Yingqiang Lou, Lijun Chen, Qingshan Zhao, Anders Pape Møller, Yun Fang, Yuehua Sun. 2022: Sexual dichromatism and assortative mating by multiple plumage color traits in wild Chestnut Thrush. Avian Research, 13(1): 100033. DOI: 10.1016/j.avrs.2022.100033
Citation:
Yingqiang Lou, Lijun Chen, Qingshan Zhao, Anders Pape Møller, Yun Fang, Yuehua Sun. 2022: Sexual dichromatism and assortative mating by multiple plumage color traits in wild Chestnut Thrush. Avian Research, 13(1): 100033. DOI: 10.1016/j.avrs.2022.100033
Yingqiang Lou, Lijun Chen, Qingshan Zhao, Anders Pape Møller, Yun Fang, Yuehua Sun. 2022: Sexual dichromatism and assortative mating by multiple plumage color traits in wild Chestnut Thrush. Avian Research, 13(1): 100033. DOI: 10.1016/j.avrs.2022.100033
Citation:
Yingqiang Lou, Lijun Chen, Qingshan Zhao, Anders Pape Møller, Yun Fang, Yuehua Sun. 2022: Sexual dichromatism and assortative mating by multiple plumage color traits in wild Chestnut Thrush. Avian Research, 13(1): 100033. DOI: 10.1016/j.avrs.2022.100033
Key Laboratory of Animal Ecology and Conservation Biology, Institute of Zoology, Chinese Academy of Sciences, Beijing, 100101, China
b.
State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing, 100085, China
Sex differences in plumage color are common in bird species. Some bird species are regarded as sexually monochromatic in human visual systems, and in recent years, some species are found to be of cryptic (to human) sexual dichromatism by spectrophotometric techniques. However, the functions of plumage color are still less understood in these species. Here, we focused on plumage color traits in the Chestnut Thrush (Turdus rubrocanus), which is considered as a sexually monochromatic bird by human observers. We used spectrometer analyses and avian visual modeling to investigate the color traits of males and females, and whether these color traits are involved in assortative mating. We found that Chestnut Thrush showed sexual dichromatism in bill, throat and wing, and pairs mated assortatively with colorations of throat, chest, crown and wing. We also found that color of tarsus was different between two consecutive years. These results revealed that Chestnut Thrush is sexually dichromatic in the avian visual system, and plumage color traits play important roles in mate choice.
Sexual dichromatism is defined as the coloration of males and females being different in the same species (Dale et al., 2015), which is often generated by structural, carotenoid- and melanin-based coloration (Shawkey and Hill, 2005; Hubbard et al., 2010). Previous studies showed that bird species can perceive a greater diversity of color than humans (Goldsmith, 1994), such as ultraviolet (UV), and then the plumage colors of birds should be different between humans and bird species. With the development of avian visual models and reflectance spectrophotometry, it is easier to distinguish the sexual dichromatism in bird species which are monochromatic to humans (Håstad and Ödeen, 2008; Dale et al., 2015). By using this method, Burns and Shultz (2012) found that 152 of 163 monochromatic species by human's visual systems are dichromatic for the avian visual system.
Plumage color plays an important role in sexual selection in birds (Hill and McGraw, 2006), these conspicuous colorations reflecting the quality of individuals, which may enhance mating opportunities or mating success (Andersson, 1994; Dale et al., 2015). Assortative mating is one of the most commonly mating patterns in animals (Macdougall and Montgomerie, 2003). It could occur when males and females with preferred certain traits choose each other (Kraaijeveld et al., 2007), such as body size, body condition, social status and plumage color traits (review in Jiang et al., 2013), and then leaving less preferred individuals to mate with others (Jiang et al., 2013). For example, the Northern Cardinals (Cardinalis cardinalis) mate assortatively by red plumage and bill color (Jawor et al., 2003). However, most studies of plumage color traits have focused on species with obvious sexual dichromatism from human visual system, with less attention being shown to species with cryptic sexual dichromatism (Clutton-Brock, 2009; Burns and Shultz, 2012). Therefore, the roles of plumage color of species with cryptic sexual dichromatism in mate choice are still unclear (Rull et al., 2016).
In many species, plumage coloration can also be related to age (Dreiss and Roulin, 2010; Edler and Friedl, 2010). Older individuals may have more experience in foraging, allowing them to be in better condition during the moulting period (Doucet and Montgomerie, 2003), and they can have superior plumage color. However, age-related differences in plumage color are not consistent among species. For example, Bitton et al. (2008) found that older male Tree Swallows (Tachycineta bicolor) are brighter and more blue than younger individuals. The Eastern Bluebird (Sialia sialis) did not change the hue of plumage color with age (Siefferman and Hill, 2005).
The Chestnut Thrush (Turdus rubrocanus) is a medium sized passerine with biparental care, and it is widespread and locally common in Southwest China (Zheng, 2017). This bird was considered as sexually monomorphic from human visual system (Zhao, 2001), and both males and females exhibit only melanin-based plumage traits. Here, we used spectrophotometry to measure plumage reflectance in wild Chestnut Thrushes based on avian visual system. Our aims of this study were three-fold: first, we investigated whether the Chestnut Thrush is sexually dichromatic from avian visual system; second, we investigated whether Chestnut Thrush mate assortatively with respect to coloration traits; third, we investigated whether the coloration traits vary with age.
2.
Materials and methods
2.1
Study area
We conducted the field work at Badu village (2200–2400 m above sea level) of Lianhuashan Nature Reserve, Gansu Province, China (34°54′17″–35°01′43″ N, 103°39′59″–103°50′26″ E), from April to August 2015–2016 and 2019–2020. The study site is mainly covered by agricultural land, woodland and bushes. More detailed information can be found in Sun et al. (2003).
2.2
Data collection
We used mist nets to capture adult Chestnut Thrushes during the breeding period, and each individual was marked by a mental and some color bands. The sex was determined by the presence or absence of a brood patch (only females incubate the eggs in this species) (Lou et al., 2021). Twenty-seven individuals were caught before incubation period whose sex could not be distinguished in the field, then we drew blood from the vein in the wing, and stored them in anhydrous alcohol. Polymerase chain reaction (PCR) was used to amplify part of the chromo-helicase-DNA binding protein (CHD) gene by the primers IntP2/IntP8 in the lab. The identities of mating pairs were confirmed by the video of provisioning behaviors during nestling provisioning.
2.3
Color measurement
We collected plumage reflectance data of the Chestnut Thrush using the spectrometer (AvaSpec-ULS2048L-USB2, Avantes Inc., Netherlands), with a AvaLight-DHc (Combined deuterium-halogen) light source and UV-VIS fibre-optic reflectance probe (FCR-7UV400-2-ME). To standardize measuring distance (3 mm) and keep out the ambient light, a black metal block was fixed on the probe. All measurements were made relative to a white WS-2 reference tile (Avantes BV, Eerbeek, Netherlands) and were dark-corrected. A black cloth was used as background to reduce the influence of the reflection curve by other objects (Stoddard and Prum, 2008). The spectrometer was set to an integration time of 625.91 ms. The diameter of the measured circular region was 6.5 mm, and the distance between probe and plumage was 3 mm. Each reading was made at an angle of 90° to the surface of the plumage. All reflectance curves were stored by Avasoft 8.1.4 software. Reflectances of captured Chestnut Thrushes were measured at eight patches: two carotenoid-based (bill and tarsus) and six melanin-based plumage (crown, back, tail, chest, throat and wing). We obtained five points per patch from each bird, and we used the average of these five curves for each individual to analyze the reflectance in the avian visible spectrum (300–700 nm).
2.4
Visual modeling
We chose three plumage color traits: hue, chroma and brightness for each patch. Hue was calculated as the wavelength of median reflectance between 300 and 700 nm (Rowe and Weatherhead, 2011). Chroma was calculated by maximum-minimum reflectance/mean brightness (Andersson et al., 2002). Brightness was the mean relative reflectance over the entire spectral range measured. Low values of brightness represent dark colors.
We applied the Starling (Sturnus vulgaris) visual system to calculate the full-spectrum (300–700 nm) reflectance curves through models of avian vision. We used the 'pavo' package in the R software (Maia et al., 2013; R Core Team, 2015), which used tetrahedral color space models to plot reflectance spectra (Stoddard and Prum, 2008). We also set the models assuming standardized daylight illumination (D65) for open habitat ambient light (Wyszecki and Stiles, 2000), which is similar to the habitat of Chestnut Thrush.
2.5
Statistical analysis
Each patch was combined in principal component analyses with varimax rotation. We kept the first component extracted with eigenvalues > 1, and most of them explained > 50% of the variation (Table 1). We used the factor scores from PCA (hereafter PC1 score) for each individual in the further analyses. For the crown and tail, high PC1 values correspond to high chroma and brightness. For the throat, high PC1 values correspond to high brightness. For chest and back, high PC1 values correspond to high hue and low hue, respectively. For the wing, high PC1 value correspond to low chroma and brightness. For the bill, high PC1 values correspond to low chroma and high brightness. For the tarsus, PC1 values correspond to high chroma and low brightness.
Table
1.
Principal component analysis (PCA) of color for each plumage patch.
One-way ANOVA was used to compare the color variables between males and females. To test whether the plumage color traits of each patch were assortative within pairs, Pearson correlation test and Spearman correlation test were used to analyze the normally distributed data and non-normal distributions, respectively. Paired t-test was used to analyze whether plumage color traits of the same individuals changed between the first time caught and the subsequent year. All statistical analyses were conducted in R (R Core Team, 2015).
3.
Results
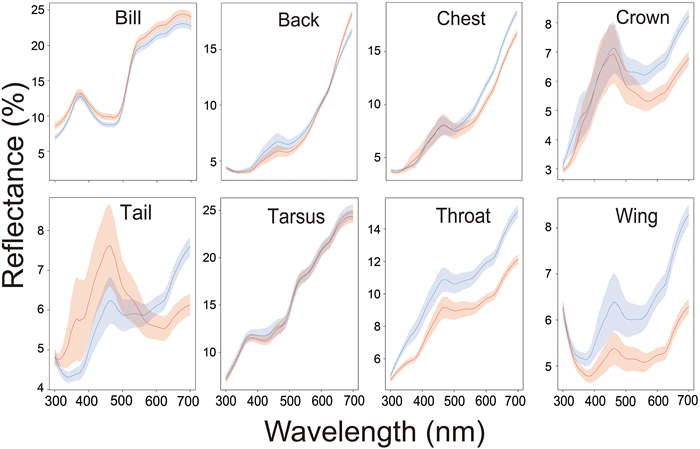
In total, 323 Chestnut Thrushes (154 males and 169 females) were measured from 2014 to 2015 and 2019–2020 in our study. The comparisons of reflectance spectra between males and females for 8 body patches were shown in Fig. 1. There were no significant differences in PC1 score of back, crown, chest, tail and tarsus (Table 2). We found sexual differences in PC1 score of throat, wing and bill (Table 2): males were brighter in throat (higher in PC1 score), less chromatic and darker in wing (higher in PC1 score), and more chromatic and darker in bill (lower in PC1 score).
Figure
1.
Reflectance spectra (300–700 nm) of Chestnut Thrushes in eight plumage color patches. The blue lines and areas represent females, and the pink lines and areas represent males. Each spectrum represents the mean reflectance ± SE.
Table
2.
Sex differences in color patches of Chestnut Thrushes.
Color patch
Males
Females
F
P
Back (PC)
−0.119 ± 1.341
0.110 ± 1.225
1.780
0.183
Crown (PC)
−0.128 ± 1.366
0.119 ± 1.226
2.935
0.087
Throat (PC)
0.334 ± 1.864
−0.310 ± 1.488
11.750
< 0.001
Chest (PC)
0.100 ± 1.381
−0.093 ± 1.246
1.880
0.171
Wing (PC)
0.231 ± 0.994
−0.215 ± 1.415
10.200
0.002
Tail (PC)
−0.053 ± 1.594
0.049 ± 0.930
0.591
0.443
Bill (PC)
−0.239 ± 1.450
0.222 ± 0.979
11.970
0.001
Tarsus (PC)
−0.049 ± 1.135
0.045 ± 1.256
0.271
0.603
The values are mean ± SD. The first principal components (PC) from the principal component analysis of each color patch are shown in the table. Significant results are shown in bold.
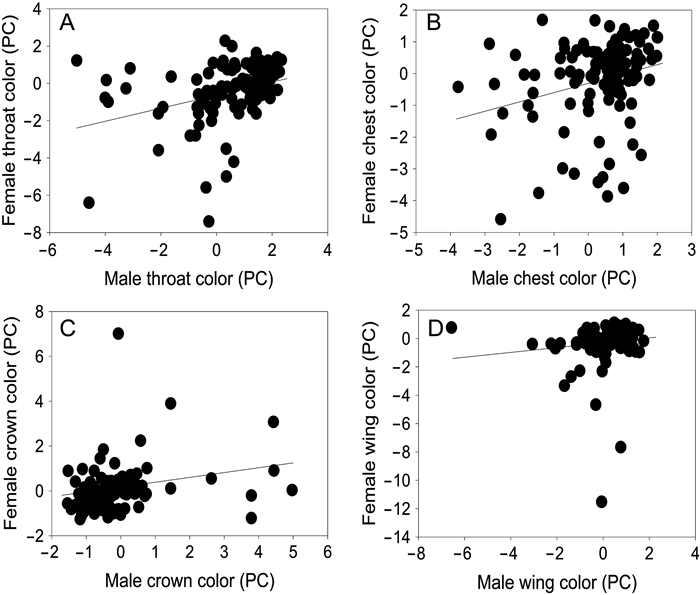
In total we measured color traits of 114 pairs, and Chestnut Thrush pairs were positive assortative mating based on plumage color of throat (r = 0.427, P < 0.001; Fig. 2A), chest (r = 0.295, P = 0.002; Fig. 2B), crown (r = 0.337, P < 0.001; Fig. 2C), and wing (r = 0.276, P = 0.003; Fig. 2D), but not in other color traits (Table 3).
Figure
2.
Assortativemating by plumage color in Chestnut Thrushes. Each black circle represents a pair and the solid line indicates the equal expression. Each color was analyzed as the first principal component (PC) from principal component analyses.
As to 15 individuals (8 males and 7 females) which were measured in two consecutive years, we found that the second-year individuals had lower PC1 scores of tarsus than the first-year individuals: second-year individuals were less chromatic and brighter in tarsus (Table 4).
Table
4.
Differences in plumage color of the same individuals in two continuous years.
Color patch
First year
Second year
t
P
Back (PC)
−0.170 ± 1.077
−0.429 ± 0.826
0.926
0.370
Crown (PC)
−0.193 ± 0.676
0.025 ± 0.695
−1.052
0.311
Throat (PC)
0.214 ± 1.042
0.006 ± 1.311
0.732
0.476
Chest (PC)
0.118 ± 1.164
0.150 ± 1.234
−0.118
0.908
Wing (PC)
0.115 ± 0.855
0.065 ± 0.995
0.202
0.843
Tail (PC)
−0.041 ± 0.525
0.261 ± 1.124
−1.235
0.237
Bill (PC)
0.252 ± 0.556
0.339 ± 0.500
−1.040
0.316
Tarsus (PC)
0.875 ± 0.694
−0.194 ± 1.445
2.280
0.039
The values are mean ± SD. The first principal components (PC) from the principal component analysis of each color patch are shown in the table. Significant results are shown in bold.
Our study found avian-perceivable sexual dichromatism in Chestnut Thrushes. Chestnut Thrush pairs showed assortative mating by multiple plumage patches (throat, chest, crown and wing), suggesting that these patches may function as sexual signals in both sexes. We also found the color trait of tarsus changed across years, but not in other plumage color traits.
We found that Chestnut Thrushes were sexually dichromatic from avian visual system: males had brighter throat, darker wing and bill, less chromatic wing and more chromatic bill than females. Similar results were also found in Whiteheads (Mohoua albicilla), i.e., males had brighter head and chest feather than females (Igic et al., 2010). Differences between males and females may be affected by natural selection and sexual selection (Baker and Parker, 1979; Dunn et al., 2015). For example, female plumage color traits were negatively associated with nest predation (Martin and Badyaev, 1996; Ekanayake et al., 2015), and females with less chromatic and brighter plumage color may reduce the nest predation. However, the exact causes of sexual dichromatism in Chestnut Thrushes require further investigations.
The Chestnut Thrush paired assortatively by multiple plumage traits. Similar results were also found in other species (Jacobs et al., 2015; Rull et al., 2016; Zwaan et al., 2019), for example, American Robins (Turdus migratorius) chose mates based on the brightness of chest (Rowe and Weatherhead, 2011). Assortative mating can result from mutual mate choice (Jiang et al., 2013), and mutual selection is likely to arise in species which have similar reproductive roles in the two sexes (Clutton-Brock, 2009). In the Chestnut Thrush, both parents participate in parental care, such as nest defense and offspring provisioning. Furthermore, the population of Chestnut Thrush is relatively large in our study area and they lack breeding territoriality. In this situation, males and females may encounter other potential mates, resulting in mutual mate choice from plumage color.
We found the color variable of tarsus changed between two years, and this result may indicate that tarsus varies with age. However, the tarsus is carotenoid-based, which could also be affected by carotenoid level in food resources (George et al., 2017), environmental toxins (Spickler et al., 2020) and social status (Karubian et al., 2011). Therefore, to fully explore this question, experiments with hand raised birds are needed. We failed to find any variations of melanin-based plumage between years, which was not in line with McKinnon and Robertson (2008). One possible reason may explain our results: melanin-based plumage color traits were less sensitive to diet and environmental factors (reviewed in Roulin, 2004; Guindre-Parker and Love, 2014), and they may not differ significantly between the two years' data. Further studies on Chestnut Thrushes would be necessary to test the relationship between melanin-based plumage and age.
5.
Conclusions
Our results showed the basic information on color traits and revealed the sex-dependent color traits in the Chestnut Thrush. We also found that assortative mating pattern based on multiple plumage color traits existed in cryptically dichromatic species. Further studies should focus on the roles of plumage colorations in signalling, and investigate how they influence reproductive fitness.
Ethics statement
The present study complies with the current laws of China and it was approved by the Animal Care and Use Committee of the Institute of Zoology, Chinese Academy of Sciences (Permission No. 2013/108). Birds were caught only during days without rain and with low wind speed during the capture. Birds were trapped and released into the wild near the trapping locations. No adults died during the experimental periods.
Author contributions
YS conceived and designed the study. YL, LC, QZ, YF conducted the experiments in the field. YL and LC analyzed the data and wrote the manuscript. APM helped in the manuscript preparation. All authors read and approved the final manuscript.
Declaration of competing interest
The authors declare that they have no competing interests.
Acknowledgements
We thank Chunlei Jing, Yumeng Tian and Huan Liu for their assistance in the field. We thank the anonymous reviewers for comments on the manuscript. This research was supported by the National Natural Science Foundation of China (No. 32070452to YS).
Andersson, M., 1994. Sexual Selection. Princeton University Press, Princeton
Andersson, S., Pryke, S.R., Ornborg, J., Lawes, M.J., Lawes, M.J., Andersson, M., 2002. Multiple receivers, multiple ornaments, and a trade-off between agonistic and epigamic signaling in a widowbird. Am. Nat. 160, 683-691
Baker, R.R., Parker, G., 1979. The evolution of bird coloration. Phil. Trans. R. Soc. Lond. B. 287, 63-130
Bitton, P.P., Dawson, R.D., Ochs, C.L., 2008. Plumage characteristics, reproductive investment and assortative mating in tree swallows Tachycineta bicolor. Behav. Ecol. Sociobiol. 62, 1543-1550
Burns, K.J., Shultz, A.J., 2012. Widespread cryptic dichromatism and ultraviolet reflectance in the largest radiation of Neotropical Songbirds: implications of accounting for avian vision in the study of plumage evolution. Auk 3, 211-221
Clutton-Brock, T., 2009. Sexual selection in females. Anim. Behav. 77, 3-11
Dale, J., Dey, C.J., Delhey, K., Kempenaers, B., Valcu, M., 2015. The effects of life history and sexual selection on male and female plumage colouration. Nature 527, 367-370
Doucet, S.M., Montgomerie, R., 2003. Multiple sexual ornaments in satin bowerbirds: ultraviolet plumage and bowers signal different aspects of male quality. Behav. Ecol. 14, 503-509
Dreiss, A.N., Roulin, A., 2010. Age-related change in melanin-based coloration of Barn Owls (Tyto alba): females that become more female-like and males that become more male-like perform better. Biol. Linn. Soc. 101, 689-704
Dunn, P.O., Armenta, J.K., Whittingham, L.A., 2015. Natural and sexual selection act on different axes of variation in avian plumage color. Sci. Adv. 1, e1400155
Edler, A.U., Friedl, T., 2010. Plumage colouration, age, testosterone and dominance in male Red Bishops (Euplectes orix): a laboratory experiment. Ethology 116, 806-820
Ekanayake, K.B., Whisson, D.A., Tan, L.X., Weston, M.A., 2015. Intense predation of non-colonial, ground-nesting bird eggs by corvid and mammalian predators. Wildlife Res. 42, 518-528
George, D.B., Schneider, B.C., McGraw, K.J., Ardia, D.R., 2017. Carotenoids buffer the acute phase response on fever, sickness behavior and rapid bill color change in zebra finches. J. Exp. Biol. 220, 2957-2964
Goldsmith, T.H., 1994. The perception of color. In: Gouras, P. (Ed.), Vision and Visual Dysfunction. Macmillan, London, pp. 62-89
Guindre-Parker, S., Love, O.P., 2014. Revisiting the condition-dependence of melanin-based plumage. J. Avian Biol. 45, 29-33
Hastad, O., Odeen, A., 2008. Different ranking of avian colors predicted by modeling of retinal function in humans and birds. Am. Nat. 171, 831-838
Hill, G.E., McGraw, K.J., 2006. Bird Coloration, Volume 1: Mechanisms and Measurements. Harvard University Press, Cambridge
Igic, B., Leuschner, N., Parker, K.A., Ismar, S.M.H., Gill, B.J., Lovegrove, T.G., et al., 2010. Size dimorphism and avian-perceived sexual dichromatism in a New Zealand endemic bird, the Whitehead Mohoua albicilla. J. Morphol. 271, 697-704
Jacobs, A.C., Fair, J.M., Zuk, M., Ebensperger, L., 2015. Coloration, paternity, and assortative mating in western bluebirds. Ethology 121, 176-186
Jawor, J.M., Linville, S.U., Beall, S.M., Breitwisch, R., 2003. Assortative mating by multiple ornaments in northern cardinals (Cardinalis cardinalis). Behav. Ecol. 14, 515-520
Jiang, Y., Bolnick, D.I., Kirkpatrick, M., 2013. Assortative mating in animals. Am. Nat. 181, E125-E138
Karubian, J., Lindsay, W.R., Schwabl, H., Webster, M.S., 2011. Bill coloration, a flexible signal in a tropical passerine bird, is regulated by social environment and androgens. Anim. Behav. 81, 795-800
Kraaijeveld, K., Kraaijeveld-Smit, F.J.L., Komdeur, J., 2007. The evolution of mutual ornamentation. Anim. Behav. 74, 657-677
Lou, Y.Q., Chen, L.J., Hu, Y.B., Komdeur, J., Sun, Y.H., Lyu, N., 2021. Animal personality can modulate sexual conflict over offspring provisioning. Anim. Behav. 177, 91–97
Macdougall, A.K., Montgomerie, R., 2003. Assortative mating by carotenoid-based plumage colour: a quality indicator in American goldfinches, Carduelis tristis. Naturwissenschaften 90, 464-467
Maia, R., Eliason, C.M., Bitton, P.P., 2013. pavo: an R package for the analysis, visualization and organization of spectral data. Method. Ecol. Evol. 4, 906-913
Martin, T.E., Badyaev, A.V., 1996. Sexual dichromatism in birds: importance of nest predation and nest location for females versus males. Evolution 50, 2454-2460
McKinnon, E.A., Robertson, R.J., 2008. The signal function of a melanin-based plumage ornament in golden-winged warblers. Wilson J. Ornithol. 120, 366–370
R Development Core Team, 2013. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Roulin, A., 2004. The evolution, maintenance and adaptive function of genetic colour polymorphism in bird. Biol. Rev. 79, 815-848
Rull, I.L., Nicolas, LNicolas, L., Neri-Vera, N., Argaez, V., Martinez, M., Torres, R., 2016. Assortative mating by multiple skin color traits in a seabird with cryptic sexual dichromatism. J. Ornithol. 157, 1049-1062
Rowe, K.M.C., Weatherhead, P.J., 2011. Assortative mating in relation to plumage traits shared by male and female American Robins. Condor 2011, 881-889
Shawkey, M.D., Hill, G.E., 2005. Caroteniods need structural colours to shine. Biol. Lett. 1, 121-124
Siefferman, L., Hill, G.E., 2005. Blue structural coloration of male eastern bluebirds Sialia sialis predicts incubation provisioning to females. J. Avian Biol. 36, 488-493
Spickler, J.L., Swaddle, J.P., Gilson, R.L., Varian-Ramos, C.W., Cristol, D.A., 2020. Sexually selected traits as bioindicators: exposure to mercury affects carotenoid-based male bill color in zebra finches. Ecotoxicology 29, 1138-1147
Stoddard, M.C., Prum, R.O., 2008. Evolution of avian plumage color in a tetrahedral color space: a phylogenetic analysis of new world buntings. Am. Nat. 171, 755-776
Sun, Y.H., Swenson, J.E., Fang, Y., Klaus, S., Scherzinger, W., 2003. Population ecology of the Chinese grouse, Bonasa sewerzowi, in a fragmented landscape. Biol. Conserv. 110, 177-184
Wyszecki, G., Stiles, W.S., 2000. Color Science, Concepts and Methods. Quantitative Data and Formulas. 2nd edn. Wiley, New York, NY
Zhao, Z., 2001. A Handbook of the Birds of China: Vol. 2 Passerines. Jilin Science and Technology Press, Changchun
Zheng, G., 2017. A Checklist on the Classification and Distribution of the Birds of China. third ed. Science Press, Beijing
Zwaan, D.R.D., Barnes, S., Martin, K., 2019. Plumage melanism is linked to male quality, female parental investment and assortative mating in an alpine songbird. Anim. Behav. 156, 41-49
Table
2.
Sex differences in color patches of Chestnut Thrushes.
Color patch
Males
Females
F
P
Back (PC)
−0.119 ± 1.341
0.110 ± 1.225
1.780
0.183
Crown (PC)
−0.128 ± 1.366
0.119 ± 1.226
2.935
0.087
Throat (PC)
0.334 ± 1.864
−0.310 ± 1.488
11.750
< 0.001
Chest (PC)
0.100 ± 1.381
−0.093 ± 1.246
1.880
0.171
Wing (PC)
0.231 ± 0.994
−0.215 ± 1.415
10.200
0.002
Tail (PC)
−0.053 ± 1.594
0.049 ± 0.930
0.591
0.443
Bill (PC)
−0.239 ± 1.450
0.222 ± 0.979
11.970
0.001
Tarsus (PC)
−0.049 ± 1.135
0.045 ± 1.256
0.271
0.603
The values are mean ± SD. The first principal components (PC) from the principal component analysis of each color patch are shown in the table. Significant results are shown in bold.
Table
4.
Differences in plumage color of the same individuals in two continuous years.
Color patch
First year
Second year
t
P
Back (PC)
−0.170 ± 1.077
−0.429 ± 0.826
0.926
0.370
Crown (PC)
−0.193 ± 0.676
0.025 ± 0.695
−1.052
0.311
Throat (PC)
0.214 ± 1.042
0.006 ± 1.311
0.732
0.476
Chest (PC)
0.118 ± 1.164
0.150 ± 1.234
−0.118
0.908
Wing (PC)
0.115 ± 0.855
0.065 ± 0.995
0.202
0.843
Tail (PC)
−0.041 ± 0.525
0.261 ± 1.124
−1.235
0.237
Bill (PC)
0.252 ± 0.556
0.339 ± 0.500
−1.040
0.316
Tarsus (PC)
0.875 ± 0.694
−0.194 ± 1.445
2.280
0.039
The values are mean ± SD. The first principal components (PC) from the principal component analysis of each color patch are shown in the table. Significant results are shown in bold.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds