Takehiko Shimizu, Masayuki Senzaki, Yuichiro Fujioka, Satoquo Seino. 2023: Relative importance of tidal flats and artificial habitats for two spoonbill species and related interspecific differences. Avian Research, 14(1): 100107. DOI: 10.1016/j.avrs.2023.100107
Citation:
Takehiko Shimizu, Masayuki Senzaki, Yuichiro Fujioka, Satoquo Seino. 2023: Relative importance of tidal flats and artificial habitats for two spoonbill species and related interspecific differences. Avian Research, 14(1): 100107. DOI: 10.1016/j.avrs.2023.100107
Takehiko Shimizu, Masayuki Senzaki, Yuichiro Fujioka, Satoquo Seino. 2023: Relative importance of tidal flats and artificial habitats for two spoonbill species and related interspecific differences. Avian Research, 14(1): 100107. DOI: 10.1016/j.avrs.2023.100107
Citation:
Takehiko Shimizu, Masayuki Senzaki, Yuichiro Fujioka, Satoquo Seino. 2023: Relative importance of tidal flats and artificial habitats for two spoonbill species and related interspecific differences. Avian Research, 14(1): 100107. DOI: 10.1016/j.avrs.2023.100107
Artificial/seminatural environments, such as aquacultural ponds, saltpans, and croplands, have recently been acknowledged as important habitats for coastal waterbirds. Although coastal waterbirds tend to use artificial habitats around tidal flats as roosting sites during high-tide, it remains unclear whether the importance of surrounding habitats relative to tidal flats varies among landscape types, seasons, species, or tidal conditions. The Black-faced Spoonbill (Platalea minor) and Eurasian Spoonbill (P. leucorodia) are two closely related sympatric species in East Asia with narrow and wide distribution ranges and habitat requirements, respectively. We therefore expect that both species will use surrounding artificial habitats across seasons at high tides, but Black-faced Spoonbills will use them less frequently than Eurasian Spoonbills. Here, we address these hypotheses in the Imazu tidal flat and its surrounding environments in southern Japan. We investigated the habitat use and behavioral patterns of both species through route and behavioral surveys during the fall migration and wintering seasons in 2021. We found that both species used surrounding habitats including artificial ones more frequently than the tidal flat regardless of the tidal condition or season, but spoonbills used these habitats more frequently in winter than in autumn. We also found that Eurasian Spoonbills foraged in surrounding artificial habitats more frequently than Black-faced Spoonbills. These results not only demonstrate how coastal waterbirds exploit surrounding habitats relative to tidal flats but also suggest that the importance of surrounding habitats varies among species and seasons. Our study thus emphasizes that valuing and managing surrounding habitats in addition to tidal flats are key to conserving globally declining waterbirds.
Previous research has shown that waterbird abundance in artificial habitats varies with physical features and tidal conditions (Rogers et al., 2006; Sripanomyom et al., 2011; Jackson et al., 2019). For example, when the surrounding tidal flat is submerged or unavailable, the number of waterbirds tends to increase in those artificial wetlands with shallow water areas and bare lands (i.e., high-tide roosts) (Rogers, 2003; Jackson et al., 2019). Moreover, preferred artificial wetlands have been suggested to be those located less than 5 km adjacent to natural wetlands (Dias et al., 2006; Jackson et al., 2019) or free from predation pressure/human disturbance (Rosa et al., 2006; Jackson et al., 2019). Nevertheless, most research has briefly described waterbird preferences for artificial habitats during a limited period of the year (Jackson et al., 2020), and we know little about how those preferences and waterbirds’ behaviors vary with seasons or habitat types.
Waterbirds may also have species-specific habitat use patterns in artificial wetlands (Yasué and Dearden, 2009; Li et al., 2013; Jackson et al., 2019, 2020). For instance, specialist species (e.g., species with narrow distribution ranges and habitat requirements) may use artificial habitats less frequently than generalist species (e.g., species with wide distribution ranges and habitat requirements) because of their strict habitat requirements and vulnerabilities to disturbances (Jackson et al., 2021; Wang et al., 2022). However, it remains unknown how the habitat use patterns in artificial environments differ between specialist and generalist species, especially between closely related sympatric species. Clarifying this is important for conservation because such knowledge will allow us to identify indispensable habitats, particularly for specialist species.
The purposes of this study were: (1) to clarify the detailed habitat use and behavioral patterns of Black-faced Spoonbills and Eurasian Spoonbills in a tidal flat and the surrounding environments in two different seasons; and (2) to examine whether there are different habitat use patterns between the two species. We expect that spoonbills will use artificial habitats when the surrounding tidal flats are unavailable due to high tide. Based on the differences in their distribution ranges and breeding habitat requirements, Black-faced and Eurasian Spoonbills can be viewed as specialist and generalist species, respectively. Therefore, we expect Black-faced Spoonbills to use artificial habitats less frequently or more limited types of artificial habitats than Eurasian Spoonbills. In this study, we employed two different methods to investigate habitat use and behavioral patterns, respectively, around the Imazu tidal flat and its surrounding environments in southern Japan. We also examined the relationship between the spoonbills’ habitat use patterns and environmental characteristics.
2.
Methods
2.1
Study area
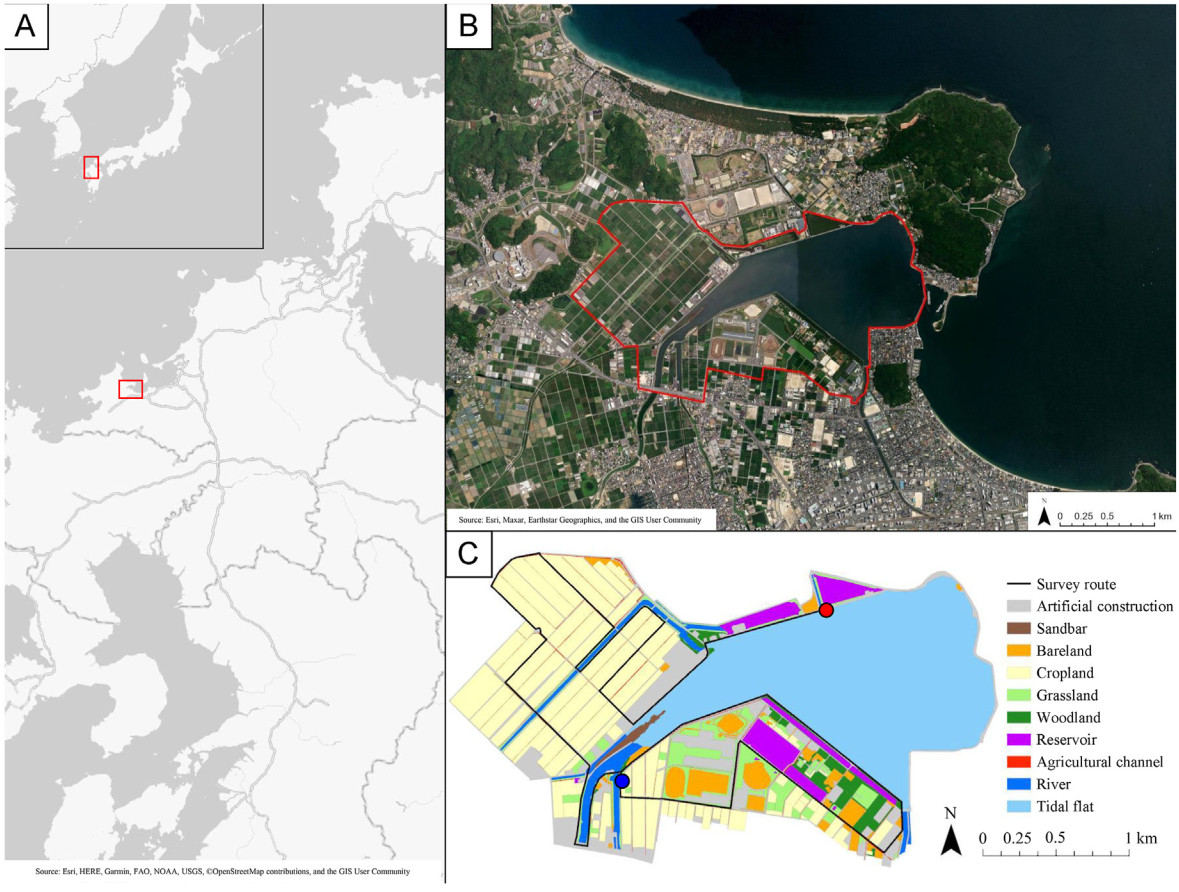
This study was conducted in the Imazu tidal flat and its surrounding environments, located in Fukuoka Prefecture in the northern Kyushu region, Japan (33.59–33.60° N, 130.23–130.27° E; 5.37 km2) (Fig. 1). We focused on this area for two reasons. The first is that seawater covers the full range of the tidal flat at high tides even in neap tides, which forces spoonbills to leave the tidal flat, making it an advantageous site to identify the surrounding habitats utilized by spoonbills. Second, there are similar numbers of Black-faced Spoonbills and Eurasian Spoonbills at the site, which makes it easy to identify the interspecific differences in habitat use patterns. The Imazu tidal flat is a small mudflat whose area is 1.8 km2, which includes permanently inundated areas, and is located at the endpoints of four rivers. The coastlines of the Imazu tidal flat are protected by concrete walls. There is a sandbar connected to the road in the estuary of the Imazu tidal flat. This sandbar is covered with shrubs, grasses, and rocks, but not covered in water even at high tides. The surrounding environments contain natural ones like the sandbar and artificial ones like croplands, artificial buildings, and water reservoirs in which the reed (Phragmites australis) is grown (see 2.3.1. Landscape classification for the detailed landscape types). Most croplands are utilized as paddy fields, and agricultural channels for entering and draining waters are established between croplands.
Figure
1.
Target area (A and B) and landscape classification (C). The red rectangle in the upper left square of (A) shows the overall range of (A). The red rectangle in the overall (A) is the range of (B). The red enclosure in (B) is the survey range. The red circle and blue circle in (C) are the start and endpoint of route surveys, respectively. The image in (B) is the satellite image on September 22, 2021, in ArcGIS. (For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)
We investigated the habitat use patterns of spoonbills and its differences between species or seasons by route surveys in two periods: from January to April 2021 and from October to December 2021. The periods were determined according to the migration schedules of spoonbills; the two target species initiate a southward migration to nonbreeding grounds from September to November, stay at wintering sites from December to February, and return to breeding sites after March to May (Chen et al., 2021; Xi et al., 2021). This migration schedule is not definite and rather different among individuals and regions; thus, we regarded the period from January to April as the wintering period and the period from October to December 2021 as the migration period.
We established the route to cover the whole study area and traveled the route repeatedly (route distance: 14 km, mean travel speed: 16.8 km/h, mean survey time: 50 min) (Fig. 1C). To make seasonal comparison reasonable, surveys were conducted in the daytime without rainfall at a similar proportion of different tidal situations (high and low tides) and time zones (morning, afternoon, and crepuscular) during each season (Table 1). Because our route surveys started from the west bank of the tidal flat and finished at the east bank (Fig. 1C), it was possible to double-count the same individuals in the tidal flat. Since we can survey the tidal flat more inclusively and clearly at the east bank than at the west bank, we conducted the counts from the east bank. The same single surveyor observed spoonbills with a binocular (Canon 15 × 50 IS) and recorded the observation time, number of spoonbills, and location of spoonbills on a paper map. After the route surveys were conducted, we obtained the tidal levels (cm) at the times corresponding to the observations from the 30-s-interval tidal data in Hakata managed by the North East Asian Regional Global Ocean Observing System (NEAR-GOOS, 2021).
Table
1.
Summary of the route surveys in the two periods.
Total
Tidal situation
Time zone
High tide
Low tide
Crepuscular
Morning
Afternoon
Wintering period
Survey times
59
27
32
17
20
22
Survey days
40
26
28
16
19
19
Rate to total survey times
45.8%
54.2%
28.8%
33.9%
37.3%
Migration period
Survey times
34
16
18
13
9
12
Survey days
30
16
18
12
9
12
Rate to total survey times
47.1%
52.9%
38.2%
26.5%
35.3%
The tidal situation is divided into two based on 190-cm tide level since spoonbills could forage in the tidal flat below 190 cm. The time period is divided into three (National Astronomical Observatory of Japan, 2021): crepuscular (1 h before dusk and after dawn), morning (after crepuscular before noon) and afternoon (after noon before crepuscular).
Observed spoonbills were instantly classified as Black-faced Spoonbills, Eurasian Spoonbills, or unidentified spoonbills. Black-faced Spoonbills and Eurasian Spoonbills can be distinguished by the body size and color of the bill tip (Hancock et al., 2011), but these features greatly differ among individuals and could overlap between the two species. Therefore, we identified all spoonbills based only on the lore color pattern; Black-faced and Eurasian Spoonbills have black and white lores, respectively (Hancock et al., 2011). For this reason, when the lore color could not be observed because spoonbills sleep with their bills tucked into their feathers, they were recorded as unidentified.
2.2.2
Behavioral surveys
We recorded all spoonbills’ behaviors at 5-min intervals (Choi et al., 2007) over 2675 and 1330 min (24 and 6 days) during the wintering and migration periods, respectively. Behavioral surveys were conducted during the daytime without rainfall at the same period, but at different times in the route surveys. We observed spoonbills’ behaviors in the following landscape types, where spoonbills had frequently been observed in the route surveys (Fig. 2): tidal flat, river, agricultural channel, reservoir, woodland, and sandbar. The total survey minutes were varied among landscape types and environmental conditions such as tides and time zones (Appendix Table S1) since we randomly searched for spoonbills and recorded spoonbills’ behaviors (see below for the specific behavioral categories). Additionally, one survey time was largely different from survey to survey (range: 5–180 min (1–36 continuous observations), average: 62.2 min) because we sometimes stopped recordings when all observed spoonbills moved to other remote places.
Figure
2.
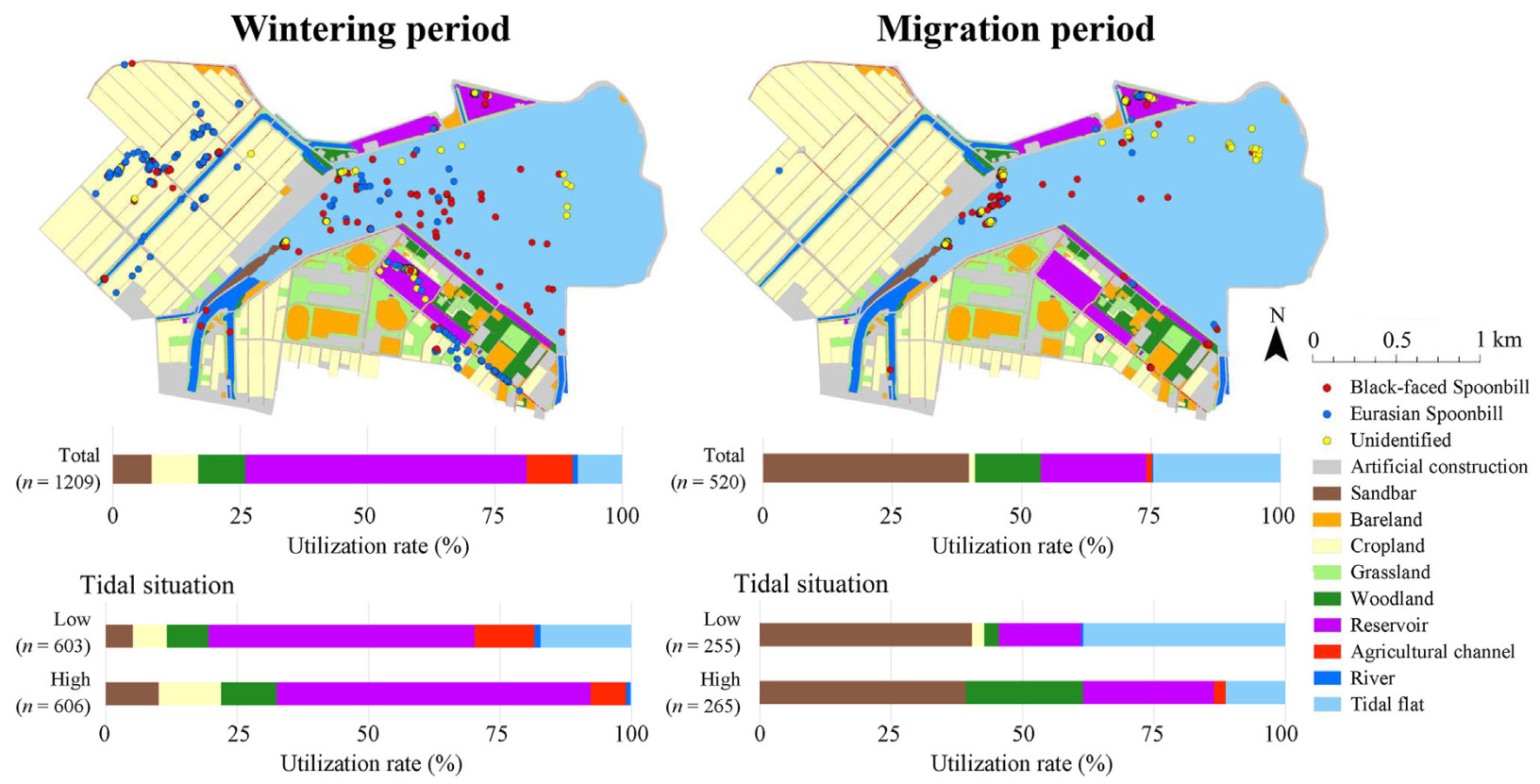
Habitat types utilized by spoonbills in the two periods; wintering (Jan–Apr) and migration (Oct–Dec) periods. The number in parentheses in the bar charts shows the count of observed individuals.
Spoonbill behaviors were classified into seven categories: sleeping, loafing, comforting, social activity, foraging, ground locomotion, and flying locomotion (Wei et al., 2005; Choi et al., 2007; Sullender et al., 2016). Sleeping is resting with the bill tucked into the feathers (Hancock et al., 2011), while loafing is standing with the bill out of the feathers (Wei et al., 2005; Sullender et al., 2016). Comforting is body maintenance activities such as preening and bathing by one individual (Choi et al., 2007; Sullender et al., 2016). Social activity is explicit interacting behaviors among multiple individuals, such as the preening between two birds (Sullender et al., 2016). Foraging is sweeping movements with the bill into the water for probing prey (Choi et al., 2007; Hancock et al., 2011; Sullender et al., 2016). Ground locomotion is walking on the ground with the bill out of the water (Sullender et al., 2016). Flying locomotion is flying movements to/from observing sites (Sullender et al., 2016). We identified spoonbills based on the same criterion used in the route surveys. Observations were performed with a binocular (Canon 15 × 50 IS) and a field scope (KOWA TSN-884 PROMINAR). Surveys were executed by one person in the wintering period and two persons in the migration period.
2.3
Data analysis
2.3.1
Landscape classification
We visually and manually classified the landscape within the survey range by land use category based on the satellite image in ArcGIS with observations (ground-truthing) of the landscape in the target area (Fig. 1C; Table 2). The landscape was largely divided into water and land areas. Water areas were further classified as tidal flat, river, agricultural channel, and reservoir. Land areas were also further classified as cropland, bareland, grassland, sandbar, woodland, and artificial construction. In this study, surrounding environments were defined as any landscape types except for tidal flat. Additionally, artificial environments were defined as both artificially used and seminatural landscape types: agricultural channel, reservoir, cropland, artificial construction, bareland, grassland, and woodland (Table 2). Landscape classification was performed in ArcGIS Pro (ESRI, 2022; Version 2.7.0).
Table
2.
Environmental variables for the MaxEnt models.
Environmental variable
Abbreviation
Explanation
Landscape classification
Categorical classification of the landscape in the survey area
Tidal flat
TF
Broad water areas connected to ocean and strongly influenced by tidal movements
River
RI
Broad natural channels permanently filled with either brackish or fresh water
Agricultural channel
AG
Artificially made narrow water channels for agriculture
Reservoir
RE
Broad water areas where water is permanently retained for human livelihood, such as agriculture and controlling flood
Woodland
WL
Land areas where some short and tall trees or shrubs grow, previously used for human activities
Grassland
GL
Land areas where grasses grow, such as parks and abandoned croplands
Cropland
CL
Land areas used for agriculture
Bareland
BL
Land areas where no plant covers lands
Sandbar
SB
Narrow land areas located in the estuary of the Imazu tidal flat
Artificial construction
AR
Land areas covered with artificial constructions, such as buildings and impervious roads
Agricultural channel classification
Categorical classification of the agricultural channels in landscape classification
Agricultural irrigation
AI
Narrowest channels enclosed by concrete walls for channeling water into croplands
Small channel
SC
Channels broader than AI but narrower than midsized and large channels
Large channel
LC
Largest channels among all types
Midsized shallow channel
MS
Midsized channel whose concrete walls are low and upper parts of walls are covered with plants
Midsized deep channel
MD
Midsized channel whose concrete walls are high and upper parts of walls are not covered with plants
Distance to tidal flat
Euclidean distance to tidal flat in landscape classification
Distance to artificial construction
Euclidean distance to artificial construction in landscape classification
Distance to reed beds
Euclidean distance to the reeds grown in water zones
We first performed a descriptive analysis of the data from route and behavioral surveys. We tested the independence between species and observed environments or behaviors by the chi-square test to clarify the interspecific difference in habitat use patterns. Since our dataset includes a considerable amount of data labeled unidentified, as spoonbills were frequently observed sleeping and their faces were not seen, the test was only applied to the landscape types and behaviors in which the number of unidentified spoonbills was few. That is, for habitat use patterns, we compared paddy fields (agricultural channel and cropland) with other landscape types in the wintering period (Appendix Fig. S1). For behavioral patterns, we compared foraging with the other behaviors in reservoir for both periods as enough individual behaviors were observed (n > 200) (Appendix Fig. S2). Descriptive analysis and plotting were carried out in R (R Core Team, 2022; Version 4.2.2).
We next examined how environmental features determined habitat use patterns in spoonbills using the species distribution models (SDMs). SDMs can calculate the probability of occupancy rates for any habitat/environmental variables with various forms, such as linear, logistic, and concave curves (Guisan et al., 2017). SDMs require species observation data (e.g., presence/absence or presence-only) and environmental data. Among many different SDMs, we employed the MaxEnt model, which follows the principle of maximum entropy (Jaynes, 1957), because it can deal with our presence-only data and approve a small sample situation as one of the generative models, unlike discriminative models (Phillips et al., 2006). Because of its versatility, the MaxEnt model has been used in many studies, including those focusing on waterbirds (Arakida et al., 2011; Wang et al., 2020) and Black-faced Spoonbills (Pickett et al., 2018).
For the wintering and migration periods, we constructed one MaxEnt model to examine the effects of environmental features on the spoonbill habitat use in each period. Hence, we contained all spoonbill counts as species data. Calculations were performed in the software MaxEnt (Phillips et al., 2022; Version 3.4.4), and the software setting used was basically the default (Phillips, 2017). The number of background points was 10,000. When a single grid had several presence points, we randomly kept just one point datum to minimize spatial autocorrelation. The regularization parameter was set to 1 as the default number. Models were calculated in 10-fold cross-validation. The effects of environmental features on the spoonbill habitat use were examined by response curves. We evaluated the MaxEnt model by the area under the curve (AUC) of the receiver operating characteristic (ROC) (Phillips et al., 2006). AUC has a value between 0 and 1, and a value close to 1 indicates reasonable/reliable performance, while a value < 0.5 indicates lower performance.
2.3.3
Environmental variables
We selected five environmental variables for the MaxEnt models, all of which can directly influence the habitat use patterns of spoonbills (Table 2): (1) landscape classification; (2) agricultural channel classification; (3) distance to tidal flat; (4) distance to artificial construction; and (5) distance to reed beds. Landscape classification was the same as in section 2.3.1. Agricultural channel classification was included to evaluate the types of agricultural channels because some spoonbills selectively used agricultural channels. In the classification, we conducted clustering analysis for 126 channels with 4 variables: width between walls, depth from the ground to the upper wall, presence/absence of vegetation, and channel wall features (Onikura et al., 2007) (Appendix Figs. S3 and S4). These four metrics were measured in the migration period and were used for the analysis in both migration and wintering periods. This is because the values of these metrics would not largely differ between and within seasons (Shimizu personal observation). More specifically, the presence/absence of vegetation in our study area was primarily decided by the substrate properties of agricultural channels such as soil (presence) and concrete (absence), and those substrate properties did not change throughout the study period. As a consequence of clustering, agricultural channels could be classified into five categories (Table 2; Appendix Figs. S5 and S6). Distances to preferable and undesirable environments likely affect habitat use patterns of spoonbills. We regarded the tidal flat and reed beds as preferable habitats since the tidal flat is the main foraging site and spoonbills frequently use shallow water zones with reed beds as roosting sites in Japan (Chan et al., 2010). We also regarded artificial construction as an undesirable environment for the primary sources of artificial disturbances (Yu and Swennen, 2004a). These three distance metrics were calculated using the Euclidean distance. The spatial resolution of all variables was 2 m × 2 m. Before implementing the MaxEnt model, the collinearity between variables was evaluated by the variance inflation factor (VIF). Because the VIF values among variables were < 2.2, we kept all variables in the MaxEnt models.
3.
Results
3.1
Habitat use and behavioral patterns of spoonbills
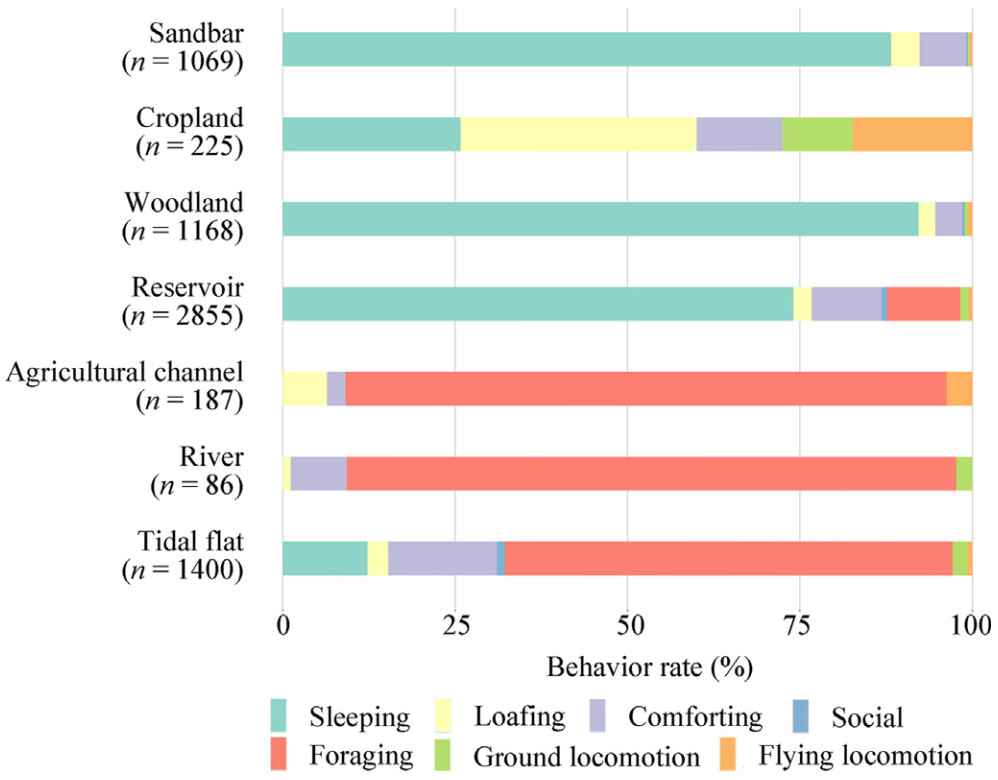
We observed a total of 1209 and 520 spoonbills in the route surveys and 4009 and 2924 spoonbills in behavioral surveys in the wintering and migration periods, respectively. The mean number of observed spoonbills per survey time in route surveys was 20.5 (±5.9 SD, range: 9–32) and 15.3 (±7.0 SD, range: 1–30) in the wintering and migration periods, respectively. A total of 91.3% and 75.4% of spoonbills were observed in the surrounding environments in the wintering and migration periods, respectively. Even when the tidal situation was low tides, 82.8% and 61.6% of spoonbills were observed in the surrounding environments in the wintering and migration periods, respectively (Fig. 2). Spoonbills used multiple types of artificial habitats in both periods: cropland, woodland, reservoir, and agricultural channel (Fig. 2). While foraging behavior was dominant in river (88.4%), agricultural channel (87.2%), and tidal flat (64.9%), sleeping behavior was dominant in woodland (92.2%), sandbar (88.2%), and reservoir (74.0%) (Fig. 3).
Figure
3.
Rates of different spoonbill behaviors in each landscape type. The number in parentheses in the bar charts shows the count of observed individuals.
However, habitat use patterns varied slightly between the periods. In the wintering period, 9.2% and 9.1% of spoonbills were observed in cropland and agricultural channel, respectively, but in the migration period, only 1.2% and 1.2% of spoonbills were observed in these environments. Furthermore, in a reservoir located south of the Imazu tidal flat, 45.3% of spoonbills were observed in the wintering period, but no spoonbills were observed in the migration period (Fig. 2), possibly because the water depth was too deep to utilize the reservoir in the migration period (Shimizu personal observation).
3.2
Relationship between environmental variables and habitat use patterns
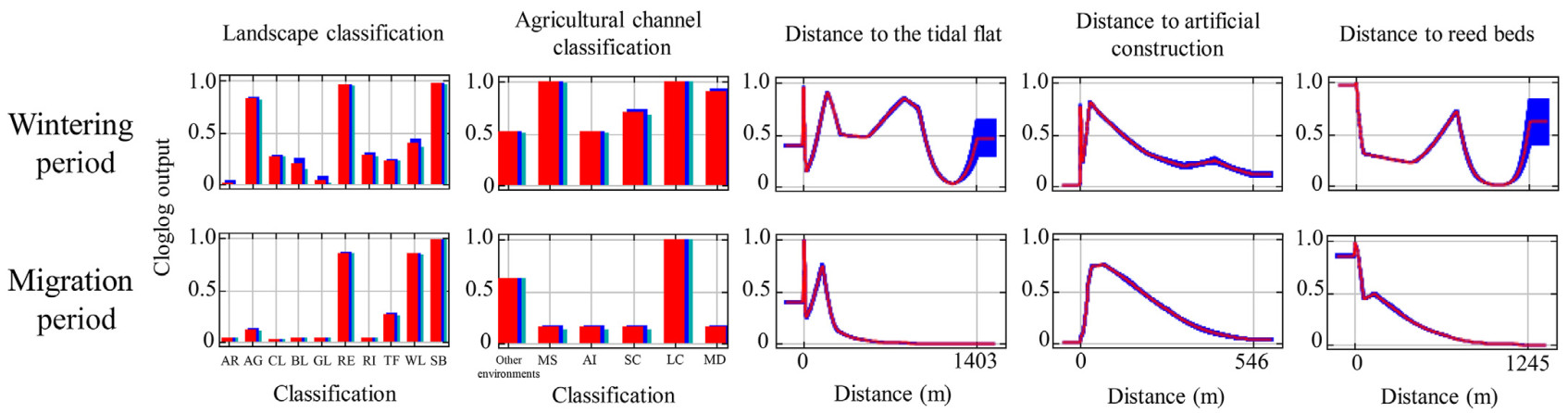
The average AUC values of the MaxEnt models (±SD) were 0.886 ± 0.021 in the wintering period and 0.939 ± 0.017 in the migration period. For the wintering period, the variables with higher contribution rates were landscape classification (32.1%), distance to reed beds (29.6%), distance to artificial construction (16.3%), agricultural channel classification (12.7%), and distance to tidal flat (9.3%). The most suitable landscape types were agricultural channel, reservoir, and sandbar (Fig. 4). For agricultural channel classification, midsized shallow channel and large channel showed relatively high suitability. The response curve of distance to tidal flat and reed beds indicated that the habitat suitability gradually declined away from tidal flat and reed beds but rapidly increased at some distances (Fig. 4). Regarding distance to artificial construction, spoonbills favored habitats close to artificial construction (Fig. 4).
Figure
4.
Response curve of each environmental variable in the wintering period and migration period. For the categorical variables, red bars show the mean values by 10-fold cross-validation, and blue and green bars represent the positive and negative standard deviations, respectively. For continuous variables, red lines show the mean values by 10-fold cross-validation, and blue lines are±standard deviations. (For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)
For the migration period, the variables with higher contribution rates were landscape classification (45.8%), distance to artificial construction (21.7%), distance to reed beds (18.9%), distance to tidal flat (10.2%), and agricultural channel classification (3.5%). The most suitable landscape type was sandbar, followed by reservoir and woodland (Fig. 4). Although the contribution of agricultural channel classification was low, large channel showed the highest suitability. Overall, habitat suitability rapidly decreased away from tidal flat, artificial construction, and reed beds compared with the wintering period (Fig. 4).
3.3
Interspecific differences in habitat use and behavioral patterns
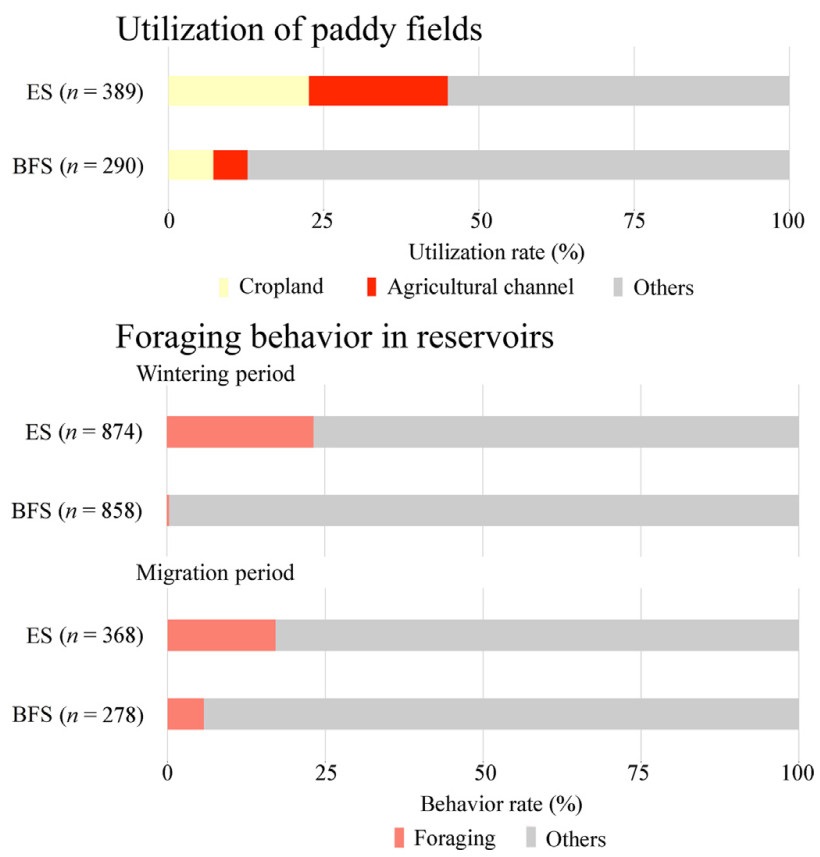
Considering that paddy fields consist of cropland and agricultural channel, there was a significant interspecific difference in habitat use patterns (i.e., paddy fields vs. others) in the wintering period (chi-square test, p < 0.01). Specifically, while 22.6% and 22.4% of Eurasian Spoonbills were observed in cropland and agricultural channel, respectively, only 7.2% and 5.5% of Black-faced Spoonbills were observed in these landscape types (Fig. 5). Moreover, there was a significant interspecific difference in foraging behavior (i.e., foraging vs. other behaviors) in reservoir in both periods (Chi-square test, p < 0.01). That is, while 23.4% and 17.1% of Eurasian Spoonbills were foraging in reservoir in the wintering and migration periods, respectively, only 0.4% and 5.8% of Black-faced Spoonbills were foraging in the landscape type (Fig. 5). On the other hand, 20.0% and 27.8% of Black-faced Spoonbills were observed in tidal flat in the wintering and migration periods, respectively, while 4.4% and 17.7% of Eurasian Spoonbills were observed in the landscape type (Appendix Fig. S1). However, we should note that the proportion of unidentified spoonbills was relatively high in tidal flat, especially during the migration period (Appendix Fig. S1). Additionally, no other landscape types were used for foraging by Black-faced Spoonbills more than Eurasian Spoonbills (Appendix Fig. S2).
Figure
5.
Interspecific difference in habitat use and behavioral patterns. ES means Eurasian Spoonbill, and BFS means Black-faced Spoonbill. The number in parentheses in the bar charts shows the count of observed individuals.
We also observed two types of intra- and interspecific interactions. The first was an aggressive behavior between two Eurasian Spoonbills (Appendix video 1). More specifically, an adult Eurasian Spoonbill drove away conspecific juveniles among 20 spoonbills in the reservoir. The second was an intraspecific interaction in roosting. We observed that a black-faced and Eurasian Spoonbill preened each other (Appendix video 2).
This study documented habitat use patterns between a tidal flat and the surrounding multitype habitats in two spoonbill species during the migration and wintering periods. We found that spoonbills relied on the surrounding habitats including artificial ones more frequently than the tidal flat regardless of tidal conditions in both periods. Additionally, Eurasian Spoonbills used croplands and agricultural channels and foraged in reservoirs more frequently than Black-faced Spoonbills.
4.1
Importance of the habitats surrounding the tidal flat
Previous research has shown that coastal waterbirds utilize various types of surrounding artificial habitats for resting and foraging when tidal flats are unavailable during high tide (Rogers, 2003; Sripanomyom et al., 2011; Bai et al., 2018). However, our results showed that spoonbills forage and rest in surrounding habitats even at low tides. Two reasons would explain our results. First, the activities of spoonbills are not tightly linked to tidal cycles; they are most active to forage during crepuscular times (Yu and Swennen, 2004b) and rest during the remaining daytime. Second, surrounding habitats are not subject to tidal disturbance, and hence would be more suitable roosting sites than the tidal flat. Therefore, our research suggests that some types of surrounding habitats would play a critical role in not only gaining food but saving energy expenditure for waterbirds throughout the day or a longer period (Rogers, 2003; Chen et al., 2021).
Consistent with previous results, our study showed that spoonbills frequently used artificial wetlands with shallow water depths below the tibia of spoonbills adjacent to reed beds and lands with no or low vegetation (Yu and Swennen, 2004a; Chan et al., 2010; Pigniczki and Végvári, 2015). In addition to these, we found that spoonbills also used other types of habitats, such as woodlands, croplands, and agricultural channels. In particular, foraging in agricultural channels in the wintering season was rarely documented (but see Fujioka et al., 2010). This seasonal difference of the agricultural channel utilization in spoonbills might be explained by three possible factors or combinations of them: 1) the low prey availability of the tidal flat caused by low biomass in winter (Beukema, 1974); 2) the high availability of flooded agricultural channels in winter (e.g., high concentration of fish in small areas) (Onikura et al., 2007; Fujioka et al., 2010); and 3) different habitat preferences between migration and wintering spoonbill populations (Takano et al., 2014; Jia et al., 2020). While Black-faced Spoonbills tend to feed in aquacultural ponds in other regions such as Chinese mainland and Taiwan (Sung et al., 2018; Cheng et al., 2022), water reservoirs in the Imazu region were not primarily foraging sites, especially for Black-faced Spoonbills. This regional difference might result from the variations of prey availability in these habitats; water reservoirs in Japan are simply established to reserve waters and are not made for farming fish or crustaceans, unlike aquacultural ponds in other countries. This suggests that water reservoirs in Japan would be unsuitable for foraging of spoonbills. Additional work needs to determine how habitat use patterns of spoonbills in the study area are related to prey availability in different types of artificial wetlands including agricultural channels and water reservoirs. Furthermore, we should elucidate how and why these relationships are different between regions/countries.
4.2
Interspecific differences in habitat use patterns
As we expected, Eurasian Spoonbills used various surrounding artificial habitats more frequently than Black-faced Spoonbills, especially for foraging. Eurasian Spoonbills breed in inland natural wetlands (Pigniczki and Végvári, 2015; Tiunov, 2021), which may connect them to utilize or forage at various artificial habitats in addition to coastal wetlands in non-breeding seasons. Although aquacultural ponds can supply Black-faced Spoonbills with enough amount of prey items in some regions (Sung et al., 2018; Cheng et al., 2022), our results suggest that degradation of tidal flats would have a larger negative impact on Black-faced Spoonbills than Eurasian Spoonbills, especially in regions without suitable aquacultural ponds. Therefore, practical conservation should concurrently focus on tidal flats for foraging and the surrounding habitats for resting, especially for Black-faced Spoonbills.
Although some individuals of Eurasian Spoonbills and Black-faced Spoonbills are known to use inland areas (Wood et al., 2013; Son et al., 2020; Chen et al., 2021; Xi et al., 2021), the difference in habitat use patterns between the two sympatric species has not been clarified before. This interspecific difference might be caused by interspecific variations in preferred prey items because prey distributions between tidal flats and surrounding habitats are likely to differ considerably. However, no information about this hypothesis is available. In contrast, few interspecific differences in using roosting sites may be explained by the narrow variations in roost site environments, which might allow two similar body-sized species to roost in the same site.
4.3
Limitations
This study has some limitations regarding the study design and inferences of statistical models. For the study design, we should point out three limitations. First, our research was conducted in a single site although the results agreed with earlier research in Japan (Chan et al., 2010). Since compositions and configurations of surrounding environments vary not only between East Asian countries but also between different sites in Japan, it is important to investigate habitat use patterns of spoonbills in other regions. Second, our surveys recorded many unidentified spoonbills when spoonbills were sleeping (Appendix Figs. S1 and S2). Therefore, we may have underestimated the interspecific differences in habitat use patterns, especially at the roosting sites. Third, we could not record the ages of spoonbills, which is an important factor for habitat use in spoonbills. For example, juvenile birds are known to use various habitats more than adult birds (Pigniczki and Végvári, 2015; Chen et al., 2021). Therefore, we cannot exclude the possibility that the documented interspecific difference in habitat use patterns was caused by the different age structures between the two focal species.
The limitation regarding statistical model inferences concerns the violated assumptions of our MaxEnt models, which might bias the model outputs (Guisan et al., 2017). For example, we did not account for spatial and temporal autocorrelation due to technical constraints. However, the influence of autocorrelation on models remains unknown, especially when focusing on species forming aggregations and narrow survey areas (Phillips et al., 2017), and fine-scale research can improve the model accuracy due to precise locations and environmental data (Gottschalk et al., 2011). Furthermore, because autocorrelation is important to explain the processes of ecological systems (MacKenzie et al., 2018), the results are reliable for understanding the habitat use patterns of spoonbills.
4.4
Conclusion and implications for conservation
This research showed that the two spoonbill species frequently used various surrounding habitats including artificial ones around the tidal flat for different behavioral purposes. This suggests that we should concurrently conserve tidal flats and surrounding habitats for the daily activities of those species, especially Black-faced Spoonbills. Notwithstanding, the surrounding habitat conditions around the Imazu region have not been improved but rather have been degraded. For example, garbage, including lures and fishing nets, accumulates in surrounding habitats, which injures spoonbills. Another issue is sandbar shrinking possibly due to the unbalanced inflow and outflow of sediments. Moreover, croplands have been shrinking due to the conversion to residential buildings and solar panels for electricity generation. These issues may be common in other regions of Japan and across the world (Sakamura et al., 2014; Murray et al., 2015; Melville et al., 2016). It is important to evaluate the impact of habitat loss and degradation of surrounding habitats on coastal waterbird populations at broader spatial scales.
On the other hand, effective management and conservation interventions can improve the quality of artificial habitats (Cheng et al., 2022; Cheng and Ma, 2023). Previous research showed that the number of shorebirds increased by manipulating the water depth of aquacultural ponds in China (Dai et al., 2021). Hence, knowing about further detailed habitat use patterns in artificial environments would enable us to make the conservation interventions more effective. Specific future challenges relevant to this include how functions of surrounding habitats (e.g., roosting sites) vary between the day and night and how waterbirds respond to interventions in artificial habitats (e.g., controlling the water depth in water reservoirs, planting reeds, and establishing wide non-concrete agricultural channels). Regardless of the outcomes of these challenges, our research emphasizes that conservation interventions of tidal flats as well as surrounding artificial habitats may maximize the effects of total conservation outcomes for various coastal waterbirds.
Authors’ contributions
TS and YF conceived the study. SS gathered important information and connected to key people. TS collected data and carried out the analyses. TS and MS wrote the first draft of the manuscript. YF and SS revised the manuscript. All authors read and approved the final manuscript.
Ethics statement
Observations were only carried out on public roads, and observers did not obstruct any passage or traffic. Disturbances due to observations were kept as low as possible.
Availability of data and materials
The raw sequence data reported in this papar have been deposited in the Figshare (Shimizu et al., 2022).
Declaration of competing interest
We have no competing interest to be declared.
Acknowledgments
We thank Satoru Matsumoto (Japan Black-faced Spoonbill Network) for help during some parts of the surveys. We are also grateful to Dr. Toru Nakahara for advice on the research plan.
Amano, T., Székely, T., Koyama, K., Amano, H., Sutherland, W.J., 2010. A framework for monitoring the status of populations: An example from wader populations in the East Asian-Australasian flyway. Biol. Conserv. 143, 2238–2247. .
Arakida, H., Mitsuhashi, H., Kamada, M., Koyama, K., 2011. Mapping the potential distribution of shorebirds in Japan: the importance of landscape-level coastal geomorphology. Aquat. Conserv. 21, 553–563. .
Bai, M.L., Chih, W.C., Lai, Y.C., Lee, P.F., Lien, Y.Y., 2018. Aquaculture ponds as important high-tide habitats for waterbirds along the west coast of Taiwan. Ornithol. Sci. 17, 55–67. .
Beukema, J.J., 1974. Seasonal changes in the biomass of the macro-benthos of a tidal flat area in the Dutch Wadden Sea. Neth. J. Sea Res. 8, 94–107. .
BirdLife International, 2017. Platalea minor. The IUCN Red List of Threatened Species 2017: e.T22697568A119347801. (Accessed 12 December 2022).
BirdLife International, 2019. Platalea leucorodia (Amended Version of 2016 Assessment). . The IUCN Red List of Threatened Species 2019: eT22697555A155460986. (Accessed 12 December 2022).
Chan, S., Fang, W.H., Lee, K.S., Yamada, Y., Yu, Y.T., 2010. International Single Species Action Plan for the Conservation of the Black-Faced Spoonbill (Platalea Minor). BirdLife International Asia Division, Tokyo. CMS Secretariat, Bonn. .
Chen, Y., Yu, Y.T., Meng, F., Deng, X., Cao, L., Fox, A.D., 2021. Migration routes, population status and important sites used by the globally threatened Black-faced Spoonbill (Platalea minor): a synthesis of surveys and tracking studies. Avian Res. 12, 74. .
Cheng, C., Liu, J., Ma, Z., 2022. Effects of aquaculture on the maintenance of waterbird populations. Conserv. Biol. 36, e13913. .
Cheng, C., Ma, Z., 2023. Conservation interventions are required to improve bird breeding performance in artificial wetlands. Biol. Conserv. 278, 109872. .
Choi, C.Y., Nam, H.Y., Lee, W.S., 2007. Measuring the behaviors of wintering Black-faced Spoonbills (Platalea minor): comparison of behavioral sampling techniques. Waterbirds 30, 310–316. .
Clemens, R., Rogers, D.I., Hansen, B.D., Gosbell, K., Minton, C.D.T., Straw, P., et al., 2016. Continental-scale decreases in shorebird populations in Australia. Emu 116, 119–135. .
Dai, W., Goodale, E., He, R., Mammides, C., Liu, S., Zhou, L., et al., 2021. An eco-compensation policy increases shorebird diversity during the non-farming period for aquaculture. Wetlands 41, 4. .
Dias, M.P., Granadeiro, J.P., Lecoq, M., Santos, C.D., Palmeirim, J.M., 2006. Distance to high-tide roosts constrains the use of foraging areas by dunlins: Implications for the management of estuarine wetlands. Biol. Conserv. 131, 446–452. .
ESRI, 2022. ArcGIS Pro, Version 2.7.0. . (Accessed 12 December 2022).
Fujioka Jr., M., W, A.J., Yoshida, H., Maeda, T., 2001. Value of fallow farmlands as summer habitats for waterbirds in a Japanese rural area. Ecol. Res. 16, 555–567. .
Fujioka, M., Lee, S.D., Kurechi, M., Yoshida, H., 2010. Bird use of rice fields in Korea and Japan. Waterbirds 33, 8–29. .
Gottschalk, T.K., Aue, B., Hotes, S., Ekschmitt, K., 2011. Influence of grain size on species-habitat models. Ecol. Model. 222, 3403–3412. .
Guisan, A., Thuiller, W., Zimmermann, N.E., 2017. Habitat Suitability and Distribution Models with Applications in R. Cambridge University Press, Cambridge. https://www.google.co.jp/books/edition/Habitat_Suitability_and_Distribution_Mod/rYswDwAAQBAJ?hl=ja&gbpv=0.
Hancock, J., Kushlan, J.A., Kahl, M.P., 2011. Storks, Ibises and Spoonbills of the World, Digital. Christopher Helms Publisher, London. https://www.google.co.jp/books/edition/Storks_Ibises_and_Spoonbills_of_the_Worl/w2lCFQ7-Il8C?hl=ja&gbpv=0.
Hong Kong Bird Watching Society, 2022. Black-faced Spoonbill Population Hits Record High of 6,000. . (Accessed 12 December 2022).
Jackson, M.V., Carrasco, L.R., Choi, C.Y., Li, J., Ma, Z., Melville, D.S., et al., 2019. Multiple habitat use by declining migratory birds necessitates joined up conservation. Ecol. Evol. 9, 2505–2515. .
Jackson, M.V., Choi, C.Y., Amano, T., Estrella, S.M., Lei, W., Moores, N., et al., 2020. Navigating coasts of concrete: Pervasive use of artificial habitats by shorebirds in the Asia-Pacific. Biol. Conserv. 247, 108591. .
Jackson, M.V., Woodworth, B.K., Bush, R., Clemens, R.S., Fuller, R.A., Garnett, S.T., et al., 2021. Widespread use of artificial habitats by shorebirds in Australia. Emu 121, 187–197. .
Jaynes, E.T., 1957. Information theory and statistical mechanics. Phys. Rev. 106, 620–630. .
Jia, R., Liu, D., Lu, J., Zhang, G., 2020. Wetland destruction on migration routes threatens a breeding population of the endangered black-faced spoonbill (Platalea minor). Glob. Ecol. Conserv. 23, e01105. .
Kang, J.H., Kim, I.K., Lee, K.S., Lee, H., Rhim, S.J., 2016. Distribution, breeding status, and conservation of the black-faced spoonbill (Platalea minor) in South Korea. For. Sci. Technol. 12, 162–166. .
Li, D., Chen, S., Lloyd, H., Zhu, S., Shan, K., Zhang, Z., 2013. The importance of artificial habitats to migratory waterbirds within a natural/artificial wetland mosaic, Yellow River Delta, China. Bird Conserv. Int. 23, 184–198. .
Ma, Z., Li, B., Zhao, B., Jing, K., Tang, S., Chen, J., 2004. Are artificial wetlands good alternatives to natural wetlands for waterbirds? - A case study on Chongming Island, China. Biodivers. Conserv. 13, 333–350. .
MacKenzie, D.I., Nichols, J.D., Royle, J.A., Pollock, K.H., Bailey, L.L., Hine, J.E., 2018. Occupancy Estimation and Modeling: Inferring Patterns and Dynamics of Species Occurrence, second ed. Academic Press, London https://www.google.co.jp/books/edition/Occupancy_Estimation_and_Modeling/hs2cBAAAQBAJ?hl=ja&gbpv=0.
Melville, D.S., Chen, Y., Ma, Z., 2016. Shorebirds along the Yellow Sea coast of China face an uncertain future-a review of threats. Emu 116, 100–110. .
Murray, N.J., Ma, Z., Fuller, R.A., 2015. Tidal flats of the Yellow Sea: A review of ecosystem status and anthropogenic threats. Austral Ecol. 40, 472–481. .
National Astronomical Observatory of Japan, 2021. Local Calendar for Fukuoka (Fukuoka). . (Accessed 12 December 2022).
NEAR-GOOS, 2021. 30 Sec. Interval Tide Data at Hakata, RDMDB Data Retrieval System. . (Accessed 20 February 2023) (In Japanese).
Onikura, N., Nakajima, J., Eguchi, K., Miyake, T., Nishida, T., Inui, R., et al., 2007. Relationships between presence and populations of freshwater fishes and revetment conditions in creeks around sea of Ariake, northwestern kyushu, Japan. J. Jpn. Soc. Water Environ. 30, 277–282. (In Japanese).
Phillips, S.J., Anderson, R.P., Schapire, R.E., 2006. Maximum entropy modeling of species geographic distributions. Ecol. Model. 190, 231–259. .
Phillips, S.J., Anderson, R.P., Dudík, M., Schapire, R.E., Blair, M.E., 2017. Opening the black box: an open-source release of Maxent. Ecography 40, 887–893. .
Phillips, S.J., 2017. A Brief Tutorial on Maxent. . (Accessed 12 December 2022).
Phillips, S.J., Dudík, M., Schapire, R.E., 2022. Maxent Software for Modeling Species Niches and Distribution, Version 3.4.4. . (Accessed 12 December 2022).
Pickett, E.J., Chan, M., Cheng, W., Allcock, J., Chan, S., Hu, J., et al., 2018. Cryptic and cumulative impacts on the wintering habitat of the endangered black-faced spoonbill (Platalea minor) risk its long-term viability. Environ. Conserv. 45, 147–154. .
Pigniczki, C., Végvári, Z., 2015. Dispersal of the central European population of the Eurasian spoonbill Platalea leucorodia. Ardeola 62, 219–236. .
R Core Team, 2022. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Version 4.2.2. . (Accessed 12 December 2022).
Ramsar Convention on Wetlands, 2018. Global Wetland Outlook: State of the World's Wetlands and Their Services to People. Ramsar Convention Secretariat. Gland. .
Rogers, D.I., 2003. High-tide roost choice by coastal waders. Wader Study Group Bull. 100, 73–79. .
Rogers, D.I., Battley, P.F., Piersma, T., Van Gils, J.A., Rogers, K.G., 2006. High-tide habitat choice: insights from modelling roost selection by shorebirds around a tropical bay. Anim. Behav. 72, 563–575. .
Rosa, S., Encarnação, A.L., Granadeiro, J.P., Palmeirim, J.M., 2006. High water roost selection by waders: maximizing feeding opportunities or avoiding predation? Ibis 148, 88–97. .
Sakamura, K., Kaneko, T., Numata, M., Nakai, N., 2014. Study on the site character of the ground standing Photovoltaic Power Generating System. J. City Plan. Inst. Jpn. 49, 633–638. (In Japanese).
Son, S.J., Kang, J.H., Lee, S.K., Kim, I.K., Yoo, J.C., 2020. Breeding and wintering home ranges of the black-faced spoonbill Platalea minor. J. Asia Pac. Bus. 13, 7–12. .
Sripanomyom, S., Round, P.D., Savini, T., Trisurat, Y., Gale, G.A., 2011. Traditional salt-pans hold major concentrations of overwintering shorebirds in Southeast Asia. Biol. Conserv. 144, 526–537. .
Studds, C.E., Kendall, B.E., Murray, N.J., Wilson, H.B., Rogers, D.I., Clemens, R.S., et al., 2017. Rapid population decline in migratory shorebirds relying on Yellow Sea tidal mudflats as stopover sites. Nat. Commun. 8, 14895. .
Sullender, B.K., Barzen, J., Silbernagel, J., 2016. Foraging success and habitat selection of the Eurasian Spoonbill (Platalea leucorodia) at Poyang Lake, China. Waterbirds 39, 356–364. .
Sung, Y.H., Tse, I.W.L., Yu, Y.T., 2018. Population trends of the Black-faced Spoonbill Platalea minor: analysis of data from international synchronised censuses. Bird Conserv. Int. 28, 157–167. .
Shimizu, T., Senzaki, M., Fujioka, Y., Seino, S., 2022. Dataset for spoonbills habitat use in the Imazu tidal flat and surrounding environments. Figshare. .
Takano, S., Takeshita, F., Henmi, Y., 2014. Spatiotemporal utilization of feeding sites by the black-faced spoonbill Platalea minor in the Yatsushiro Sea, Japan. Ornithol. Sci. 13, 59–66. .
Tiunov, I., 2021. Nesting of the black-faced spoonbill (Platalea minor) (Pelecaniformes: Threskiornithidae) and the hybrid partner on the coasts of Khanka Lake (the Russian Federation). J. Asia-Pac. Biodivers. 14, 111–115. .
Wang, G., Wang, C., Guo, Z., Dai, L., Wu, Y., Liu, H., et al., 2020. A multiscale approach to identifying spatiotemporal pattern of habitat selection for red-crowned cranes. Sci. Total Environ. 739, 139980. .
Wang, X., Li, X., Ren, X., Jackson, M.V., Fuller, R.A., Melville, D.S., et al., 2022. Effects of anthropogenic landscapes on population maintenance of waterbirds. Conserv. Biol. 36, e13808. .
Wei, G.-A., Lei, F.-M., Yin, Z.-H., Ding, C.-Q., Ding, W.-N., 2005. Nesting and disturbance of the Black-faced Spoonbill in Liaoning Province, China. Waterbirds 28, 420–425. .
Wood, C., Tomida, H., Jin-Han, K., Lee, K.S., Cho, H.J., Nishida, S., et al., 2013. New perspectives on habitat selection by the Black-faced Spoonbill Platalea minor based upon satellite telemetry. Bird Conserv. Int. 23, 495–501. .
Xi, J., Deng, X., Zhao, G., Batbayar, N., Damba, I., Zhao, Q., et al., 2021. Migration routes, behavior and protection status of Eurasian spoonbills (Platalea leucorodia) wintering in China. Avian Res. 12, 70. .
Yasué, M., Dearden, P., 2009. The importance of supratidal habitats for wintering shorebirds and the potential impacts of shrimp aquaculture. Environ. Manag. 43, 1108–1121. .
Yu, Y.T., Swennen, C.K., 2004a. Habitat use of the Black-faced Spoonbill. Waterbirds 27, 129–134. .
Yu, Y.T., Swennen, C.K., 2004b. Feeding of wintering Black-faced Spoonbills in Hong Kong: when and how long? Waterbirds 27, 135–140. .
Table
1.
Summary of the route surveys in the two periods.
Total
Tidal situation
Time zone
High tide
Low tide
Crepuscular
Morning
Afternoon
Wintering period
Survey times
59
27
32
17
20
22
Survey days
40
26
28
16
19
19
Rate to total survey times
45.8%
54.2%
28.8%
33.9%
37.3%
Migration period
Survey times
34
16
18
13
9
12
Survey days
30
16
18
12
9
12
Rate to total survey times
47.1%
52.9%
38.2%
26.5%
35.3%
The tidal situation is divided into two based on 190-cm tide level since spoonbills could forage in the tidal flat below 190 cm. The time period is divided into three (National Astronomical Observatory of Japan, 2021): crepuscular (1 h before dusk and after dawn), morning (after crepuscular before noon) and afternoon (after noon before crepuscular).

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds