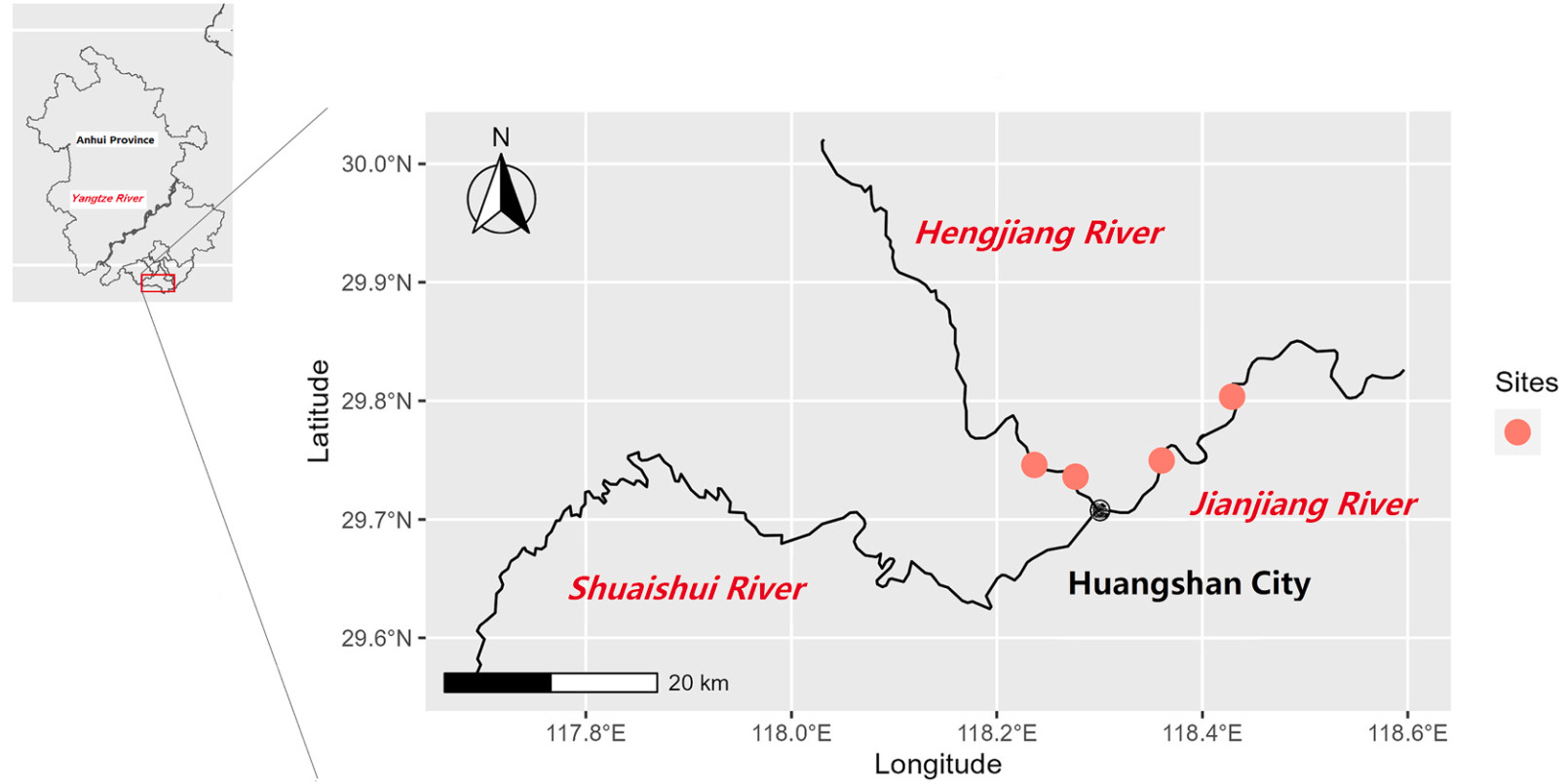
Observational studies were carried out in two rivers in the Xin'an River Basin: Jianjiang River (29.71–29.72° N, 118.30–118.72° E) and Hengjiang River (29.71–30.01° N, 117.95–118.30° E), Huangshan, Anhui, China (Fig. 1). These rivers are traditionally wintering refuges and are supplements on habitats for waterbirds on the East Asian–Australasian Flyway. Waterbirds inhabited several locations in Xin'an River Basin, and ducks observed foraging in these sites included the Spot-billed Duck (Anas poecilorhyncha), Common Teal (Anas crecca), and Mallard (Anas platyrhynchos). The two rivers investigated cross dozens of kilometers with villages and towns scattered on relatively flat river plain. The Jianjiang River is wider, with less human activity and mainly grassland landscape on both sides. The width of the Hengjiang River is relatively narrow (about 150–200 m), with grassland on one side and a stone dam on the other. Residents use the shores of these rivers for activities such as washing, recreational walking, and pole or net fishing. This means that there was substantial impact from human activities on the waterbirds' habitats.
Figure
1.
Habitats of wintering Spot-billed Ducks in the Xin’an River Basin in Huangshan, Anhui Province, China. Red dots indicate the study sites.
The study was conducted from October 2021 to March 2022, which is the wintering period of waterbirds. The instantaneous scanning method was used to record individual behavior types at intervals of 30 min. Behaviors included foraging, resting, comfort, alert (in ducks this was defined as stretching their neck upright and turning their head around, sometimes accompanied by vocalization and swimming), swimming, sociality, flying, and walking. Spot-billed Ducks generally inhabited the rivers for 3–10 h from morning to evening; however, the ducks flew away from the rivers if disturbances were serious. The observations occurred from 7:30 to 17:30 under normal circumstances or whenever the areas of the river with disturbances were inhabited. A total of 129 scanning samples were taken over 15 days.
The types of disturbances recorded included washing, grazing, pedestrians, and fishermen. We also recorded the disturbance distance (the shortest distance between the disturbance source and the Spot-billed Duck) and disturbance number (the number of disturbance sources appearing at the duck horizon). The disturbance distance was measured using a laser rangefinder telescope (Zeiss Victory Scope, 10 × 42, Jena, Germany). If any disturbance source in the horizon of the Spot-billed Duck was scanned, the record was regarded as a disturbance sample; otherwise, it was considered as a normal sample.
We recorded the relative water velocity, river width, and water depth where the Spot-billed Duck was located as well as the air temperature and wind speed (sourced from http://www.weather.com.cn/) when scanning behaviors of ducks once. We estimated water depth by the degree the legs or body of a Great Egret (Ardea alba) or White Egret (Egretta garzetta) near a fixed target (such as a stone in river) were submerged, following He et al. (2016), and divided it into five grades: < 0.1, 0.1–0.2, 0.2–0.3, 0.3–0.4, and 0.4–0.5 m. The relative water velocity was measured by a flowmeter (Xiangruide, LS1206b, Nanjing, China) with a lower detection limit of 0.06 m/s. Because ducks are distributed in large areas, the water velocity varied at different locations, making it difficult to obtain precise water velocity; therefore, we put the flowmeter at sites 5 m from the fixed spot on the shore, which gained relative water velocity without disturbing the ducks. The relative water velocity was divided into seven grades: < 0.06, 0.06–0.12, 0.12–0.18, 0.18–0.24, 0.24–0.30, 0.30–0.36, and > 0.36 m/s.
2.3
Statistical analysis
We calculated the proportion of different behavior types exhibited in the population (number of ducks exhibiting the behavior type/population number) and obtained the allocation percentage for each behavior in the observed sample unit (a scan sample). On this basis, the diurnal activity time allocation of each behavior was calculated by multiplying the inhabiting duration by each behavior allocation percentage of the sample. We then multiplied that result with the resting metabolic rate (RMR, 514.98 kJ/24 h) (Jones et al., 2014) and its multiple of each behavior. We used the following formula:
(1)
where xe is the activity energy expenditure of behavior x; i is the allocation percentage of behavior x in scan example; xr is the multiple of RMR for the behavior x; and T is the inhabiting duration of waterbirds in rivers observed in a day. The multiple of RMR of each behavior was 1.2 for resting and foraging, 2.2 for swimming and alert, 2.1 for comfort, 12.5 for flying, 2.4 for sociality, and 1.7 for walking (Wooley and Owen, 1977; Jones et al., 2014). Diurnal energy expenditure was then the sum of the energy expenditure of different behavioral types during the day.
We analyzed the normal distributions and homoscedasticity in the allocation of time and energy expenditure of behaviors. Subsequently, we compared these distributions using either Student's t-test or nonparametric tests for normally or non-normally distributed parameters, respectively. Spearman's tests were used to test the relative correlations between the disturbance distance, disturbance number, water depth, relative water velocity, river width, air temperature, and wind velocity with the allocation of time and energy expenditure of behaviors. Generalized linear models were assessed to identify the microhabitat factors influencing total energy expenditure to different extents.
Total energy expenditure was Log10 transformed and fitted to the predictors of environmental factors relative to total energy expenditure. When the Akaike weight of the best-ranked model (using the corrected Akaike information criterion; AICc), considerably surpassed that of the subsequent model, inferences were solely drawn from the former (Symonds and Moussalli, 2011). Otherwise, all models with Δi < 4 were averaged and the relative importance of the predictor variables was estimated by summing the Akaike weights of the variables that appeared in each model (Symonds and Moussalli, 2011). We used the multi-model inference package 'MuMIn' (Bartoń, 2015) in R (version 4.1.2) (R Core Team, 2023) to apply the selected models and average the results.
Flying, walking, and social behaviors occurred less frequently and are expressed as mean ± standard error, while other behaviors are expressed as mean ± standard deviation. The significance level for all statistical tests was set at p < 0.05.
3.
Results
3.1
Time allocation of diurnal behaviors
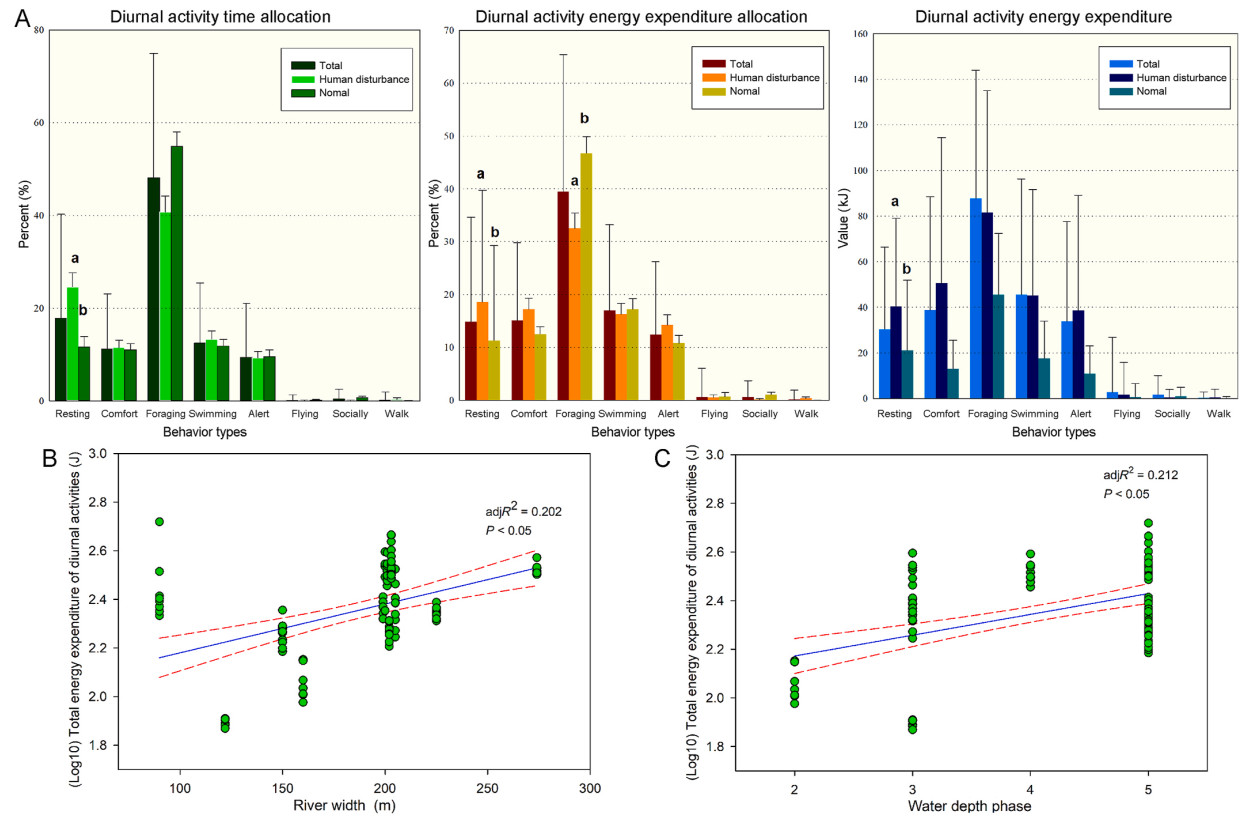
The study involved 129 scanning samples (62 disturbed and 67 normal samples) of Spot-billed Ducks in the two rivers. Foraging was the most frequently observed behavior, and it was observed less in a disturbed environment than in a normal environment (F = 0.376, p = 0.004). In contrast, resting behavior was observed more frequently in a disturbed environment than in a normal environment (Z = −2.397, p = 0.017). Of the other main behaviors, no significant difference was observed between disturbed and normal environments for swimming (Z = −0.178, p = 0.859), comfort (Z = −0.088, p = 0.93) and alert (Z = −0.076, p = 0.939) (Fig. 2; Table 1).
Figure
2.
Diurnal activity time and energy expenditure allocation depending on behaviors, water depth, and river width in Xin'an River Basin indicate that Spot-billed Ducks employ flexible time and energy expenditure strategies to adapt to heterogeneous environmental conditions. (A) The variation in diurnal activity time, energy expenditure allocation and energy expenditure values, different letters (a, b) indicate significant differences at p < 0.05. (B) The total energy expenditure of diurnal activities changed with water depth. The green points represent water samples. The red lines represent 95% confidence intervals. (C) Spot-billed Ducks showed different total energy expenditure on diurnal activities as a function of river width. The green points represent water samples. The red lines represent 95% confidence intervals.
Table
1.
The allocation of time and energy expenditure of behaviors for the Spotted-billed Ducks in disturbed and normal habitats at Xin'an River Basin in 2021–2022.
3.2
Energy expenditure allocation of diurnal behaviors
The total diurnal energy expenditure of Spot-billed Ducks was 239.972 ± 94.444 kJ. Under disturbed conditions, the total diurnal energy expenditure was 260.749 ± 85.517 kJ, which was greater than that in normal conditions (220.745 ± 98.786 kJ) (Z = −2.218, p = 0.027). The most energy-consuming behavior was foraging, and its contribution to total diurnal energy expenditure in the disturbed environment was lower than that in the normal environment (F = −2.718, p = 0.007). In contrast, the contribution of resting behavior to total diurnal energy expenditure was higher in the disturbed environment than that in the normal environment (Z = −2.435, p = 0.015). No significant difference in the total diurnal energy expenditure contribution was observed between disturbed and normal environments for the other behaviors, such as comfort (Z = −1.235, p = 0.217), swimming (Z = −0.463, p = 0.643), and alert (Z = −1.114, p = 0.265) (Fig. 2; Table 1).
When total energy expenditure increased, the percentage of foraging (r = −0.288, p = 0.001) and resting energy expenditure (r = −0.297, p = 0.001) decreased. In contrast, the percentage of comfort (r = 0.181, p = 0.004), swimming (r = 0.314, p < 0.001), and alert (r = 0.268, p = 0.002) energy expenditure increased. Meanwhile, foraging time percentage did not change (r = −0.122, p = 0.170), resting time percentage decreased (r = −0.176, p = 0.046); however, swimming (r = 0.238, p = 0.007) and alert time percentage (r = 0.281, p = 0.001) increased. These results indicated that when total energy expenditure increased, the ducks adjusted energy expenditure and time allocation; more energy was allocated to alert and swimming, which directly relates to safety and comfort, but not to foraging.
3.3
Relationship between diurnal energy expenditure and environment factors
When the water depth increased, the total energy expenditure increased (r = 0.282, p = 0.003). Analysis of individual behaviors showed that alert (r = 0.212, p = 0.027), foraging (r = 0.188, p = 0.05), and swimming (r = 0.219, p = 0.022) energy expenditure percentage increased as the water depth increased, as well as the swimming time percentage (r = 0.227, p = 0.017). However, resting energy expenditure percentage decreased (r = −0.321, p = 0.001) as the water depth increased. As relative water velocity increased, diurnal total energy expenditure (r = 0.299, p = 0.002) increased. Alert energy expenditure (r = 0.347, p < 0.001) and time percentage (r = 0.332, p < 0.001) also increased as the relative water velocity increased. In contrast, both foraging behaviors decreased with an increase in the relative water velocity (renergy expenditure percentage = −0.292, p = 0.002; rtime percentage = −0.418, p < 0.001). River width only had a positive relationship with diurnal total energy expenditure (r = 0.396, p < 0.001). The comfort behavior time percentage (r = −0.288, p = 0.032) was negatively correlated to water depth. We found that both water depth (r = 0.298, p = 0.026) and river width (r = 0.370, p = 0.005) positively affected total energy expenditure in the normal environment. Diurnal energy expenditure of Spot-billed Ducks had no relationship with wind velocity (r = −0.145, p = 0.141), but rather with foraging time (r = 0.198, p = 0.043). Diurnal total energy expenditure percentage was had no relationship with air temperature (r = −0.155, p = 0.117), but did have a positive relationship with resting energy expenditure percentage (r = 0.342, p < 0.001), foraging showed the opposite trend (r = −0.526, p = 0.000). Disturbance distance had a negative relationship with foraging behavior (renergy expenditure percentage = −0.393, p = 0.004; rtime percentage = −0.374, p = 0.006), whereas no relationship was observed between disturbance distance (r = 0.261, p = 0.062), which meant that when the disturbance distance became longer, the diurnal total energy consumption would be not changed, foraging energy consumption and time percentage were less. Disturbance number and diurnal total energy expenditure had no relationship (r = 0.101, p = 0.256). However, diurnal total energy expenditure increased with fishermen disturbance number (r = 0.585, p = 0.014). Foraging time percentage (r = −0.210, p = 0.017) decreased with disturbance number. In contrast, both resting behaviors increased with disturbance number (renergy expenditure percentage = 0.204, p = 0.002; rtime percentage = 0.284, p = 0.001). In general, when the water depth and velocity increased, alert (2.2 multiple of RMR) or swimming (2.2 multiple of RMR) behaviors with high cost were used instead of low-cost resting (1.2 multiple of RMR) or foraging (1.2 multiple of RMR). When the disturbance number and air temperature increased, resting was observed more than foraging (same cost: 1.2 multiple of RMR). The diurnal total energy expenditure increased with river width, but there was no substitution between the behaviors.
Energy expenditure fit the predictors of water depth, relative water velocity, and river width. Eight models were tested and ranked according to the AICc (Table 2). Candidate models were set to include all predictors. The cumulative weight of these models (Δi < 4) was 1.00. The best model which included potential predictors of water depth and river width; with a model weight of 0.52, water depth and river width were significant in the model. The standardized and model-averaged parameter estimates for the models are presented in Table 3. The optimal energy expenditure model included two variables: river width and water depth. Their coefficients were positive, indicating that the energy expenditure increased with these environmental factors.
Table
2.
Model selection tables-ranked generalized linear mixed models in the candidate set for the diurnal total energy expenditure of wintering Spot-billed Ducks at Xin'an River Basin.
Candidate models
df
AICc
Δi
ωi
acc ωi
Log10 (Diurnal total energy expenditure) = Water depth + River width + Relative water velocity
1 Water depth + River width
4
−94.69
0
0.52
0.52
2 Water depth + River width + Relative water velocity
5
−94.55
0.13
0.48
1
3 River width + relative water velocity
4
−83.46
11.22
0
1
4 Water depth
3
−73.42
21.27
0
1
5 Water depth + Relative water velocity
4
−72.32
22.37
0
1
6 River width
3
−71.96
22.73
0
1
7 Relative water velocity
3
−58.27
36.42
0
1
8 Null
2
−48.49
46.2
0
1
Δi, difference in AICc between model and top model; ωi, Aikaike model weight; acc ωi, cumulative model weight.
Table
3.
Parameter estimates, standard errors, and confidence intervals for the potential predictors of the diurnal total energy expenditure of wintering Spot-billed Ducks. Dependent variables have been Log10 transformed.
Predictor
Relative importance
Estimate
S.E.
95% confidence interval
Lower limit
Upper limit
Intercept
1.736
0.081
1.575
1.898
Water depth
1
0.067
0.016
0.034
0.099
River width
1
0.002
0
0.001
0.002
Relative water velocity
0.48
0.005
0.007
−0.004
0.025
Relative importance: sum of the Akaike weights for each variable across all the models where it occurred (see Materials and methods); S.E.: standard error.
In this study, we rejected the compensatory foraging hypothesis that ducks compensate for their diurnal total energy cost by allocating more time and energy for foraging. We found that the ducks increased their energy cost percentage directly related to safety, but not foraging and resting behavior, which indicated their flexible energy expenditure strategies. Factors such as river depth and width positively impacted the diurnal total energy cost.
4.1
Diurnal behavior time allocation
Behaviors related to foraging are important for wintering waterbirds, and are based on energy intake demand. Foraging, resting, swimming, and alert were the main behaviors of Spot-billed Ducks in different environments. Foraging behavior in normal environments was greater than that in disturbed environments, likely because waterbirds use the winter to recuperate lost energy from the summer (Wan et al., 2016; Yu et al., 2019b). Increased resting behaviors in disturbed environments may have occurred because resting decreases total energy expenditure and preserves strength, similar to the behavior of farm-raised chickens (Forslind et al., 2021), which allows the waterbirds to compensate for the lack of energy increase if they are unable to forage effectively in a disturbed environment that does not directly threaten their safety. In addition, ducks usually rested in the warmth at noon, which was when there was also a higher occurrence of washing, a common type of disturbance in the study rivers. Swimming and alert behaviors were direct responses to avoid distractions in a disturbing environment, while swimming was also frequent between foraging bouts and to avoid crashes or conflict with other ducks (Yu et al., 2019b).
4.2
Diurnal behavior energy expenditure allocation
Waterbirds have varying energy expenditures in different environments. Although foraging, comfort, swimming, and alert energy expenditure percentage accounted for more than 10% of all behavioral energy expenditure in the disturbed environments, we found foraging energy expenditure decreased and resting increased in disturbed environments compared with normal environments. These results suggest that wintering waterbirds concentrate their time and energy on resting in areas with high disturbance, transferring the high energy expenditure of foraging to other low-energy behaviors when it cannot directly replenish energy by foraging (Zimmer et al., 2011). The energy expenditure of comfort behavior was similar in normal and disturbed environments in our study; however, previous studies have shown that birds emerge from preening (Zhao et al., 2020), bathing, and other compensatory behaviors after disturbances, resulting in less energy expenditure in comfort behavior in disturbed environments (Zimmer et al., 2011). In this study, we found that the diurnal total energy expenditure in the disturbance environment was greater than that in the normal environment. When diurnal total energy expenditure increased, the ducks adjusted the energy expenditure allocation percentage of comfort, alert, and swimming. These behaviors directly relate to safety, not to foraging, which does not conform to the compensatory hypothesis, possibly because there were too few daylight hours for activity (Magella et al., 2012). At the study sites, washers, fishermen, and walkers were always present at some point during the day, which stressed nearby ducks. Safety is a foundational benefit for waterbirds; therefore, they invest more energy in behaviors related to avoiding potential danger and ensuring security (Collop et al., 2016; Li et al., 2015).
4.3
Environment factors affecting diurnal energy expenditure
The diurnal energy expenditure of the Spot-billed Ducks increased with increasing water depth, river velocity and river width. An increase in water velocity and depth leads to increased energy use as the waterbirds are required to swim more to resist the current and dive deeper to find food (Nolet et al., 2001; Wood et al., 2013). The increase in alert, foraging, and swimming resulted in an increase in the expenditure ratio of high-energy foraging and a decrease in low-energy resting, resulting in an overall increase in diurnal total energy expenditure. The diurnal energy expenditure of Spot-billed Ducks increased with fishermen disturbance, mainly because fishermen were present in the middle of rivers, were mobile, and had a shorter disturbance distance. This caused waterbirds to abandon low-energy resting in favor of other behaviors related to safety that had a higher energy expenditure rate, resulting in an overall increase in diurnal energy expenditure. This finding is supported by a previous study (Schummer and Eddleman, 2003). The factors in the optimal model influencing the total diurnal energy expenditure were river width and water depth. As river depth increased, the proportion of swimming time allocation increased, because ducks must engage in swimming behavior when performing any other behaviors, such as foraging or resting. While there was no relationship between water width and time allocation of behaviors, river width had a positive correlation to foraging and swimming energy consumption, which resulted in an increase in total energy consumption. River width and depth were both positively related to energy consumption, and their impacts were usually mixed, especially in the Xin'an River Basin. Because the channel was narrow and the banks were stone or artificial dikes, when the river width was wider, the water was deeper, and the ducks needed more energy to forage effectively.
4.4
Limitations and future studies
There were a number of limitations to this study. First, the fieldwork methods employed meant that the time and distance of a duck's flight could not be determined; therefore, the energy consumption of flight was not calculated. The energy consumption of flight is higher than that of non-flight energy expenditure (Bairlein et al., 2015). Changes to flight behaviors in disturbed environments compared with normal environments could have impacted the diurnal total energy expenditure (Nolet et al., 2016). Second, we could not determine the type of food the ducks were consuming; therefore, energy intake was not considered. There is a close relationship between energy expenditure and energy intake (Elliott et al., 2008b).
For future studies, we aim to determine the nutrient intake to better understand the balance between energy expenditure and energy intake in disturbed environments compared with normal environments.
5.
Conclusion
This study demonstrated the effects of river width and water depth on the diurnal total energy expenditure patterns of Spot-billed Ducks. The results indicated diverse activity time and energy expenditure patterns in response to changes in river factors in heterogeneous river habitats. The ducks did not compensate for their overall energy cost with more time allocation for foraging. Instead, they increased their total energy cost percentage directly related to safety, which indicated their flexible energy expenditure strategy. The diurnal total energy cost increased with river depth and width. This study suggests that at the energy level, energy expenditure may provide an alternative for understanding behavior adaption strategies. This study provides a better understanding of energy allocation strategies underlying the superficial time allocation of wintering waterbirds according to environmental conditions. To improve habitat management, there should be a focus on river depth and width as the key factors for river habitat conservation.
CRediT authorship contribution statement
Chao Yu: Writing – review & editing, Writing – original draft, Visualization, Validation, Resources, Project administration, Methodology, Investigation, Funding acquisition, Formal analysis, Data curation, Conceptualization. Xuying Lu: Writing – original draft, Software, Methodology, Formal analysis, Data curation. Deli Sun: Software, Methodology, Data curation. Mengnan Chu: Software, Investigation, Data curation. Xueyun Li: Investigation, Data curation. Qun Li: Investigation, Data curation.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
We thank the people who helped collect the behavior and microhabitat data in Shuaishui, Hengjiang, Jianjiang, including Letun Zhang, Yi Ni, Shen Liu, Dequan Zeng, to Ying Yu, Lei Cheng for statistical assistance, Shunqing Lv and Wei Li for their invaluable suggestions and revisions to the original manuscript, and constructive comments of two anonymous reviewers. We would like to thank Editage (www.editage.cn) for English language editing.
Bairlein, F., Fritz, J., Scope, A., Schwendenwein, I., Stanclova, G., Dijk, G.V., et al., 2015. Energy expenditure and metabolic changes of free-flying migrating northern Bald Ibis. PLoS One 10, e0134433.
Bartoń, K., 2015. MuMIn: Multi-Model Inference. R Package Version 1.40.4. Available at: .
Collop, C., Stillman, R.A., Garbutt, A., Yates, M.G., Rispin, E., Yates, T., 2016. Variability in the area, energy and time costs of wintering waders responding to disturbance. Ibis 158, 711-725.
Efrat, R., Harel, R., Alexandrou, O., Catsadorakis, G., Nathan, R., 2019. Seasonal differences in energy expenditure, flight characteristics and spatial utilization of Dalmatian Pelicans Pelecanus crispus in Greece. Ibis 161, 415-427.
Elliott, K.H., Davoren, G.K., Gaston, A.J., 2008a. Increasing energy expenditure for a deep-diving bird alters time allocation during the dive cycle. Anim. Behav. 75, 1311-1317.
Elliott, K.H., Davoren, G.K., Gaston, A.J., 2008b. Time allocation by a deep-diving bird reflects prey type and energy gain. Anim. Behav. 75, 1301-1310.
Engelhardt, K.A.M., Ritchie, M.E., Powell, J.A., 2014. Body size mediated coexistence in swans. Sci. World J. 2014, 643694.
Fattorini, N., Brunetti, C., Baruzzi, C., Chiatante, G., Lovari, S., Ferretti, F., 2019. Temporal variation in foraging activity and grouping patterns in a mountain-dwelling herbivore: environmental and endogenous drivers. Behav. Process. 167, 103909.
Fletcher, Q.E., Speakman, J.R., Boutin, S., Lane, J.E., Mcadam, A.G., Gorrell, J.C., et al., 2014. Daily energy expenditure during lactation is strongly selected in a free-living mammal. Funct. Ecol. 29, 195-208.
Forslind, S., Blokhuis, H.J., Riber, A.B., 2021. Disturbance of resting behaviour of broilers under different environmental conditions. Appl. Anim. Behav. Sci. 242, 105425.
Goldstein, D.L., 1988. Estimates of daily energy expenditure in birds: the time-energy budget as an integrator of laboratory and field studies. Am. Zool. 28, 829-844.
Gyimesi, A., Franken, M.S., Feige, N., Nolet, B.A., 2012. Human disturbance of Bewick's Swans is reflected in giving-up net energy intake rate, but not in giving-up food density. Ibis 154, 781-790.
He, F., Wu, Q., Zhu, J., Su, L., Zou, H., 2016. The effect of measurement method on recorded water depths used by wetland Grallatores. Chin. J. Wildlife 37, 346-350.
Ito, H., 2019. Risk sensitivity of a forager with limited energy reserves in stochastic environments. Ecol. Res. 34, 9-17.
Jones, O.E., Williams, C.K., Castelli, P.M., 2014. A 24-hour time-energy budget for wintering American Black Ducks (Anas rubripes) and its comparison to allometric estimations. Waterbirds 37, 264-273.
Li, C., Zhou, L., Xu, L., Zhao, N., Beauchamp, G., 2015. Vigilance and activity time-budget adjustments of wintering Hooded Cranes Grus monacha in human-dominated foraging habitats. PLoS One 10, e0118928.
Magella, G., Samantha, E.R., Steven, J.P., Patrick, J.B., 2012. Behavioural compensation reduces energy expenditure during migration hyperphagia in a large bird. Funct. Ecol. 26, 876-883.
Montgomery, M., Baumgardt, B., 1965. Regulation of food intake in ruminants. 1. Pelleted rations varying in energy concentration. J. Dairy Sci. 48, 569-574.
Navedo, J.G., Verdugo, C., Rodríguez-jorquera, I.A., Abad-gómez, J.M., Suazo, C.G., Castañeda, L.E., et al., 2019. Assessing the effects of human activities on the foraging opportunities of migratory shorebirds in Austral high-latitude bays. PLoS One 14, e0212441.
Nolet, B.A., Kölzsch, A., Elderenbosch, M., Noordwijk, A.J., 2016. Scaring waterfowl as a management tool: how much more do geese forage after disturbance? J. Appl. Ecol. 53, 1413-1421.
Nolet, B.A., Langevoord, O., Bevan, R.M., Engelaar, K.R., Klaassen, M., Mulder, R.J.W., et al., 2001. Spatial variation in tuber depletion by swans explained by differences in net intake rates. Ecology 82, 1655-1667.
Peace, A., Wang, H., 2019. Compensatory foraging in stoichiometric producer-grazer models. Bull. Math. Biol. 81, 4932-4950.
Pelletier, D., Guillemette, M., Grandbois, J., Butler, P.J., 2008. To fly or not to fly: high flight costs in a large sea duck do not imply an expensive lifestyle. Proc. R. Soc. B 275, 2117-2124.
Rizzolo, J.D., Schmutz, A.J., Speakman, R.J., 2015. Fast and efficient: postnatal growth and energy expenditure in an Arctic-breeding waterbird, the Red-throated Loon (Gavia stellata). Auk 132, 657-670.
Roby, D.D., Ricklefs, R.E., 2015. Energy expenditure in adult least Auklets and diving Petrels during the chick-rearing period. Physiol. Biochem. Zool. 59, 661-678.
Schummer, M.L., Eddleman, W.R., 2003. Effects of disturbance on activity and energy budgets of migrating waterbirds in South-Central Oklahoma. J. Wildl. Manag. 67, 789-795.
Shankar, A., Graham, C.H., Canepa, J.R., Wethington, S.M., Powers, D.R., 2019. Hummingbirds budget energy flexibly in response to changing resources. Funct. Ecol. 33, 1904-1916.
Symonds, M.R.E., Moussalli, A., 2011. A brief guide to model selection, multimodel inference and model averaging in behavioural ecology using Akaike's information criterion. Behav. Ecol. Sociobiol. 65, 13-21.
Wan, W., Zhou, L., Song, Y., 2016. Shifts in foraging behavior of wintering Hooded Cranes (Grus monacha) in three different habitats at Shengjin Lake, China. Avian Res. 7, 13.
Wei, Z., Zheng, M., Zhou, L., Xu, W., 2020. Flexible foraging response of wintering Hooded Cranes (Grus monacha) to food availability in the lakes of the Yangtze River Floodplain, China. Animals 10, 568.
Weimerskirch, H., Martin, J., Clerquin, Y., Alexandre, P., Jiraskova, S., 2001. Energy saving in flight formation. Nature 413, 697-698.
Wood, K.A., Stillman, R.A., Wheeler, D., Groves, S., Hambly, C., Speakman, J.R., et al., 2013. Go with the flow: water velocity regulates herbivore foraging decisions in river catchments. Oikos 122, 1720-1729.
Wooley, J.B., Owen, R.B., 1977. Metabolic rates and heart rate-metabolism relationships in the Black Duck (Anas rubripes). Comp. Biochem. Physiol. A 57, 363-367.
Yu, C., Zhou, L., Mahtab, N., Fan, S., Song, Y., 2020. Microhabitat variables explain patch switching by wintering Bewick's Swans through giving-up net energy intake rates. Environ. Sci. Pollut. Res. Int. 27, 18843-18852.
Yu, C., Zhou, L., Song, Y., 2019a. The effects of human activities on diurnal energy expenditure of Tundra Swan Cygnus columbianus foraging in paddy field during the late wintering period. J. Lake Sci. 31: 195-201.
Yu, C., Zhou, L., Mahtab, N., Fan, S., Song, Y., 2019b. The influence of food density, flock size, and disturbance on the functional response of Bewick's Swans (Cygnus columbianus bewickii) in Wintering Habitats. Animals 9, 946.
Zhang, D., Zhou, L., Song, Y., 2015. Effect of water level fluctuations on temporal-spatial patterns of foraging activities by the wintering Hooded Crane (Grus monacha). Avian Res. 6, 16.
Zhao, J., Zhang, J., Zhao, Y., Zhang, Z., Godefroit, P., 2020. Shaking the wings and preening feathers with the beak help a bird to recover its ruffled feather vane. Mater. Design 187, 108410.
Zimmer, C., Boos, M., Bertrand, F., Robin, J., Petit, O., 2011. Behavioural adjustment in response to increased predation risk: a study in three duck species. PLoS One 6, e18977.
Table
1.
The allocation of time and energy expenditure of behaviors for the Spotted-billed Ducks in disturbed and normal habitats at Xin'an River Basin in 2021–2022.
Table
2.
Model selection tables-ranked generalized linear mixed models in the candidate set for the diurnal total energy expenditure of wintering Spot-billed Ducks at Xin'an River Basin.
Candidate models
df
AICc
Δi
ωi
acc ωi
Log10 (Diurnal total energy expenditure) = Water depth + River width + Relative water velocity
1 Water depth + River width
4
−94.69
0
0.52
0.52
2 Water depth + River width + Relative water velocity
5
−94.55
0.13
0.48
1
3 River width + relative water velocity
4
−83.46
11.22
0
1
4 Water depth
3
−73.42
21.27
0
1
5 Water depth + Relative water velocity
4
−72.32
22.37
0
1
6 River width
3
−71.96
22.73
0
1
7 Relative water velocity
3
−58.27
36.42
0
1
8 Null
2
−48.49
46.2
0
1
Δi, difference in AICc between model and top model; ωi, Aikaike model weight; acc ωi, cumulative model weight.
Table
3.
Parameter estimates, standard errors, and confidence intervals for the potential predictors of the diurnal total energy expenditure of wintering Spot-billed Ducks. Dependent variables have been Log10 transformed.
Predictor
Relative importance
Estimate
S.E.
95% confidence interval
Lower limit
Upper limit
Intercept
1.736
0.081
1.575
1.898
Water depth
1
0.067
0.016
0.034
0.099
River width
1
0.002
0
0.001
0.002
Relative water velocity
0.48
0.005
0.007
−0.004
0.025
Relative importance: sum of the Akaike weights for each variable across all the models where it occurred (see Materials and methods); S.E.: standard error.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds