
Figure
1.
Examples of combinations of experimental host nests used in this study.
| Citation: |
Longwu Wang, Gangbin He, Yuhan Zhang, Wei Liang, William E. Feeney. 2022: Experimental evidence that cuckoos preferentially parasitize host nests early in their laying cycle. Avian Research, 13(1): 100042. DOI: 10.1016/j.avrs.2022.100042

|
Brood parasites, such as the Common Cuckoo (Cuculus canorus), rely on manipulating hosts from other species to raise their offspring. Selection should favor individuals that are able to identify and choose host nests that closely match the incubation stage of their own eggs. While intuitive, and supported by the findings of numerous long-term monitoring studies, experimental evidence of this remains sparse. By using video monitoring, we used three experimental groups to explore whether Common Cuckoos preferentially choose host Oriental Reed Warbler (Acrocephalus orientalis) nests that are early in their laying cycle. We found that Common Cuckoos preferentially parasitize nests of the host Oriental Reed Warbler that are earlier in their laying (less vs. more eggs) or nesting cycle (eggs vs. chicks), and videos of our experiments provide direct insights into their decision making process. Our results provide strong experimental evidence that Common Cuckoos are able to assess nesting stage and strategically choose nests that increase the likelihood of successful parasitism.
Brood parasites, such as the Common Cuckoo (Cuculus canorus, hereafter 'cuckoo'), lay their eggs in the nests of other birds and impose the cost of parental care to the host (Davies, 2000; Feeney et al., 2014; Soler, 2014). Following incubation, cuckoo eggs typically hatch before the host's own eggs and the newly hatched cuckoo evicts the host's eggs and/or nestlings (Davies, 2000). A cuckoo chick's survival prospects are severely diminished if it is forced to cohabit and compete with the host's own chicks (Rutila et al., 2002; Grim et al., 2009). This highlights that accurate timing of egg deposition by the adult cuckoo and early hatching of the cuckoo's egg are important adaptations that underpin successful brood parasitism.
Cuckoo females usually parasitize in the early stage of egg-laying (0–3 eggs) (Moskát and Honza, 2002; Moskát et al., 2009; Honza et al., 2020; Wang et al., 2020a, b). Wyllie (1981) found that the majority of cuckoo eggs were laid during the egg laying period (92%, 83/90), and observed no cases of parasitism after incubation began. Similarly, parasitism of the Reed Warbler (Acrocephalus scirpaccus) shows that no parasitism occurred before or after laying, and all parasitic eggs (N = 45) were laid during the laying stage of the host (Davies and Brooke, 1988). It can be seen from the above studies that cuckoos prefer to parasitize in the early laying stage of the host, which may be beneficial for the hatching of parasitic eggs. Further field observations indicate that cuckoo eggs that were laid before the Great Reed Warbler (Acrocephalus arundinaceus) began to incubate hatch earlier than host eggs. In contrast, one-third of the cuckoo eggs that were laid after the host eggs began to hatch did not have this advantage (Geltsch et al., 2016). In the Great Reed Warbler, the fourth egg is considered as a dividing line such that cuckoo eggs laid prior to it being laid often hatch, while those laid after are less likely to be successful (Geltsch et al., 2016). Therefore, for the cuckoo, being able to sense the number of eggs in the host's nest, or being able to know the host's egg-laying stage (early or late), plays a key role in the success of the parasitism.
Selection should favor brood parasites that identify and lay in host nests that maximize their likelihood of successful parasitism. Indeed, it is well-established that brood parasites choose nests based off a variety of cues, such as nest location (Øien et al., 1996), host behavior (Banks and Martin, 2001), host group size (Feeney et al., 2013), and the degree to which the parasite's eggs resemble those of the host (Yang et al., 2016, 2017; Ma and Liang, 2021). Likewise, the results of numerous field-based monitoring efforts near unanimously report that brood parasites preferentially lay in nests during the early-stages of the host's laying cycle (e.g. Mermoz and Reboreda, 1999; Moskát et al., 2006; Honza et al., 2020) and that eggs laid too early or late tend to be unsuccessful (Langmore et al., 2003). Interestingly, while laying synchrony between the parasite and host is key to successful parasitism, only one recent study (Jelínek et al., 2014) has provided observational data in support of this.
At our research site, cuckoos mainly use the Oriental Reed Warbler (Acrocephalus orientalis) as its primary host. Previous work found that cuckoo females can always parasitize at the appropriate time (1–3 eggs in the nest) (Yang et al., 2016, 2017; Wang et al., 2020a, b, 2021), so we speculate that the cuckoo can evaluate the situation of the target nest, including whether it contains eggs or nestlings, which suggests that they know the stage of the nest (early or late), and finally choose the right time for parasitism. In this study, to verify our speculation, we experimentally investigated whether Common Cuckoos preferentially target Oriental Reed Warbler nests at an earlier nesting stage, as well as whether they preferentially target nests earlier in the egg-laying cycle. As our previous work indicates that female cuckoos preferentially target hosts early in their egg-laying cycle (see Wang et al., 2020a, b), we set comparison of nests with chicks compared to nests with eggs and hypothesized that cuckoos preferentially target nests that contain eggs rather than chicks, and that they also target nests with fewer eggs than more eggs as this suggests that they are earlier in the egg-laying cycle.
This study was performed in Zhalong National Nature Reserve (46°48′–47°31′ N, 123°51′–124°37′ E) located in Heilongjiang Province, Northeast China. Field experiments were carried out during the breeding season (June to August) between 2017–2020. At our study site, habitats are primarily reed swamps, and to our knowledge, the Common Cuckoo only parasitizes the Oriental Reed Warbler (clutch size: 4–6 eggs) (Wang et al., 2020a; b, 2021). Our previous work indicates that parasitism rates are high, ranging from 34.3% to 65.5% per year (Yang et al., 2016, 2017).
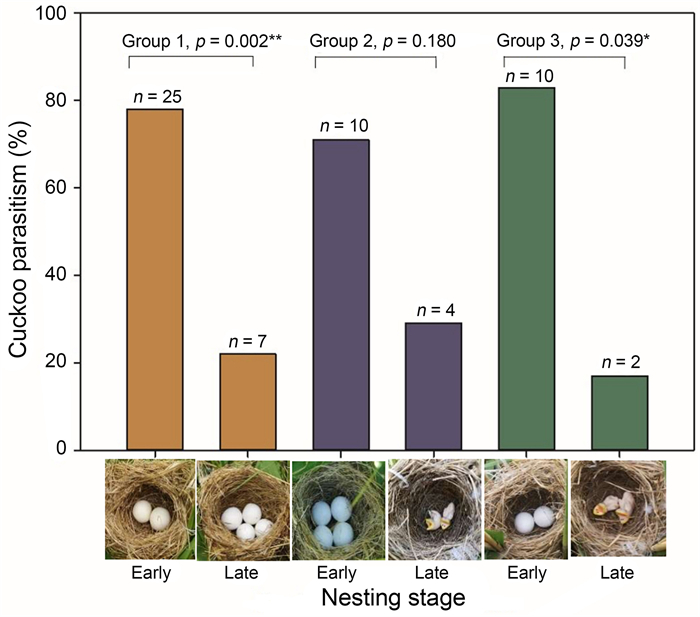
We systematically searched for warbler nests in the reed beds by monitoring the activities of adult birds during the breeding season. Nests were monitored daily to ascertain the date that the first egg was laid, clutch size, parasitism status and nest fate. Once a warbler laid the first egg in their nest, two additional artificial nests were set up approximately 1 m away from the host nest and assigned to one of three experimental groups: 1) two and four model eggs placed in the experimental nests (Fig. 1; group 1; N = 55); 2) four model eggs and two model nestlings (Fig. 1; group 2; N = 30); and 3) two model eggs and two model nestlings (Fig. 1; group 3; N = 23). Experimental nests were old warbler nests that we collected in the field during the previous year after breeding had finished, and we combined them using twine (as per Wang et al., 2020a). During experiments these nests were placed nearby host nests (Fig. 1) and slightly higher than the host's nest, as cuckoos preferentially target more visible nests (Patten et al., 2011). We camouflaged the camera with reed leaves and wrapped it in plastic to avoid rain damage. Model eggs (length = 24.18 ± 0.50 mm; width = 21.56 ± 0.63 mm; mass = 7.64 ± 0.13 g) and chicks (length = 20.93 ± 0.42 mm; width = 16.01 ± 0.35 mm; mass = 5.86 ± 0.17 g) were made of a white synthetic clay (Fig. 1). Experimental nests were monitored daily for six days to record whether a parasitism event occurred. All nests were also video recorded from dawn until dusk each day during the experiment. We used mini digital cameras (Uniscom-T71, 70 mm × 26 mm × 12 mm; Mymahdi Technology Co. Ltd., Shenzhen, China) to capture cuckoo behaviour at host nests. To achieve longer battery life, the camcorders were equipped with external power supply (20,000 mAh, Romoss-Sense 6; Romoss Technology Co. Ltd., Shenzhen, China).
During experiments, all parasitism events occurred during the host's laying period, we only observed parasitism of the experimental nests, and no hosts abandoned their nests during the experiments. Three example video recordings showing cuckoo laying behavior in group 1 (Appendix Video S1), group 2 (Appendix Video S2), and group 3 (Appendix Video S3), are provided as supplementary files.
Supplementary video related to this article can be found at https://doi.org/10.1016/j.avrs.2022.100042
Binomial tests (two-tailed test) were used to test for the choice of nests by cuckoo females. For all experimental nests, if one parasitism event at a particular nest was recorded, the cuckoo egg was removed, and the original experimental status was re-established to ensure that all combinations of nests were parasitized with similar probability. Difference was considered to be significant at p < 0.05, highly significant at p < 0.01, and not significant at p > 0.05. Statistical analysis was conducted using IBM SPSS Version 25.0.
For group 1, 55 pairs of nests were set up, 32 of which were parasitized (58%). Cuckoos were significantly more likely to parasitize the nests containing 2 eggs (25 of 32, 78%) compared to 4 eggs (7 of 32, 22%) (Binomial test, p = 0.002; Fig. 2; group 1). For group 2, 30 nests were set up, 14 of which were parasitized (47%). We found that cuckoos were not more likely to parasitize nests containing 4 eggs (10 of 14, 71%) compared to 2 nestlings (4 of 14, 29%) (Binomial test, p = 0.180; Fig. 2; group 2). For group 3, 23 nests were set up, 12 of which were parasitized (52%). Cuckoos were significantly more likely to parasitize the nests containing 2 eggs (10 of 12, 83%) compared to 2 nestlings (2 of 12, 17%) (Binomial test, p = 0.039; Fig. 2; group 3).
Our results show that Common Cuckoo females preferentially parasitized nests with incomplete clutches of eggs rather than nestlings or complete clutches. This result suggests that cuckoos are able to evaluate which nests are more suitable for parasitism based on nesting stage and the number of eggs within the host nest. While observational data near unanimously indicate that brood parasites target nests that maximize incubation synchrony between their own and host's eggs (e.g., Moskát et al., 2006; Honza et al., 2020), to our knowledge these data comprise the first experimental evidence that cuckoos show preferences for nests at an earlier nesting stage and earlier in the laying cycle.
Videos of our experiments provide direct insights into the decision-making process that cuckoos face when choosing a nest to lay in. For instance, when posed with a choice between nests containing either two or four eggs, we found that one cuckoo initially landed on the rim of the nest with four eggs and after approximately 10 s of inspecting the nest contents, moved across to the nest with two eggs before laying (Appendix Video S1). In another instance, the cuckoo initially approached the nest containing two nestlings, but moved across to the nest with two eggs before laying (Appendix Video S3). Although specifically selected, these videos appear to provide direct evidence of the cuckoos assessing nest contents prior to laying their egg, preferring nests with eggs over chicks and nests with fewer eggs compared to more eggs in a manner that would maximize their likelihood of synchronizing the incubation of the own egg with that of their host. For the model nestling experimental treatment, the results were a little not satisfying our expectations in group 2, which may be caused by fewer samples in the fieldwork.
Overall, our study provides experimental results supporting the widely held presumption that cuckoos, like other brood parasites, strategically choose nests to maximize synchrony with their hosts, and highlights the key role that information and strategic decisions play in underpinning successful brood parasitism.
WL and LW designed the study, LW, GH and YZ carried out field experiments, LW and WEF wrote the draft manuscript, and WL involved in discussion and improved the manuscript. All authors read and approved the final manuscript.
The experiments comply with the current laws of China. Fieldwork was performed with permission (No. ZL-GZNU-2019-06) from Zhalong National Nature Reserve, Heilongjiang, China. Experimental procedures were in accordance with the Animal Research Ethics Committee of Guizhou Normal University (No. GZNUECEE-2022-001).
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
We are grateful to Wenfeng Wang and Jianhua Ma for their assistance with fieldwork. We also thank the Zhalong National Nature Reserve for their support and permission to carry out this study. This work was supported by the National Natural Science Foundation of China (Nos. 31960105 to LW, and 31970427 to WL) and the specific research fund of the Innovation Platform for Academicians of Hainan Province. LW was funded by the Guizhou Natural Science Foundation (No. ZK[2022]-316).
|
Davies, N.B., 2000. Cuckoos, Cowbirds and Other Cheats. T & AD Poyser, London.
|
|
Wyllie, I., 1981. The Cuckoo. Batsford, London.
|
Figures(2)





 DownLoad:
DownLoad:
 Email Alerts
Email Alerts RSS Feeds
RSS Feeds