The spatial distribution and breeding behavior of neighboring birds (conspecific and heterospecific) may influence reproduction and the effects differ across species. In this study, we investigated intra- and inter-specific spatial distribution of breeding via artificial nestboxes and examined whether overlap with neighbors affected reproductive success of the focal breeding pair. Regarding spatial distribution, Varied Tits (Sittiparus varius) and Cinereous Tits (Parus cinereus) showed similar pattern: distances of nestboxes containing conspecific tits were greater than those with heterospecific ones. In terms of breeding behavior, reproductive success was not significantly related to the distance from their neighbor’s nest. The reproductive success of Varied Tits was significantly related to that of their neighbors, while in Cinereous Tits, no strong link was found to any recorded reproductive behavior of the neighbors. These findings explain spatial variation in nest site selection of Varied Tits and Cinereous Tits. Reproductive success rate of Varied Tits is affected by that of the neighbors while that of Cinereous Tits is not. The results highlight that there are differences in reproductive outcome among related species and offer suggestions on the use of artificial nestboxes in experiment and conservation programs.
There often is competition for limited resources when species live in the same area, especially in the breeding season. Individuals need suitable nest sites (Martin, 1993) and sufficient food resources for survival and reproduction (Martin, 1987). They also want to avoid losses caused by interference competition (Minot and Perrins, 1986; review in Dhondt, 2012).
A large number of experimental studies have shown that intra- and inter-specific competition can reduce breeding success and population size (Schoener, 1983; Finch, 1990; Gurevitch et al., 1992). House Wrens (Troglodytes aedon) that co-habit with Tree Swallows (Tachycineta bicolor) have lower reproductive success rate than wrens living in swallow-free plots (Finch, 1990). The first egg laying date of Eurasian Blue Tits (Cyanistes caeruleus) is affected by intra-specific and inter-specific interference, but no such relationship has been found in a closely related species, the Great Tit (Parus major, Møller et al., 2018). The reproductive output of the central nest is adversely affected by their short-distance neighbors containing conspecifics but not heterospecific ones (Deeming et al., 2017). Most studies have concentrated on the breeding behavior of conspecific or heterospecific neighbors to evaluate intra- and inter-specific competition. Breeding synchrony may also be an important environmental indicator as needs for resources vary hugely as nestlings grow. Competition might lead greater intervals among neighbors in terms of delay in breeding and might have effects on reproduction in neighbors.
Some species are sufficiently abundant and show no obvious spatial heterogeneity, while others are range-restricted and influenced by both habitat composition and local environment (Laube, 2011). Regarding the question of what affects the size of the distribution, most studies concentrated on non-behavioral environmental or spatial factors (e.g. Szabo et al., 2009; Boucher-Lalonde et al., 2014) but few have investigated behaviors. Zhang et al. (2019) attributed the difference in the distribution size of Cinereous (Parus cinereus) and Varied Tits (Sittiparus varius) to their personalities because the relatively wider-spread Cinereous Tit is more exploratory, active, and risk-prone than the Varied Tit, a locally distributed species, is. We predict that anti-interference capability of the narrowly-distributed Varied Tit may be weaker than that of the relatively widely-distributed Cinereous Tit.
Artificial nestboxes are widely used in studies of birds, particularly in field research, because they attract breeding birds. As a result, it is an effective tool to gather reproduction data (Møller et al., 2014) to study demographic shift, life history evolution, quantitative genetics, and sexual selection (Evans et al., 2002). To ensure sufficiency, usually large numbers of artificial nestboxes are provided, but only a small number are occupied every year (Mänd et al., 2005).
In this study, we measured intra- and inter-specific spatial distribution of used artificial nestboxes and investigated whether the intersection of neighbors’ living area affected the breeding outcome of the focal breeding pair. Two hole-nesting passerine species, Cinereous Tit and Varied Tit, were concentrated on. Based on which we proposed three hypotheses: (1) the distance of nests within species is longer than that between them, (2) conspecific and heterospecific neighbors both have a negative impact on the focal nest, and (3) the range-restricted species, Varied Tits, are more vulnerable to neighbor influence than the relatively wider-spread Cinereous Tits.
2.
Materials and methods
2.1
Study area and data collection
Starting from 2016, this study was carried out every year from early April through mid-July for three consecutive years in Liaoning Xianrendong National Nature Reserve (39°54ʹ‒40°03ʹ N, 122°53ʹ‒123°03ʹ E). Yet ten years before this study began artificial nestboxes had already been installed and supplemented or replaced every year. All boxes had a base dimension of 15 cm × 15 cm and a height of 30 cm. The opening was round, with a diameter of 3 cm or 3.5 cm, and located about 7 cm below the box cover. The artificial nestboxes were hung mainly on Pinus densiflora, Quercus mongolica and Pinus koraiensis, at a height of 2–3 m from the ground.
All boxes were hung randomly on both sides along infrequent paths in the reserve. Their locations were recorded using OvitalMap (Beijing Ovital Software Co. Ltd. 2016). The average distance between two nearest ones was around 30 m. Weekly recording included the laying date, the species (available since incubation) and reproductive parameters such as clutch size, the number of hatchlings, ten-day-old nestlings and fledglings. We then calculated the hatching rate (the number of hatchlings/clutch size) and reproductive success rate (the number of fledglings/clutch size). The distances from the central nestbox to (1) the nearest ones, (2) the nearest nestbox occupied by a conspecific, and (3) the nearest nestbox occupied by a heterospecific were all measured using OvitalMap.
2.2
Statistical analyses
The data was analyzed in SPSS version 21.0 (IBMCorp. Armonk, NY). An index of dispersion (Fowler et al., 1997) was calculated to ascertain the box distribution pattern. Mixed effect regression was used to test factors (the explanatory variable: neighbor type; and the random factors: box number and year) that affected the intra- and inter-specific nest spacing (the response variables: the distance between the focal nest and its nearest conspecific/heterospecific nest). We used mixed effect models to explore the influence of spatial distribution on the hatching rate of the focal nest (the explanatory variables: conspecific distance, heterospecific distance; and the random factors: box number and year) and linear models to explore the influence of spatial distribution on the reproductive success rate of the focal nest (the explanatory variables: conspecific distance, heterospecific distance). We used mixed effect models to explore the influence of breeding behavior on the hatching rate of the focal nest (the explanatory variables: neighbor type, neighbor hatching rate, and breeding synchrony; and the random factors: box number and year) and linear models to explore the influence of breeding behavior on the reproductive success rate of the focal nest (the explanatory variables: neighbor type, neighbor reproductive success rate and breeding synchrony). All analyses were considered significant at a probability value of <0.05.
3.
Results
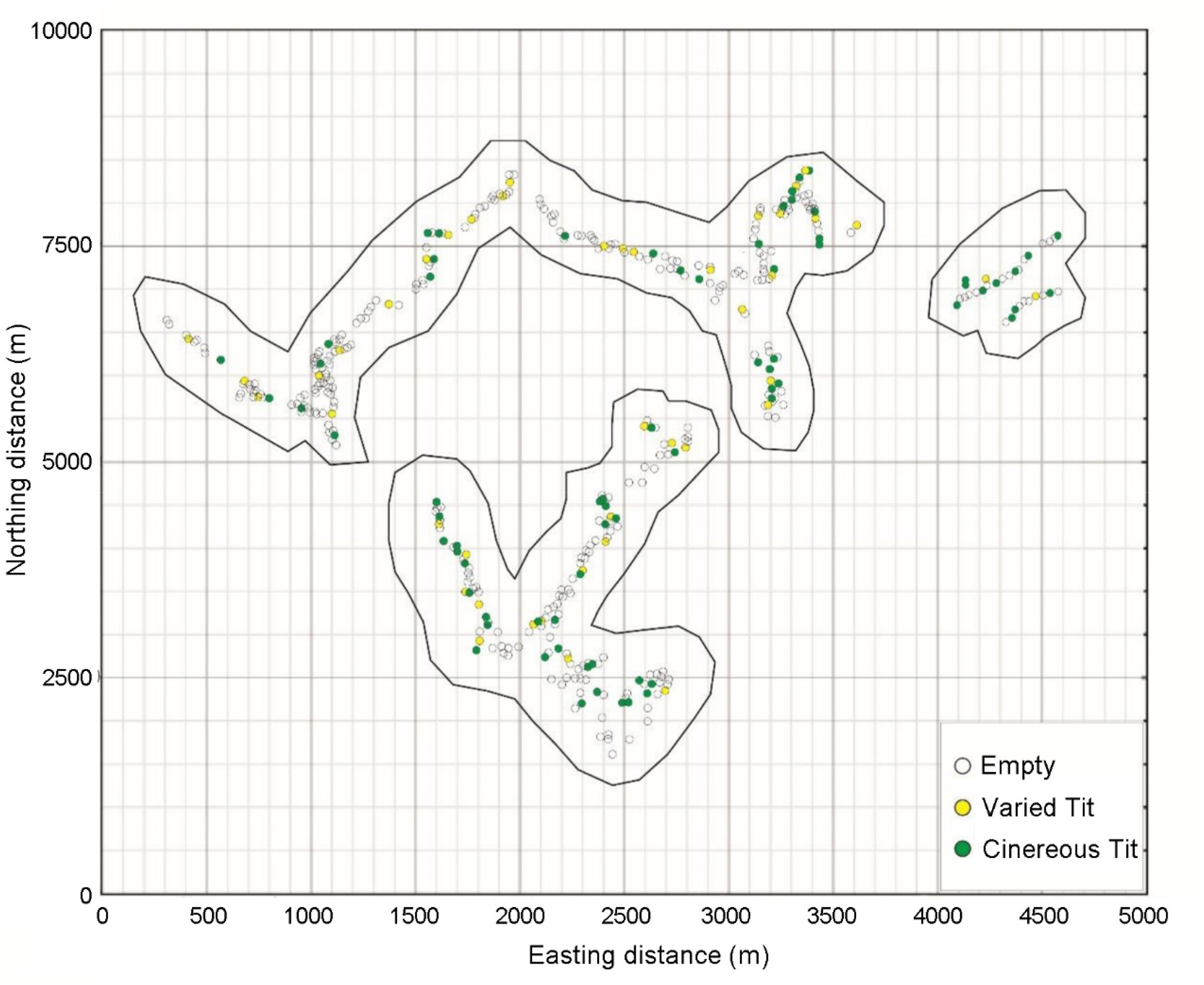
In total, 340, 451 and 434 nestboxes were investigated in 2016, 2017 and 2018, respectively. Each year less than half were occupied (Table 1) mainly by Cinereous Tits and Varied Tits, while few were occupied by other species such as Marsh Tits (Poecile palustris) and Daurian Redstarts (Phoenicurus auroreus). Over the 3 years, 20 boxes were used twice by Cinereous Tits, 17 boxes were used twice by Varied Tits, and 1 box was used thrice by each. All boxes were randomly dispersed within 500 m on both sides of the path (e.g., Fig. 1, dispersion indices χ2 = 13.46, df = 19), as were those occupied by both Varied Tits (χ2 = 8.68, df = 19) and Cinereous Tits (χ2 = 18.02, df = 19).
Table
1.
The total number of nestboxes placed, occupied and the occupancy rate in 2016, 2017, and 2018. The main species that used the artificial nestbox were Varied Tit and Cinereous Tit.
Figure
1.
Maps of Liaoning Xianrendong National Nature Reserve showing the distribution of the nestboxes. Solid black lines suggest the range on both sides of the paths. Circles indicate position of nestboxes in 2018 and the species occupying them.
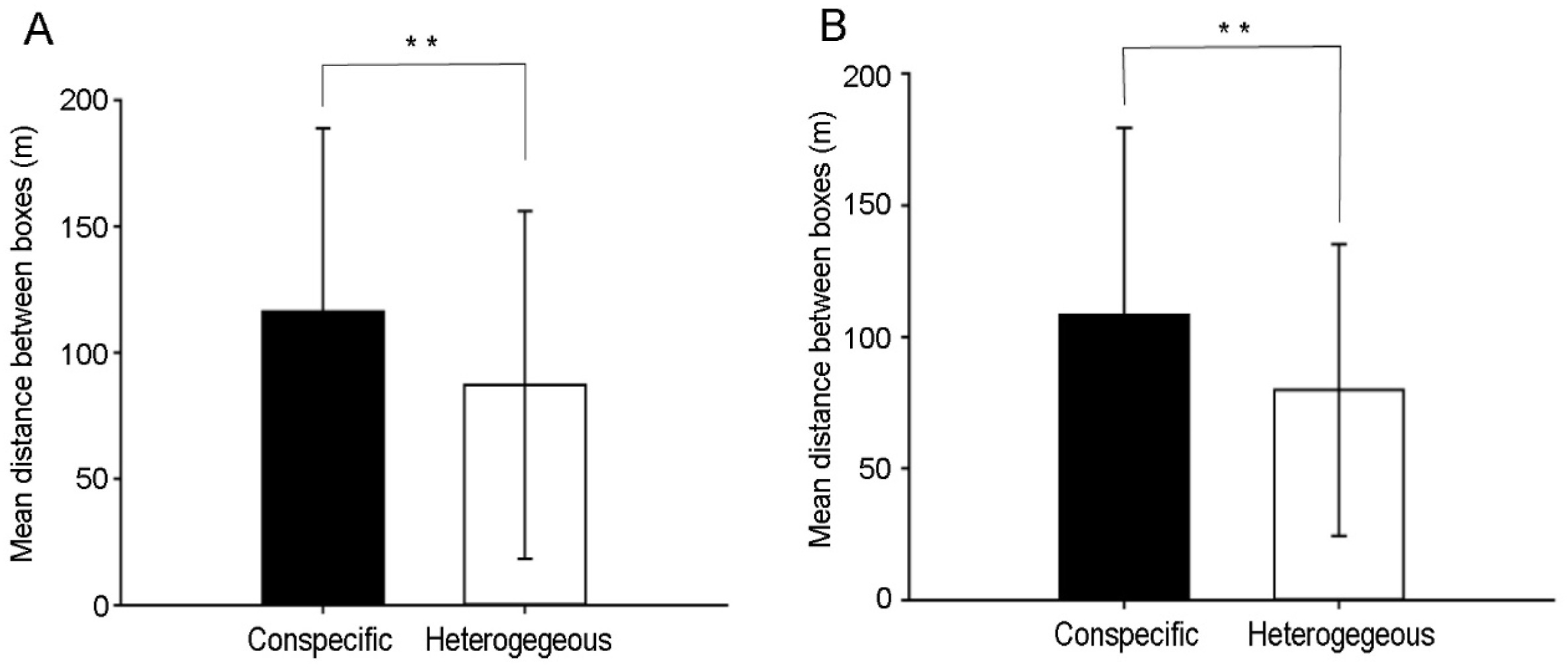
The distance between breeding nests was significantly affected by neighbor type (F = 29.652, P < 0.001), but not species (F = 1.304, P = 0.254). The distance between intraspecific breeding nests of Varied Tits was significantly longer than that between interspecific nests (Fig. 2A). For Cinereous Tits, the same was true (Fig. 2B). However, reproduction of Varied and Cinereous Tits was not significantly relevant to the distance from their neighbor’s nest (Table 2).
Figure
2.
Using mixed effect models to test the relationship between box distance and neighbor type (conspecific/heterspecific), box number and year. (A) Mean distance between the focal nest of Varied Tit (Sittiparus varius) and its conspecific and heterospecific neighbors. The distance between intraspecific breeding nests (116.28 ± 72.5 m, n = 140) was significantly longer than that between interspecific nests (87.17 ± 68.78 m, t = −3.537, P = 0.001). The mixed effect model had the statistics F3,311 = 6.419, r2 = 0.059, P < 0.001. (B) Mean distance between the focal nest of Cinereous Tit (Parus cinereus) and its conspecific and heterospecific neighbors. The distance between intraspecific nests (108.40 ± 71.04 m, n = 155) was significantly longer than that between interspecific nests (79.80 ± 55.50 m, t = −4.108, P < 0.001). The mixed effect model had the statistics F3,311 = 5.769, r2 = 0.053, P < 0.001.
Table
2.
Statistical results of the effect of conspecific and heterospecific neighboring nest distance on the focal nest’s hatching rate (mixed effect models) and reproductive success (linear models). The mixed effect model for the Cinereous Tit had the statistics n = 70, r2 = 0.055, and for the Varied Tit n = 72, r2 = 0.026. The linear model for the Cinereous Tit had the statistics n = 53, F2,14 = 0.121, r2 = 0.017, and for the Varied Tit n = 53, F2,16 = 0.282, r2 = 0.034.
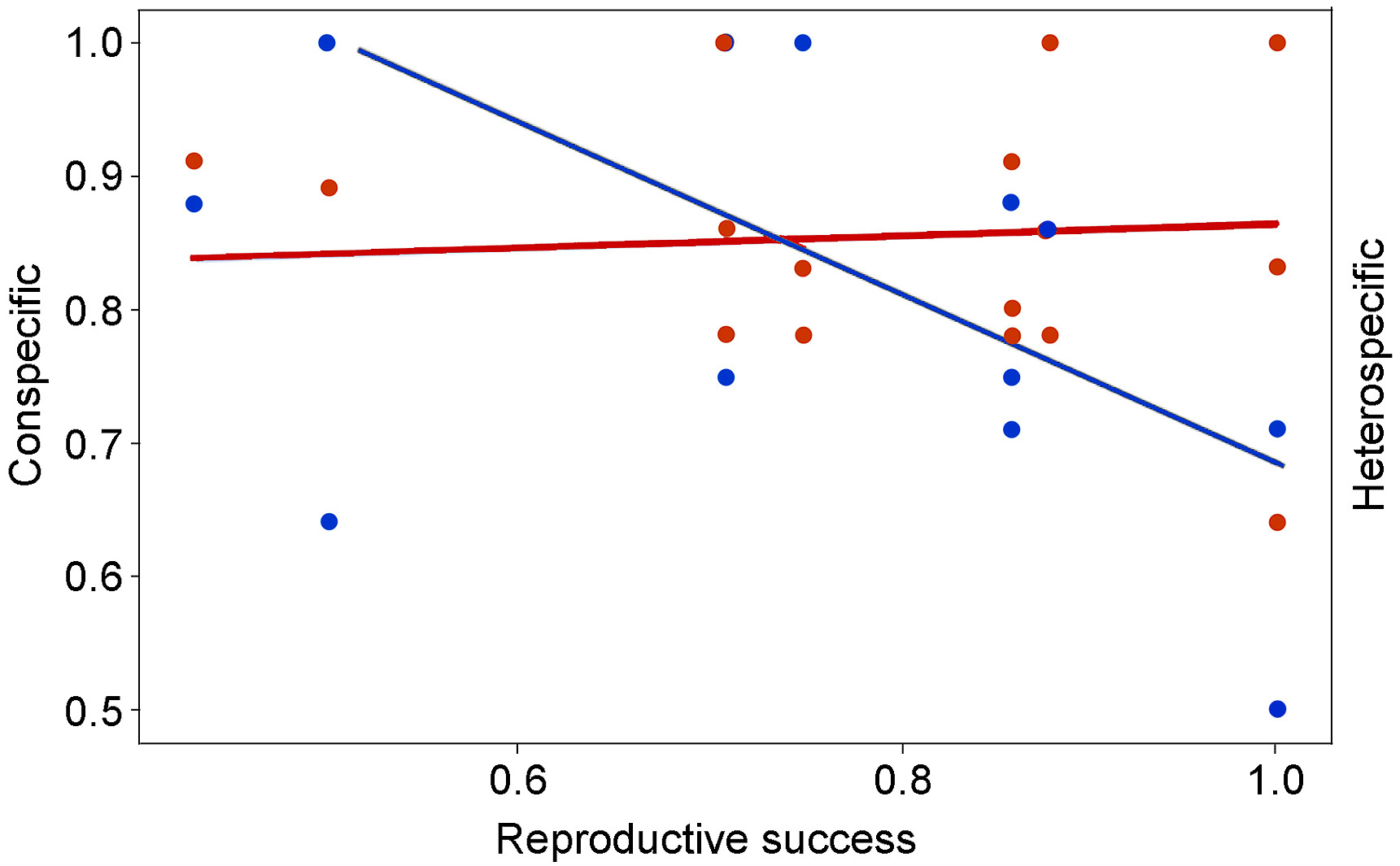
The hatching rate of Varied Tits (n = 144, r2 = 0.892) was not related to recorded breeding behavior of their neighbors (hatching rate, t = 0.773, P = 0.982; and breeding synchrony, t = −0.419, P = 0.333). Its reproductive success rate (n = 38, r2 = 0.270, F5,32 = 2.362, P = 0.062) was negatively related to that of their neighbors (t = −2.677, P = 0.012) and positively related to the interaction between neighbor type and the reproductive success rate of the neighbor (t = 2.142, P = 0.040). Separate analyses on intraspecific neighbor and interspecific neighbor showed that the reproductive success of Varied Tits was negatively related to that of intraspecific neighbor (t = −2.889, P = 0.011, Fig. 3) and not significantly related to that of interspecific neighbor (t = −0.086, P = 0.932, Fig. 3). The reproductive effect of Cinereous Tits (hatching rate, n = 140, r2 = 0.758; and reproductive success rate, n = 34, r2 = 0.201, F5,28 = 1.404, P = 0.253) was not significantly affected by the recorded breeding behavior of their neighbors.
Figure
3.
Using linear regression to test the relationship between the reproductive success of the Varied Tit and that of its intraspecific neighbor (Varied Tit) and interspecific neighbor (Cinereous Tit), respectively. The reproductive success of the Varied Tits decreased with increasing reproductive success of intraspecific neighbor (t = −2.889, P = 0.011) and was not significantly related to that of interspecific neighbor (t = −0.086, P = 0.932). The blue and red spots represented intra- and inter-specific neighbors. The solid lines were the linear regression lines.
There was spatially separated nest sites selection for Varied Tits and Cinereous Tits. The distance between nests was longer within than between species. While both inter- and intra-specific competitions affect site selection (Tarjuelo et al., 2017) and food abundance (Teather, 1992), intraspecific competition in particular also impacts mate choice (Martínez-Rivera and Gerhardt, 2008) and extra-pair mating (Birkhead et al., 1985). To reduce such competition, individuals of the same species may space each other's nesting sites farther apart. Thus, intra-specific nests were more sparsely spaced. Deeming (2017) also shows that the conspecific nest distance is longer than the heterospecific one for Blue Tits and Great Tits. These results support that intraspecific competition in nest selection is more intense than interspecific competition.
Regarding spatial distribution, Varied and Cinereous Tits showed a similar pattern. The distance to their neighbor’s nest was not significantly relevant to reproduction. But intense competition arises from the relatively short distance between two nests of the same species, which may adversely affect the reproduction of the breeding nest (Deeming et al., 2017). In our study, fewer than half of the installed boxes were occupied. The average distance between two nearest nestboxes was around 30 m, about twice the distance between two nearest neighbors. The space interval was relatively sufficient, and that was not so drastic to affect the reproductive success.
The breeding success of Varied Tits may or may not be affected by that of their neighbors. The type matters. Between conspecific neighbors there is a negative correlation whereas between heterospecific ones, no identifiable correlation. Competition can reduce the number of offsprings (Gurevitch et al., 1992). In resource-restricted habitat, we are likely to detect the effect of density-dependence on reproduction in territorial species (Dhondt, 2010). When the conspecifics reproduce in the same area, there will be severe competition for food (Krüger et al., 2012). Food shortage is known to disrupt reproduction.
However, Cinereous Tits did not show significant correlation to any recorded reproductive behavior of their neighbors. The results are supported by Møller’s (2018) finding that the breeding behavior of Blue Tits is affected by conspecifics but not Great Tits. In other words, the effect of competition on demographic variables has interspecific divergence. This may result from the either the scramble or exploitation-type competition for food both within and between species, and interference competition between different body sizes (Dhondt, 1977, 2010), as Cinereous Tits are larger than Varied Tits. Interspecific divergence may be related to the variances in clutch size of the two species. In our research site, the clutch size of Varied Tits is mostly 7–9, and the clutch size of Cinereous Tits is 7–13. Flexible adjustment of clutch size might be one of the ways for big tits to reduce competition. Behavioral differences may also contribute to the divergence. Active individuals can get more food resources under the same circumstance (David et al., 2011). So Varied Tits, which are less exploratory, active, and risk-taking than Cinereous Tits (Zhang et al., 2019), get less. Reproductive success of Cinereus Tits is not significantly affected.
This study focused on one option of breeding site, namely through artificial nestboxes, rather than exhausting all possibilities. For example, nests in natural cavities are not considered. This is because on the reserve, where dead wood is removed in a timely manner, natural cavities are in insufficient supply. Before the breeding season, in March, we ringed the birds on the experimental site. During the entire breeding test period, unringed individuals were rarely spotted. Therefore, we assume that, at least in the area where artificial nest boxes were hung, natural nests were negligible.
Boxes that are too densely distributed can even negatively affect the breeding behavior of birds, especially in areas where natural nests are extremely scarce. Thus, artificial nestboxes usually have low occupancy rate (Mänd et al., 2005; Deeming et al., 2017). This knowledge can be applied to the arrangement of nestboxes for bird study and conservation.
5.
Conclusions
Our results show that intra- and inter-specific competition affects the nest distribution of two hole-nesting species. The effect on reproductive outcome has interspecific divergence. The reproductive outcome of Varied Tits is significantly affected by its conspecific and heterospecific neighbors, but that of Cinereous Tits is not affected.
Authors’ contributions
YJ conceived the experiments, analyzed the data, wrote the manuscript, and reviewed the drafts; YB conceived and designed the experiments, performed the experiments, analyzed the data, and wrote the manuscript; RM analyzed the data; JZ performed the experiments; DW conceived and designed the experiments, and reviewed drafts of the paper. All authors read and approved the final manuscript.
Ethics statement
All our study procedures were approved by Liaoning Xianrendong National Nature Reserve.
Declaration of competing interest
The authors declare that they have no competing interests.
Acknowledgements
Thanks for the support of Liaoning Xianrendong National Nature Reserve. This work was supported by the National Natural Science Foundation of China (No. 31872231 to DW, No. 32000316 to YJ).
Birkhead TR, Johnson SD, Nettleship DN. Extra-pair matings and mate guarding in the Common Murre Uria Aalge. Anim Behav. 1985; 33: 608-619
Boucher-Lalonde V, Morin A, Currie DJ. A consistent occupancy-climate relationship across birds and mammals of the Americas. Oikos. 2014; 123:1029-1036
David M, Auclair Y, Cezilly F. Personality predicts social dominance in Female Zebra Finches, Taeniopygia guttata, in a feeding context. Anim Behav. 2011; 81:219-224
Deeming DC, Biddle LE, Christopher RF. Interspecific and intraspecific spatial separation by birds breeding in nest boxes. Avian Conserv Ecol. 2017; 12:1
Dhondt, A.A., 1977. Interspecific competition between Great and Blue Tit. Nature 268, 521–523
Dhondt AA. Effects of competition on great and blue tit reproduction: Intensity and importance in relation to habitat quality. J Anim Ecol. 2010; 79: 257-265
Dhondt, A.A., 2012. Interspecific competition in birds. In: Birkhead, T.R. (Ed.), Oxford Avian Biology Series. Oxford University Press, Oxford, UK.
Evans MR, Lank DB, Boyd WS, Cooke FA. Comparison of the characteristics and fate of Barrow’s Goldeneye and Bufflehead nests in nest boxes and natural cavities. The Condor. 2002; 104:610-619
Finch DM. Effects of predation and competitor interference on nesting success of house wrens and tree swallows. The Condor. 1990; 92:674-687
Fowler, J., Cohen, L., Jarvis, P., 1998. Practical Statics for Field Biology, second ed. Wiley, Chichester, UK.
Gurevitch J, Morrow LL, Wallace A, Walsh JS. A meta-analysis of competition in field experiments. Am Nat. 1992; 140:539-572
Kruger O, Chakarov N, Nielsen JT, Looft V, Grunkorn T, Struwe-Juhl B, et al. Population regulation by habitat heterogeneity or individual adjustment? J Anim Ecol. 2012; 81:330-340
Laube, I., 2011. Avian Range Dynamics: Traits, Biotic Interactions and Niches. Ph.D thesis. Goethe University Frankfurt, Frankfurt.
Mand RV, Tilgar A, Lohmus, Leivits A. Providing nest boxes for hole-nesting birds - does habitat matter? Biodiver Conserv. 2005; 14:1823-1840
Martin TE. Nest predation and nest sites new perspectives on old patterns. BioScience. 1993; 43:523-532
Martin TE. Food as a limit on breeding birds: A life-history perspective. Ann Rev Ecol System. 1987; 18:453-487
Martinez-Rivera CC, Gerhardt HC. Advertisement-call modification, male competition, and female preference in the bird-voiced treefrog Hla avivoca. Behav Ecol Sociobiol. 2008; 63:195-208
Minot EO, Perrins CM . Interspecific interference competition - nest sites for blue and great tits. J Anim Ecol. 1986; 55: 331-350
Moeller AP, Adriaensen F, Artemyev A, Banbura J, Barba E, Biard C. Clutch-size variation in western Palaearctic secondary hole-nesting passerine birds in relation to nest box design. Methods Ecol Evol. 2014; 5:353-362
Moeller AP, Balbontin J, Dhondt AA, Remes V, Adriaensen F, Biard C. Effects of interspecific coexistence on laying date and clutch size in two closely related species of hole-nesting birds. J Anim Ecol. 2018; 87:1738-1748
Schoener TW. Field experiments on interspecific competition. Am Nat. 1983; 122:240-285
Szabo ND, Algar AC, Kerr JT. Reconciling topographic and climatic effects on widespread and range-restricted species richness. Glob Ecol Biogeogr. 2009; 18:735-744
Tarjuelo R, Morales MB, Arroyo B, Manosa S, Bota G, Casas F. Intraspecific and interspecific competition induces dependent habitat niche shifts in an endangered steppe bird. Ecol Evol. 2017; 7:9720-9730
Teather KL. An experimental study of competition for food between male and female nestlings of the red-winged blackbird. Behav Ecol Sociobiol. 1992; 31:81-87
Zhang L, Wang J, Wan DM, Bi YJ, Zhang L. Narrow or wide distribution? A comparison of behavioral traits between the Varied Tit (Sittiparus varius) and Great Tit (Parus cinereus). Chin J Zool. 2019; 54:471-477
Table
1.
The total number of nestboxes placed, occupied and the occupancy rate in 2016, 2017, and 2018. The main species that used the artificial nestbox were Varied Tit and Cinereous Tit.
Table
2.
Statistical results of the effect of conspecific and heterospecific neighboring nest distance on the focal nest’s hatching rate (mixed effect models) and reproductive success (linear models). The mixed effect model for the Cinereous Tit had the statistics n = 70, r2 = 0.055, and for the Varied Tit n = 72, r2 = 0.026. The linear model for the Cinereous Tit had the statistics n = 53, F2,14 = 0.121, r2 = 0.017, and for the Varied Tit n = 53, F2,16 = 0.282, r2 = 0.034.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds