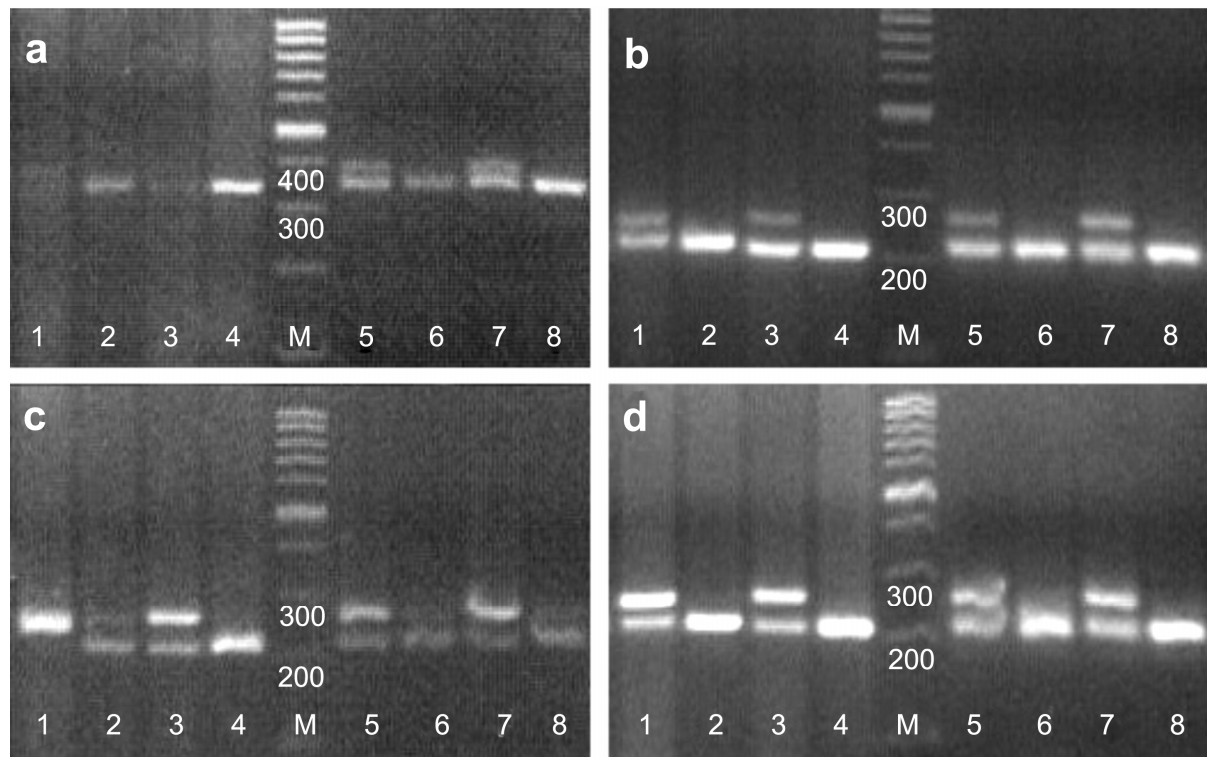
The primer pair sex1/sex2, which can be widely applied for sex identification in Galliform species, was used to determine the sex of 17 Passeriform species. As CHD-W fragments tended to be preferentially amplified, which may cause unnecessary misidentification in bird species with little difference between CHD-Z and CHD-W, we modified sex1 and sex2, obtaining sex1' and sex-mix respectively. Primer sets were then recombined to conduct sex identification. After testing several Passeriforme birds of known sex, we found that the primer pair sex1'/sex2 was better at limiting the preferential amplification of CHD-W fragments. As they are being frequently used in sex allocation study of Aegithalos concinnus and song learning research of Lonchura striata, we can expect more applications of this primer pair to further studies in Passeriformes.
Occurrence and abundance of birds at typical feeders in gardens has been studied in detail for many decades. In this study, motion triggered infrared camera traps have been used to analyze discovery of a novel food source and circadian patterns of feeding. Camera traps are just at the rise of being used for bird studies (e.g., Randler and Kalb 2018; Hillemann et al. 2019). Two main lines of research have been addressed in this study. First, I analyzed at what time discovery of a novel food source occurs. Second, the more general questions are addressed, which species discover novel food sources first, and which species visit those food sources. Also, the duration until detection is analyzed.
Turning to the first aspect, predation influences activities, such as feeding or maintenance behaviour (Randler 2006). Small songbirds, therefore, have to trade off starvation risk against predation. Depending on predation risk, individuals may reduce their fat storage against their escape or flight ability and hence, the trade-off is shifted away from foraging (Gosler et al. 1995; Gentle and Gosler 2001). Farine and Lang (2013) proposed the hypothesis that individuals should prioritize discovery and assessment of potential resources early in the day, before switching to exploitation as the day progresses. Bonter et al. (2013) found that birds generally started to feed before sunrise and continued to forage at an increasing rate throughout the day. Therefore, I hypothesize that discovery of novel food resources should occur during the morning hours.
Food availability is an important environmental cue and adaptations that allow discovery and use of ephemeral food should be favored by natural selection (Ducatez et al. 2015; Tryjanowski et al. 2015a). As discovery of novel food is important for survival in urban and rural habitats (Tryjanowski et al. 2015a), it should also be important in natural environments. If food sources are clumped, the first individual arriving may be often the winner in terms of food quality and quantity (Tryjanowski et al. 2015a, 2017).
This study differs in some respect from previous work, making a new contribution to the field. First, previous studies have been carried out during the winter months (Bonter et al. 2013; Farine and Land 2013; Tryjanowski et al. 2015a, b, 2016, 2017; Moiron et al. 2018). During the winter, birds are more time constrained than during summer, and colder temperatures lead to a higher energy demand. Second, the study was carried out in a natural environment outside from cities and villages where no artificial bird feeding has happened before (see e.g., Farine and Land2013). To the best of my knowledge, there are no tests of how fast birds recognize novel food sources in a natural environment.
The study was conducted on a small mountain range, the Spitzberg, in SW Germany (Baden-Württemberg). The Spitzberg (48°30 N, 9°00 E) is located between the city of Tübingen in the east and Rottenburg-Wurmlingen in the west, extending in length about 6 km and with the widest N-S extension of about 2 km (Gottschalk and Randler 2019). The highest point is the Kapellenberg near Wurmlingen with a height of 475 m. On the southern slopes, the forest is almost completely cleared and terraces with dry stone walls were cultivated for winegrowing. The largest part, however, is covered by woods, including the heights and the northerly slopes. The forest is characterized mainly by the Scots Pine (Pinea sylvestris), different oak species (Quercus sp.) and Beech (Fagus sylvatica). The study was carried out in the wooden areas.
Great Tits (Parus major) are among the most common species in the study area with about 370‒390 breeding pairs (Gottschalk and Randler 2019; see Table 1). Also, in urban and suburban areas, Great Tits usually frequent feeders regularly (Tryjanowski et al. 2015a, b). Predators in the area are diurnal and nocturnal mammalian and avian species. For example, 1‒2 breeding pairs of Sparrowhawk (Accipiter nisus) nest in the area, as well as Common Buzzards (Buteo buteo; 7‒10 pairs), Red Kite (Milvus milvus; 2‒4 pairs), and Tawny Owls (Strix aluco; Gottschalk and Randler 2019).
Table
1.
a Average time taken for a species to discover a novel food source (expressed in hours), b Average clock time taken when a species discovered a novel food source (expressed in clock times), c Population size of the species in the study area
The study took place between 29 June and 5 October, 2018. For this study, camera traps with a special macro lens have been deployed, allowing close-up photographs. We used six different Bushnell Natureview cameras simultaneously at different places, all with the same macro lens and of the same model (model 119740). The cameras were placed at a distance of 0.6 m near a feeding station and the field of view covered about 50 cm × 35 cm (0.175 m2) of the study location. Previous work has assessed this as a reliable distance between small birds and different camera traps including the model used here (see Randler and Kalb 2018).
I set the trigger sensor level on the highest level, the number of images released to three in a row. I set up the cameras and immediately afterwards released the trigger to test functionality. Also, when returning back to check the SD cards and batteries, I approached the camera in a manner that should trigger photos. This was used as some kind of test to check if the cameras are still working.
I applied a variety of food to attract birds: apples, apple juice, honey, peanut mousse, raisins, prunes, bird food, and sunflower seeds. All feeding places were baited with the same variety of food to avoid any influence of different foods on the results. Thus, a standardized food mix was applied. The food was replenished every second or third day to provide a continuous food supply. Food was presented in open, unsheltered feeding places allowing approaches from above, below and from all sides.
There were 41 active camera locations with bait stations. Following Meek et al. (2012) an active camera set was defined as a bait station with non-toxic bait used to attract animals to within the detection zone of a camera trap. On six out of the camera trap places, no bird species occurred leaving 35 locations for analysis. Data were screened and date, time, taxon/species and number of individuals were transferred into an Excel sheet. A temporal buffer of 5 min was used to distinguish between consecutive events within species at a camera trapping station (Meek et al. 2012). This was based on Meek et al. (2012) who suggested using 1–5 min for small mammals. Total trap days in the current study were 457 days (mean 13.1 ± 5.4 nights per camera location), corresponding to a total observation time of 10, 968 h with 1951 total bird events.
The species that first discovered a novel food source was labelled as explorer species, and species that did not discover the food source but used them after another species has discovered it were assigned visitors. The number of feeders with the discovery by a species and the number of feeders visited by a species were correlated with their population size using Kendall's tau. Sample sizes were number of species (N = 18) and basis for the assignment was number of feeders (N = 35). Therefore, a species can be an explorer in 35 cases and a visitor also in 35 cases.
Eighteen species occurred at the feeders (Fig. 1). A total of nine species were explorers and discovered the novel food source first: Great Tit, Robin (Erithacus rubecula), Nuthatch (Sitta europaea), and Blackbird (Turdusmerula) discovered novel food sources most often as first arriving species. Similarly, these species visited most of the food sources. There was a moderate correlation in these 18 species between population size and first discovery (Kendall-tau: 0.508, p = 0.007) and between population size and visitation of the feeders (Kendall-tau: 0.500, p = 0.006). Great Tits were among the most common visitors and the ones that discovered most of the 35 novel food sources first (Fig. 1). Considering all visitations, Great Tits occurred at 80% of the feeders (Fig. 1). Concerning the first visit, Table 1 shows mean, standard deviation and median of the first arrivals of a given bird species to a novel food source. The time to first discovery was rather long. Usually, it took some days until the feeders were discovered (about 3‒5 days in Great Tits). The shortest detection times were found in Great Tit, Robin, and Winter Wren (Troglodytes troglodytes). The average clock time of a first discovery was before noon in most species (Table 1). In Great Tits, first discoveries were more common before noon (18 before, 10 after noon; Fig. 2).
Figure
1.
Abundance and frequency of first bird visiting a novel food source according to species and total number of feeders visited. Maximum number of feeders was N = 35
Food was discovered more in the morning than in the afternoon, concerning all species as well as Great Tits separately (Farine and Lang2013). This supports the hypothesis that time available in the morning should be spent on exploration and exploitation/visitation should occur later during the day. In most cases the first species discovering the novel food was the Great Tit. This is similar to the results of Tryjanowski et al. (2015a, b, 2017), who carried out their study in human settlements. Thus, the present data expand this finding to Great Tits in a natural environment without any previous artificial bird feeding activities and allows characterizing Great Tits as an explorative species in general. Turdus philomelos and Poecile palustris were species that discovered no novel food patch but used/visited them after its has been discovered by other species. Interestingly, population size was a predictor how many feeders were discovered and used by a species; thus it is difficult to assess whether these two species were no discoverers just because of their population size or because they are somewhat parasitic in exploiting food resources found by other species.
The mean latency time to discovery was about 3‒5 days for all species. This is strikingly different to the other studies. For example, in Tryjanowski et al. (2017), the mean latency was 24.8 min for all species and 21.5 min for Great Tits. Hillemann et al. (2019) reported that their feeders were usually discovered during the first hours of the day. Similarly, novel food patches were discovered quickly (Farine and Lang 2013). However, those studies placed a novel feeder only a few hundred meters away from previous ones and birds were habituated to regular feeding. In my study plot, no regular feeding occurs because it is in a natural environment without any feeders. Therefore, this study adds to the previous ones about the discovery of novel food in an unmanipulated environment.
There are some limitations in this study. First, predation events were not directly or indirectly assessed and there were no experiments to simulate predation or predation risk and its effects on feeding habits. Sample sizes in this study are small, which is owed to the design of the study, but results should not be overstated. As the feeders were placed far away from another and given the size of the breeding population of the different species, pseudo-replication, i.e., discovery of different feeders by the same individual seems highly unlikely. Future studies should replicate the findings using a considerable higher amount of camera traps and feeders. This would allow more complex and rigorous statistical analysis going beyond these more descriptive analyses. Additionally, environmental variables could be included in further studies.
Acknowledgements
I am grateful to Jochen Kalb, MSc, for checking all images and establishing the database.
Authors' contributions
The author read and approved the final manuscript.
Bensch S, Westerdahl H, Hansson B, Hasselquist D. 1999. Do females adjust the sex of their offspring in relation to the breeding sex ratio? J Evol Biol, 12: 1104–1109
Cockburn A, Double MC. 2008. Cooperatively breeding superb fairy-wrens show no facultative manipulation of offspring sex ratio despite plausible benefits. Behav Ecol Sociobiol, 62: 681–688
Dreiss A, Richard M, Moyen F, White J, Moller AP, Danchin E. 2006. Sex ratio and male sexual characters in a population of blue tits, Parus caeruleus. Behav Ecol, 17: 13–19
Dubiec A, Zagalska-Neubauer M. 2006. Molecular techniques for sex identification in birds. Biol Lett, 43: 3–12
Ellegren H. 1996. First gene on the avian W chromosome (CHD) provides a tag for universal sexing of non-ratite birds. Proc R Soc Lond B, 263: 1635–1641
Ellegren H, Sheldon BC. 1997. New tools for sex identification and the study of sex allocation in birds. Trends Ecol Evol, 12: 255–259
Ellegren H. 2002. Dosage compensation: do birds do it as well? Trends Genet, 18: 25–28
Fiala KL. 1980. On estimating the primary sex ratio from incomplete data. Am Nat, 115: 442–444
Frank SA. 1990. Sex allocation theory for birds and mammals. Ann Rev Ecol Syst, 21: 13–55
Fridolfsson AK, Ellegren H. 1999. A simple and universal method for molecular sexing of non-ratite birds. J Avian Biol, 30: 116–121
Griffiths R, Double MC, Orr K, Dawson RJG. 1998. A DNA test to sex most birds. Mol Ecol, 7: 1071–1075
Hatchwell BJ, Russell AF. 1996. Provisioning rules in cooperatively breeding long-tailed tits Aegithalos caudatus: An experimental study. Proc R Soc Lond B Biol Sci, 263: 83–88
Kahn NW, John JS, Quinn TW. 1998. Chromosome-specific intron size differences in the avian CHD gene provide an efficient method for sex identification in birds. Auk, 115: 1074–1078
Lessells K, Mateman C. 1996. Molecular sexing of birds. Nature, 383: 761–762
McGowan A, Hatchwell BJ, Woodburn RJW. 2003. The effect of helping behaviour on the survival of juvenile and adult long-tailed tits Aegithalos caudatus. J Anim Ecol, 72: 491–499
McQueen HA, McBride D, Miele G, Bird AP, Clinton M. 2001. Dosage compensation in birds. Curr Biol, 11: 253–257
Nakagawa S. 2004. Is avian sex determination unique?: clues from a warbler and from chickens. Trends Genet, 20: 479–480
Quinn TW, Cooke F, White BN. 1990. Molecular sexing of geese using a cloned Z chromosomal sequence with homology to the W chromosome. Auk, 107: 199–202
Saino N, de Ayala RM, Martinelli R, Boncoraglio G. 2008. Male-biased brood sex ratio depresses average phenotypic quality of barn swallow nestlings under experimentally harsh conditions. Oecologia, 156: 441–453
Wang N, Zhang ZW. 2009. The novel primers for sex identification in the brown eared-pheasant and their application to other species. Mol Ecol Resour, 9: 186–188
West SA, Sheldon BC. 2002. Constraints in the evolution of sex ratio adjustment. Science, 295: 1685–1688
Whittingham LA, Dunn PO. 2000. Offspring sex ratios in tree swallows: females in better condition produce more sons. Mol Ecol, 9: 1123–1129
Woxvold IA, Magrath MJL. 2008. Sex-biases in the hatching sequence of cooperatively breeding apostlebirds Struthidea cinerea. Evol Ecol, 22: 139–151
Michael A. Weston, Nick Porch, Desley A. Whisson, et al. Do different camera trap lures result in different detection rates of vertebrates because of their attractiveness to invertebrates?. Ecological Management & Restoration, 2024.
DOI:10.1111/emr.12603
2.
Emilia Grzędzicka. Bird Feeder Explorers Are Not Attracted by the Seeds of Invasive Weeds in Winter. Diversity, 2024, 16(2): 81.
DOI:10.3390/d16020081
3.
Christoph Randler, Nadine Kalb. Circadian activity of the fat dormouse Glis glis measured with camera traps at bait stations. Mammal Research, 2021, 66(4): 657.
DOI:10.1007/s13364-021-00583-6
Table
1.
a Average time taken for a species to discover a novel food source (expressed in hours), b Average clock time taken when a species discovered a novel food source (expressed in clock times), c Population size of the species in the study area

 DownLoad:
DownLoad:







 Email Alerts
Email Alerts RSS Feeds
RSS Feeds