Hem Bahadur Katuwal, Hari Prasad Sharma, Prashant Rokka, Krishna Prasad Bhusal, Bishnu Prasad Bhattarai, Sabina Koirala, Sandeep Chhetri Luitel, Shailendra Yadav, Ganesh Sah, Hem Sagar Baral, Laxman Prasad Poudyal, Lin Wang, Rui-Chang Quan. 2023: The effects of climate and land use change on the potential distribution and nesting habitat of the Lesser Adjutant in Nepal. Avian Research, 14(1): 100105. DOI: 10.1016/j.avrs.2023.100105
Citation:
Hem Bahadur Katuwal, Hari Prasad Sharma, Prashant Rokka, Krishna Prasad Bhusal, Bishnu Prasad Bhattarai, Sabina Koirala, Sandeep Chhetri Luitel, Shailendra Yadav, Ganesh Sah, Hem Sagar Baral, Laxman Prasad Poudyal, Lin Wang, Rui-Chang Quan. 2023: The effects of climate and land use change on the potential distribution and nesting habitat of the Lesser Adjutant in Nepal. Avian Research, 14(1): 100105. DOI: 10.1016/j.avrs.2023.100105
Hem Bahadur Katuwal, Hari Prasad Sharma, Prashant Rokka, Krishna Prasad Bhusal, Bishnu Prasad Bhattarai, Sabina Koirala, Sandeep Chhetri Luitel, Shailendra Yadav, Ganesh Sah, Hem Sagar Baral, Laxman Prasad Poudyal, Lin Wang, Rui-Chang Quan. 2023: The effects of climate and land use change on the potential distribution and nesting habitat of the Lesser Adjutant in Nepal. Avian Research, 14(1): 100105. DOI: 10.1016/j.avrs.2023.100105
Citation:
Hem Bahadur Katuwal, Hari Prasad Sharma, Prashant Rokka, Krishna Prasad Bhusal, Bishnu Prasad Bhattarai, Sabina Koirala, Sandeep Chhetri Luitel, Shailendra Yadav, Ganesh Sah, Hem Sagar Baral, Laxman Prasad Poudyal, Lin Wang, Rui-Chang Quan. 2023: The effects of climate and land use change on the potential distribution and nesting habitat of the Lesser Adjutant in Nepal. Avian Research, 14(1): 100105. DOI: 10.1016/j.avrs.2023.100105
Center for Integrative Conservation, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Mengla, Yunnan, 666303, China. E-mail address: quanrc@xtbg.ac.cn (R.-C. Quan)
Climate change and land use change pose a threat to the world's biodiversity and have significant impacts on the geographic distribution and composition of many bird species, but little is known about how they affect threatened large-sized waterbird species that rely on agricultural landscapes. To address this gap, we investigated how climate and land use changes influence the distribution and nesting habitats of the globally vulnerable Lesser Adjutant (Leptoptilos javanicus) in Nepal. Between 2012 and 2023, we collected distribution data from 24 districts and nesting site information from 18 districts. In a nation-wide breeding survey conducted in 2020, we documented a total of 581 fledglings from 346 nests in 109 colonies. The ensemble model predicted a current potential distribution of 15% (21,637 km2) and a potential nesting habitat of 13% (19,651 km2) for the species in Nepal. The highest predicted current suitable distribution and nesting habitat was in Madhesh Province, while none was predicted in Karnali Province. The majority of this predicted distributional and nesting habitat falls on agricultural landscapes (> 70%). Our model showed a likely range expansion of up to 15% (21,573 km2) for the distribution and up to 12% (17,482 km2) for the nesting habitat under SSP5–8.5 scenarios for the 2070s. The range expansion is expected to occur mainly within the current distribution and breeding range (Tarai and some regions of Siwalk), particularly in Lumbini and Sudurpashchim provinces, and extend to the northern portions (Siwalik and Mid-hill regions) in other provinces. However, the current Protected Areas and Important Bird and Biodiversity Areas are inadequate for providing optimal habitats for the species. Although the model suggests range expansion, the use of such novel habitats is primarily contingent on the availability and protection of large-sized trees (particularly Bombax ceiba, observed in 65% of colonies) in agricultural regions where nesting occurs. Therefore, our research suggests that agricultural landscapes should be prioritized in management plans for the conservation of the Lesser Adjutant in Nepal.
Climate change, coupled with land use change, poses a threat to the world's biodiversity (Jetz et al., 2007; Garcia et al., 2014; Fusco et al., 2021; Kattel, 2022). These changes have not only affected the current geographic range and composition of wildlife species, but also their future distribution (Walther et al., 2002; Bellard et al., 2012; Trautmann, 2018; Adhikari et al., 2022). Climate and land use change are anticipated to have far-reaching impacts on both tropical and Himalayan ecosystems, posing significant conservation challenges for many species (Jetz et al., 2007; Wiens, 2016; Sharma et al., 2020; Baral et al., 2023). The foraging and breeding patterns of farmland birds have already been affected (Santangeli et al., 2018; Liu et al., 2021). However, the effect of climate and land use change-driven habitat modification on farmland bird distributions, particularly large waterbirds such as storks in agricultural landscapes, is still poorly understood. This knowledge gap has led to several conservation challenges for the stork species (BirdLife International, 2021; Gula et al., 2023). Therefore, understanding the impact of climate and land use change on large breeding waterbirds would help in implementing conservation strategies in agricultural landscapes.
The Lesser Adjutant (Leptoptilos javanicus; Fig. 1) is one of the largest storks native to South and Southeast Asia (Elliott et al., 2020; BirdLife International, 2021). The species is globally listed as vulnerable due to rapid population declines caused by several anthropogenic threats, including hunting, loss of nesting trees, conversion and degradation of wetlands, and agricultural changes and intensification (BirdLife International, 2021). Contrary to past studies, which indicated that the species mostly favors marsh and forest habitats, recent study in Nepal reveals that it prefers agricultural landscapes for foraging and breeding (Sundar et al., 2016; Koju et al., 2019; Katuwal et al., 2022b). It is a carnivorous species that feeds mostly on fish, frogs, snakes, rodents, larger invertebrates such as locusts and crabs, and sometimes carrion (Elliott et al., 2020). Breeding begins in July, and chicks fledge by December, but some chicks hatched later may still be observed in trees until March (Katuwal et al., 2022b). Nests are built on tall trees, such as Bombax ceiba, Haldina cordifolia, and Ficus religiosa, which are dispersed over agricultural areas (Karki and Thapa, 2013; Koju et al., 2019; Bhattarai et al., 2021; Katuwal et al., 2022b; Wagle et al., 2022). However, due to unplanned urbanization, agricultural landscapes are shrinking and are expected to decline further in the future (D'Amour et al., 2017; Rimal et al., 2020). In addition, large trees are felled for commercial and construction purposes. Thus, Lesser Adjutants face numerous anthropogenic threats, which are anticipated to rise in the future.
Figure
1.
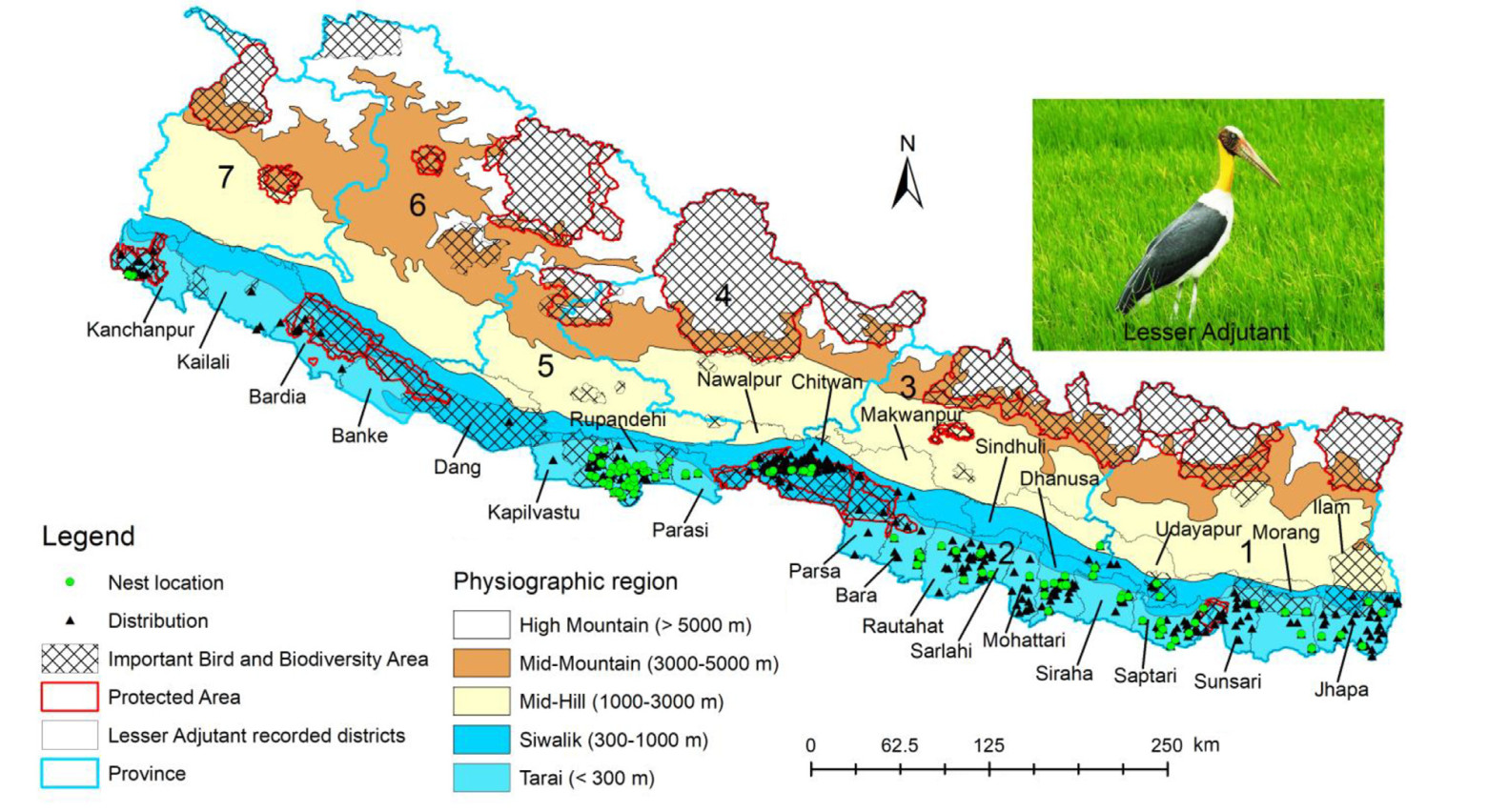
The distribution and nesting sites of Lesser Adjutants in Nepal from 2012 to 2023. Lesser Adjutants were recorded from 24 districts, whereas nesting sites from 18 districts. Despite the fact that Lesser Adjutant has an east-west distribution range, we were unable to find any recent records in Banke. Although there was a record in 1983 in eBird, we did not use this historical record as most of the environmental factors we considered have significantly changed since then. Additionally, we did not find any nest sites in the Parsa, Dang, Banke, Bardia and Kailali districts. In the map, 1–7 are the provinces of Nepal which also apply to Figs. 3–6 (1, Koshi Province; 2, Madhesh Province; 3, Bagmati Province; 4, Gandaki Province; 5, Lumbini Province; 6, Karnali Province; 7, Sudurpashchim Province).
Nepal holds the smallest breeding habitat for the Lesser Adjutant within South Asia, but unexpectedly, the majority of studies have been undertaken in Nepal (Katuwal et al., 2022b). Until recently, most Lesser Adjutant research focused on determining population status, breeding ecology, and threats in isolated locations (Inskipp et al., 2016; Katuwal et al., 2022b). However, little is known about other potential habitats, less information exists on its preferred nesting trees and breeding metrics across the Nepal. Moreover, the role of Protected Areas (PAs) and Important Bird and Biodiversity Areas (IBAs) in providing suitable habitats for the species are not well known, despite their establishment with the aim of conserving bird. In addition, emerging risks such as climatic and other anthropogenic threats have been underestimated in the case of the Lesser Adjutant. Other tropical species such as Sarus Cranes (Grus antigone) are already experiencing climate change impact (Sundar et al., 2018), but little is known about the Lesser Adjutant although it is expected to be affected by climate change (Khadka and Pandey, 2014). Because its breeding typically coincides with the onset of the monsoon season, the offspring of this species are vulnerable to colder temperatures and rainfall during the winter months (Khadka and Pandey, 2014; Katuwal et al., 2022b). Furthermore, its foraging and breeding habitat—which includes agricultural landscapes and wetlands—is expected to be negatively impacted by climate change and other human activities (Karki and Gurung, 2012; Lamsal et al., 2017; Chhogyel and Kumar, 2018; Rimal et al., 2020). As climate change is already impacting the monsoon and winter rain and Nepal is experiencing unpredictable weather patterns (Karki et al., 2017; Hamal et al., 2020) and both temperature and precipitation are expected to increases in future (Ghimire, 2019; Chapagain et al., 2021), it is crucial to monitor the effects of climate change on the Lesser Adjutant. Predicting the potential change of species distribution and responses to climate change coupled with other factors like land use change can provide information on the ecological monitoring for species conservation and management (Moradi et al., 2019; Chhetri et al., 2021).
This study aimed to investigate the nesting tree characteristics and breeding metrics of the Lesser Adjutant, as well as to gain a better understanding of the factors influencing the species' distribution and nesting habitat in Nepal. Specifically, our objectives were to: 1) predict the current suitable distribution and nesting habitat of the species across Nepal using bioclimatic and land use variables; 2) forecast the potential future distribution and nesting habitat based on various climate and land use change scenarios; and 3) evaluate the effectiveness of PAs and IBAs in providing optimal habitat for the species. Given the adverse impacts of climate and land use change on various bird species (Langham et al., 2015; Trautmann, 2018; Tehrani et al., 2020), we predicted that the distribution and nesting habitat of the Lesser Adjutant in Nepal would experience range contraction. This study may be the first large-scale investigation of Lesser Adjutants in their nesting territory. The findings will provide important information for the development of management and conservation plans for the species in Nepal, and may also serve as a model for other range countries.
2.
Materials and methods
2.1
Study area
Physiographically, Nepal is divided into five regions: Tarai (< 300 m), Siwalik (300–1000 m), Mid-Hill (1000–3000 m), Mid-Mountain (3000–5000 m), and High Mountain (above 5000 m; Fig. 1; LRMP, 1986). We surveyed the occurrences of the Lesser Adjutant throughout Nepal from 2012 to early 2023 using systematic field survey, opportunistic recordings, discussion with bird experts, and online databases. However, we confirmed their occurrences primarily in southern lowland Nepal, below 500 m elevation, because Lesser Adjutants are known to occur and breed at this elevation (60–400 m; Fig. 1; Inskipp et al., 2016; Baral et al., 2020). Although Nepal has a total of 20 PAs (core and buffer zone area) and 37 IBAs (DNPWC, 2022), only seven PAs and 16 IBAs fall within the breeding range of the species (Fig. 1).
The lowland landscape often includes flood plains of many river systems such as Koshi, Narayani, and Karnali, receiving a huge quantity of water during the monsoon season. Agriculture is the major source of income of people in this region, with some individuals working in animal husbandry, business, and overseas jobs. During the monsoon season (June–September), rice is the major crop grown throughout the region, but summer and winter crops differ (Katuwal et al., 2022a). In the winter, wheat, mustard, and a variety of vegetables are farmed, whereas in the summer, maize and rice are cultivated; sugarcane is only grown in certain places. After harvest in the winter or summer, fields are also kept fallow until the monsoon season. Shorea robusta is the dominant vegetation, Trewia nudiflora, Acacia catechu, and Bombax ceiba in riverine forest, and Saccharum and Themeda species are present in periodically flooded tall grasslands. Agroforestry is practiced by growing Eucalyptus spp., Bombax ceiba, Ficus spp., and other orchards in agricultural areas (Koju et al., 2019). The climate in the east is relatively wetter than in the west largely dictated by monsoonal rain. The monsoon season begins in June and lasts until September. The winter season begins after November and lasts until February. The winter is characterized by thick fog for a few weeks and, cold and rain for a few days.
2.2
Species occurrence data
2.2.1
Distributional habitat
We collected distribution data by systematic field surveys, opportunistic field surveys, and the ebird portal. During the Farmland Bird Survey Program from 2018 to 2019, we established 116 transects in farmland, river, and forest habitat in four districts (Kapilvastu, Chitwan, Sarlahi and Sunsari) of lowland Nepal (Katuwal et al., 2022a). The farmland transects were re-monitored in 2021 and 2022. This systematic investigation resulted in a total of 61 unique distribution points. In addition, we obtained 500 occurrence data from opportunistic sightings that we observed during the systematic survey, nesting survey, and other bird watching activities across Nepal between 2018 and 2023. We acquired some occurrences (n = 49) from ebird (https://ebird.org/species/lesadj1) from 2012 to 2023 to fill in the location gaps in a few mid (Dang and Bardia) western and far western (Kailali and Kanchanpur) and eastern (Morang and Jhapa) regions. We verified the uncertainties seen in the records with the authors before using the data. However, we did not find the recent record of Lesser Adjutant in Banke district, although ebird shows the presence in 1983. But, we did not use this historical record because most of the environmental factors we used have changed significantly. Altogether, we collected a total of 610 occurrences of Lesser Adjutant from 24 districts throughout six provinces (not found in Karnali Province; Fig. 1). However, we only used one point within 1 km × 1 km grid to avoid the spatial bias and spatial autocorrelation (Boria et al., 2014), which resulted a total of 425 occurrences points for modeling distribution.
2.2.2
Nesting site
Since Lesser Adjutants relocate some of their nesting sites within their habitat annually (HBK and PR personal observations), we included all colonies we identified between 2018 and 2023. A colony is defined as a nest or a group of nests with at least one chick in a single or clump of trees with contiguous canopies (Sundar et al., 2019; Katuwal et al., 2022b). In 2018, we monitored nine colonies in Chitwan (Bhattarai et al., 2021), and in 2019, we systematically examined 65 colonies from six districts (Kapilvstu, Rupandehi, Bara, Sarlahi, Udayapur, and Morang) and opportunistically examined 13 colonies from five other districts (Chitwan, Siraha, Saptari, Sunsari, and Jhapa) (Katuwal et al., 2022b). In addition, we conducted a nation-wide breeding survey for the Lesser Adjutant in 2020, including areas examined in 2018 and 2019. We monitored the colonies from July to December, visiting 1–2 times per month, and recorded opportunistic colonies until March. During the nation-wide survey, we discovered a total of 109 colonies in lowland Nepal (Fig. 1). Furthermore, we identified and monitored 72 colonies from 2021 to 2023, including previously known sites and new ones like Mohattari. For our analysis, we also included one location from Sindhuli (Baral et al., 2020). However, despite the range of distribution, we were unable to identify any nesting sites during our field surveys or in literature in four mid and far-western districts (Dang, Banke, Bardia, and Kailali) and one central district (Parsa) of Nepal (Fig. 1). Overall, we collected data on 269 nesting sites or colonies from 18 districts in six provinces (not found in Karnali Province) between 2018 and 2023 (Fig. 1). However, to avoid spatial bias and autocorrelation (Boria et al., 2014), we only used one point within a 1 km × 1 km grid. This resulted in a total of 122 nest site occurrences for modeling.
2.3
Environmental parameters
2.3.1
Current parameters
Bioclimatic factors reflect annual patterns, seasonality, and temperature and precipitation extremes that influence the occurrences of species (Hijmans et al., 2005). We downloaded 19 bioclimatic data (average for the years 1970–2000) from Worldclim version 2 (Fick and Hijmans, 2017) at a resolution of 30 s (~1 km2) to model the current distributional and nesting habitat of the Lesser Adjutant in Nepal. In addition, land use (e.g., urban and wetland) are important factors affecting the distribution and nest-site selection of large-sized waterbirds including Lesser Adjutant (Sundar et al., 2019; Katuwal et al., 2022b). So, we downloaded the Moderate Resolution Imaging Spectroradiometer (MODIS) global land use and land cover (hereafter land use) map at same resolution (Li et al., 2017) and extracted the raster files for Euclidian distance from the nearest wetland (all water bodies) and urban pixels across the study area in ArcMap (10.4). Then, we calculated the raster file for the proportion of farmland and forest area using a moving window (5 × 5) in raster package (Hijmans, 2022).
We extracted the spatial values from all the layers using the occurrence data in the usdm package (Naimi et al., 2014) and performed the Pearson correlation test to avoid multicollinearity in the predictors. We excluded highly correlated variables (r > 0.7). The remaining four ecologically significant bioclimatic variables (Bio-1: annual mean temperature; Bio-4: temperature seasonality (standard deviation × 100); Bio-12: annual precipitation; and Bio-19: precipitation of coldest quarter), distance to wetland, distance to urban and farmland proportion were then used for modeling.
2.3.2
Future parameters
We used the sixth version of the Model for Interdisciplinary Research on Climatic (MIROC), also known as MIROC6, of the general circulation model to estimate how future climate changes may affect the distribution and nesting areas of the Lesser Adjutant. MIROC6 was chosen because it is the most current version of MIROC5 and offers improved global repeatability of mean climate and internal climatic variability compared to previous version (Tatebe et al., 2019). Furthermore, MIROC's circulation was used as it better captures the rainfall and temperature fluctuation across South Asia (Mishra et al., 2014). Data are available for four Shared Socioeconomic Pathways (SSPs) for 2050 (2041–2060) and 2070 (2061–2080) to account for potential impacts of climate change on the species. To encompass the full range of predicted climate change scenarios, we used SSP1–2.6, which is based on a lower emission scenario and predicts a mean warming well below 2 ℃ by the end of century, and SSP5–8.5, which is based on the highest emission scenario and predicts a mean warming of around 5.1 ℃ by the end of this century (Riahi et al., 2017; Hausfather, 2018). In addition, to evaluate how climate and land use change collectively affect the future range of the species, we used a 2050-simulated MODIS land use map (Li et al., 2017) and included two land use change scenarios: A1B, which depicts rapid economic growth with a balance on all energy sources, and A2, which emphasizes on development, leads to higher greenhouse emission, and adversely affect the environment. To determine the cumulative effects of climate and land use changes on habitat suitability for 2050 and 2070, we paired SSP1–2.6 with the A1B scenario and SSP5–8.5 with the A2 scenario following Pant et al. (2021) and Baral et al. (2023). Since there was no simulated MODIS land use change scenario for 2070, we ran the model for 2070 climatic data using the closest simulated data for 2050.
2.4
Species distribution modeling
Species distribution modeling (SDM) establishes a relationship between species and environment to explain and predict a species' likely current and future distribution (Elith and Leathwick, 2009; Thuiller et al., 2009; Naimi and Araújo, 2016). It enables the projection of species distribution in both time and space, which can assist in identifying suitable habitats for species and mitigating the risk of the local extinction of several threatened taxa within their native geographical range (Fois et al., 2016). Furthermore, SDM can aid in guiding management and restoration decisions by providing future projections of suitable habitats, minimizing potential risk (Riordan et al., 2018; Chhetri et al., 2021).
We used ensemble modeling, a machine learning technique that use multiple algorithms to forecast the species distribution, which is widely used in the field of ecology due to its ability to enhance overall accuracy beyond the capabilities of single algorithms (Araújo and New, 2007; Oppel et al., 2012; Sagi and Rokach, 2018). We selected five algorithms that are widely used in the SDM for birds, such as the generalized linear model (GLM), the generalized additive model (GAM), the multivariate adaptive regression spline (MARS), the boosted regression tree (BRT), and the random forest (RF; see Oppel et al., 2012; Tehrani et al., 2020; Awan et al., 2021; Brambilla et al., 2022). We used the sdm package (Naimi and Araújo, 2016) for ensemble modeling in the R program (R Core Team, 2021).
We used four bioclimatic (Bio-1, Bio-4, Bio-12, and Bio-19), and three land use variables (distance to wetland, distance to urban and farmland proportion) to predict current and future distribution and nesting habitats. We randomly generated 5000 pseudo-absence points below 3000 m. The presence and pseudo-absence points were divided into training sets (70%) and testing sets (30%). We ran 10 replications with a subsampling procedure using the five model algorithms, resulting in a total of 50 individual models. We constructed an ensemble model using the weighted mean approach, which improved model precision by maximizing the sum of sensitivity-specificity using True Skill Statistic (TSS) optimization (Marmion et al., 2009; Naimi and Araújo, 2016). We also determined the relative importance of each predictor variable used in the models.
We used the receiver operating characteristic (ROC) plot and calculated the area under the curve (AUC) to assess the model's performances (Bradley, 1997). AUC values ranging from 0.5 to 1.0 indicated three distinct prediction accuracies: excellent at > 0.90, good between 0.70 and 0.90, and uninformative at 0.70. Due to criticisms of the use of AUC for model evaluation (Lobo et al., 2008), we additionally examined model performance using the TSS (Allouche et al., 2006). TSS values vary from −1 to 1, with the highest value indicating good model performance with perfect discrimination and a value of zero or below suggesting random performance. To convert the predicted probabilities into presence and absence, we used the "Maximum Sensitivity plus Specificity" threshold (Liu et al., 2005; Naimi and Araújo, 2016). We then calculated the area of predicted current distribution and nesting habitat of the species, as well as the changes in these habitats between the current and future predictions scenarios relative to the area of Nepal.
2.5
Conservation gap analysis
We applied gap analysis, a widely used method for evaluating the effectiveness of PAs for species conservation (Fonseca and Venticinque, 2018; Yang et al., 2020). We projected the current and future predicted maps of the species into Nepal's PAs to assess their potential for safeguarding the species. In addition, we digitized a map of Nepal's IBAs (DNPWC, 2022) to assess their suitability as habitats for the species.
3.
Results
3.1
Distribution and breeding status
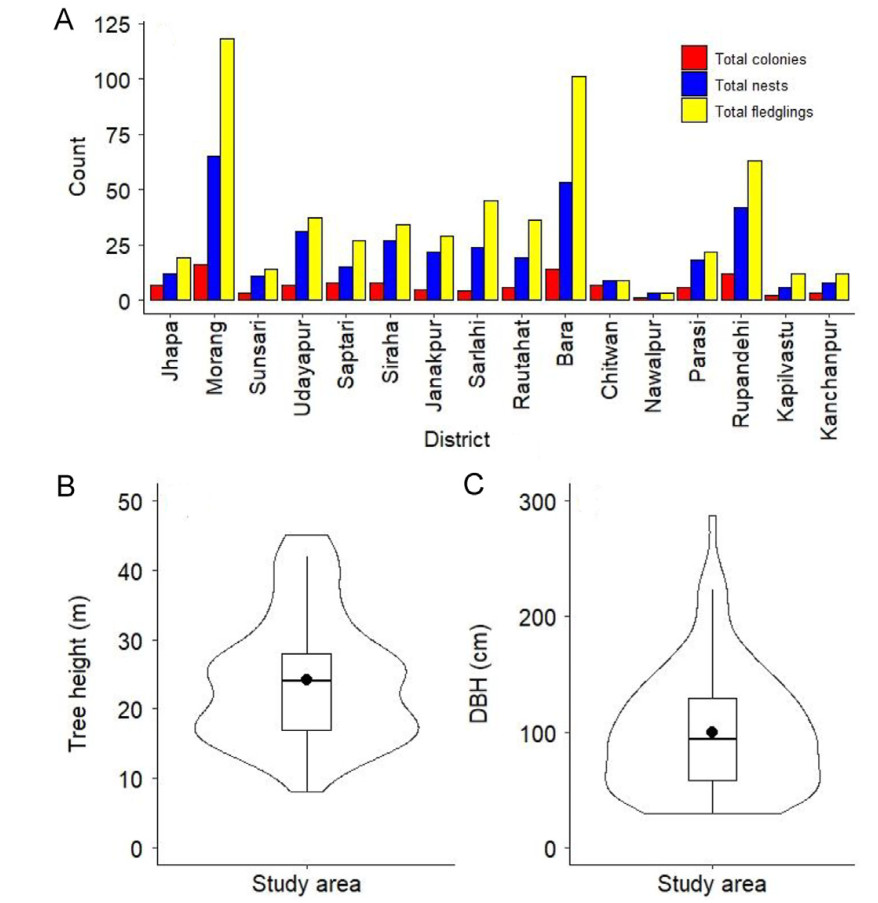
We confirmed the distribution of the Lesser Adjutant in 24 districts and documented nesting sites in 18 districts across six provinces (Fig. 1). Lesser Adjutant was observed sporadically west of Kapilvastu, but we were unable to locate nesting sites in Dang, Banke, Bardia and Kailali districts, except in Kanchanpur (a very few colonies exist in Shuklaphanta National Park) and also in Parsa district in central Nepal (Fig. 1). However, in our nation-wide breeding survey conducted in 2020, we documented 109 colonies of Lesser Adjutant across 16 districts, with total of 595 chicks (mean ± SD: 5.45 ± 5.13) in 346 nests (mean ± SD: 3.34 ± 3.0; Fig. 2A). Majority of the chicks (97.5%, n = 581, mean ± SD: 5.33 ± 5.13) successfully fledged from these nests. Higher number of colonies, nests and fledglings were recorded in Morang, followed by Bara and Rupandehi districts (Fig. 2A). Chicks' mortality was 2.5% (n = 14) primarily due to felling of nesting trees (n = 12), and unknown reasons. During the 2020 survey, we documented a total of 1273 Lesser Adjutants (adults and young).
Figure
2.
Breeding colonies and metrics across different districts (A) and the tree dimension (B, C) of the nesting colonies of the Lesser Adjutant in 2020 nation-wide survey in Nepal. B and C are the violin plots, while the black line in the box plot represents the median, and the dot represents the mean value.
In the 2020 nation-wide breeding survey, we discovered that Lesser Adjutants used 11 trees species for nesting across 16 districts. Bombax ceiba was the most frequently used species, accounting for 65% of the colonies (n = 71), followed by Haldina cordifolia with 13% (n = 14), Trewia nudiflora with 6.5% (n = 7), and Breonia chinensis with 4% (n = 4). Other species, including Shorea robusta, Mangifera indica, Tamarindus spp., Populus ciliata, Ficus religiosa, Celtis australis, and Albizia lucidior, had fewer colonies (n = 1–3 each). The mean (±SD) height of nesting trees was 24.23 ± 9.03 m and DBH was 100.09 ± 52.46 cm (Fig. 2B and C). The height and DHB of the nesting trees also varied across the study area (Fig. 2B and C).
3.2
Model performance
The ensemble model demonstrated good performance for both distribution (AUC = 0.94 ± 0.1, TSS = 0.82 ± 0.01) and nesting site (AUC = 0.95 ± 0.01, TSS = 0.80 ± 0.03). Annual mean temperature (Bio-1) was the most influential predictive variable for distribution (relative variable importance = 60%), followed by precipitation of coldest quarter (Bio-19; 22%), distance to urban area (16%), and temperature seasonality (Bio-4; 13%); the other variables had less than 10% contribution on the distribution of the species (Appendix Fig. S1A). Similarly, for nest site, annual mean temperature (Bio-1) also had the highest influence (38%), followed by precipitation of coldest quarter (Bio-19; 30%), farmland (24%), distance to urban (13%), and temperature seasonality (Bio-4; 12%), whereas, the others had less than 10% contribution (Appendix Fig. S1B). Response curves of the models indicated that the probablity of the distribution and nesting habitat suitability increased with increasing annual mean temperature (Bio-1) and farmland proportion, whereas, decreased with increasing precipitation of coldest quarter (Bio-19) and distance to urban area (Appendix Fig. S2).
3.3
Predicted habitat suitability for Lesser Adjutant in Nepal
3.3.1
Current prediction
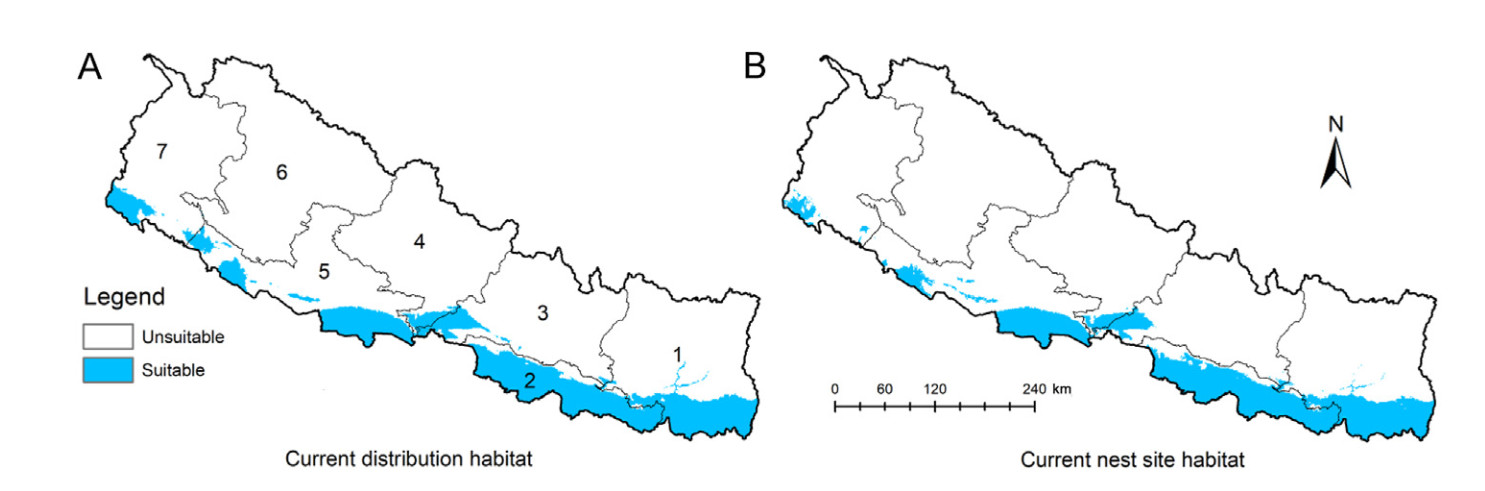
Approximately 15% (21,637 km2) and 13% (19,651 km2) of Nepal's total area are potentially suitable for the current distribution and nesting sites of the species, respectively (Table 1; Fig. 3). The highest suitable habitat for both distribution and nesting sites was predicted to be in Madhesh Province, while none was predicted in Karnali Province (Fig. 3). The suitable distribution and nesting habitat were fewer and patchily predicted in the west of Kapilvastu district of Lumbini Province (Fig. 3). Our analysis revealed that most of the predicted current habitat for distribution (71%) and nesting site (74%) was located in farmlands, followed by forests (distribution = 19%, nesting site = 17%), and a few other land use categories.
Table
1.
Predicted suitable area (km2) for distribution and nesting site for Lesser Adjutant in Nepal under current and future climatic and land use change scenarios.
Current
2050
2070
Parameters
SSP1–2.6
SSP5–8.5
SSP1–2.6
SSP5–8.5
Distribution
21,637 (14.64%)
32,165 (21.76%)
37,076 (25.08%)
33,183 (22.45%)
43,210 (29.24%)
Nesting site
19,651 (13.29%)
29,074 (19.67%)
32,685 (22.11%)
29,763 (20.14%)
37,133 (25.12%)
The percentage in the parentheses represents the predicted area relative to the area of Nepal.
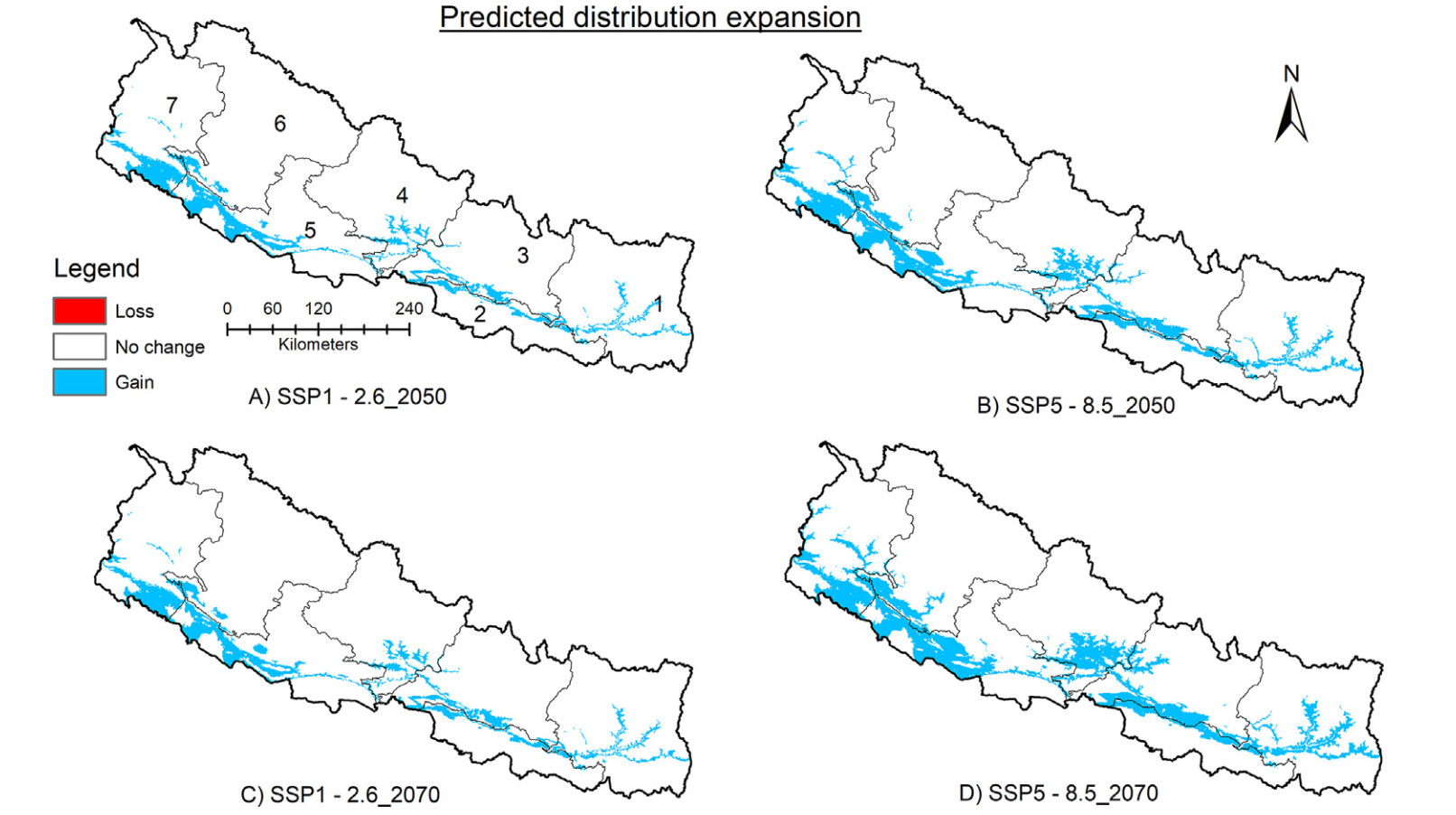
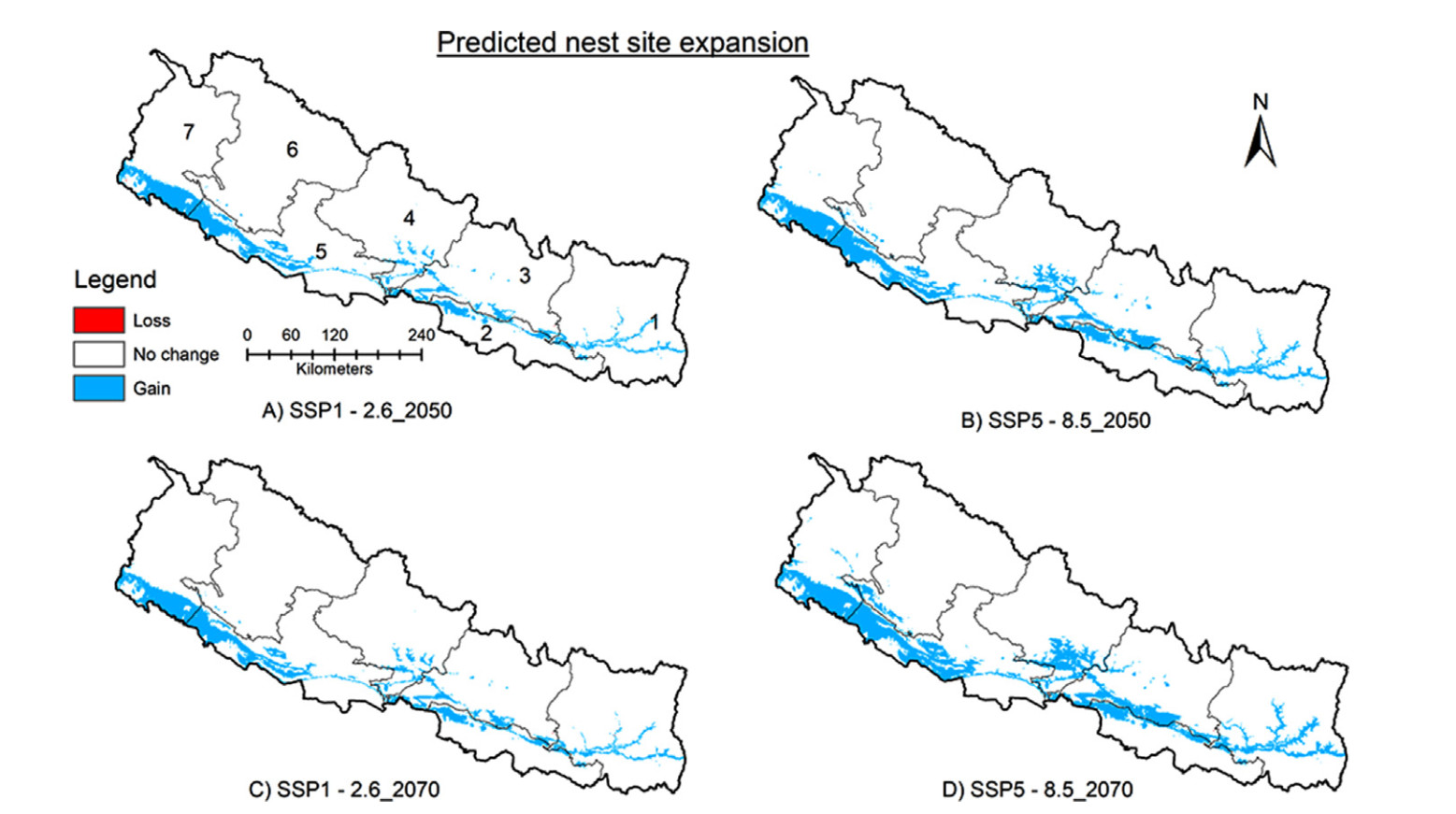
According to the model, the species' distribution range could expand by a maximum of around 10% (15,439 km2) and 15% (21,573 km2) under SSP5–8.5 scenarios in 2050 and 2070, respectively (Table 1; Fig. 4). On the other hand, range contraction was predicted to be very minimal (< 0.002) for both scenarios in both years (Table 1; Fig. 4). The model predicted that the range expansion would mostly occur within the existing distribution range, particularly in Lumbini and Sudurpashchim Provinces (Tarai and some regions of Siwalk), and extend to the northern portions (Siwalik and Mid-hill regions) of other provinces (Fig. 4). The predicted range expansion was greater for SSP5–8.5 scenarios than for SSP1–2.6 scenarios in both years (Table 1).
Figure
4.
Predicted range expansion for distribution habitat of Lesser Adjutant in 2050 and 2070. Refer to Fig. 1 for names of provinces.
The model predicted a maximum range expansion of around 9% (13,034 km2) and 12% (17,482 km2) for nesting sites under SSP5–8.5 scenarios for 2050 and 2070, respectively (Table 1; Fig. 5). Range contraction was almost negligible for all scenarios (< 0.02%; Fig. 5). The nesting range expansion was predicted to be mostly confined to the existing distribution range in Lumbini and Sudurpashchim Provinces (Tarai and some parts of Siwalik area) but was also predicted to extend to the northern portions of other provinces, particularly in Siwalik and Mid-hill regions (see Fig. 5). In both years, the predicted range expansion was slightly greater for SSP5–8.5 scenarios than for SSP1–2.6 scenarios (Table 1; Fig. 5).
Figure
5.
Predicted range expansion for nesting site of Lesser Adjutant in 2050 and 2070. Refer to Fig. 1 for names of provinces.
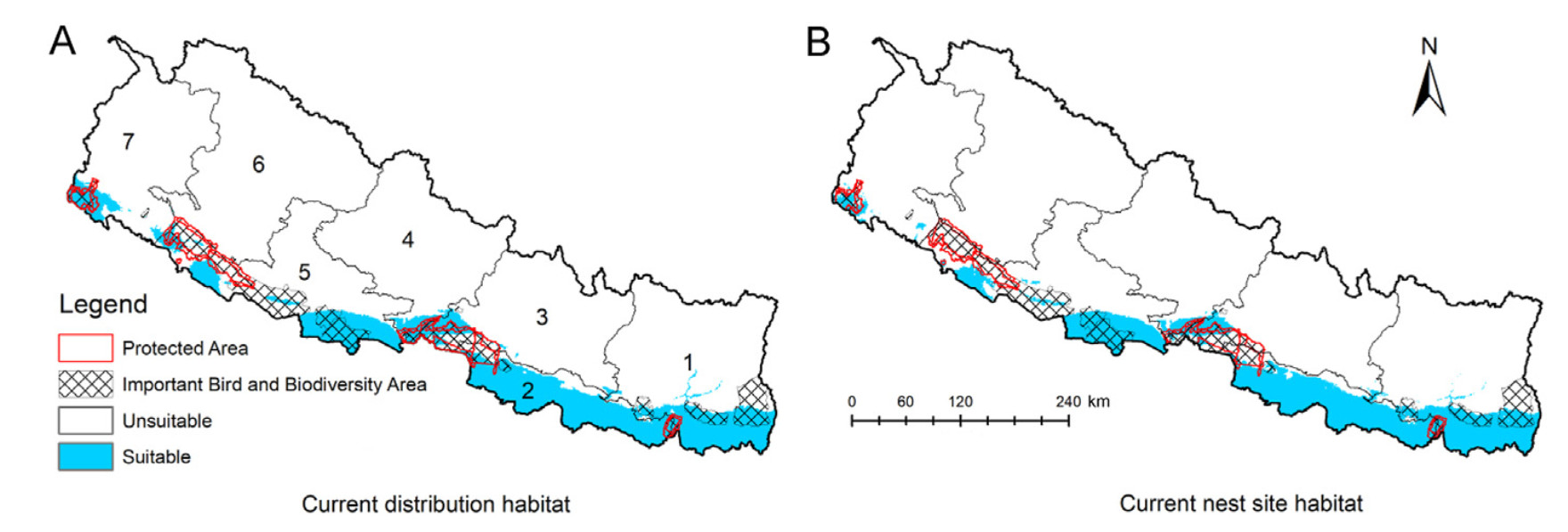
The majority of potential distribution habitats and nesting locations for the Lesser Adjutant were found outside of Nepal's protected areas (Fig. 6). Only 12% of the current suitable distribution habitat and 9% of the current suitable nesting habitat were located within protected areas (Fig. 6). However, predictions indicate that a maximum of 15% of future suitable distribution habitat and 14% of suitable nesting habitat (under SSP5–8.5 scenario for 2050) are likely to be within protected areas, but this is predicted to a slight decline in the 2070s. Additionally, 28% of the current suitable distribution habitat and 25% of suitable nesting sites were located within IBAs (Fig. 6). Predictions suggest that a maximum of 29% of suitable distribution habitat and 29% of suitable nesting habitat under SSP5–8.5 for 2050s are likely to fall inside IBAs in the future, but this is predicted to a slight decline in the 2070s.
Figure
6.
Current suitable distribution and nesting site of the Lesser Adjutant within Protected Areas and Important Bird and Biodiversity Areas of Nepal. Refer to Fig. 1 for names of provinces.
This study represents the first comprehensive evaluation of the distribution and nesting ecology of the Lesser Adjutant in Nepal. Our investigation of the breeding colonies and distribution of the Lesser Adjutants in Nepal demonstrates that the species is more widely distributed than initially believed. Although previous estimates suggested a population size of around 1000 individuals (Inskipp et al., 2016), our findings revealed the highest recorded number of fledglings and adults in Nepal. We discovered larger breeding populations in Morang, Bara and Rupandehi districts; however, due to COVID-19, we were unable to explore some sections of the Kapilvastu and also Rupandehi districts in 2020, where greater colonies (n = 29) and fledglings (n = 114) were recorded in 2019 (Katuwal et al., 2022b). The fledgling success rate across Nepal was greater than that reported in previous studies (Karki and Thapa, 2013; Sundar et al., 2016; Bhattarai et al., 2021; Katuwal et al., 2022b). Since the Lesser Adjutant has a higher fledgling success rate than many other waterbirds (Sundar, 2011; Ramamohan and Rao, 2017; Pang et al., 2019; Gulati and Rana, 2021; Kar et al., 2021), and if we had surveyed all areas, the population in Nepal would be even larger than what has been guesstimated.
The habitat suitability map for species distribution was wider than the nesting site, because Lesser Adjutants forage over a vast area (Sundar et al., 2016), but their nesting sites are localized in patches (Inskipp et al., 2016; Katuwal et al., 2022b). Our habitat suitability maps indicated that the Lesser Adjutant prefers agricultural regions over forest habitats for both distribution and breeding. Although Lesser Adjutants employed less forest habitat for distribution and breeding than agricultural areas (Koju et al., 2019; Katuwal et al., 2022b), our model predicted that some forest area, primarily in the Tarai-Siwalik regions, would be exploited as nesting and foraging habitats. However, it should be noted that the species' habitat utilization is highly dependent on the availability of open spaces or wetlands in these forested regions (HBK personal observation). Although the model indicated that some parts of Lumbini and Madhesh Province and Koshi Province would be particularly suitable for nest site, the use of such habitat is largely reliant on the availability of the most popular nesting trees, such as Bombax ceiba, Haldina cordifolia, and Trewia nudiflora, among others, because such trees have a beneficial effect on colony size or nesting rate, and thus on breeding success (Hilaluddin Shah and Shawl, 2003; Koju et al., 2019). However, there are fewer of these trees in the landscape and tree felling is commonly observed, limiting nest site choices in the region (Katuwal et al., 2022b). Protection of these trees across Nepal will be beneficial for the Lesser Adjutant, but removing them poses significant challenges to the species.
The model indicated that only a few areas in mid-western and far-western Nepal (west of Kapilvastu in Lumbini Province, such as Dang, Banke, and Bardia, and Sudurpashchim Province, such as Kailali and Kanchanpur) were suitable habitats for the Lesser Adjutant. While breeding adults have been sighted in multiple areas in this low suitable habitat, breeding has only been confirmed in Kanchanpur (Sukalaphata National Park) in far-western Nepal. In 2019 and 2020, we conducted several trips to search for Lesser Adjutant nesting sites in these places, but we were unsuccessful in finding any. Although ebird indicated the presence of a Lesser Adjutant in Banke in the 1980s, we could not locate any during our visits or expert contacts. Therefore, compared to other provinces, there were fewer nesting sites in Sudurpashchim and Lumbini Province (west of Kapilvastu; Sharma, 2006; Inskipp et al., 2016). It is also possible that the species could breed on the Indian side, which has more agricultural areas than the Nepali side. Additionally, it should be noted that the species has rarely been recorded west of the Nepal border, which marks its northen-westernmost distribution range (https://ebird.org/species/lesadj1; Elliott et al., 2020; BirdLife International, 2021).
The Lesser Adjutant's known distribution in Nepal is primarily below 400 m (Inskipp et al., 2016; Baral et al., 2020). However, our model predicted a few suitable habitats at somewhat higher elevations (< 1000 m), possibly due to the availability of similar climatic conditions. Nevertheless, majority of their predicted habitat lies in lowland due to larger agricultural areas and wetland habitats suitable for foraging and breeding, making them a tropical species. Our model also found that annual mean temperature (Bio-1) and precipitation during the coldest quarter (Bio-19) contributed to the distribution and nesting habitat suitability. However, Khadka and Pandey (2014) identified annual precipitation and minimum temperature of the coldest month as the most influential variables on the Lesser Adjutant's distribution. As Lesser Adjutant fledglings are observed throughout the winter months, increased precipitation and temperature anomalies during this season may impact their breeding success rate. We observed fatalities of Lesser Adjutant chicks due to excessively low temperatures and unseasonal precipitation in our study area (Katuwal et al., 2022b). Changing monsoonal patterns have also resulted in unseasonal nesting of the Sarus Crane in India (Sundar et al., 2018). However, a comprehensive study is necessary to understand the impact of precipitation and temperature anomalies on every aspect of the Lesser Adjutant's breeding cycle, including the timing of egg laying, hatching propensity, and fledgling success. Our model also detected a positive influence of farmland on nesting habitat and on distribution, suggesting that Lesser Adjutants prefer to nest and forage in farmlands over other habitats. In addition, the suitability of distribution and nesting habitats were higher nearer to urban areas, indicating that the species is found in human-dominated landscapes; however, further research is required to understand how urban areas influence their breeding ecology.
4.2
Future habitat suitability
In the previous study, Khadka and Pandey (2014) predicted that the current distribution habitat of the Lesser Adjutant has shifted towards the northern region compared to its historical distribution, owing to climate change. Our models also showed that the current distribution and nesting habitats of the species are further expected to expand to higher elevations in the mid and far-western regions (Tarai and a few Siwalik regions) as well as towards the northern regions (Siwalik and Mid-Hill regions) by 2050 and 2070s. These findings demonstrate that the Lesser Adjutants are susceptible to the impacts of climate change. Although climate change is a threat to many species (Jetz et al., 2007; Langham et al., 2015; Tehrani et al., 2020), a small number of them also benefit. Many bird species, such as pheasants in the Himalayas (Chhetri et al., 2021) and the Black-necked Crane (Grus nigricollis) in China, are expected to experience range expansion (Li et al., 2022). Conversely, some species, such as the Oriental Honey-buzzard (Pernis ptilorhynchus), are predicted to experience range contraction due to climate change (Condro et al., 2022). While our research suggests a range extension based on future climate and land use scenarios, the colonization of the new areas by the Lesser Adjutant is contingent on the availability of large nesting trees in agricultural areas, and large wetlands, and agricultural areas for foraging. Because the trees commonly utilized by the species are scarce in the landscapes (Katuwal et al., 2022b). Hence, conserving large trees and their habitats within the species' range will be essential for their survival and range expansion, especially if they migrate to higher elevations beyond their current range.
4.3
Protected areas and IBAs versus predicted distribution
Protected areas have been established globally to safeguard biodiversity under threat (Coetzee et al., 2014; Ghosh-Harihar et al., 2019). However, existing protected areas often fail to provide adequate habitats for a significant number of threatened species (Coad et al., 2019; Kearney et al., 2020), as a large proportion of them are found outside these areas. This is particularly true for the Lesser Adjutant, whose habitat extends mostly outside protected areas, particularly in agricultural regions (Sundar et al., 2016; Koju et al., 2019; Katuwal et al., 2022b). Moreover, although IBAs have been established worldwide to promote bird conservation, they are not representative geographically in Nepal as there are gaps in the lowlands of Madhesh and Koshi Provinces (DNPWC, 2022). Most suitable habitats for the Lesser Adjutant are situated outside IBAs, and even where they are located inside IBAs, they are not adequately monitored, posing a threat to this important bird habitat. For instance, the Urlabari Forest Grooves IBA in Morang district of Koshi Province was established especially for the conservation of the Lesser Adjutant (Baral and Inskipp, 2005), however it has been removed from the revised list of IBAs (DNPWC, 2022), because the species was believed to have disappeared from the area, potentially as a result of sporadic monitoring. But, we discovered some thriving breeding colonies in the area and other parts of the district. So, to safeguard the critical habitat of this globally vulnerable species, continuous monitoring and conservation efforts should focus on IBAs outside of protected areas, and the establishing additional IBAs in the lowlands of Madhesh and Koshi Provinces could be considered to address gaps in the IBA network.
4.4
Conservation implications
Our field observations and models indicate that the suitable habitat of the Lesser Adjutant is primarily found in agricultural landscapes outside of protected areas between Lumbini and Koshi Provinces, specifically from the Kapilvastu to Jhapa districts. Future projections suggest an expansion of suitable habitats. Therefore, we recommend that the local, provincial, and central governments of Nepal prioritize agricultural lands as suitable habitats for the species and promote agricultural policies that benefit threatened species such as the Lesser Adjutant. Further research is needed in the Siwalik areas and river valleys of Lumbini (west of Kapilvastu) and Sudhurpashchim Provinces to identify breeding sites of the species. It is essential to establish IBAs, particularly in Madhesh and Koshi Provinces, to protect the species' natural habitats. Cutting trees is frequently observed in the study area, so the breeding success of the species within these IBAs, must be monitored regularly. The Department of Forests and Soil Conservation and Division Forest Offices (through provincial ministries) should continue to safeguard large-sized nesting trees, particularly by delisting nesting-preferred tree species from harvesting. Controlling agrochemicals concurrently with the protection of large nesting trees is essential for improving the breeding performance of the species in agricultural areas (Mitra et al., 2011; Koju et al., 2019). Monitoring known nesting habitats and newly discovered colonies are essential for their conservation. We advocate for land zoning policies that limit the unplanned urbanization of agricultural areas. Finally, the ecological significance of the species in agricultural landscapes must be conveyed to local farmers and students.
5.
Conclusions
We discovered a greater distribution and nesting area for the Lesser Adjutant than was previously reported in Nepal. Our research has shown that the species is likely to have higher suitable habitat in the agricultural landscapes of the western to eastern regions of Nepal (Kapilvastu to Jhapa districts), but scarce in the mid and far-western region (west of Kapilvastu). While many studies focus on the negative effects of climate and land use change on species, our research suggests that the distribution and nesting habitats of the Lesser Adjutants are likely to expand in the future. Nonetheless, the utilization of any habitat for nesting relies heavily on the presence of large-size trees within or nearby agricultural landscapes. Therefore, we recommend continued monitoring of the distribution and nesting habitat of the species in Nepal.
Authors' contributions
HBK, HPS, and RCQ conceptualized the study; HBK, HPS, BPB, SK, and RCQ acquired funding, HBK, PR, KPB, BPB, SCL, SY, GS, HSB, and LPP collected data; HBK, HPS, LW, and RCQ developed methods and performed data analysis; HBK wrote the first draft of the paper. All authors read and approved the final manuscript.
Ethics statement
We did not handle the species, so do not require any ethical clearance.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
This work has been supported by CAS-SEABRI (Y4ZK111B01), International Science, and Technology Commissioner of Yunnan Province (202203AK140027), Yunnan Province Science and Technology Department (202203AP140007), Rufford Small Grants Foundation (31372–2), and Tribhuvan University National Priority Area Research Grant (TU-NPAR-2078/79-ERG-04). We thank the Department of Forests and Soil Conservation and the Department of National Parks and Wildlife Conservation, Nepal for providing permissions to carry out this work. We would like to thank Sandeep Regmi, Amrit Nepali, Tek Gharti Magar, Nikeet Pradhan, Ramji Gautam, Sabin K.C., Yubin Raj Shrestha, Tek Bahadur Gurung, Jaya Bhandari, Manav Chaudhary, Dibya Raj Dahal, Amit Chaudhary, Hari Sharan Giri, Yam Mahato, Netra Koirala, Anis Timsina, Prashant Ghimire, K.S. Gopi Sundar, Asmit Limbu, Dev Narayan Mandal, Ram Shahi, Manju Mahatara, Deep Shahi, Pawan Gautam, Hirulal Dungura, Deu Bahadur Rana, Ishwari Prasad Chaudhary, Bird Conservation Nepal, Nitesh Das, Tapil Prakash Rai, Devraj Joshi, Laxmi Raj Joshi, National Trust for Nature Conservation, Ganesh Tamang, and Wyclife Agumba Oluoch for their help, cooperation, and sharing information during the study period. We would also like to thank Suraj Baral for suggestions on modeling and Jon E. Swenson for revising the English of the manuscript. Also, we would like to thank two reviewers for their constructive comments on the manuscript.
Adhikari, P., Kim, B.J., Hong, S.H., Lee, D.H., 2022. Climate change induced habitat expansion of nutria (Myocastor coypus) in South Korea. Sci. Rep. 12, 3300.
Allouche, O., Tsoar, A., Kadmon, R., 2006. Assessing the accuracy of species distribution models: prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 43, 1223–1232.
Araújo, M.B., New, M., 2007. Ensemble forecasting of species distributions. Trends Ecol. Evol. 22, 42–47.
Awan, M.N., Saqib, Z., Buner, F., Lee, D.C., Pervez, A., 2021. Using ensemble modeling to predict breeding habitat of the red-listed Western Tragopan (Tragopan melanocephalus) in the Western Himalayas of Pakistan. Glob. Ecol. Conserv. 31, e01864.
Baral, B., Bhandari, S., Koirala, S., Bhandari, P., Magar, G., Basnet, D.R., et al., 2020. First distributional record of the lesser adjutant Leptoptilos javanicus Horsfield, 1821 (Ciconiiformes: Ciconiidae) from Sindhuli district, Nepal. J. Threat. Taxa 12, 17028–17031.
Baral, H.S., Inskipp, C., 2005. Important bird areas in Nepal: key sites for conservation. In: Bird Conservation Nepal and BirdLife International (Kathmandu and Cambridge).
Baral, S., Kunwar, A., Adhikari, D., Kandel, K., Mandal, D.N., Thapa, A., et al., 2023. The potential distribution of the yellow monitor, Varanus flavescens (Hardwick & Gray) under multiple climate, land cover and dispersal scenarios in Nepal. Wildl. Res. .
Bellard, C., Bertelsmeier, C., Leadley, P., Thuiller, W., Courchamp, F., 2012. Impacts of climate change on the future of biodiversity. Ecol. Lett. 15, 365–377.
Bhattarai, B.P., Adhikari, J.N., Rijal, M., 2021. Nesting habitat selection and challenges of conservation of the vulnerable Lesser Adjutant Leptoptilos javanicus (Horsfield, 1821) in the Chitwan National Park, Nepal. Ornis Hung. 29, 33–46.
BirdLife International, 2021. Species factsheet: Leptoptilos javanicus. . (Accessed 3 August 2021).
Boria, R.A., Olson, L.E., Goodman, S.M., Anderson, R.P., 2014. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 275, 73–77.
Bradley, A.P., 1997. The use of the area under the ROC curve in the evaluation of machine learning algorithms. Pattern Recogn. 30, 1145–1159.
Brambilla, M., Rubolini, D., Appukuttan, O., Calvi, G., Karger, D.N., Kmecl, P., et al., 2022. Identifying climate refugia for high-elevation alpine birds under current climate warming predictions. Global Change Biol. 28, 4276–4291.
Chapagain, D., Dhaubanjar, S., Bharati, L., 2021. Unpacking future climate extremes and their sectoral implications in western Nepal. Clim. Change 168, 8.
Chhetri, B., Badola, H.K., Barat, S., 2021. Modelling climate change impacts on distribution of Himalayan pheasants. Ecol. Indicat. 123, 107368.
Chhogyel, N., Kumar, L., 2018. Climate change and potential impacts on agriculture in Bhutan: a discussion of pertinent issues. Agric. Food Secur. 7, 79.
Coad, L., Watson, J.E.M., Geldmann, J., Burgess, N.D., Leverington, F., Hockings, M., et al., 2019. Widespread shortfalls in protected area resourcing undermine efforts to conserve biodiversity. Front. Ecol. Environ. 17, 259–264.
Coetzee, B.W.T., Gaston, K.J., Chown, S.L., 2014. Local scale comparisons of biodiversity as a test for global protected area ecological performance: a meta-analysis. PLoS One 9, e105824.
Condro, A.A., Syartinilia Higuchi, H., Mulyani, Y.A., Raffiudin, R., Rusniarsyah, L., Setiawan, Y., et al., 2022. Climate change leads to range contraction for Japanese population of the Oriental Honey-Buzzards: implications for future conservation strategies. Glob. Ecol. Conserv. 34, e02044.
D'Amour, C.B., Reitsma, F., Baiocchi, G., Barthel, S., Güneralp, B., Erb, K.H., et al., 2017. Future urban land expansion and implications for global croplands. Proc. Nat. Acad. Sci. USA 114, 8939–8944.
DNPWC, BCN, 2022. Birds of Nepal: an Official Checklist. Department of National Parks and Wildlife Conservation and Bird Conservation Nepal, Kathmandu, Nepal.
Elith, J., Leathwick, J.R., 2009. Species distribution models: ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 40, 677–697.
Elliott, A., Garcia, E.F.J., Boesman, P.F.D., 2020. Lesser adjutant (Leptoptilos javanicus), version 1.0. In: del Hoyo, J., Elliott, A., Saragata, J., Christie, D.A., de Juana, E. (Eds.), Birds of the World. Cornell Lab of Ornithology, Ithaca, NY.
Fick, S.E., Hijmans, R.J., 2017. WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315.
Fois, M., Cuena-Lombraña, A., Fenu, G., Cogoni, D., Bacchetta, G., 2016. The reliability of conservation status assessments at regional level: past, present and future perspectives on Gentiana lutea L. ssp. lutea in Sardinia. J. Nat. Conserv. 33, 1–9.
Fonseca, C.R., Venticinque, E.M., 2018. Biodiversity conservation gaps in Brazil: a role for systematic conservation planning. Perspect. Ecol. Conserv. 16, 61–67.
Fusco, J., Walker, E., Papaïx, J., Debolini, M., Bondeau, A., Barnagaud, J.Y., 2021. Land use changes threaten bird taxonomic and functional diversity across the Mediterranean Basin: a spatial analysis to prioritize monitoring for conservation. Front. Ecol. Evol. 9, 612356.
Garcia, R.A., Cabeza, M., Rahbek, C., Araújo, M.B., 2014. Multiple dimensions of climate change and their implications for biodiversity. Science 344, 1247579.
Ghimire, P., 2019. A review of studies on climate change in Nepal. Geogr. Base 6, 11–20.
Ghosh-Harihar, M., An, R., Athreya, R., Borthakur, U., Chanchani, P., Chetry, D., et al., 2019. Protected areas and biodiversity conservation in India. Biol. Conserv. 237, 114–124.
Gula, J., Sundar, K.S.G., Willows-Munro, S., Downs, C.T., 2023. The state of stork research globally: a systematic review. Biol. Conserv. 280, 109969.
Gulati, H., Rana, S., 2021. Nest characteristics and breeding success of Sarus Crane, Antigone antigone (Linnaeus, 1758) (Aves: Gruidae) in different habitats at Dhanauri Wetland, Uttar Pradesh, India. Record Zool. Surv. India 121, 205–210.
Hamal, K., Sharma, S., Baniya, B., Khadka, N., Zhou, X., 2020. Inter-annual variability of winter precipitation over Nepal coupled with ocean-atmospheric patterns during 1987-2015. Front. Earth Sci. 8, 161.
Hijmans, R.J., 2022. raster: geographic data analysis and modeling. R package version 3.5-29. .
Hijmans, R.J., Cameron, S.E., Parra, J.L., Jones, P.G., Jarvis, A., 2005. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 25, 1965–1978.
Hilaluddin Shah, J.N., Shawl, T.A., 2003. Nest site selection and breeding success by Cattle Egret and Little Egret in Amroha, Uttar Pradesh, India. Waterbirds 26, 444–448.
Inskipp, C., Baral, H.S., Phuyal, S., Bhatt, T., Khatiwada, M., Inskipp, T., et al., 2016. The Status of Nepal's Birds: the National Red List Series. Zoological Society of London, United Kingdom.
Jetz, W., Wilcove, D.S., Dobson, A.P., 2007. Projected impacts of climate and land-use change on the global diversity of birds. PLoS Biol. 5, 1211–1219.
Kar, T., Debata, S., Mundkur, T., 2021. Population, breeding phenology, and factors affecting breeding success of River Tern (Sterna aurantia) in Eastern India. Waterbirds 44, 308–316.
Karki, R., Gurung, A., 2012. An overview of climate change and its impact on agriculture: a review from least developing country, Nepal. Int. J. Ecosyst. 2, 19–24.
Karki, R., ul Hasson, S., Schickhoff, U., Scholten, T., Böhner, J., 2017. Rising precipitation extremes across Nepal. Climate 5, 4.
Karki, S., Thapa, T.B., 2013. Population status, nesting habitat selection and conservation threats of lesser adjutant stork (Leptoptilos javanicus) in the eastern lowlands of Nepal. Conserv. Sci. 1, 27–35.
Kattel, G.R., 2022. Climate warming in the Himalayas threatens biodiversity, ecosystem functioning and ecosystem services in the 21st century: is there a better solution? Biodivers. Conserv. 31, 2017–2044.
Katuwal, H.B., Rai, J., Tomlinson, K., Rimal, B., Sharma, H.P., Baral, H.S., et al., 2022a. Seasonal variation and crop diversity shape the composition of bird communities in agricultural landscapes in Nepal. Agric. Ecosyst. Environ. 333, 107973.
Katuwal, H.B., Sundar, K.S.G., Zhang, M., Rimal, B., Baral, H.S., Sharma, H.P., et al., 2022b. Factors affecting the breeding ecology of the globally threatened Lesser Adjutant Leptoptilos javanicus in agricultural landscapes of Nepal. Avian Conserv. Ecol. 17, 15.
Kearney, S.G., Adams, V.M., Fuller, R.A., Possingham, H.P., Watson, J.E.M., 2020. Estimating the benefit of well-managed protected areas for threatened species conservation. Oryx 54, 276–284.
Khadka, K.K., Pandey, R., 2014. Changes in the distribution of Lesser Adjutant Storks (Leptoptilos javanicus) in South and Southeast Asia: a plausible evidnce of global climate change and land-use change effect. Int. J. Zool. Res. 10, 9–14.
Koju, R., Maharjan, B., Gosai, K.R., Kittur, S., Sundar, K.S.G., 2019. Ciconiiformes nesting on trees in cereal-dominated farmlands: importance of scattered trees for heronries in lowland Nepal. Waterbirds 42, 355–365.
Lamsal, P., Kumar, L., Atreya, K., Pant, K.P., 2017. Vulnerability and impacts of climate change on forest and freshwater wetland ecosystems in Nepal: a review. Ambio 46, 915–930.
Langham, G.M., Schuetz, J.G., Distler, T., Soykan, C.U., Wilsey, C., 2015. Conservation status of North American birds in the face of future climate change. PLoS One 10, e0135350.
Li, M., Zhou, H., Bai, J., Zhang, T., Liu, Y., Ran, J., 2022. Distribution of breeding population and predicting future habitat under climate change of Black-Necked Crane (Grus nigricollis Przevalski, 1876) in Shaluli Mountains. Animals 12, 2594.
Li, X., Chen, G., Liu, X., Liang, X., Wang, S., Chen, Y., et al., 2017. A new global land-use and land-cover change product at a 1-km resolution for 2010 to 2100 based on human-environment interactions. Ann. Assoc. Am. Geogr. 107, 1040–1059.
Liu, C., Berry, P.M., Dawson, T.P., Pearson, R.G., 2005. Selecting thresholds of occurrence in the prediction of species distributions. Ecography 28, 385–393.
Liu, C.Y., Gélin, U., He, R.C., Li, H., Quan, R.C., 2021. Flexible breeding performance under unstable climatic conditions in a tropical passerine in Southwest China. Zool. Res. 42, 221–226.
Lobo, J.M., Jiménez-valverde, A., Real, R., 2008. AUC: a misleading measure of the performance of predictive distribution models. Global Ecol. Biogeogr. 17, 145–151.
LRMP, 1986. Land Resources Mapping Project. Kenting Earth Sciences, HMGN and Government of Canada, Land Resources Mapping Project, Kathmandu.
Marmion, M., Parviainen, M., Luoto, M., Heikkinen, R.K., Thuiller, W., 2009. Evaluation of consensus methods in predictive species distribution modelling. Divers. Distrib. 15, 59–69.
Mishra, V., Kumar, D., Ganguly, A.R., Sanjay, J., Mujumdar, M., Krishnan, R., et al., 2014. Reliability of regional and global climate models to simulate precipitation extremes over India. J. Geophys. Res. Atmos. 119, 9301–9323.
Mitra, A., Chatterjee, C., Mandal, F.B., 2011. Synthetic chemical pesticides and their effects on birds. Res. J. Environ. Toxicol. 5, 81–96.
Moradi, S., Sheykhi Ilanloo, S., Kafash, A., Yousefi, M., 2019. Identifying high-priority conservation areas for avian biodiversity using species distribution modeling. Ecol. Indicat. 97, 159–164.
Naimi, B., Araújo, M.B., 2016. Sdm: a reproducible and extensible R platform for species distribution modelling. Ecography 39, 368–375.
Naimi, B., Hamm, N.A.S., Groen, T.A., Skidmore, A.K., Toxopeus, A.G., 2014. Where is positional uncertainty a problem for species distribution modelling? Ecography 37, 191–203.
Oppel, S., Meirinho, A., Ramírez, I., Gardner, B., O'Connell, A.F., Miller, P.I., et al., 2012. Comparison of five modelling techniques to predict the spatial distribution and abundance of seabirds. Biol. Conserv. 156, 94–104.
Pang, R.H., Yu, T.L., Busam, M., 2019. Low breeding success of the little egret (Egretta garzetta) near residential areas and in colonies exposed to gales: a comparison of colony in Sichuan, Southwest China, with literature. Anim. Cell. Syst. 23, 235–240.
Pant, G., Maraseni, T., Apan, A., Allen, B.L., 2021. Predicted declines in suitable habitat for greater one-horned rhinoceros (Rhinoceros unicornis) under future climate and land use change scenarios. Ecol. Evol. 11, 18288–18304.
R Core Team, 2021. R: a Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. .
Ramamohan, H., Rao, K.K., 2017. Breeding Success and Mortality Rates in the Spot-Billed Pelican (Pelecanus philippensis), at Telineelapuram Bird Protected Areas, (An IBA Site; in 229) Srikakulam District, Andra Pradesh, India, vol. 143. Indian For, pp. 1064–1070.
Riahi, K., van Vuuren, D.P., Kriegler, E., Edmonds, J., O'Neill, B.C., Fujimori, S., et al., 2017. The shared socioeconomic pathways and their energy, land use, and greenhouse gas emissions implications: an overview. Global Environ. Change 42, 153–168.
Rimal, B., Sloan, S., Keshtkar, H., Sharma, R., Rijal, S., Shrestha, U.B., 2020. Patterns of historical and future urban expansion in Nepal. Rem. Sens. 12, 628.
Riordan, E.C., Montalvo, A.M., Beyers, J.L., 2018. Using species distribution models with climate change scenarios to aid ecological restoration decisionmaking for southern California shrublands. In: Research Paper—Pacific Southwest Research Station. USDA Forest Service. PSW-RP-270.
Sagi, O., Rokach, L., 2018. Ensemble learning: a survey. Wiley Interdiscip. Rev. Data Min. Knowl. Discov. 8, e1249.
Santangeli, A., Lehikoinen, A., Bock, A., Peltonen-Sainio, P., Jauhiainen, L., Girardello, M., et al., 2018. Stronger response of farmland birds than farmers to climate change leads to the emergence of an ecological trap. Biol. Conserv. 217, 166–172.
Sharma, H.P., Rimal, B., Zhang, M., Sharma, S., Poudyal, L.P., Maharjan, S., et al., 2020. Potential distribution of the critically endangered Chinese pangolin (Manis pentadactyla) in different land covers of Nepal: implications for conservation. Sustainability 12, 1282.
Sharma, S., 2006. Population status and distribution of Lesser Adjutant (Leptoptilos javanicus) in far-western lowland Nepal. Tigerpaper 33, 9–11.
Sundar, K.S.G., 2011. Agricultural intensification, rainfall patterns, and large waterbird breeding success in the extensively cultivated landscape of Uttar Pradesh, India. Biol. Conserv. 144, 3055–3063.
Sundar, K.S.G., Koju, R., Maharjan, B., Marcot Bruce, G., Kittur, S., Gosai, K.R., 2019. First assessment of factors affecting the breeding success of two stork species in lowland Nepal using Bayesian network models. Wildfowl 69, 45–69.
Sundar, K.S.G., Maharjan, B., Koju, R., Kittur, S., Gosai, K.R., 2016. Factors affecting provisioning times of two stork species in lowland Nepal. Waterbirds 39, 365–374.
Sundar, K.S.G., Yaseen, M., Kathju, K., 2018. The role of artificial habitats and rainfall patterns in the unseasonal nesting of Sarus Cranes (Antigone antigone) in South Asia. Waterbirds 41, 80–86.
Tatebe, H., Ogura, T., Nitta, T., Komuro, Y., Ogochi, K., Takemura, T., et al., 2019. Description and basic evaluation of simulated mean state, internal variability, and climate sensitivity in MIROC6. Geosci. Model Dev. (GMD) 12, 2727–2765.
Tehrani, N., Naimi, B., Jaboyedoff, M., 2020. Toward community predictions: multi-scale modelling of mountain breeding birds' habitat suitability, landscape preferences, and environmental drivers. Ecol. Evol. 10, 5544–5557.
Thuiller, W., Lafourcade, B., Engler, R., Araújo, M.B., 2009. BIOMOD: a platform for ensemble forecasting of species distributions. Ecography 32, 369–373.
Trautmann, S., 2018. Climate change impacts on bird species. In: Tietze, D.T. (Ed.), Bird Species, How They Arise, Modify and Vanish. Springer Nature, Switzerland, pp. 217–234.
Wagle, Y., Bhattarai, B.P., Adhikari, J.N., 2022. Factors influencing distribution and habitat utilisation of Leptoptilos javanicus in and around Barandabhar Corridor Forest, Chitwan, Nepal. Nat. Conserv. Res. 7, 19–26.
Walther, G., Post, E., Convey, P., Menzel, A., Parmesank, C., Beebee, T.J.C., et al., 2002. Ecological response to recent climate cnahge. Nature 416, 389–395.
Wiens, J.J., 2016. Climate-related local extinctions are already widespread among plant and animal species. PLoS Biol. 14, e2001104.
Yang, B., Qin, S., Xu, W., Busch, J., Yang, X., Gu, X., et al., 2020. Gap analysis of Giant Panda conservation as an example for planning China's National Park System. Curr. Biol. 30, 1287–1291.
Table
1.
Predicted suitable area (km2) for distribution and nesting site for Lesser Adjutant in Nepal under current and future climatic and land use change scenarios.
Current
2050
2070
Parameters
SSP1–2.6
SSP5–8.5
SSP1–2.6
SSP5–8.5
Distribution
21,637 (14.64%)
32,165 (21.76%)
37,076 (25.08%)
33,183 (22.45%)
43,210 (29.24%)
Nesting site
19,651 (13.29%)
29,074 (19.67%)
32,685 (22.11%)
29,763 (20.14%)
37,133 (25.12%)
The percentage in the parentheses represents the predicted area relative to the area of Nepal.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds