Jorge Garrido-Bautista, Antonio Soria, Cristina E. Trenzado, Amalia Pérez-Jiménez, Eliana Pintus, José Luis Ros-Santaella, Nicola Bernardo, Mar Comas, Stanislav Kolenčík, Gregorio Moreno-Rueda. 2022: Within-brood body size and immunological differences in Blue Tit (Cyanistes caeruleus) nestlings relative to ectoparasitism. Avian Research, 13(1): 100038. DOI: 10.1016/j.avrs.2022.100038
Citation:
Jorge Garrido-Bautista, Antonio Soria, Cristina E. Trenzado, Amalia Pérez-Jiménez, Eliana Pintus, José Luis Ros-Santaella, Nicola Bernardo, Mar Comas, Stanislav Kolenčík, Gregorio Moreno-Rueda. 2022: Within-brood body size and immunological differences in Blue Tit (Cyanistes caeruleus) nestlings relative to ectoparasitism. Avian Research, 13(1): 100038. DOI: 10.1016/j.avrs.2022.100038
Jorge Garrido-Bautista, Antonio Soria, Cristina E. Trenzado, Amalia Pérez-Jiménez, Eliana Pintus, José Luis Ros-Santaella, Nicola Bernardo, Mar Comas, Stanislav Kolenčík, Gregorio Moreno-Rueda. 2022: Within-brood body size and immunological differences in Blue Tit (Cyanistes caeruleus) nestlings relative to ectoparasitism. Avian Research, 13(1): 100038. DOI: 10.1016/j.avrs.2022.100038
Citation:
Jorge Garrido-Bautista, Antonio Soria, Cristina E. Trenzado, Amalia Pérez-Jiménez, Eliana Pintus, José Luis Ros-Santaella, Nicola Bernardo, Mar Comas, Stanislav Kolenčík, Gregorio Moreno-Rueda. 2022: Within-brood body size and immunological differences in Blue Tit (Cyanistes caeruleus) nestlings relative to ectoparasitism. Avian Research, 13(1): 100038. DOI: 10.1016/j.avrs.2022.100038
Departamento de Zoología, Facultad de Ciencias, Universidad de Granada, 18071, Granada, Spain
b.
Departamento de Biología Celular, Facultad de Ciencias, Universidad de Granada, 18071, Granada, Spain
c.
Department of Veterinary Sciences, Faculty of Agrobiology, Food and Natural Resources, Czech University of Life Sciences Prague, Kamýcká 129, 16500, Prague 6, Suchdol, Czech Republic
d.
Estación Biológica de Doñana, EBD-CSIC, Av. Américo Vespucio 26, 41092, Seville, Spain
e.
Department of Biological Sciences, Dartmouth College, NH, 03755, Hanover, USA
f.
Department of Biology, University of Nevada, NV, 89557, Reno, USA
Several ectoparasites parasitise nestlings decreasing their body condition, growth and survival. To minimise any loss of fitness due to ectoparasites, birds have developed a wide variety of defence mechanisms, potentially including hatching asynchrony. According to the Tasty Chick Hypothesis (TCH), the cost of parasitism would be reduced if ectoparasites tend to eat on less immunocompetent nestlings, typically the last-hatched chick in asynchronously hatched broods, as they are in poor body condition. Two predictions of the TCH are that immune capacity is lower in smaller nestlings than in larger ones and that parasites should provoke a more negative effect on smaller nestlings. Here, we test these predictions in a population of Blue Tits (Cyanistes caeruleus) whose broods are parasitised by Hen Fleas (Ceratophyllus gallinae) and Blowflies (Protocalliphora azurea). We recorded the presence of both ectoparasites and analysed the immunocompetence (number of leucocytes per 10,000 erythrocytes and cutaneous immune response to phytohaemagglutinin) and body condition of smaller and larger nestlings within individual broods. The leucocyte count was higher in smaller nestlings than in larger ones, whereas the cutaneous immune response did not differ between smaller and larger nestlings. Smaller nestlings, but not larger nestlings, had lower body mass when fleas were present. Blowflies, by contrast, had no detectable negative effect on nestlings. Overall, our findings provide partial support to the TCH. Lower immune capacity in smaller nestlings than in larger ones was not supported, but Hen Fleas seemed to negatively impact on smaller nestlings more than on larger ones.
In altricial birds, nestlings often establish a body size hierarchy, mainly due to hatching asynchrony (Clark and Wilson, 1981). Size hierarchy implies the nestlings have different reproductive values for parents, so individual broods contain both core high-value and marginal low-value nestlings (Forbes et al., 1997). This means food is distributed unequally among the nestlings, with small ones usually being underfed (Dickens and Hartley, 2007; Moreno-Rueda et al., 2009; García-Navas et al., 2014). Several hypotheses have been proposed to explain the adaptive value of hatching asynchrony, mainly in relation to resource availability (Magrath, 1990; Stenning, 1996). An example is the Tasty Chick Hypothesis (TCH), which suggests that hatching asynchrony may have evolved as an anti-ectoparasite defence (Christe et al., 1998). The TCH proposes that ectoparasites enhance their fitness by aggregating on the less immunocompetent nestling, which is presumed to be the last chick hatched in asynchronous broods and also the one with the poorest body condition due to a low nutritional status. Hence, the smallest nestling in a brood would be the most attractive (tasty) to parasites. This would have the concomitant effect of reducing the parasitic load for the remaining nestlings, which are of higher reproductive value for parents. According to the TCH, parental fitness would eventually increase with more healthy core nestlings surviving at the expense of sacrificing the highly parasitised last-hatched chick, the "tasty" chick (Christe et al., 1998). In sum, the TCH predicts that: (1) smaller nestlings are less immunocompetent against ectoparasites than larger nestlings; (2) parasites feed mainly on smaller nestlings; and (3) smaller nestlings are more negatively affected by ectoparasites than larger nestlings.
The aim of the present study was to test the aforementioned predictions 1 and 3 of the TCH in a wild population of Blue Tits parasitised by Blowfly larvae (Protocalliphora azurea) and Hen Fleas (Ceratophyllus gallinae). We did not test prediction 2 (parasites aggregate on the last-hatched nestling) given that these parasites inhabit inside the nest material, feeding on nestlings usually at night. According to the TCH, we predict that smaller nestlings in the same brood will present lower immunocompetence than larger nestlings (prediction 1). Moreover, if the TCH is applicable in this species, given that ectoparasites would feed mainly on the smaller nestlings, these nestlings will suffer a higher impact from ectoparasitism than larger nestlings. Consequently, within-brood variation in body mass, tarsus length and body condition are expected to be greater in infested compared to uninfested nests (prediction 3). Specifically, we compared body mass, tarsus length, body condition and immunocompetence (measured as relative leucocyte concentration and immune response to phytohaemagglutinin (PHA)) between the smallest and largest nestlings within a brood according to the prevalence of nest ectoparasites.
2.
Material and methods
2.1
Study area and bird species
Blue Tits are Palearctic forest insectivorous birds that nest in secondary cavities in trees, where they typically lay an average of six to nine eggs per clutch (seven in our population) in the Iberian Peninsula (Salvador, 2016). Eggs generally hatch asynchronously (hatching spread of two days; Slagsvold et al., 1995; Stenning, 2008), leading to within-brood size hierarchy (Stenning, 2018). During the breeding season, nests are commonly infested by different ectoparasites such as Blowflies, fleas and mites, which usually decreases nestling health, condition and survival (e.g. Hurtrez-Boussès et al., 1997; Tomás et al., 2007; Castaño-Vázquez et al., 2018).
In 2017, we studied a wild population of Blue Tits inhabiting two contiguous forests, located at 1700–1800 m a.s.l. in the Sierra Nevada National Park (southeastern Spain; 36°57′ N, 3°24′ W). The two study areas comprised of (1) a dry forest composed of Holm Oaks (Quercus ilex) and Pyrenean Oaks (Quercus pyrenaica), and (2) a humid forest characterised by Scots Pines (Pinus sylvestris), Holm Oaks and Pyrenean Oaks. These two forests differed in several environmental and biotic factors, such as mean temperature, humidity, irradiation, insolation time, canopy cover and parasite load (Appendix 1; also see Garrido-Bautista et al., 2021). The higher humidity of the humid forest was due to the presence of a river and a stream, and, to simplify, we referred to these two areas as humid and dry forests throughout the text. Blue Tits bred in nest boxes (ICONA C model; base area: 196 cm2; height: 20 cm; hole diameter: 3 cm; material: wood with the outer layer of plastic paint; Moreno-Rueda, 2003) hung from branches using metal hooks. They were inspected regularly to determine laying date, hatching date and brood size. Nest boxes were cleaned and their contents removed at the end of the breeding season to avoid the accumulation of ectoparasites between breeding seasons.
2.2
Morphometric measurements and leucocyte count
We considered the hatching day as the day 0. When the nestlings were 13 days old, the age at which body mass and tarsus length reach asymptotic growth (Björklund, 1996), nestlings from 37 nests (20 from the dry forest and 17 from the humid forest) were weighed with a portable, digital scale (accuracy, 0.1 g) and their tarsus length measured to the nearest 0.01 mm with a digital calliper. We calculated the body condition index (BCI) of nestlings as the residuals of regressing log body mass on log tarsus length. In 19 nests (dry forest, n = 11; humid forest, n = 8), we also took a 100 μL blood sample from the largest and smallest nestlings (~1% of nestling body mass, which was about 10 g) from the jugular vein using heparinised insulin syringes in sterile conditions (following Owen, 2011). Blood sampling has been shown to have negligible effects on tit nestling survival (Schmoll et al., 2004). Handling time was kept to a minimum to reduce nestling stress (de Jong, 2019). A drop of blood was smeared on a slide and air-dried following Owen (2011). Blood samples were fixed in absolute methanol for 2 min and stained with a Wright-Giemsa combination stain as follows: (1) Wright stain for 2 min; (2) distilled water for 2 min followed by tap water for a few seconds; (3) Giemsa stain 1:9 diluted in phosphate-buffered saline, pH 7.2, for 10 min followed by tap water for a few seconds; and (4) 0.5% acetic acid for 1 s. Smears were prepared with Eukitt® mounting medium. Samples were viewed with a Zeiss Axiophot microscope at 400× magnification and 35–40 fields per blood smear were observed (always by the same researcher, AS). Fields were photographed with a Zeiss Axiocam camera connected to the microscope. Leucocytes were counted and identified following Campbell and Ellis (2007) and erythrocytes were counted with Mizutama software (Ochoa et al., 2019). The relative leucocyte concentration for each nestling was estimated as the number of leucocytes per 10,000 erythrocytes.
2.3
Cutaneous immune response to PHA
We also measured the immune response to PHA in larger and smaller nestlings from 28 broods (dry forest, n = 15; humid forest, n = 13). When the nestlings were 12 days old, we inoculated 0.1 mg of phytohaemagglutinin (PHA-P; Sigma Aldrich, L-8754) diluted in 0.02 mL of isotonic phosphate-buffered saline in their left wing web (following Smits et al., 1999). PHA is a mitogen that provokes an inflammatory immune response involving different types of cell and can be considered a multifaceted index of cutaneous immune activity (Martin et al., 2006). The inflammatory response to PHA correlates positively with nestling survival to adulthood in Blue Tits (Cichoń and Dubiec, 2005). Prior to inoculation of PHA, we measured the wing web thickness three times with a pressure-sensitive micrometre (Mitutoyo Inc.; accuracy, 0.01 mm) and took the average. The following day, when nestlings were 13 days old, we measured the wing web thickness again following with the same procedure, calculating the inflammatory immune response as the difference between both measurements. All measurements were made by the same researcher (GMR). Some of the nestlings (n = 20) that were inoculated with PHA were measured at the next day for leucocyte count. Although the inoculation of PHA in the wing web of nestlings causes local and cutaneous inflammation, an elevation of T-cells in the bloodstream can occur (Tella et al., 2008). To control for this possible effect, we compared the leucocyte count between those nestlings that were inoculated and not inoculated with PHA.
2.4
Ectoparasite abundance and prevalence
Once nestlings fledged and left the nest, we carefully sorted through nest material and recorded the presence and number of Blowflies (puparia and larvae) per nest, as well as the presence or absence of Hen Fleas (larvae or adults). Females of the Blowfly lay their eggs in the nests of cavity-nesting birds, so their larvae inhabit inside nest material and feed intermittently on nestlings (Bennett and Whitworth, 1991). The haematophagous activity of Blowfly larvae provokes several negative impacts on nestlings (Merino and Potti, 1995; Hurtrez-Boussès et al., 1997; Puchala, 2004). Tits (Paridae) are one of the hosts where the Hen Flea achieves its higher productivity (Tripet and Richner, 1997a). Fleas inhabit inside nest material; flea larvae are saprophyte, but adults take blood from nestlings (Tripet and Richner, 1999). The blood-sucking of adult Hen Fleas has been reported to cause detrimental effects on the growth, body condition, blood count and health of Blue Tit nestlings (Tripet and Richner, 1997b; Pitala et al., 2009; Brommer et al., 2011).
2.5
Statistical analyses
Graphical inspection (following Zuur et al., 2010) revealed a normal distribution for all recorded variables, except Blowfly abundance. The variation in Blowfly and flea prevalence in relation to each forest was tested using the chi-squared test, and the abundance of Blowflies between forests was tested with the Mann-Whitney U test. We calculated the within-brood differences in relative leucocyte count, cutaneous immune response, body mass, tarsus length and body condition between larger and smaller nestlings (values for larger nestling minus smaller ones).
The differences in the number of leucocytes between larger and smaller nestlings were initially tested using a paired Student's t-test (Quinn and Keough, 2002). We assessed how the relative leucocyte count and the cutaneous immune response varied with forest type and nestling size using two separate linear mixed-effects models of restricted maximum likelihood (REML-LMM) (Zuur et al., 2009), where nest identity was the random factor and nestling rank (larger versus smaller), forest type (humid versus dry) and its interaction were the predictors. Brood size and laying date were included as covariates, but their effects were not significant and so they were removed from the final model. A REML-LMM was also used to test the relationship between body mass and leucocyte count. In this case, nest identity was the random factor and body mass and tarsus length were the covariates. We tested whether the differences in relative leucocyte count were related to Blowfly abundance using Spearman's rank correlation. We used a t-test to analyse the effect of PHA (nestlings inoculated and non-inoculated with PHA) on relative leucocyte count. We ran two separate linear models to assess how the within-brood differences in relative leucocyte count and cutaneous immune response varied with nest parasitisation by Hen Fleas and Blowflies. These models included forest type, prevalence of fleas and prevalence of Blowflies as predictors and nest identity as random factor.
We used t-tests to analyse the effect of nest parasitisation by Blowflies and fleas (parasitised or nonparasitised) on body mass, tarsus length and BCI and its within-brood differences. REML-LMM were used to test whether the observed ectoparasite-dependent within-brood differences in body mass, tarsus length and BCI were robust when controlled for forest, laying date and brood size, variables which could potentially affect any within-brood variation. In these cases, within-brood differences in the three morphological characters were the dependent variables, and flea or Blowfly prevalence, forest type, brood size and laying date were the predictors. The correlations between body mass of larger and smaller nestlings in nests parasitised and nonparasitised by fleas were established using the Pearson product-moment correlation. The basic statistics are given as mean ± SE (standard error). We used the "nlme" package (Pinheiro et al., 2019) in the software R (R Development Core Team, 2020). The data are free available (Appendix 2).
3.
Results
3.1
Immunological parameters
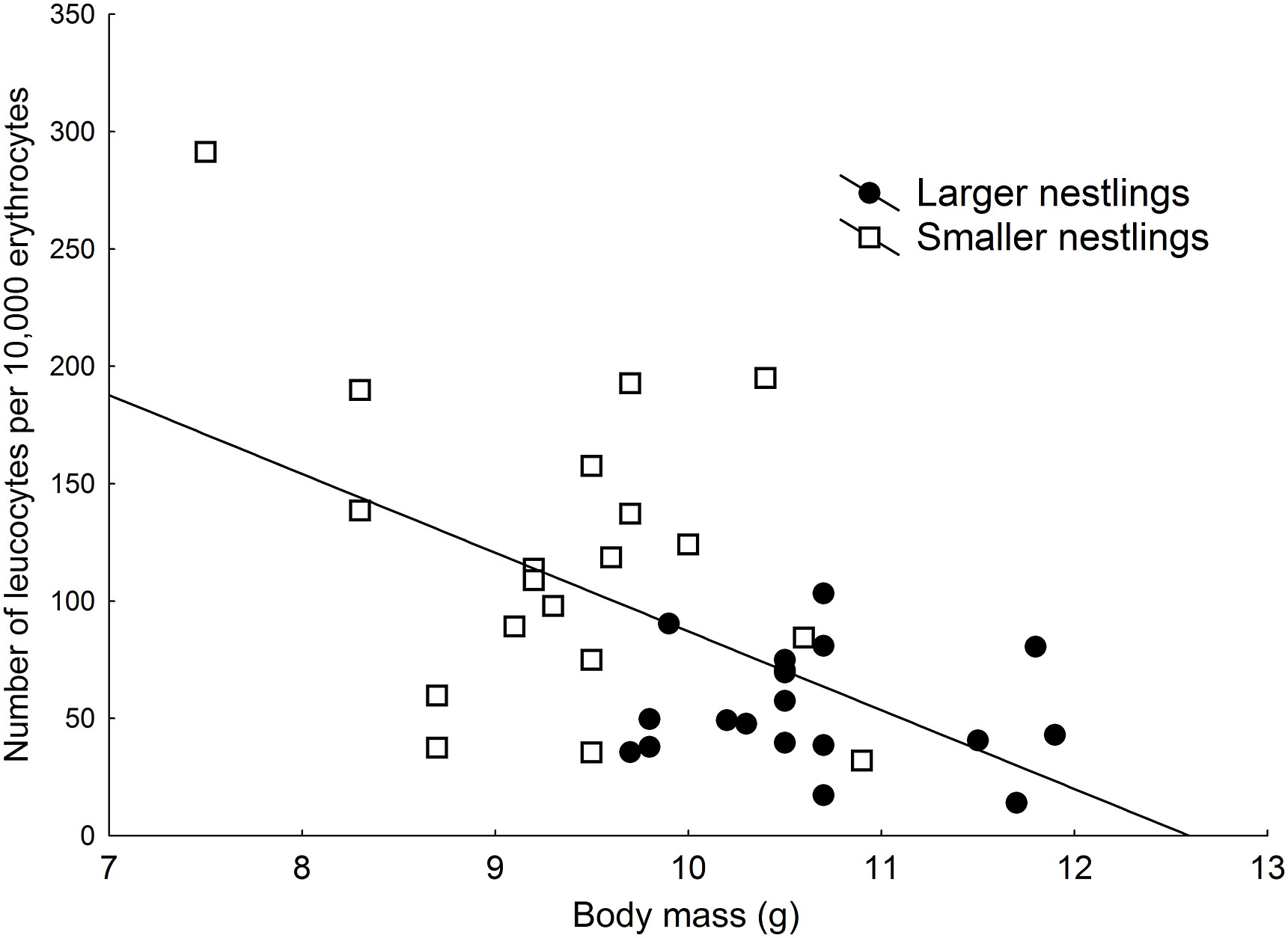
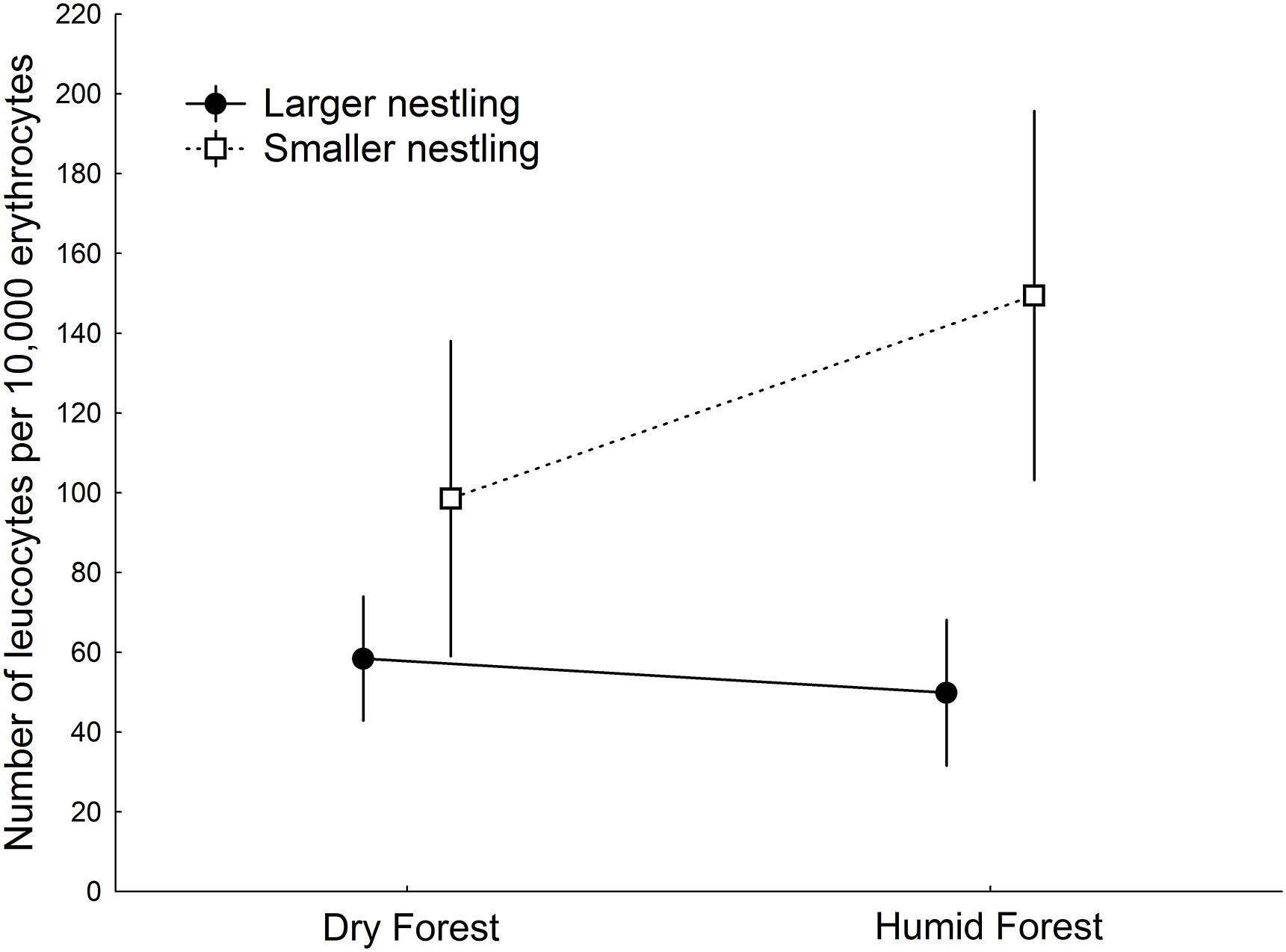
Blowfly prevalence and abundance were higher in the humid forest (14 out 17 nests infested, abundance: 10.82 ± 2.20 Blowflies) than in the dry forest (7 out of 20 nests infested, abundance: 2.45 ± 1.70 Blowflies; respectively, χ2 = 8.40, p = 0.0038, Mann-Whitney U test, z = 2.85, p = 0.003), but flea prevalence did not differ between forests (humid: 7 out of 17 nests, dry: 5 out of 20 nests; χ2 = 1.10, p = 0.29). Larger nestlings had half as many leucocytes (54.85 ± 5.54 leucocytes per 10,000 erythrocytes) as smaller nestlings (119.98 ± 15.05; t18 = −4.75, p < 0.001; Table 1). In fact, the number of leucocytes per 10,000 erythrocytes in a nestling was negatively affected by body mass (χ2 = 12.87, p < 0.001; Fig. 1). This within-brood difference was higher in the humid forest than in the dry forest (Table 1; Fig. 2). However, the within-brood difference in relative leucocyte count between larger and smaller nestlings did not vary with Blowfly abundance or prevalence (abundance: rs = 0.22, p = 0.36; Table 1) or flea prevalence (Table 1). Leucocyte count was not affected by PHA inoculation, as inoculated nestlings (n = 20) did not statistically differ in leucocyte count to non-inoculated nestlings (n = 18 nests; for larger nestlings: t17 = 0.49, p = 0.62; smaller: t17 = 0.27, p = 0.79). The cutaneous immune response did not differ significantly between larger (0.47 ± 0.04 mm) and smaller nestlings (0.46 ± 0.04 mm; Table 1). The presence of Blowflies or fleas did not affect the within-brood difference in immune response (Table 1).
Table
1.
Results of the linear mixed-effects models and linear models for the number of leucocytes, cutaneous immune response, and its within-brood differences (values for larger nestling minus smaller ones) of Blue Tit nestlings.
Variable
Chi-square (df = 1)
p-value
Number of leucocytes
Forest type
5.39
0.02
Nestling rank
6.27
0.012
Forest type*Nestling rank
5.81
0.02
Cutaneous immune response
Forest type
0.57
0.45
Nestling rank
0.19
0.66
Forest type*Nestling rank
0.24
0.62
Within-brood differences
Variable
F-value (df = 1)
P-value
Within-brood difference in leucocytes
Forest type
6.36
0.02
Flea prevalence
0.04
0.84
Blowfly prevalence
1.20
0.29
Within-brood difference in immune response
Forest type
0.59
0.45
Flea prevalence
1.17
0.29
Blowfly prevalence
0.51
0.48
Significant predictors (p < 0.05) are marked in bold.
Figure
1.
Relationship between body mass and relative leucocyte count in Blue Tit nestlings. Larger nestlings: filled circles; smaller nestlings: squares.
Figure
2.
Number of leucocytes per 10,000 erythrocytes in larger (filled circles) and smaller Blue Tit nestlings (squares) according to forest type. Bars represent the 95% confidence interval.
In flea-infested broods, smaller nestlings tended to have a lower body mass and shorter tarsus than uninfested broods, while larger nestling body mass and tarsus length did not differ with flea prevalence (Table 2). The presence of fleas also affected the within-brood difference in body mass, being higher in parasitised nests (Table 2). The effect of flea prevalence on within-brood differences in body mass remained statistically significant in a linear model when controlling for forest type, laying date, and brood size (F1, 31 = 13.09; p = 0.001; the remaining effects were not significant). In non-parasitised broods, there was a strong correlation between the body mass of larger and smaller nestlings (r = 0.93, p < 0.001); however, in broods parasitised by fleas, the body mass of larger and smaller nestlings did not present a significant correlation (r = 0.45, p = 0.15). Flea infestation was unrelated to nestling BCI (Table 2; always p > 0.15). However, within-brood differences in BCI were higher in the humid forest (0.09 ± 0.013) than in the dry forest (0.04 ± 0.013; t35 = 2.60, p = 0.014). This difference remained significant when controlling for laying date and brood size (F1, 33 = 7.02, p = 0.012; the remaining effects were not significant). Blowfly prevalence held no relation to within-brood differences in tarsus length, body mass or BCI (data not shown for simplicity).
Table
2.
Body mass, tarsus length and body condition index (BCI) of smaller and larger Blue Tit nestlings and the within-brood differences between Hen Flea-infested and uninfested broods.
Empty Cell
Infested broods (n = 12)
Uninfested broods (n = 25)
t-value
p-value
Larger nestlings
Tarsus length (mm)
16.63 ± 0.50
16.79 ± 0.52
−0.92
0.36
Body mass (g)
10.65 ± 0.58
10.62 ± 0.74
0.11
0.92
BCI (residuals)
0.045 ± 0.041
0.027 ± 0.087
0.67
0.51
Smaller nestlings
Tarsus length (mm)
15.76 ± 0.47
16.19 ± 0.45
−2.70
0.01
Body mass (g)
9.06 ± 0.71
9.54 ± 0.80
−1.78
0.08
BCI (residuals)
−0.042 ± 0.074
−0.029 ± 0.087
−0.44
0.67
Within-brood difference
Tarsus length (mm)
0.87 ± 0.62
0.60 ± 0.65
1.17
0.25
Body mass (g)
1.59 ± 0.68
1.08 ± 0.30
3.18
0.003
BCI (residuals)
0.087 ± 0.062
0.056 ± 0.058
1.46
0.15
Differences were tested using t-tests. Mean values are given with the SD.
In the present study conducted in a wild population of Blue Tits parasitised by Blowflies and fleas, we tested two predictions of the Tasty Chick Hypothesis (TCH): whether smaller nestlings within a brood have lower immunocompetence than larger nestlings (Christe et al., 1998) and whether ectoparasites had a greater negative effect on body mass and body size in smaller nestlings compared to larger nestlings (Szép and Møller, 2000). Our results lend some support to the TCH, as the presence of Hen Fleas in broods was detrimental for smaller Blue Tit nestlings (negatively affecting their body mass) but did not affect larger nestlings. However, there was no evidence that smaller nestlings are less immunocompetent than larger nestlings: the relative leucocyte count was higher in smaller than in larger nestlings and the immune response to a novel substance, phytohaemagglutinin, did not differ between larger and smaller nestlings. Moreover, the presence of fleas did not influence either of the immune response markers we measured. We did not find any evidence that Blowflies affected the body characteristics or immune status of smaller nestlings; so, this ectoparasite species does not appear to apply to the TCH (unlike the findings reported by Simon et al., 2003).
4.1
Evidence for the Tasty Chick Hypothesis
We found some evidence that the TCH might apply to the Blue Tit in our study population. Body mass and tarsus length of smaller nestlings were lower in the presence of fleas (the tarsus length, nonsignificantly), without affecting the body size and mass of larger nestlings. Consequently, within-brood differences in body mass were more marked in broods parasitised by fleas. Similarly, other studies have reported increased within-brood variance in infested nests (Merino and Potti, 1995; Christe et al., 1996; Szép and Møller, 2000; but no such effect was observed by Descamps et al., 2002; Hannam, 2006). Several studies have shown that nest-dwelling ectoparasites, including fleas, have negative effects on nestling body mass (Richner et al., 1993; Merino and Potti, 1995; Tomás et al., 2007). Therefore, if these ectoparasites tend to feed on last-hatched nestlings, smaller nestlings would be more negatively affected by parasitism (Szép and Møller, 2000). Nevertheless, this lends very indirect support to the TCH and alternative explanations should be explored. Specifically, parasitised broods may suffer a food shortage which causes parents to feed the larger nestlings preferentially (Smiseth et al., 2003), resulting in poorer growth of smaller nestlings. Parents may also feed healthy nestlings preferentially (Saino et al., 2000), hence smaller nestlings might suffer a disadvantage if parents disfavour them for being unhealthy.
Smaller nestlings had significantly more leucocytes than larger ones. This finding, a priori, contradicts a pivotal prediction of the TCH. There is compelling evidence that an increase in the leucocyte count in birds is usually associated with the infestation or infection by different ecto- and endoparasites (Norris and Evans, 2000), so the smaller nestlings' greater investment in immune function could be explained by increased exposure to parasites. Similarly, Saino et al. (2001) and Parejo et al. (2007) found that last-hatched nestlings showed a higher immunological profile (level of antibodies) than their core siblings. However, in our study, neither the presence of fleas nor Blowflies affected immune function. A possible explanation is that smaller nestlings mounted an immune response against other parasites that were not analysed in the present study, such as viruses, bacteria, coccidia, haemosporidians or their dipteran vectors. In fact, the relative leucocyte count of smaller nestlings was higher in the humid forest than in the dry forest and, in a previous study carried out in 2016, we found a higher prevalence of the haemosporidian Leucocytozoon in the humid forest (Moreno-Rueda et al. in prep.). However, we cannot discard that higher leucocyte count in smaller nestlings is due to other causes not related to TCH. For example, parents could improve the immune system of smaller nestlings via resources in ovo (Roulin et al., 2008). Therefore, the overall evidence for the TCH in our study is weak and indirect.
4.2
Evidence against the Tasty Chick Hypothesis
However, some of our results do not support the TCH: the cutaneous immune response did not differ between larger and smaller nestlings and was unaffected by ectoparasites. The fact that exposure to parasites activates the immune system means that comparing immune system parameters, such as leucocyte count, is of no use when examining differences in immunocompetence, as these parameters probably mirror a mix of immunocompetence and parasite exposure. Immunocompetence can be measured by challenging the immune system with a novel stimulus to which it has not previously been exposed. PHA is a novel stimulus widely used to quantify immunocompetence (Kennedy and Nager, 2006). It has been shown to provoke inflammation with infiltration of different immune cells (Martin et al., 2006). Various studies have found that PHA testing normally reflects nestling body condition and nutritional status (e.g. Saino et al., 1997; Westneat et al., 2004; Martínez-De La Puente et al., 2013) and is related to nestling ectoparasite resistance (Martin et al., 2001; Tschirren et al., 2007; Bize et al., 2008). Hence, the TCH predicts a weaker inflammatory response to PHA in smaller compared to larger nestlings. This has been confirmed in House Martin (Delichon urbicum; Christe et al., 1998), Black-headed Gull (Chroicocephalus ridibundus; Müller et al., 2003), Barn Owl (Tyto alba; Roulin et al., 2003), and Collared Dove (Streptopelia decaocto; Eraud et al., 2008) nestlings. However, no within-brood differences in PHA immune response have been observed in Great Tit (Parus major; Roulin et al., 2003; Kilgas et al., 2010), Alpine Swift (Tachymarptis melba; Bize et al., 2005), Red-billed Chough (Pyrrohocorax pyrrhocorax; Banda and Blanco, 2008), or Blue Tit (this study) nestlings. In fact, Saino et al. (2001) even reported a stronger immune response in smaller than in larger Barn Swallow (Hirundo rustica) nestlings. Therefore, there is a lack of evidence to support one of the main predictions of the TCH, namely that smaller nestlings have poorer immune systems than larger nestlings.
For their part, Blowflies, unlike fleas, did not affect the body mass or tarsus length of smaller nestlings. Blowfly larvae are known to have negative effects on the physiology, growth and survival of Blue Tit nestlings (Hurtrez-Boussès et al., 1997; Bouslama et al., 2001; Tomás et al., 2007; Arriero et al., 2008; Castaño-Vázquez et al., 2018). However, these negative impacts of Blowflies may vary between years and according to weather conditions, so in some years they cause relatively little harm to nestlings (Merino and Potti, 1996; Simon et al., 2004). Furthermore, there is still no consensus on whether Blowfly larvae eat preferentially on last-hatched nestlings (Descamps et al., 2002; Simon et al., 2003). Hence, the apparent absence of any impact of Blowfly larvae on nestlings in our study may be due to a lack of any significant detrimental effects on Blue Tit nestlings in our population specifically during the year the study was carried out.
4.3
Tasty Chick Hypothesis: state of the art
The TCH predicts that last-hatched (marginal) nestlings must be less immunocompetent than core nestlings. As explained above, several studies performed in various species do not support this prediction (Saino et al., 2001; Roulin et al., 2003; Bize et al., 2005; the present study). In addition, the TCH predicts that nest-dwelling ectoparasites tend to feed more on the least immunocompetent nestling. Although some studies have shown that higher host immunocompetence reduces ectoparasite feeding, breeding and survival success (Tschirren et al., 2007; Bize et al., 2008), only a few studies support this prediction. Simon et al. (2003) reported that Blowfly larvae eat more blood from smaller than larger Blue Tit nestlings. O'Connor et al. (2014) evidenced aggregation of parasitic larvae of the dipteran Philornis downsi on just one nestling per brood. However, nestlings frequently move around more when parasitised, in an attempt to escape from parasites in the nest (Simon et al., 2005; O'Connor et al., 2010), so it is unclear whether the patterns reported by Simon et al. (2003) and O'Connor et al. (2014) are due to the feeding behaviour of parasites or nestling competition for the positions least exposed to ectoparasites. Furthermore, and in clear contrast with the TCH, several studies have shown that parasites prefer to feed on large, well-nourished nestlings before last-hatched nestlings (Roulin et al., 2003; Valera et al., 2004; Bize et al., 2008; Václav et al., 2008; Václav and Valera, 2018). Therefore, the overall evidence in favour of the TCH is, at best, only weak.
4.4
The evolution of hatching asynchrony in the Blue Tit
While the TCH seems unable to explain the evolution of hatching asynchrony in the Blue Tit, other hypotheses may explain this phenomenon in said species. Indeed, hatching asynchrony has been shown to be adaptive in Blue Tits, as asynchronous broods produce heavier offspring than synchronous broods, even when there was no brood reduction (Slagsvold et al., 1995). The most classical explanation for hatching asynchrony is to facilitate adaptive brood reduction (Stenning, 1996). According to this concept, in the scenario of a temporally and spatially unpredictable environment, parents might secondarily adjust their brood size to food availability, with small chicks sacrificed to ensure the survival of the remaining offspring if food resources become scarce (Magrath, 1989). Hatching asynchrony may also have evolved to reduce sibling competition, to decrease the peak in parental workload or to minimise predation risk and egg failure in some altricial birds (review in Magrath, 1990). Accordingly, Stenning (2008) showed that Blue Tit nestlings from asynchronous broods spent less time in their nests. Moreover, marginal nestlings from asynchronous broods typically die earlier compared to those in synchronous broods, with the parents saving the energy that they would have invested in chicks that would have likely died anyway.
5.
Conclusions
In conclusion, our study provides some, albeit indirect, evidence that the TCH might be applied to the Blue Tit and a specific ectoparasite as we found that the presence of Hen Fleas was more detrimental to smaller than larger nestlings (affecting their body mass). However, most of the results in our study did not support the TCH. Blowflies did not affect nestlings' morphometry and immune system. Moreover, we did not find evidence that smaller nestlings were less immunocompetent than larger nestlings as the immune response to PHA did not differ between both type of nestlings and smaller nestlings had more leucocytes than larger ones.
Author contributions
GMR designed the study. GMR, JLRS, EP, NB and MC carried out the fieldwork and collected the field data and blood samples. AS, CET and APJ analysed the blood smears. SK identified the fleas. GMR and JGB analysed the data. JGB and GMR wrote the manuscript with input from the other authors. All authors read and approved the final manuscript.
Ethics statement
The study was carried out with permission from the Andalusian Regional Government (reference: 03-06-15-259) with respect to bioethics and animal welfare and the Espacio Natural de Sierra Nevada with respect to environmental protection.
Declaration of competing interest
The authors declare that they have no competing interests.
Acknowledgements
We are grateful to the staff of the National Park of Sierra Nevada for their constant support. This study was supported by two projects in the National Plan of the Spanish Ministry of Economy and Competition (CGL2014-55969-P and CGL2017-84938-P) and by a project of the Andalusian government (A-RNM-48-UGR20), financed with FEDER funds from the European Union (EU). JLRS and EP were funded by Erasmus+ grants from the EU. JGB was supported by a FPU pre-doctoral contract from the Spanish Ministry of Education (FPU18/03034).
Arriero, E., Moreno, J., Merino, S., Martínez, J., 2008. Habitat effects on physiological stress response in nestling blue tits are mediated through parasitism. Physiol. Biochem. Zool. 81, 195-203.
Banda, E., Blanco, G., 2008. Influence of hatching asynchrony and within-brood parental investment on size, condition, and immunocompetence in nestling red-billed choughs. Biol. J. Linn. Soc. 94, 675-684.
Bennett, G.F., Whitworth, T.L., 1991. Studies on the life history of some species of Protocalliphora (Diptera: Calliphoridae). Can. J. Zool. 69, 2048-2058.
Bize, P., Jeanneret, C., Klopfenstein, A., Roulin, A., 2008. What makes a host profitable? Parasites balance host nutritive resources against immunity. Am. Nat. 171, 107-118.
Bize, P., Roulin, A., Tella, J.L., Richner, H., 2005. Female-biased mortality in experimentally parasitized Alpine swift Apus melba nestlings. Funct. Ecol. 19, 405-413.
Björklund, M., 1996. Similarity of growth among great tits (Parus major) and blue tits (P. caeruleus). Biol. J. Linn. Soc. 58, 343-355.
Bouslama, Z., Chabi, Y., Lambrechts, M., 2001. Chicks resist high parasite intensities in an Algerian population of blue tits. Ecoscience 8, 320-324.
Brinkhof, M.W.G., Heeb, P., Kölliker, M., Richner, H., 1999. Immunocompetence of nestling great tits in relation to rearing environment and parentage. Proc. R. Soc. Lond. B. 266, 2315-2322.
Brommer, J.E., Pitala, N., Siitari, H., Kluen, E., Gustafsson, L., 2011. Body size and immune defense of nestling blue tits (Cyanistes caeruleus) in response to manipulation of ectoparasites and food supply. Auk 128, 556-563.
Brown, C.R., Brown, M.B., 1986. Ectoparasitism as a cost of coloniality in cliff swallows (Hirundo pyrrhonota). Ecology 67, 1206-1218.
Bush, S.E., Clayton, D.H., 2018. Anti-parasite behaviour of birds. Phil. Trans. R. Soc. B. 373, 20170196.
Campbell, T.W., Ellis, C.K., 2007. Avian and Exotic Animal Hematology and Citology. Blackwell Publishing Ltd, Oxford.
Castaño-Vázquez, F., Martínez, J., Merino, S., Lozano, M., 2018. Experimental manipulation of temperature reduce ectoparasites in nests of blue tits Cyanistes caeruleus. J. Avian Biol. 49, e01695.
Christe, P., Møeller, A.P., de Lope, F., 1998. Immunocompetence and nestling survival in the house martin: the tasty chick hypothesis. Oikos 83, 175-179.
Christe, P., Richner, H., Oppliger, A., 1996. Begging, food provisioning, and nestling competition in great tit broods infested with ectoparasites. Behav. Ecol. 7, 127-131.
Cichoń, M., Dubiec, A., 2005. Cell-mediated immunity predicts the probability of local recruitment in nestling blue tits. J. Evol. Biol. 18, 962-966.
de Jong, A., 2019. Less is better. Avoiding redundant measurements in studies on wild birds in accordance to the principles of the 3Rs. Front. Vet. Sci. 6, 195.
Descamps, S., Blondel, J., Lambrechts, M., Hurtrez-Boussès, S., Thomas, F., 2002. Asynchronous hatching in a blue tit population: a test of some predictions related to ectoparasites. Can. J. Zool. 80, 1480-1484.
Dickens, M., Hartley, I.R., 2007. Differences in parental food allocation rules: evidence for sexual conflict in the blue tit? Behav. Ecol. 18, 674-679.
Dubiec, A., Cichoń, M., Deptuch, K., 2006. Sex-specific development of cell-mediated immunity under experimentally altered rearing conditions in blue tit nestlings. Proc. R. Soc. B. 273, 1759-1764.
Eraud, C., Trouvé, C., Dano, S., Chastel, O., Faivre, B., 2008. Competition for resources modulates cell-mediated immunity and stress hormone level in nestling collared doves (Streptopelia decaocto). Gen. Comp. Endocr. 155, 542-551.
Fitze, P.S., Clobert, J., Richner, H., 2004. Long-term life-history consequences of ectoparasite-modulated growth and development. Ecology 85, 2018-2026.
Forbes, S., Thornton, S., Glassey, B., Forbes, M., Buckley, N.J., 1997. Why parent birds play favourites. Nature 390, 351-352.
García-Navas, V., Ferrer, E.S., Serrano-Davies, E., 2014. Experimental evidence for parental, but not parentally biased, favouritism in relation to offspring size in blue tits Cyanistes caeruleus. Ibis 156, 404-414.
Garrido-Bautista, J., Soria, A., Trenzado, C.E., Pérez-Jiménez, A., Ros-Santaella, J.L., Pintus, E., et al., 2021. Oxidative status of blue tit nestlings varies with habitat and nestling size. Comp. Biochem. Physiol. A. 258, 110986.
Hannam, K., 2006. Ectoparasitic blow flies (Protocalliphora sp. ) and nestling Eastern bluebirds (Sialia sialis): direct effects and compensatory strategies. Can. J. Zool. 84, 921-930.
Hurtrez-Boussès, S., Perret, P., Renaud, F., Blondel, J., 1997. High blowfly parasitic loads affect breeding success in a Mediterranean population of blue tits. Oecologia 112, 514-517.
Kennedy, M., Nager, R., 2006. The perils and prospects of using phytohaemagglutinin in evolutionary ecology. Trends Ecol. Evol. 21, 653-655.
Kilgas, P., Tilgar, V., Külavee, R., Saks, L., Hõrak, P., Mänd, R., 2010. Antioxidant protection, immune function and growth of nestling great tits Parus major in relation to within-brood hierarchy. Comp. Biochem. Physiol. B. 157, 288-293.
Lochmiller, R.L., Deerenberg, C., 2000. Trade-offs in evolutionary immunology: just what is the cost of immunity? Oikos 88, 87-98.
Magrath, R.D., 1989. Hatching asynchrony and reproductive success in the blackbird. Nature 339, 536-538.
Martin, L.B., Han, P., Lewittes, J., Kuhlman, J.R., Klasing, K.C., Wikelski, M., 2006. Phytohemagglutinin-induced skin swelling in birds: histological support for a classic immunoecological technique. Funct. Ecol. 20, 290-299.
Martin, T.E., Møeller, A.P., Merino, S., Clobert, J., 2001. Does clutch size evolve in response to parasites and immunocompetence? Proc. Natl. Acad. Sci. U.S.A. 98, 2071-2076.
Martínez-De La Puente, J., Martínez, J., Rivero-De-Aguilar, J., Del Cerro, S., Merino, S., 2013. Vector abundance determines Trypanosoma prevalence in nestling blue tits. Parasitology 140, 1009-1015.
Merino, S., Potti, J., 1995. Mites and blowflies decrease growth and survival in nestling pied flycatchers. Oikos 73, 95-103.
Merino, S., Potti, J., 1996. Weather dependent effects of nest ectoparasites on their bird hosts. Ecography 19, 107-113.
Møeller, A.P., Arriero, E., Lobato, E., Merino, S., 2009. A meta-analysis of parasite virulence in nestling birds. Biol. Rev. 84, 567-588.
Moreno-Rueda, G., 2003. Selección de cajas-nido por aves insectívoras en Sierra Nevada. Zool. Baetica 13/14, 131-138.
Moreno-Rueda, G., Soler, M., Martín-Vivaldi, M., Palomino, J.J., 2009. Brood provisioning rate and food allocation rules according to nestling begging in a clutch-adjusting species, the Rufous-tailed Scrub-robin Cercotrichas galactotes. Acta Ornithol. 44, 167-175.
Müller, W., Dijkstra, C., Groothuis, T.G.G., 2003. Inter-sexual differences in T-cell-mediated immunity of black-headed gull chicks (Larus ridibundus) depend on the hatching order. Behav. Ecol. Sociobiol. 55, 80-86.
Norris, K., Evans, M.R., 2000. Ecological immunology: life history trade-offs and immune defense in birds. Behav. Ecol. 11, 19-26.
O'Brien, E.L., Dawson, R.D., 2009. Palatability of passerines to parasites: within-brood variation in nestling responses to experimental parasite removal and carotenoid supplementation. Oikos 118, 1743-1751.
Ochoa, D., Redondo, T., Moreno-Rueda, G., 2019. Mizutama: a quick, easy, and accurate method for counting erythrocytes. Physiol. Biochem. Zool. 92, 206-210.
O'Connor, J.A., Robertson, J., Kleindorfer, S., 2010. Video analysis of host-parasite interactions in nests of Darwin's finches. Oryx 44, 588-594.
O'Connor, J.A., Robertson, J., Kleindorfer, S., 2014. Darwin's finch begging intensity does not honestly signal need in parasitised nests. Ethology 120, 228-237.
Owen, J.C., 2011. Collecting, processing, and storing avian blood: a review. J. Field Ornithol. 82, 339-354.
Owen, J.P., Delany, M.E., Cardona, C.J., Bickford, A.A., Mullens, B.A., 2009. Host inflammatory response governs fitness in an avian ectoparasite, the northern fowl mite (Ornithonyssus sylviarum). Int. J. Parasitol. 39, 789-799.
Parejo, D., Silva, N., Avilés, J.M., 2007. Within-brood size differences affect innate and acquired immunity in roller Coracias garrulus nestlings. J. Avian Biol. 38, 717-725.
Pinheiro, J., Bates, D., DebRoy, S., Sarkar, D., R Core Team., 2019. nlme: linear and nonlinear mixed effects models. R Package Version 3.1-141.
Pitala, N., Siitari, H., Gustafsson, L., Brommer, J.E., 2009. Ectoparasites help to maintain variation in cell-mediated immunity in the blue tit-hen flea system. Evol. Ecol. Res. 11, 79-94.
Puchala, P., 2004. Detrimental effects of larval blow flies (Protocalliphora azurea) on nestlings and breeding success of tree sparrows (Passer montanus). Can. J. Zool. 82, 1285-1290.
Quinn, G.P., Keough, M.J., 2002. Experimental Design and Data Analysis for Biologists. Cambridge University Press, Cambridge.
R Development Core Team, 2020. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna.
Richner, H., Oppliger, A., Christe, P., 1993. Effect of an ectoparasite on reproduction in great tits. J. Anim Ecol. 62, 703-710.
Richner, H., Tripet, F., 1999. Ectoparasitism and the trade-off between current and future reproduction. Oikos 86, 535-538.
Roulin, A., Brinkhof, M.W.G., Bize, P., Richner, H., Jungi, T.W., Bavoux, C., et al., 2003. Which chick is tasty to parasites? The importance of host immunology vs. parasite life history. J. Anim. Ecol. 72, 75-81.
Roulin, A., Gasparini, J., Froissart, L., 2008. Pre-hatching maternal effects and the tasty chick hypothesis. Evol. Ecol. Res. 10, 463-473.
Saino, N., Calza, S., Møeller, A.P., 1997. Immunocompetence of nestling barn swallows in relation to brood size and parental effort. J. Anim. Ecol. 66, 827-836.
Saino, N., Incagli, M., Martinelli, R., Ambrosini, R., Møeller, A.P., 2001. Immunity, growth and begging behaviour of nestling barn swallows Hirundo rustica in relation to hatching order. J. Avian Biol. 32, 263-270.
Saino, N., Ninni, P., Incagli, M., Calza, S., Sacchi, R., Møeller, A.P., 2000. Begging and parental care in relation to offspring need and condition in the barn swallow (Hirundo rustica). Am. Nat. 156, 637-649.
Salvador, A., 2016. Herrerillo Comun-Cyanistes caeruleus. Enciclopedia Virtual de Los Vertebrados Espanoles. Museo Nacional de Ciencias Naturales, Madrid.
Schmid-Hempel, P., 2011. Evolutionary Parasitology. The Integrated Study of Infections, Immunology, Ecology, and Genetics. Oxford University Press, Oxford.
Schmoll, T., Dietrich, V., Winkel, W., Lubjuhn, T., 2004. Blood sampling does not affect fledging success and fledgling local recruitment in coal tits (Parus ater). J. Ornithol. 145, 79-80.
Simon, A., Thomas, D.W., Blondel, J., Lambrechts, M., Perret, P., 2003. Within-brood distribution of ectoparasite attacks on nestling blue tits: a test of the tasty chick hypothesis using inulin as a tracer. Oikos 102, 551-558.
Simon, A., Thomas, D.W., Blondel, J., Perret, P., Lambrechts, M., 2004. Physiological ecology of Mediterranean blue tits (Parus caeruleus L. ): effects of ectoparasites (Protocalliphora spp. ) and food abundance on metabolic capacity of nestlings. Physiol. Biochem. Zool. 77, 492-501.
Simon, A., Thomas, D.W., Speakman, J.R., Blondel, J., Perret, P., Lambrechts, M., 2005. Impact of ectoparasitic blowfly larvae (Protocalliphora spp. ) on the behavior and energetics of nestling blue tits. J. Field Ornithol. 76, 402-410.
Slagsvold, T., Amundsen, T., Dale, S., 1995. Costs and benefits of hatching asynchrony in blue tits Parus caeruleus. J. Anim. Ecol. 64, 563-578.
Smiseth, P.T., Bu, R.J., Eikenæs, A.K., Amundsen, T., 2003. Food limitation in asynchronous bluethroat broods: effects on food distribution, nestling begging, and parental provisioning rules. Behav. Ecol. 14, 793-801.
Smits, J.E., Bortolotti, G.R., Tella, J.L., 1999. Simplifying the phytohaemagglutinin skin-testing technique in studies of avian immunocompetence. Funct. Ecol. 13, 567-572.
Stenning, M.J., 1996. Hatching asynchrony, brood reduction and other rapidly reproducing hypotheses. Trends Ecol. Evol. 11, 243-246.
Stenning, M.J., 2008. Hatching asynchrony and brood reduction in blue tits Cyanistes caeruleus may be a plastic response to local oak Quercus robur bud burst and caterpillar emergence. Acta Ornithol. 43, 97-106.
Stenning, M., 2018. The Blue Tit. T & AD Poyser, London.
Streby, H.M., Peterson, S.M., Kapfer, P.M., 2009. Fledging success is a poor indicator of the effects of bird blow flies on ovenbird survival. Condor 111, 193-197.
Szép, T., Møeller, A.P., 2000. Exposure to ectoparasites increases within-brood variability in size and body mass in the sand martin. Oecologia 125, 201-207.
Tella, J.L., Lemus, J.A., Carrete, M., Blanco, G., 2008. The PHA test reflects acquired T-cell mediated immunocompetence in birds. PLoS One 3, e3295.
Tomás, G., Merino, S., Moreno, J., Morales, J., 2007. Consequences of nest reuse for parasite burden and female health and condition in blue tits, Cyanistes caeruleus. Anim. Behav. 73, 805-814.
Tripet, F., Richner, H., 1997a. The coevolutionary potential of a 'generalist' parasite, the hen flea Ceratophyllus gallinae. Parasitology 115, 419-427.
Tripet, F., Richner, H., 1997b. Host responses to ectoparasites: food compensation by parent blue tits. Oikos 78, 557-561.
Tripet, F., Richner, H., 1999. Dynamics of hen flea Ceratophyllus gallinae subpopulations in blue tit nests. J. Insect. Behav. 12, 159-174.
Tschirren, B., Bischoff, L.L., Saladin, V., Richner, H., 2007. Host condition and host immunity affect parasite fitness in a bird-ectoparasite system. Funct. Ecol. 21, 372-378.
Václav, R., Calero-Torralbo, M.A., Valera, F., 2008. Ectoparasite load is linked to ontogeny and cell-mediated immunity in an avian host system with pronounced hatching asynchrony. Biol. J. Linn. Soc. 94, 463-473.
Václav, R., Valera, F., 2018. Host preference of a haematophagous avian ectoparasite: a micronutrient supplementation experiment to test an evolutionary trade-off. Biol. J. Linn. Soc. 125, 171-183.
Valera, F., Hoi, H., Darolov, A., Kristofik, J., 2004. Size versus health as a cue for host choice: a test of the tasty chick hypothesis. Parasitology 129, 59-68.
Westneat, D.F., Weiskittle, J., Edenfield, R., Kinnard, T.B., Poston, J.P., 2004. Correlates of cell-mediated immunity in nestling house sparrows. Oecologia 141, 17-23.
Zuur, A.F., Ieno, E.N., Walker, N., Saveliev, A.A., Smith, G.M., 2009. Mixed Effects Models and Extensions in Ecology with R. Springer, New York.
Zuur, A.F., Ieno, E.N., Elphick, C.S., 2010. A protocol for data exploration to avoid common statistical problems: data exploration. Method. Ecol. Evol. 1, 3-14.
Table
1.
Results of the linear mixed-effects models and linear models for the number of leucocytes, cutaneous immune response, and its within-brood differences (values for larger nestling minus smaller ones) of Blue Tit nestlings.
Variable
Chi-square (df = 1)
p-value
Number of leucocytes
Forest type
5.39
0.02
Nestling rank
6.27
0.012
Forest type*Nestling rank
5.81
0.02
Cutaneous immune response
Forest type
0.57
0.45
Nestling rank
0.19
0.66
Forest type*Nestling rank
0.24
0.62
Within-brood differences
Variable
F-value (df = 1)
P-value
Within-brood difference in leucocytes
Forest type
6.36
0.02
Flea prevalence
0.04
0.84
Blowfly prevalence
1.20
0.29
Within-brood difference in immune response
Forest type
0.59
0.45
Flea prevalence
1.17
0.29
Blowfly prevalence
0.51
0.48
Significant predictors (p < 0.05) are marked in bold.
Table
2.
Body mass, tarsus length and body condition index (BCI) of smaller and larger Blue Tit nestlings and the within-brood differences between Hen Flea-infested and uninfested broods.
Empty Cell
Infested broods (n = 12)
Uninfested broods (n = 25)
t-value
p-value
Larger nestlings
Tarsus length (mm)
16.63 ± 0.50
16.79 ± 0.52
−0.92
0.36
Body mass (g)
10.65 ± 0.58
10.62 ± 0.74
0.11
0.92
BCI (residuals)
0.045 ± 0.041
0.027 ± 0.087
0.67
0.51
Smaller nestlings
Tarsus length (mm)
15.76 ± 0.47
16.19 ± 0.45
−2.70
0.01
Body mass (g)
9.06 ± 0.71
9.54 ± 0.80
−1.78
0.08
BCI (residuals)
−0.042 ± 0.074
−0.029 ± 0.087
−0.44
0.67
Within-brood difference
Tarsus length (mm)
0.87 ± 0.62
0.60 ± 0.65
1.17
0.25
Body mass (g)
1.59 ± 0.68
1.08 ± 0.30
3.18
0.003
BCI (residuals)
0.087 ± 0.062
0.056 ± 0.058
1.46
0.15
Differences were tested using t-tests. Mean values are given with the SD.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds