Alexandra A. Grossi, Min Zhang, Fasheng Zou, Daniel R. Gustafsson. 2024: Bird species (Charadriiformes) does not impact endosymbiotic bacteria (Gammaproteobacteria) of their ectoparasites (Insecta: Phthiraptera: Ischnocera). Avian Research, 15(1): 100206. DOI: 10.1016/j.avrs.2024.100206
Citation:
Alexandra A. Grossi, Min Zhang, Fasheng Zou, Daniel R. Gustafsson. 2024: Bird species (Charadriiformes) does not impact endosymbiotic bacteria (Gammaproteobacteria) of their ectoparasites (Insecta: Phthiraptera: Ischnocera). Avian Research, 15(1): 100206. DOI: 10.1016/j.avrs.2024.100206
Alexandra A. Grossi, Min Zhang, Fasheng Zou, Daniel R. Gustafsson. 2024: Bird species (Charadriiformes) does not impact endosymbiotic bacteria (Gammaproteobacteria) of their ectoparasites (Insecta: Phthiraptera: Ischnocera). Avian Research, 15(1): 100206. DOI: 10.1016/j.avrs.2024.100206
Citation:
Alexandra A. Grossi, Min Zhang, Fasheng Zou, Daniel R. Gustafsson. 2024: Bird species (Charadriiformes) does not impact endosymbiotic bacteria (Gammaproteobacteria) of their ectoparasites (Insecta: Phthiraptera: Ischnocera). Avian Research, 15(1): 100206. DOI: 10.1016/j.avrs.2024.100206
Guangdong Key Laboratory of Animal Conservation and Resource Utilization, Guangdong Public Library of Wild Animal Conservation and Utilization, Institute of Zoology, Guangdong Academy of Sciences, Guangzhou, 510260, China
b.
Key Laboratory for Biodiversity Science and Ecological Engineering, College of Life Sciences, Beijing Normal University, Beijing, 100875, China
Funds:
the National Natural Science Foundation of China32001098
the National Natural Science Foundation of China31961123003
the Introduction of FullTime High-Level Talent Fund of the Institute of Zoology, Guangdong Academy of SciencesGIABR-GJRC201701
GDAS Special Project of Science and Technology Development2022GDASZH-2022010106
GDAS Special Project of Science and Technology Development2019GDASYL-0203001
the Foreign Young Talent PlanQN20200130012
the Pearl River Talent Recruitment Program of Guangdong Province2019QN01N968
These agencies had no influence over the design and execution of this study
Guangdong Key Laboratory of Animal Conservation and Resource Utilization, Guangdong Public Library of Wild Animal Conservation and Utilization, Institute of Zoology, Guangdong Academy of Sciences, Guangzhou, 510260, China. E-mail address: zhangm@giz.gd.cn (M. Zhang)
Endosymbiotic bacteria of insects can facilitate host expansion into novel niches by providing their host with a fitness benefit such as vitamins or amino acids that are otherwise lacking in their hosts' diet. This close association can lead to cospeciation between insects and their symbionts; however, the symbionts’ small genome size leaves it susceptible to genome derogation which can result in symbiont replacement. Here, we screen chewing lice infesting shorebirds and terns to see what endosymbiotic bacteria are present, and build a summary phylogeny that includes louse endosymbiont sequences from this study as well as those from other louse genera, insects and bacteria strains from GenBank. We found a Sodalis-allied endosymbiont in Carduiceps, Lunaceps, Quadraceps, and Saemundssonia, as well as symbionts belonging to the family Enterobacteriaceae in Lunaceps, and Quadraceps. No louse species were host to both endosymbionts; however, the birds Kentish Plover (Charadrius alexandrinus) and Greater Crested Tern (Thalasseus bergii) were host to two genera of lice, each of which was infested with a different group of endosymbionts. In the summary phylogeny the endosymbionts from shorebirds, and tern lice did not form a monophyletic group, and therefore likely acquired their bacterial endosymbionts multiple times.
Endosymbionts are commonly associated with insects and have interactions ranging from parasitic to mutualistic. For instance, the symbiont genus Wolbachia infects a wide range of insects and is able to manipulate the reproductive characteristics of their host by causing cytoplasmic incompatibility, parthenogenesis, male-killing and feminization (Werren et al., 2008; Saridaki and Bourtzis, 2010). Other endosymbiotic bacteria such as Buchnera aphidicola found in aphids are responsible for the synthesis of essential amino acids that are lacking in the aphids' diet of plant sap (Wilson et al., 2010). Therefore, bacterial acquisition can be critically important to an insect's fitness as it can facilitate expansion to new ecological niches (Douglas, 2009). The genome of an endosymbiont is often smaller than their free-living counterpart as it relies on its host for many functions (Wernegreen, 2015). This can result in mutations accumulating in the symbionts genome due to, e.g., a loss of a DNA repair system, genetic drift, and lack of recombination (McCutcheon and Moran, 2012), which in turn can lead to genome degeneration in the endosymbiont, ultimately resulting in symbiont extinction and replacement.
Some of the most prominent bacterial lineages that have become established within insects belong to the class Gammaproteobacteria (Husník et al., 2011; Santos-Garcia et al., 2017), including the genus Sodalis (Gammaproteobacteria: Enterobacterales: Pectobacteriaceae). Sodalis glossinidius was first described from the Tsetse Fly (Glossina morsitans Westwood, 1851) (Dale and Maudlin, 1999), and subsequently Sodalis or Sodalis-allied species have been found in several insect groups including aphids (Aphididae), weevils (Curculionoidea), louse flies (Hippoboscidae), and sweat bees (Halictidae) (Nováková and Hypša, 2007; Wilson et al., 2010; Toju et al., 2013; Rubin et al., 2018). Recently chewing lice (Insecta: Phthiraptera) have also been added to this list (Fukatsu et al., 2007).
Chewing lice are permeant ectoparasites of birds and mammals that have no free-living stage; therefore, they complete their whole life cycle upon their host (Marshall, 1981). Since lice are wingless, transmission between hosts generally only happens when these come into direct contact with each other. This is usually in the form of vertical transmission between parents and nestlings (Clayton and Tompkins, 1994; Lee and Clayton, 1995; Brooke, 2010), or horizontal transmission during mating (Hillgarth, 1996). However, exceptions are known, such as phoretically hitching rides between hosts on hippoboscid flies. This phoretic behavior is one way that lice may be able to come into contact with novel bird host species and could facilitate host switching (Lee et al., 2022).
Louse endosymbionts have been known since the 1930s (Ries, 1931), but only recently have DNA sequencing and phylogenetic analysis been used to analyze these bacteria. These methods revealed that the symbiont of the Slender Pigeon Louse (Columbicola columbae Linnaeus, 1758) is allied to Sodalis glossinidius from the Tsetse Fly (Fukatsu et al., 2007). Subsequently, Sodalis-allied symbionts have been found in multiple species of Columbicola, Guimaraesiella, and Brueelia (Smith et al., 2013; D’Alessio, 2023; Sweet et al., 2023; Grossi et al., 2024), as well as some bacterial lineages that fall outside Sodalis (Smith et al., 2013). Each species of chewing lice is believed to harbor a single endosymbiotic bacterial lineage that has been maternally transmitted (Smith et al., 2013). This lineage is housed in specialized cells called bacteriocytes clustered on both sides of the abdomen in males and immature females; however, in mature females the cells were only found in ovarian tissue (Smith et al., 2013).
Ischnoceran chewing lice primarily consume the feather barbs, which are composed of keratin and lack amino acids essential to the survival of the lice. This lack of amino acids caused Alickovic et al. (2021) to hypothesize that the endosymbionts of lice synthesize these amino acids for their hosts. Alickovic et al. (2021) examined the genome of the endosymbiont found in Columbicola wolffhuegeli (Eichler, 1952) parasitizing the Pied Imperial Pigeon (Ducula bicolor) but failed to identify many of the genes that would be expected to support metabolism of amino acids. The role of these endosymbiotic bacteria is thus still unclear.
The phylogenies of the Sodalis-allied bacteria from Columbicola and Brueelia both have comb-like topography with short internal nodes and long terminal branches (Smith et al., 2013; D’Alessio, 2023). This may be caused by repeated recent symbiont acquisition/replacement events from a common bacterial source, and there appears to be little signal of co-speciation in these louse-symbiont associations. However, in the songbird louse genus Guimaraesiella Eichler, 1949, there is a strong signal of co-speciation between the lice and their symbionts (Grossi et al., 2024). It thus appears that some Sodalis endosymbionts have cospeciated with their louse hosts, but the factors determining whether or not co-speciation happens are unclear.
Here we examine a fourth louse-symbiont system, that of chewing lice parasitizing shorebirds and terns (Charadriiformes). In general, each shorebird/tern species is parasitized by 1–3 species of lice in the Quadraceps-complex (in this case Lunaceps, Quadraceps, and Saemundssonia), with some species also being parasitized by the distantly related genus Carduiceps Clay & Meinertzhagen, 1939. The Quadraceps-complex includes several distinct lineages that have adapted to different microhabitats of the hosts, such as the wings and the head (Johnson et al., 2012). As multiple species of ischnoceran lice are typically easy to collect from each bird species, we ask the question: do ischnoceran lice occurring on the same host bird species share the same endosymbiotic bacteria? Additionally, does louse relatedness impact symbiont linages? That is, do closely related lice on the same or closely related bird species (e.g., two members of the Quadraceps-complex) have more similar symbiont lineages than more distally related lice parasitizing the same host species (e.g., Carduiceps and Quadraceps-complex).
2.
Materials and methods
2.1
Sample collection
Lice were collected from birds caught on the coast of China at three locations (Dongli: 20.84° N, 110.37° E; Hebei: 20.91° N, 110.16° E; Tujiao: 20.89° N, 110.16° E) on the Leizhou Peninsula in western Guangdong and on Chiyu Island (23.32° N, 117.12° E) near Shantou in eastern Guangdong in 2020, using mist nets. Birds were caught under permit 2019-48 from the Leizhou Bureau of Natural Resources. Lice were removed by fumigation following the protocol outlined by Gustafsson et al. (2019). Birds were identified using MacKinnon and Phillipps (2000) and Arlott (2017) and taxonomy follows Clements et al. (2022). Lice were stored in absolute ethanol at −80 ℃, until DNA extraction.
2.2
DNA extraction and PCR
DNA extraction followed the protocol outlined by Grossi et al. (2024). For polymerase chain reactions (PCR), we used either Cytiva PureTaq Ready-To-Go beads (GE Healthcare, Vienna, Austria) or Qiagen Hot StarTaq Master Mix Kit (Qiagen, Shanghai, China), following the manufacture's protocol for 25 μL reactions. For bacteria we targeted 16S ribosomal RNA gene (16S) using primers: SodF, and R1060 (Nováková and Hypša, 2007), and thermocycle: 95 ℃ for 5 min, 35 cycles (95 ℃ for 30 s, 56 ℃ for 1 min, 72 ℃ for 1 min), 72 ℃ for 10 min. For lice we targeted cytochrome oxidase subunit 1 (COI) using primers: L6625, and H7005 (Hafner et al., 1994), and elongation factor 1α (EF-1α) using primers: EF1-For3, and EF1-CH10 (Danforth and Ji, 1998), and thermocycle: 94 ℃ for 2 min, 35 cycles (94 ℃ for 30 s, 46 ℃ for 30 s, 72 ℃ for 30 s), 72 ℃ for 7 min. PCR products were screened using gel electrophoresis and those with satisfactory bands were sent for sequencing at Tianyi Huiyuan Gene Technology, Co. Ltd. (Guangzhou, China).
2.3
Phylogenetic analysis
Sequences were assembled using the de novo assemble tool in Geneious Prime® v.2022.1.1, and the ends were trimmed. Sequences were aligned using MUSCLE (Edgar, 2004) with the default settings and checked manually. Three phylogenetic trees were assembled: a louse tree used to confirm identity, a bacteria tree from the lice newly sequenced in this study and a summary tree composed of bacteria sequenced in this study as well as those from other louse genera, insects and bacteria strains from GenBank (https://www.ncbi.nlm.nih.gov/genbank), for accession numbers of sequences pulled from GenBank see Appendix Table S1. Only unique louse/bacteria combination from the newly sequenced bacteria tree were carried over to the summery tree. To determine how many operational taxonomic units (OTUs) were present in the bacteria tree we used a General Mixed Yule Coalescent (Fujisawa and Barraclough, 2013) in RStudio 2023.03.1 (R Core Team, 2018) the “gmyc” function in the splits package was used to determine the number of OTUs (Joseph and Vakayil, 2022). Substitution models for each gene were evaluated in MEGA11 (Kumar et al., 2021), with the best model for lice being, CO1: HKY + G + I, EF-1α: HKY + G, and for both bacteria trees: HKY + G + I.
The two louse genes were concatenated, and this data set was used for all downstream analyses. As an outgroup for lice, we used Campanulotes compar (Burmeister, 1838) (accession no. CO1: AF384997, and EF-1α: HQ332855) and for bacteria Vibrio cholerae (accession no. LC487865). Bayesian analysis was conducted using BEAST v.2.7.3 (Suchard et al., 2018) with the following settings: the appropriate model(s) for each gene or gene partition were selected (trees were linked for the louse data set), Gamma category count was set to four, Yule Model was used, Markov chain Monte Carlo (MCMC) was run for 1 × 108 generations, and sampled every 1000 trees. The log file produced by BEAST was examined in Tracer (Rambaut et al., 2018) to assess MCMC for convergence. TreeAnnotator v2.7.3 was then used for tree integration, we discarded the first 10,000 trees (10%) as burn-in and constructed a maximum clade credibility tree.
2.4
Louse identification
After DNA extraction the gut was dissected out, and the louse exoskeleton was placed into clove oil for 30 min up to 24 h before it was slide mounted in Canada balsam and allowed to cure at room temperature for 30 days. All slides were deposited at the Institute of Zoology, Guangdong Academy of Sciences, Guangzhou, China. Specimens were examined through a Nikon Eclipse Ni microscope (Nikon Corporation, Tokyo, Japan) and identified using Hopkins and Timmermann (1954), Timmermann (1954a, 1952, 1950), Gustafsson and Olsson (2012), as well as through direct comparison with previously identified specimens in cases where no adequate illustrations or descriptions have been published.
3.
Results
3.1
Sample collection
In total 85 individual birds, representing 11 species were examined for lice, of which 68 (80%) were infested by at least one species of louse. See Table 1 for a breakdown by bird and louse species.
Table
1.
Bird species examined in this study, and the number of those infested with chewing lice.
All lice were found to belong to one of four genera: Carduiceps or Lunaceps (Clay & Meinertzhagen, 1939), on sandpipers (Calidrinae), and Quadraceps Clay & Meinertzhagen, 1939, or Saemundssonia Timmermann, 1936, on the other hosts. In two cases (J4451 and J4456), Lunaceps specimens were found on plovers, presumably as a result of straggling. In addition, samples from two different specimens of Spoon-billed Sandpiper (Calidris pygmaea) were found to represent different species of Lunaceps, with one (Lunaceps schismatus Gustafsson & Olsson, 2012) presumably being a straggler. The second species of Lunaceps is morphologically close to Lunaceps falcinellus Timmermann, 1954, but genetically distinct, and may represent an undescribed species. Apart from the specimens from terns, these specimens were previously reported by Grossi et al. (2023).
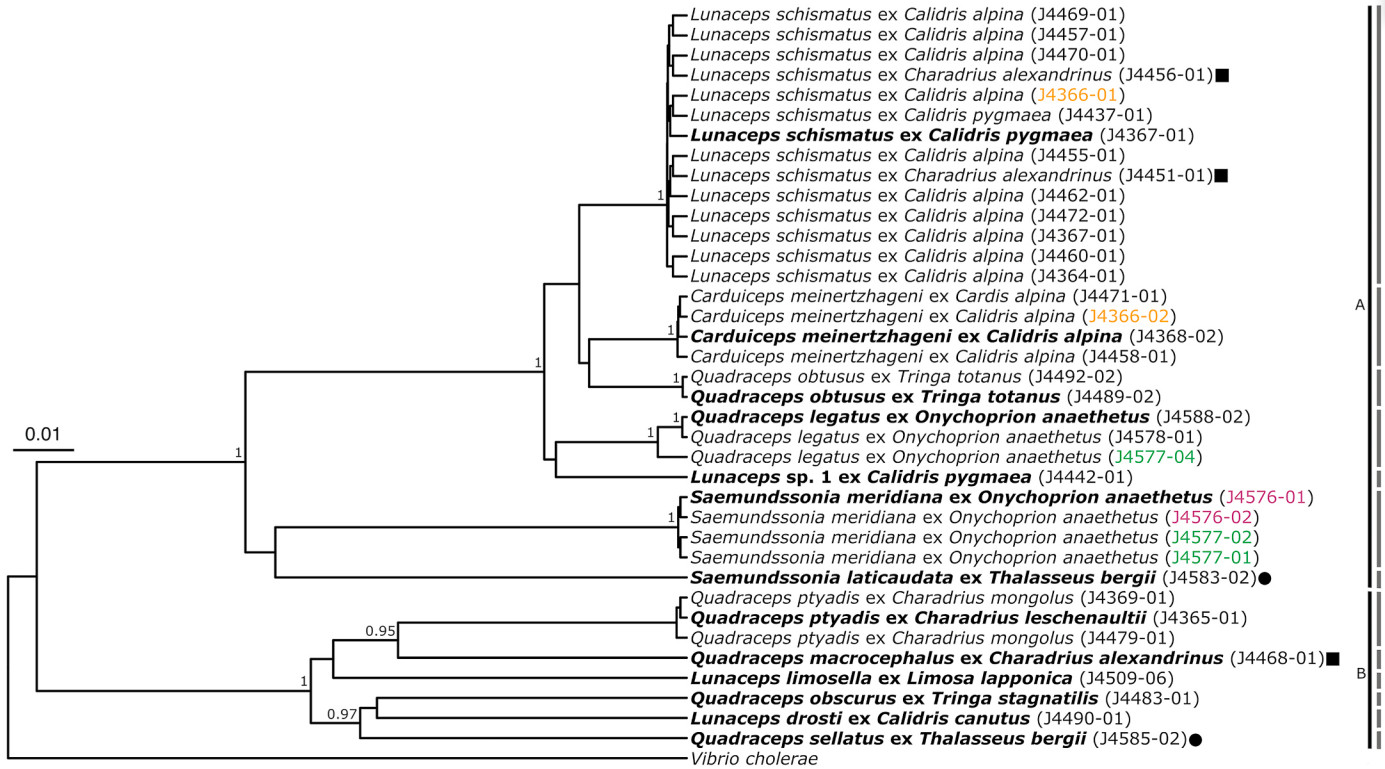
The louse phylogeny (Fig. 1) was constructed from CO1, 377 bp (203 variable sites, 192 parsimony-informative sites) and EF-1α, 348 bp (99 variable sites, 74 parsimony-informative sites). In this phylogeny, the genera Lunaceps and Saemundssonia are both recovered as monophyletic with good support, but both are nested inside a paraphyletic Quadraceps; Carduiceps is placed outside the Quadraceps-complex. Within Quadraceps, species grouped together in accordance with louse host associations.
Figure
1.
Bayesian phylogenetic tree of chewing lice infesting shorebirds and terns. Bolded taxa indicate sequences that are included in the summary phylogeny (Fig. 3), colored voucher numbers indicate lice that came from the same host individual. Only posterior probabilities greater than 0.90 were included.
We found endosymbiotic bacteria in each of the four chewing lice genera (Table 2). The phylogeny of the endosymbiotic bacteria from the lice infesting shore birds was constructed from 16S, 890 bp (242 variable sites, 167 parsimony-informative sites) and shows two distinct clades referred to as A and B (Fig. 2). The OTU analysis found 13 distinct units, see Fig. 2.
Table
2.
Collection and GenBank accession numbers for specimens included in this study.
Louse species
Bird host species
Voucher No.
Loc.
Symbiont (16S) Accession No.
Lice Accession No.
CO1
Ef-1α
Carduiceps meinertzhageni
Calidris alpina
J4366-02
L3
PP081401
PP082907
PP092242
Carduiceps meinertzhageni
Calidris alpina
J4368-02
L3
PP081403
PP082909
PP092244
Carduiceps meinertzhageni
Calidris alpina
J4458-01
L3
PP081411
PP082917
PP092252
Carduiceps meinertzhageni
Calidris alpina
J4471-01
L1
PP081418
PP082924
PP092259
Lunaceps sp.
Calidris pygmaea
J4442-01
L3
PP081406
PP082912
PP092247
Lunaceps drosti
Calidris canutus
J4490-01
L1
PP081423
PP082929
PP092264
Lunaceps limosella
Limosa lapponica
J4509-06
L1
PP081425
PP082931
PP092266
Lunaceps schismatus
Calidris pygmaea
J4437-01
L3
PP081405
PP082911
PP092246
Lunaceps schismatus
Charadrius alexandrinus
J4451-01
L3
PP081407
PP082913
PP092248
Lunaceps schismatus
Charadrius alexandrinus
J4456-01
L3
PP081409
PP082915
PP092250
Lunaceps schismatus
Calidris alpina
J4364-01
L3
PP081398
PP082904
PP092239
Lunaceps schismatus
Calidris alpina
J4366-01
L3
PP081400
PP082906
PP092241
Lunaceps schismatus
Calidris alpina
J4367-01
L3
PP081402
PP082908
PP092243
Lunaceps schismatus
Calidris alpina
J4457-01
L3
PP081410
PP082916
PP092251
Lunaceps schismatus
Calidris alpina
J4460-01
L3
PP081412
PP082918
PP092253
Lunaceps schismatus
Calidris alpina
J4462-01
L3
PP081413
PP082919
PP092254
Lunaceps schismatus
Calidris alpina
J4467-01
L1
PP081414
PP082920
PP092255
Lunaceps schismatus
Calidris alpina
J4469-01
L3
PP081416
PP082922
PP092257
Lunaceps schismatus
Calidris alpina
J4470-01
L1
PP081417
PP082923
PP092258
Lunaceps schismatus
Calidris alpina
J4472-01
L3
PP081419
PP082925
PP092260
Lunaceps schismatus
Calidris alpina
J4455-01
L3
PP081408
PP082914
PP092249
Quadraceps legatus
Onychoprion anaethetus
J4578-01
CI
PP081431
PP082937
PP092272
Quadraceps legatus
Onychoprion anaethetus
J4577-04
CI
PP081430
PP082936
PP092271
Quadraceps legatus
Onychoprion anaethetus
J4588-02
CI
PP081434
PP082940
PP092275
Quadraceps macrocephalus
Charadrius alexandrinus
J4468-01
L1
PP081415
PP082921
PP092256
Quadraceps obscurus
Tringa stagnatilis
J4483-01
L1
PP081421
PP082927
PP092262
Quadraceps obtusus
Tringa totanus
J4489-02
L1
PP081422
PP082928
PP092263
Quadraceps obtusus
Tringa totanus
J4492-02
L3
PP081424
PP082930
PP092265
Quadraceps ptyadis
Charadrius leschenaultii
J4365-01
L3
PP081399
PP082905
PP092240
Quadraceps ptyadis
Charadrius mongolus
J4369-01
L2
PP081404
PP082910
PP092245
Quadraceps ptyadis
Charadrius mongolus
J4479-01
L1
PP081420
PP082926
PP092261
Quadraceps sellatus
Thalasseus bergii
J4585-02
CI
PP081433
PP082939
PP092274
Saemundssonia laticaudata
Thalasseus bergii
J4583-02
CI
PP081432
PP082938
PP092273
Saemundssonia meridiana
Onychoprion anaethetus
J4576-01
CI
PP081426
PP082932
PP092267
Saemundssonia meridiana
Onychoprion anaethetus
J4576-02
CI
PP081427
PP082933
PP092268
Saemundssonia meridiana
Onychoprion anaethetus
J4577-01
CI
PP081428
PP082934
PP092269
Saemundssonia meridiana
Onychoprion anaethetus
J4577-02
CI
PP081429
PP082935
PP092270
Voucher number, the first four digits refer to the host individual and the last two digits refer to a unique louse. Thus, J4366–01 and J4366-02 are different lice from the same host individual. Location abbreviation: CI – Chiyu Island, Shantou; L1 – Dongli, Leizhou; L2 – Hebei, Leizhou; L3 – Tujiao, Leizhou, Guangdong, China.
Figure
2.
Bayesian phylogenetic tree of endosymbiotic bacteria from chewing lice infesting shorebirds and terns. Bolded taxa indicate sequences that are included in the summary phylogeny (Fig. 3), colored voucher numbers indicate lice that came from the same host individual, symbols denote bird species that were infested with chewing lice with endosymbionts found in both Clades A and B. Only posterior probabilities greater than 0.90 were included. Black lines denote clades referenced to in text and grey lines indicate operational taxonomic units.
Each louse species in this study has endosymbiotic bacteria that falls into either clade A or B, and no louse species has endosymbiotic bacteria from both clades. In two cases (Calidris alpina J4366 and Onychoprion anaethetus J4577), lice of different genera found on the same bird host individual tested positive for bacteria from the same clade. However, in the case of the lice from Thalasseus bergii (from two different host individuals J4583, J4585), the two louse species obtained tested positive for symbionts belonging to different clades. The two louse species obtained from Charadrius alexandrinus also tested positive for different symbiont clades, but in this case the Lunaceps samples are likely stragglers. Notably, each bacterial OTU was associated with exactly one louse species, but symbionts from closely related louse species did not always group together.
Prevalence of infection for clade A: Carduiceps 80% (4/5), Lunaceps 50% (15/30), Quadraceps 11% (4/36) and Saemundssonia 63% (5/8). Prevalence of infection for clade B: Carduiceps 0%, Lunaceps 7% (2/30), Quadraceps 19% (7/36) and Saemundssonia 0%.
3.4
Summary tree
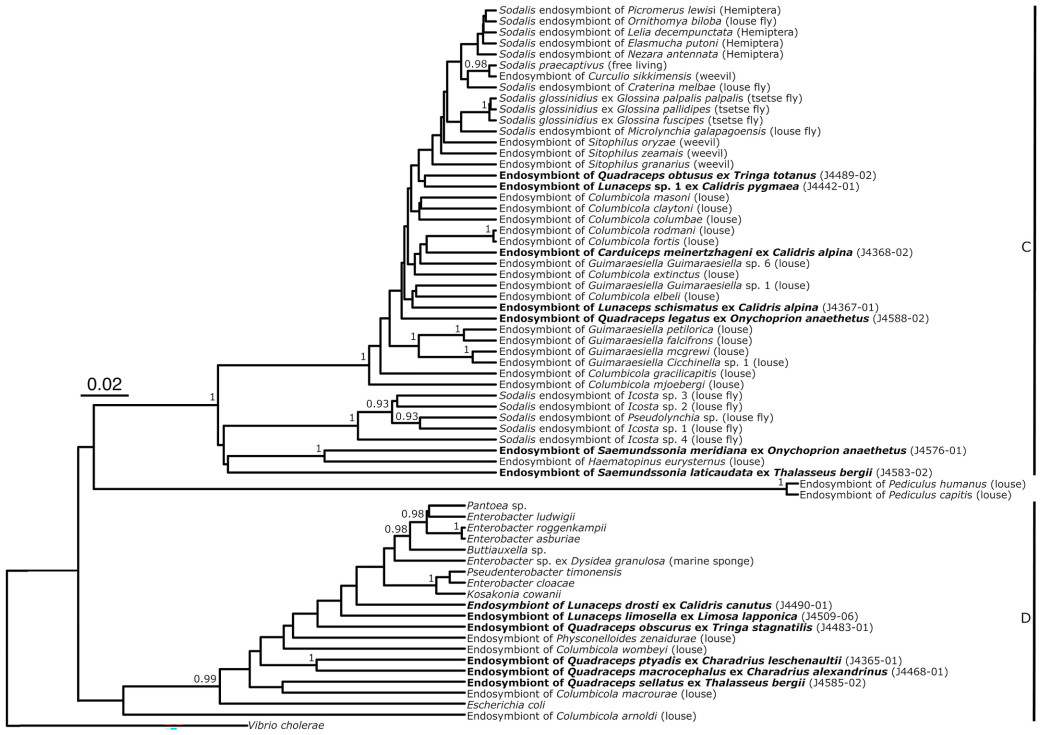
One symbiont sequence from each OTU was selected for inclusion in a larger analysis of related bacteria from other louse groups as well as other insects (Fig. 3), constructed from 16S, 848 bp (316 variable sites, 220 parsimony-informative sites). Most deeper nodes of this tree are unsupported, but the division between the same two clades as in Fig. 2 is evident. Symbionts from shorebird lice are scattered throughout both clades, and few shorebird louse symbionts appear to be closely related. Notably, Sodalis symbionts from hippoboscid flies were not placed close to symbionts from Guimaraesiella spp., known to be phoretic on these flies. Clade D in this tree also include bacteria belonging to genera other than Sodalis.
Figure
3.
Bayesian phylogenetic tree of endosymbiotic bacteria from chewing lice infesting shorebirds and terns, other genera of chewing lice, insects, and bacteria strains from GenBank (see Appendix Table S1 for accession numbers). Bolded taxa indicate newly sequenced endosymbiotic bacteria from this study. Only posterior probabilities greater than 0.90 were included. Black lines denote clades referenced to in text.
The ischnoceran chewing lice of most shorebirds belong to the Quadraceps-complex, a morphologically diverse group of lice that includes several microhabitat specialists. Notably, whereas e.g., the head and body lice of many songbirds are distantly related (e.g., de Moya et al., 2019), those of many shorebirds are thus closely related. One notable exception to this is the small group of birds that are parasitized by lice in the genus Carduiceps, which are distantly related to the Quadraceps-complex, and possibly more closely related to lice from other host groups (Gustafsson and Olsson, 2017). This variety of relationships between lice within a relatively small group of bird hosts makes shorebird and tern lice ideal for examining the distribution patterns of their bacterial symbionts.
The louse-bacteria systems studied to date show two almost diametrically opposite patterns: in Columbicola there appears to be little co-speciation and wide-spread replacement of bacterial strains, whereas in Guimaraesiella the symbionts appear to co-speciate extensively with their hosts. In shorebird and tern lice, we find that bacteria from each of the four louse genera examined are mixed together, with little resemblance between the louse phylogeny (Fig. 1) and that of their bacteria (Fig. 2). Put into a larger context, the bacteria of shorebird lice are scattered throughout the tree (Fig. 3), and thus potentially more reminiscent of those of Columbicola spp.
The symbionts recovered from shorebird lice fell into two different clades. Sodalis-allied symbionts were present in all four genera examined (Carduiceps, Lunaceps, Quadraceps and Saemundssonia), whereas endosymbionts belonging to the family Enterobacteriaceae were only found in Lunaceps and Quadraceps. Potentially, this is due to the larger number of Lunaceps and Quadraceps examined, but more data is needed to evaluate whether both these clades are more widely distributed across shorebird lice. Judging from the summary tree (Fig. 3), both clades seem to be established in many different louse groups. Both of these endosymbiont groups have previously been recorded from Columbicola (Smith et al., 2013); however, only Sodalis-allied symbionts have been recorded from Brueelia and Guimaraesiella (D’Alessio, 2023; Sweet et al., 2023; Grossi et al., 2024).
Each louse species was associated with a single bacterial OTU, even in cases where the same louse species occurs on multiple bird host species (e.g., J4365 and J4479 in Fig. 2). Thus, it does not appear that the bird hosts are the source of the endosymbiotic bacteria. However, when comparing different louse species associated with the same host species, different patterns appear. First, in the case of lice from Dunlins (Calidris alpina), each louse species was associated with a single bacterial OTU, but these were relatively closely related. Second, in the case of lice from Bridled Terns (Onychoprion anaethetus), each louse species was associated with a different bacterial symbiont lineage, and while they both were placed in Clade A in Fig. 2, they are not closely related within this clade. Finally, the two louse species from the Greater Crested Tern (Thalasseus bergii) were even more distantly related, with one symbiont lineage in each of Clades A and B (Fig. 2). Thus, host associations of the lice do not seem to predict relationships of their bacterial symbionts.
Notably, when seen in isolation, the shorebird and tern louse symbionts appear to show a mixture of the two speciation types seen in other examined louse symbionts. The evolutionary history of the symbionts of Columbicola is characterized by short terminal branches and longer internal branches. Smith et al. (2013) ran multiple simulations and found that this typology was formed by repeated endosymbiont acquisition. Repeated replacement of Sodalis symbionts is not unique to chewing lice, but is found in many insects such as planthoppers, mealybugs, weevils and louse flies (Toju et al., 2013; Husnik and McCutcheon, 2016; Šochová et al., 2017; Michalik et al., 2021). This pattern is also seen in Clade B here, and in the two symbiont OTUs from Saemundssonia spp. In contrast, much of the rest of Clade A has shorter internal branches, more reminiscent of those found in the symbionts of Guimaraesiella spp. (Grossi et al., 2024). However, in the summary tree (Fig. 3), these differences seem to disappear, and each of the Clade A OTUs are connected to purportedly closely related species by longer internal branches.
The bird hosts sampled for this study were all caught at similar latitudes in the same climatic zone and often in the same locality, most of which represent wintering grounds for the hosts. The exception to this is the samples from tern species, which were caught at breeding sites in a different part of Guangdong. In both cases, this leaves parts of the hosts’ ranges unsampled; however, it is unlikely that more expansive sampling would change our conclusion that bird host species does not impact the endosymbiont present in the louse. Above all, the associations between lice and their bacteria appear to be stable over very long time periods, and unlikely to change over the migration or breeding cycle of a single bird. Grossi et al. (2024) examined the endosymbiont of Guimaraesiella mcgrewi infesting four bird host species across five locations in southern China and found all G. mcgrewi harbored the same endosymbiotic strain. While there is evidence for Sodalis in the genus Columbicola to be of recent origin (∼0.4 My from free-living species [Smith et al., 2013]) these are still long-established relationships that do not change seasonally. However, it is possible that other populations of the same bird species breeding and migrating in different parts of the world may be parasitized with lice associated with other bacterial strains. More data from other host flyways are needed to address this.
CRediT authorship contribution statement
Alexandra A. Grossi: Writing – original draft, Methodology, Investigation, Formal analysis. Min Zhang: Writing – review & editing, Resources, Investigation, Funding acquisition. Fasheng Zou: Supervision, Resources, Project administration, Funding acquisition. Daniel R. Gustafsson: Writing – review & editing, Supervision, Resources, Investigation, Funding acquisition, Conceptualization.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
We thank Chi-Yeung Choi from Duke Kunshan University, and Chunpo Tian from Shaanxi Normal University, as well as all our past colleagues for their assistance in collecting the lice used in this study.
Alickovic, L., Johnson, K.P., Boyd, B.M., 2021. The reduced genome of a heritable symbiont from an ectoparasitic feather feeding louse. BMC Ecol. Evol. 21, 108.
Arlott, N., 2017. Birds of South-East Asia. Collins Field Guide. Harper Collins, London.
Brooke, M.D.L., 2010. Vertical transmission of feather lice between adult blackbirds Turdus merula and their nestlings: a lousy perspective. J. Parasitol. 96, 1076–1080.
Clayton, D.H., Tompkins, D.M., 1994. Ectoparasite virulence is linked to mode of transmission. Proc. R. Soc. B 256, 211–217.
Clements, J.F., Schulenberg, T.S., Iliff, M.J., Fredericks, T.A., Gerbracht, J.A., Lepage, D., et al., 2022. The eBird/Clements checklist of Birds of the World: v2022. . (Accessed 2 January 2023).
Dale, C., Maudlin, I., 1999. Sodalis gen. nov. and Sodalis glossinidius sp. nov., a microaerophilic secondary endosymbiont of the tsetse fly Glossina morsitans morsitans. Int. J. Syst. Bacteriol. 49, 267–275.
D’Alessio, L.M., 2023. Cophylogenetics of the Brueelia-Complex (Phthiraptera: Ischnocera) with avian hosts and bacterial endosymbionts. University of Illinois, Urbana-Champaign, Urbana, Illinois. Master’s Thesis.
Danforth, B.N., Ji, S., 1998. Elongation factor-1 alpha occurs as two copies in bees: implications for phylogenetic analysis of EF-1 alpha sequences in insects. Mol. Biol. Evol. 15, 225–235.
de Moya, R.S., Allen, J.M., Sweet, A.D., Walden, K.K.O., Palma, R.L., Smith, V.S., et al., 2019. Extensive host-switching of avian feather lice following the Cretaceous-Paleogene mass extinction event. Commun. Biol. 2, 445.
Douglas, A.E., 2009. The microbial dimension in insect nutritional ecology. Funct. Ecol. 23, 38–47.
Edgar, R.C., 2004. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797.
Fujisawa, T., Barraclough, T.G., 2013. Delimiting species using single-locus data and the generalized mixed yule coalescent approach: a revised method and evaluation on simulated data sets. Syst. Biol. 62, 707–724.
Fukatsu, T., Koga, R., Smith, W.A., Tanaka, K., Nikoh, N., Sasaki-Fukatsu, K., et al., 2007. Bacterial endosymbiont of the slender pigeon louse, Columbicola columbae, allied to endosymbionts of grain weevils and tsetse flies. Appl. Environ. Microbiol. 73, 6660–6668.
Grossi, A.A., Tian, C., Ren, M., Zou, F., Gustafsson, D.R., 2024. Co-phylogeny of a hyper-symbiotic system: endosymbiotic bacteria (Gammaproteobacteria), chewing lice (Insecta: Phthiraptera) and birds (Passeriformes). Mol. Phylogenet. Evol. 190, 107957.
Grossi, A.A., Lee, M.B., Tian, C., Zou, F., Choi, C.Y., Gustafsson, D.R., 2023. Host migration and size do not influence the prevalence of most chewing lice (Phthiraptera: amblycera and Ischnocera) on shorebirds (Aves: Charadriiformes) across the world. Diversity 15, 200.
Gustafsson, D.R., Lei, L., Luo, K., Chu, X., Zhao, X., Zhang, Q., et al., 2019. Chewing lice from high-altitude and migrating birds in Yunnan, China, with descriptions of two new species of Guimaraesiella. Med. Vet. Entomol. 33, 407–419.
Gustafsson, D.R., Olsson, U., 2012. The “very thankless task”: revision of Lunaceps Clay and Meinertzhagen, 1939 (Insecta: Phthiraptera: Ischnocera: Philopteridae), with descriptions of six new species and one new subspecies. Zootaxa 3377, 1–85.
Gustafsson, D.R., Olsson, U., 2017. Unexpected distribution patterns of Carduiceps feather lice (Phthiraptera: Ischnocera: Philopteridae) on sandpipers (Aves: Charadriiformes: Scolopacidae). Syst. Entomol. 42, 509–522.
Hafner, M.S., Sudman, P.D., Villablanca, F.X., Spradling, T.A., Demastes, J.W., Nadler, S.A., 1994. Disparate rates of molecular evolution in cospeciating hosts and parasites. Science 265, 1087–1090.
Hillgarth, N., 1996. Ectoparasite transfer during mating in ringed-necked pheasants Phasianus colchicus. J. Avian Biol. 27, 260–262.
Hopkins, G.H.E., Timmermann, G., 1954. A revision of the species of Quadraceps (Mallophaga) parasitic on Tringinae. Trans. R. Ent. Soc. Lond. 105, 131–150.
Husník, F., Chrudimský, T., Hypša, V., 2011. Multiple origins of endosymbiosis within the Enterobacteriaceae (γ-Proteobacteria): convergence of complex phylogenetic approaches. BMC Biol. 9, 87.
Husnik, F., McCutcheon, J.P., 2016. Repeated replacement of an intrabacterial symbiont in the tripartite nested mealybug symbiosis. Proc. Natl. Acad. Sci. U.S.A. 113, E5416–E5424.
Joseph, V.R., Vakayil, A., 2022. SPlit: an optimal method for data splitting. Technometrics 64, 166–176.
Kumar, S., Stecher, G., Tamura, K., 2021. MEGA11: molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 38, 3022–3027.
Lee, L.I., Tan, D.J.X., Obona, J., Gustafsson, D.R., Ang, Y., Meier, R., 2022. Hitchhiking into the future on a fly: toward a better understanding of phoresy and avian louse evolution (Phthiraptera) by screening bird carcasses for phoretic lice on hippoboscid flies (Diptera). Syst. Entomol. 47, 420–429.
Lee, P.L.M., Clayton, D.H., 1995. Population biology of swift (Apus apus) ectoparasites in relation to host reproductive success. Ecol. Entomol. 20, 43–50.
MacKinnon, J., Phillipps, K., 2000. A Field Guide to the Birds of China. Oxford University Press, Oxford.
Marshall, A., 1981. The Ecology of Ectoparasitic Insects. Academic Press, London.
Michalik, A., Franco, D.C., Kobiałka, M., Szklarzewicz, T., Stroinski, A., Łukasik, P., 2021. Alternative transmission patterns in independently acquired nutritional cosymbionts of Dictyopharidae planthoppers. mBio 12, e0122821.
Nováková, E., Hypša, V., 2007. A new Sodalis lineage from bloodsucking fly Craterina melbae (Diptera, Hippoboscoidea) originated independently of the tsetse flies symbiont Sodalis glossinidius. FEMS Microbiol. Lett. 269, 131–135.
R Core Team, 2018. R: A Language and Environment for statistical computing. Foundation for Statistical Computing. .
Rambaut, A., Drummond, A.J., Xie, D., Baele, G., Suchard, M.A., 2018. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 67, 901–904.
Ries, E., 1931. Die symbiose der läuse und federlinge. Z. Morph. Okol. Tiere 20, 233–367.
Rubin, B.E.R., Sanders, J.G., Turner, K.M., Pierce, N.E., Kocher, S.D., 2018. Social behaviour in bees influences the abundance of Sodalis (Enterobacteriaceae) symbionts. R. Soc. Open Sci. 5, 180369.
Santos-Garcia, D., Silva, F.J., Morin, S., Dettner, K., Kuechler, S.M., 2017. The all-rounder Sodalis: a new bacteriome-associated endosymbiont of the Lygaeoid Bug Henestaris halophilus (Heteroptera: Henestarinae) and a critical examination of its evolution. Genome Biol. Evol. 9, 2893–2910.
Saridaki, A., Bourtzis, K., 2010. Wolbachia: more than just a bug in insects genitals. Curr. Opin. Microbiol. 13, 67–72.
Smith, W.A., Oakeson, K.F., Johnson, K.P., Reed, D.L., Carter, T., Smith, K.L., et al., 2013. Phylogenetic analysis of symbionts in feather-feeding lice of the genus Columbicola: evidence for repeated symbiont replacements. BMC Evol. Biol. 13, 109.
Šochová, E., Husník, F., Nováková, E., Halajian, A., Hypša, V., 2017. Arsenophonus and Sodalis replacements shape evolution of symbiosis in louse flies. PeerJ 5, e4099.
Suchard, M.A., Lemey, P., Baele, G., Ayres, D.L., Drummond, A.J., Rambaut, A., 2018. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 4, vey016.
Sweet, A.D., Browne, D.R., Hernandez, A.G., Johnson, K.P., Cameron, S.L., 2023. Draft genome assemblies of the avian louse Brueelia nebulosa and its associates using long-read sequencing from an individual specimen. G3 Gen. Genom. Genet. 13, jkad030.
Timmermann, G., 1950. Saemundssonia meridiana n. sp., eine bemerkenswerte neue Mallophagenart von Sterna anaethetus Scopoli. Parasitol. News 1, 1–4.
Timmermann, G., 1952. Revision der bei Seeschwalben schmarotzenden Kletterfederlinge der Gattung Quadraceps. Zool. Anz. 148, 71–87.
Timmermann, G., 1954. A revision of the genus Carduiceps Clay & meinertzhagen, 1939 (mallophaga). Ann. Mag. Nat. Hist. 12, 40–48.
Toju, H., Tanabe, A.S., Notsu, Y., Sota, T., Fukatsu, T., 2013. Diversification of endosymbiosis: replacements, co-speciation and promiscuity of bacteriocyte symbionts in weevils. ISME J. 7, 1378–1390.
Wernegreen, J.J., 2015. Endosymbiont evolution: predictions from theory and surprises from genomes. Ann. N. Y. Acad. Sci. 1360, 16–35.
Wilson, A.C.C., Ashton, P.D., Calevro, F., Charles, H., Colella, S., Febvay, G., et al., 2010. Genomic insight into the amino acid relations of the pea aphid, Acyrthosiphon pisum, with its symbiotic bacterium Buchnera aphidicola. Insect Mol. Biol. 19, 249–258.
Shanrui Wu, Ru Jia, Ying Wang, et al. Prevalence, Diversity, and Virulence of Campylobacter Carried by Migratory Birds at Four Major Habitats in China. Pathogens, 2024, 13(3): 230.
DOI:10.3390/pathogens13030230
2.
Zoljargal Purevdorj, Namsrai Jargal, Onolragchaa Ganbold, et al. Spatial and Temporal Variations in Waterfowl Assemblage Structures in Mongolian Lakes and the Changes Linked to the Gradient of Lake Surface Areas. Diversity, 2023, 15(3): 334.
DOI:10.3390/d15030334
3.
Zhongrong Wu, Xiulin Ye, Zhongfan Kuang, et al. Positive Effects of Land Use Change on Wintering Bar-Headed Geese between 2010 and 2021. Animals, 2022, 12(22): 3142.
DOI:10.3390/ani12223142
4.
MARY ANNE BISHOP, DONGPING LIU, GUOGANG ZHANG, et al. Rapid growth of the Bar-headed Goose Anser indicus wintering population in Tibet, China: 1991–2017. Bird Conservation International, 2021.
DOI:10.1017/S0959270921000265
5.
Wen Wang, Fang Wang, Rongkai Hao, et al. First de novo whole genome sequencing and assembly of the bar-headed goose. PeerJ, 2020, 8: e8914.
DOI:10.7717/peerj.8914
6.
Junjian Zhang, Yanbo Xie, Laixing Li, et al. Assessing site-safeguard effectiveness and habitat preferences of Bar-headed Geese (Anser indicus) at their stopover sites within the Qinghai-Tibet Plateau using GPS/GSM telemetry. Avian Research, 2020, 11(1)
DOI:10.1186/s40657-020-00230-9
7.
Ruobing Zheng, Lacy Smith, Diann Prosser, et al. Investigating Home Range, Movement Pattern, and Habitat Selection of Bar-headed Geese during Breeding Season at Qinghai Lake, China. Animals, 2018, 8(10): 182.
DOI:10.3390/ani8100182
Table
2.
Collection and GenBank accession numbers for specimens included in this study.
Louse species
Bird host species
Voucher No.
Loc.
Symbiont (16S) Accession No.
Lice Accession No.
CO1
Ef-1α
Carduiceps meinertzhageni
Calidris alpina
J4366-02
L3
PP081401
PP082907
PP092242
Carduiceps meinertzhageni
Calidris alpina
J4368-02
L3
PP081403
PP082909
PP092244
Carduiceps meinertzhageni
Calidris alpina
J4458-01
L3
PP081411
PP082917
PP092252
Carduiceps meinertzhageni
Calidris alpina
J4471-01
L1
PP081418
PP082924
PP092259
Lunaceps sp.
Calidris pygmaea
J4442-01
L3
PP081406
PP082912
PP092247
Lunaceps drosti
Calidris canutus
J4490-01
L1
PP081423
PP082929
PP092264
Lunaceps limosella
Limosa lapponica
J4509-06
L1
PP081425
PP082931
PP092266
Lunaceps schismatus
Calidris pygmaea
J4437-01
L3
PP081405
PP082911
PP092246
Lunaceps schismatus
Charadrius alexandrinus
J4451-01
L3
PP081407
PP082913
PP092248
Lunaceps schismatus
Charadrius alexandrinus
J4456-01
L3
PP081409
PP082915
PP092250
Lunaceps schismatus
Calidris alpina
J4364-01
L3
PP081398
PP082904
PP092239
Lunaceps schismatus
Calidris alpina
J4366-01
L3
PP081400
PP082906
PP092241
Lunaceps schismatus
Calidris alpina
J4367-01
L3
PP081402
PP082908
PP092243
Lunaceps schismatus
Calidris alpina
J4457-01
L3
PP081410
PP082916
PP092251
Lunaceps schismatus
Calidris alpina
J4460-01
L3
PP081412
PP082918
PP092253
Lunaceps schismatus
Calidris alpina
J4462-01
L3
PP081413
PP082919
PP092254
Lunaceps schismatus
Calidris alpina
J4467-01
L1
PP081414
PP082920
PP092255
Lunaceps schismatus
Calidris alpina
J4469-01
L3
PP081416
PP082922
PP092257
Lunaceps schismatus
Calidris alpina
J4470-01
L1
PP081417
PP082923
PP092258
Lunaceps schismatus
Calidris alpina
J4472-01
L3
PP081419
PP082925
PP092260
Lunaceps schismatus
Calidris alpina
J4455-01
L3
PP081408
PP082914
PP092249
Quadraceps legatus
Onychoprion anaethetus
J4578-01
CI
PP081431
PP082937
PP092272
Quadraceps legatus
Onychoprion anaethetus
J4577-04
CI
PP081430
PP082936
PP092271
Quadraceps legatus
Onychoprion anaethetus
J4588-02
CI
PP081434
PP082940
PP092275
Quadraceps macrocephalus
Charadrius alexandrinus
J4468-01
L1
PP081415
PP082921
PP092256
Quadraceps obscurus
Tringa stagnatilis
J4483-01
L1
PP081421
PP082927
PP092262
Quadraceps obtusus
Tringa totanus
J4489-02
L1
PP081422
PP082928
PP092263
Quadraceps obtusus
Tringa totanus
J4492-02
L3
PP081424
PP082930
PP092265
Quadraceps ptyadis
Charadrius leschenaultii
J4365-01
L3
PP081399
PP082905
PP092240
Quadraceps ptyadis
Charadrius mongolus
J4369-01
L2
PP081404
PP082910
PP092245
Quadraceps ptyadis
Charadrius mongolus
J4479-01
L1
PP081420
PP082926
PP092261
Quadraceps sellatus
Thalasseus bergii
J4585-02
CI
PP081433
PP082939
PP092274
Saemundssonia laticaudata
Thalasseus bergii
J4583-02
CI
PP081432
PP082938
PP092273
Saemundssonia meridiana
Onychoprion anaethetus
J4576-01
CI
PP081426
PP082932
PP092267
Saemundssonia meridiana
Onychoprion anaethetus
J4576-02
CI
PP081427
PP082933
PP092268
Saemundssonia meridiana
Onychoprion anaethetus
J4577-01
CI
PP081428
PP082934
PP092269
Saemundssonia meridiana
Onychoprion anaethetus
J4577-02
CI
PP081429
PP082935
PP092270
Voucher number, the first four digits refer to the host individual and the last two digits refer to a unique louse. Thus, J4366–01 and J4366-02 are different lice from the same host individual. Location abbreviation: CI – Chiyu Island, Shantou; L1 – Dongli, Leizhou; L2 – Hebei, Leizhou; L3 – Tujiao, Leizhou, Guangdong, China.

 DownLoad:
DownLoad:






 Email Alerts
Email Alerts RSS Feeds
RSS Feeds