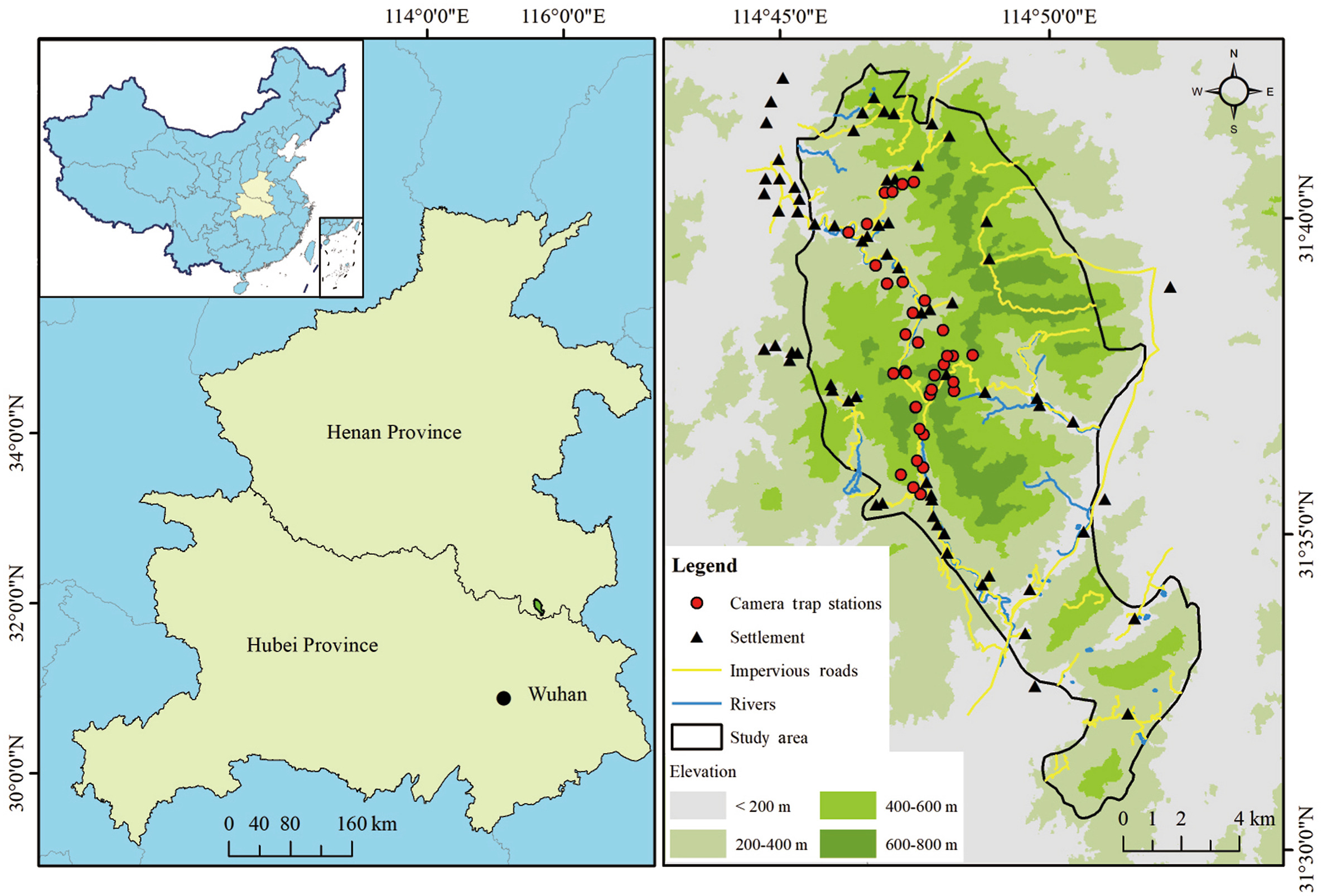
The outbreak of the COVID-19 pandemic has brought massive shifts in human activities through a global blockade, directly affecting wildlife survival. However, the indirect impacts of changes in human activities are often easily overlooked. We conducted surveys of Reeves’s Pheasant (Syrmaticus reevesii) and its sympatric species by camera traps in forest-type nature reserves in three different scenarios: pre-lockdown, lockdown and post-lockdown. An increase in livestock activities observed during the lockdown and post-lockdown period in our study area provided us an opportunity to investigate the indirect impact of the lockdown on wildlife. The pre-lockdown period was used as a baseline to compare any changes in trends of relative abundance index, activity patterns and temporal spacing of targeted species and livestock. During the lockdown period, the relative abundance index of livestock increased by 50% and there was an increase in daytime activity. Reeves's Pheasant showed avoidance responses to almost all sympatric species and livestock in three different periods, and the livestock avoidance level of Reeves's Pheasant during the lockdown period was significantly and positively correlated with the relative abundance index of livestock. Species-specific changes in activity patterns of study species were observed, with reduced daytime activities of Hog Badger and Raccoon Dog during and after the confinement periods. This study highlights the effect of the COVID-19 lockdown on the responses of wildlife by considering the changes in their temporal and spatial use before, during and after lockdown. The knowledge gained on wildlife during reduced human mobility because of the pandemic aids in understanding the effect of human disturbances and developing future conservation strategies in the shared space, to manage both wildlife and livestock.
Interspecific avian brood parasites lay their eggs in nests of other species, the hosts, which provide parental care to the parasitic eggs and chicks (Payne, 1977; Rothstein and Robinson, 1998; Davies, 2000). Brood parasitism reduces the reproductive success of hosts (Rothstein and Robinson, 1998), which select for the evolution of antiparasitic defenses (i.e., host traits that reduce the impact of parasitism, which have evolved in response to, or are currently maintained by, selection pressures arising from parasitism; Rothstein, 1990; Krüger, 2007). In turn, host defenses may select for parasite's counterdefenses and potentially create a co-evolutionary arms race (Dawkins and Krebs, 1979; Rothstein, 1990; Davies, 2011).
Interspecific avian brood parasitism has evolved independently at least seven times in birds (Sorenson and Payne, 2002): three times among the cuckoos, once in each of the honeyguides and waterfowl, and twice among songbirds: the African brood parasitic finches and the cowbirds. The cowbirds comprise a monophyletic group (Lanyon, 1992), which is comprised of five species that vary greatly in the degree of host specificity (Ortega, 1998). The basal species of this clade, the Screaming Cowbird (Molothrus rufoaxillaris) separated from the lineage leading to other parasitic cowbirds 2.8–3.8 Mya (Rothstein et al., 2002) and is one of the most specialized brood parasites, using almost exclusively the Baywing (Agelaioides badius) (Friedmann, 1929; Fraga, 1998; De Mársico et al., 2010a). By contrast, the two most recently derived species of the clade (i.e., 0.8–1.2 Mya; Rothstein et al., 2002), the Shiny Cowbird (M. bonariensis) and the Brown-headed Cowbird (M. ater), are extreme generalist parasites at the population level, with nearly 250 host species documented for each species (Lowther, 2011) and ca. 150 host species recorded rearing cowbirds. The Giant Cowbird was the first species that split from the ancestral Screaming Cowbird (Lanyon, 1992), which involved a range expansion from southern South America to northern South America and Central America (Rothstein et al., 2002). It is known presently to parasitize 11 species, mainly caciques and oropendolas (Lowther, 2011).
Hosts may exhibit various lines of defense against brood parasitism that yield different benefits to their reproductive output (Table 1). As a first line of defense, hosts may increase nest attentiveness or aggressively mob cowbirds to prevent them from gaining access to the nest (Robertson and Norman, 1976; Neudorf and Sealy, 1994; Sealy et al., 1998; Burhans et al., 2001). Nest defense would allow hosts to avoid most costs of parasitism, but in practice it appears to have limited effectiveness to prevent egg-laying by cowbird females (Neudorf and Sealy, 1994; Sealy et al., 1998; Ellison and Sealy, 2007; Gloag et al., 2012) and can bear additional costs if increased host activity at the nest facilitates its detection by brood parasites or nest predators (Robertson and Norman, 1977; Smith et al., 1984; Uyehara and Narins, 1995, Gill et al., 1997; but see Clotfelter, 1998). A second and more effective line of defense is ejection of parasitic eggs (Rohwer and Spaw, 1988; Peer et al., 2000; Lorenzana and Sealy, 2001), which eliminates the costs of a decrease in host's hatching success and chick survival. Egg ejection can be nearly cost-free if hosts are able to reliably discriminate the parasitic eggs and remove them from the nest without damaging their own eggs (Lorenzana and Sealy, 2001; Rasmussen et al., 2009). This is more likely for large-billed hosts, which can remove the cowbird egg by grasping it whole between their mandibles. For small-billed hosts, which must eject cowbird eggs by piercing the shell, ejection can be costly because hosts may accidentally damage one or more of their own eggs while attempting to remove a larger and thick-shelled cowbird egg (Rothstein, 1977; Rohwer and Spaw, 1988; Røskaft et al., 1993). Alternatively, hosts may reject parasitism by abandoning the nest or burying the parasitized clutch (Sealy, 1995;
Goguen and Mathews, 1996; Guigueno and Sealy, 2010). If subsequent re-nesting attempts are less likely to be parasitized, these tactics may rid hosts from parasitism, though at the expense of losing a complete clutch and delaying reproduction (Goguen and Mathews, 1996;
Budnik et al., 2001; Guigueno and Sealy, 2010). However, whether the abandonment of parasitized clutches represents an evolved antiparasitic defense (Hosoi and Rothstein, 2000; Guigueno and Sealy, 2010, 2011) or a generalized response to nest disturbance (Hill and Sealy, 1994; Goguen and Mathews, 1996; Kosciuch et al., 2006) is still a matter of debate. Finally, as a last line of defense hosts may reject the parasitic young. Although by the nestling stage hosts have already paid most costs of parasitism (but see Rasmussen and Sealy, 2006), they might still save time and energy for current or future reproduction by avoiding prolonged parental care of unrelated nestlings and fledglings. Puzzlingly, discrimination and/or rejection of brood parasitic young is far less common than host defenses against adult parasites and eggs, in spite of the striking differences in appearance between parasite and host nestlings in several hostparasite systems (Grim, 2006). Among cuckoo hosts, antiparasitic defenses beyond the egg stage include the desertion of parasitized broods (Langmore et al., 2003), eviction of parasitic nestlings from the nest (Sato et al., 2010a; Tokue and Ueda, 2010), and time-limited provision of parental care (Grim, 2007).
Table
1.
Summary of main antiparasitic defenses shown by hosts, costs avoided by the host as a result of the defense, costs already paid by the host at the time the defense acted and potential costs associated with the defense
Host defense
Costs avoided by the defense
Costs incurred before the defense has acted
Costs associated with the defense
Nest defense and aggression towards the parasite
Egg losses, lower hatchability, lower chick and young survival, lower future reproductive success of parents
None
Time, increased detectability of the nest by other brood parasites or predators
Egg ejection
Lower hatchability, lower chick and young survival, lower future reproductive success of parents
Egg losses
Recognition errors, breakage of own eggs, retaliation by the parasite ("mafia" behavior)
Nest abandonment
Lower hatchability, lower chick and young survival, lower future reproductive success of parents
Egg losses
Time and energy for renesting, lower reproductive success with time of breeding
Chick discrimination
Lower chick and young survival, lower future reproductive success of parents
Egg losses, lower hatchability
Recognition errors
Young discrimination
Lower young survival, lower future reproductive success of parents
Given the substantial costs of brood parasitism to host's fitness, it continues to be intriguing why some hosts have not evolved effective defenses against parasitic eggs or young. The evolutionary lag hypothesis states that defenses, though adaptive, may be absent because there has not been sufficient time for those behaviors to spread within the host population (Rothstein, 1975). In support of this hypothesis, some studies have revealed differences in the response to parasitism between host populations with varying degrees of sympatry with, or exposure to the parasite (Soler and Møller, 1990;
Robert and Sorci, 1999; Hosoi and Rothstein, 2000). Alternatively, the equilibrium hypothesis proposes that antiparasitic defenses are not always adaptive as these may entail costs that outweigh the costs of parasitism. Putative costs of host's defenses include recognition errors (i.e., mistakenly rejecting an own egg or chick at unparasitized nests [Davies and Brooke, 1988; Marchetti, 1992; Lotem et al., 1995; Sato et al., 2010a]), rejection errors (i.e., mistakenly rejecting an own egg or chick at parasitized nests [Davies and Brooke, 1988; Davies et al., 1996; Lorenzana and Sealy, 2001]), accidental damage to host eggs during ejection attempts (Rohwer et al., 1989; Røskaft et al., 1993; Underwood and Sealy, 2006;
Antonov et al., 2009), increased risk of nest failure following the desertion of parasitized nests (Krüger, 2011), and possible retaliation by adult parasites after the rejection of parasite eggs (Hoover and Robinson, 2007). Evidence for the equilibrium hypothesis is indirect and based on studies documenting the existence of costs associated to hosts' antiparasitic defenses (Rohwer et al., 1989; Lotem et al., 1992; Antonov et al., 2009; Krüger, 2011).
The different lines of defenses by hosts of the Brownheaded Cowbird have been relatively well studied and reviewed in several works (i.e., Rothstein, 1975; Sealy et al., 1998; Hosoi and Rothstein, 2000; Peer and Sealy, 2004). On the contrary, antiparasitic defenses by hosts of South American cowbirds have been less studied and, so far, the results have not been comprehensively reviewed. In this work we present new experimental data on egg rejection in a host of the Shiny Cowbird, the House Wren (Troglodytes aedon), review the different lines of defense against parasitism evolved by hosts of South American cowbirds, and examine whether hosts of the most derived cowbird species differ from those of the more ancestral ones in the type and extent of defenses evolved. In addition, we discuss potential constraints and costs associated with antiparasitic behaviors that could explain why some hosts lack defenses against cowbird parasitism.
Methods
study area
Experiments that simulated parasitism on House Wrens were conducted at "Reserva de Fauna y Flora El Destino" (35°08′S, 57°23′W) near the town of Magdalena, province of Buenos Aires, Argentina. The study site is within the Biosphere Reserve "Parque Costero del Sur"
(MAB-UNESCO) and consists of semi-open grassland with patches of xeric thorny woodlands arranged in several strips, 20–100 m in width and up to several kilometers in length, parallel to the edge of the "de la Plata" river. These patches are mainly dominated by native tree species such as Celtis tala, Scutia buxifolia and Schinus longifolius.
study species
House Wrens nest in natural and artificial cavities. To facilitate data collection, we placed approximately 100 nest-boxes of 25 × 17 × 13 cm (height, width and depth) in trees at heights of 1.5–1.8 m and at least 20 m apart. At our study site, House Wrens breed from early October until mid-January and it is a common host of Shiny Cowbirds, with a frequency of parasitism of 60% and an intensity of 1.7 eggs per parasitized nest (Tuero et al., 2007). Wrens lay a modal clutch size of 5 eggs and 57% of parasitism occurs during host laying (Fiorini et al., 2009a).
At our study site, Shiny Cowbirds lay eggs from late September to mid-January, mainly in nests of Chalkbrowed Mockingbirds (Mimus saturninus) (frequency of parasitism 60–70% [Fiorini et al., 2009a, Gloag et al., 2012]) and House Wrens. In eastern Argentina and neighboring parts of Uruguay and Brazil, Shiny Cowbird eggs are immaculate white or spotted, with a highly variable ground color and size of the spots (Mahler et al., 2008).
Experimental parasitism
We experimentally parasitized 42 House Wren nests with unincubated Shiny Cowbird eggs collected from active Chalk-browed Mockingbird nests. As mockingbirds eject white immaculate Shiny Cowbird eggs (see results), we created eggs of the white morph by coating spotted cowbird eggs with white acrylic paint, matching the appearance of real eggs (de la Colina et al., 2012). Similarly, to standardize the type of spots in the treatment with those on spotted eggs we painted the eggs with the same background color and spotting pattern. We conducted the experiments during host's laying period without removing a host's egg in conjunction with experimental parasitism because Shiny Cowbirds do not remove host eggs in association with laying. They peck (and sometimes puncture) one or more host eggs, instead, which afterwards are removed by the host (Astié and Reboreda, 2006). We checked nests daily for egg ejection and at each visit we examined host and parasite eggs for cracks or punctures and determined whether the nest was active or abandoned. We considered a parasitic egg ejected if it was gone, and accepted if it remained in the nest for two days after its introduction. We considered that the egg was rejected by nest desertion if the nest was abandoned (i.e., no longer attended by adults) the day after the event of experimental parasitism. We conducted another 16 experiments with natural Shiny Cowbird eggs (8 with white painted spotted eggs and 8 with naturally spotted eggs) in which we left the parasite egg in the nest of House Wrens for five days and in none of these cases the egg was rejected (data not shown).
Analyses of data from other studies
We reviewed the literature on antiparasitic defenses in hosts of Screaming, Giant and Shiny cowbirds. To analyze nest defenses we considered studies in which hosts were presented simultaneously or sequentially with taxidermic models of the female parasite and a control species (i.e., Gill and Sealy, 1996; Røskaft et al., 2002). We considered three categories of nest defensive behaviors performed by either member of the pair: (1) attacks and close passes directed to the model (attack),
(2) approaches towards the model (approach), and (3) moving to the nest and sitting in it (staying in the nest). We considered that any of these nest defensive behaviors was an antiparasitic defense when the level of response towards the parasite model was significantly higher than that directed towards the control model. Most of these studies used one model per species only and, therefore, they cannot discriminate whether the differential responses towards models where because they were different species or different models. Because of the limited experimental data available for hosts of the Giant Cowbird, we also included observations of nest defense from those species.
To analyze egg rejection we considered studies with experiments that simulated parasitism with at least 10 replicates. For hosts of the Shiny Cowbird, we did not include studies conducted in the Caribbean (i.e., Post et al., 1990), because sympatry between Shiny Cowbirds and hosts' populations in this region is recent (i.e., less than 100 years; Post and Wiley, 1977a) and therefore, the time of interaction between host and parasites may not have been long enough for the evolution of antiparasitic defenses. In addition, we preferentially included studies that tested rejection of white immaculate and spotted Shiny Cowbird eggs (the only exception was Kattan's [1998] study) because we wanted to test whether the level of mismatching in shell coloration between host and parasite eggs was a cue for eggrejection. These studies considered that the parasite egg was accepted if it remained in the nest for at least five days after the experimental introduction, or otherwise rejected (Rothstein, 1975). In addition, the egg was considered rejected by nest desertion if the eggs were cold and adults no longer attended the nest the day after the event of experimental parasitism.
Results
Nest defense
Screaming Cowbird
De Mársico and Reboreda (2008a) tested nest defenses in Baywings by presenting dummy models of female Screaming and Shiny cowbirds, and a female Whitebrowed Blackbird (Sturnella supercilliaris), as a control species. Baywings attacked the Screaming Cowbird model more than the control model, and approached that model more often and spent a greater proportion of time close to it than to the control model. The rate of attacks, number of approaches and time spent close to the Shiny Cowbird model were intermediate, but did not differ significantly from those of the other two models.
Giant Cowbird
Observations at four hosts (two caciques and two oropendolas) showed that Giant Cowbirds were attacked by hosts when approaching their nests. Robinson (1988) observed that Yellow-rumped Caciques (Cacicus cela) and Russet-backed Oropendolas (Psarocolius angustifrons) were invariably aggressive to Giant Cowbirds.
Similarly, Webster (1994) reported that Giant Cowbirds that approached colonies of Montezuma Oropendolas (P. montezuma) were attacked and driven away from the colony, and Fraga (2011) reported attacks and chases by male and female Red-rumped Caciques (C. haemorrhous) on Giant Cowbirds perched near the colony.
Shiny Cowbird
Table 2 shows the results of studies that evaluated nest defenses against taxidermic models of Shiny Cowbirds: attacks and approaches towards the model and staying in the nest. Only three of the eight hosts tested in those studies, the House Wren, Chalk-browed Mockingbird and Brown-and-yellow Marshbird (Pseudoleistes virescens) attacked Shiny Cowbird models at a higher rate than control models. Mockingbirds also approached the Shiny Cowbird model at a higher rate than the control model.
Baywings ejected all parasite eggs (whether Screaming Cowbird or Shiny Cowbird) laid before their own first egg and when already heavily parasitized (i.e., 3–4 parasite eggs), they ejected the entire clutch and laid a replacement clutch in the same nest (Fraga, 1998, De Mársico et al., in press). Although ejected and replacement clutches did not differ in parasitism frequencies, the latter received significantly fewer cowbird eggs and the clutch contained more Baywing eggs (De Mársico et al., in press). Video recordings showed that Baywings consistently ejected intact eggs by kicking them out of the nest cup using their feet (De Mársico et al., in press). Regarding the two alternative hosts of Screaming Cowbirds, Brown-and-yellow Marshbird and Chopi Blackbird (Gnorimopsar chopi), although there are no experimental studies analyzing egg rejection by these hosts, observational data indicate that they do not eject Screaming Cowbird eggs (Mermoz and Fernández, 2003; De Mársico et al., 2010b).
Giant Cowbird
In a controversial study, Smith (1968) reported that two hosts of the Giant Cowbird, Chestnut-headed Oropendola (P. wagleri) and Yellow-rumped Cacique, either ejected non-mimetic eggs (i.e., discriminators) or accepted them (i.e., non-discriminators), depending on whether they nested in colonies near nests of aggressive wasps, which protected colonies from botfly (Diptera: Philornis) parasitism. Smith recorded that Giant Cowbird nestlings removed subcutaneous botfly larvae from themselves and from their nest mates, therefore, benefiting the host because this in turn favored acceptance of parasitic eggs. Cunningham and Lewis (2006), in a study of egg rejection in Montezuma Oropendola, compared egg rejection rates in four experimental treatments in which a Giant Cowbird egg was added to clutches of one or two host eggs, with and without removal of one host egg. The overall frequency of rejection was 72%, regardless of whether there was another egg in the nest to which females could compare the model cowbird egg.
Shiny Cowbird
Table 3 shows the results of studies evaluating egg rejection of white immaculate and spotted Shiny Cowbird eggs in 14 hosts. Two hosts (Crested Cardinal Paroaria coronata and Rufous Hornero Furnarius rufus) ejected Shiny Cowbird eggs of the white and spotted morphs, four hosts ejected only white eggs and 7 hosts accepted all eggs of both morphs. The remaining host, the Baywing accepted white and spotted eggs, except when nests were already heavily parasitized, in which case the entire clutches were ejected. In the Crested Cardinal the cue used to eject parasite eggs was divergence in color or a combination of color and shape (Segura and Reboreda, 2012), whereas in the Rufous Hornero egg size was the cue (they ejected eggs with a width of less than 88% that of their own eggs [Mason and Rothstein, 1986]). The selective pressure imposed by Rufous Horneros has favored, as a counterdefense, the increase in the size of parasitic eggs in areas where they are commonly parasitized by Shiny Cowbirds (Mason and Rothstein, 1986; Di Giacomo AG, Massoni V, Reboreda JC, unpublished data). The four host species that ejected white Shiny Cowbird eggs used divergence in color as a cue as all of them have spotted eggs. Of the six ejector species, five were grasp-ejectors, whereas one, the Crested Cardinal, was a puncture-ejector. Most non-ejector species weighed less than Shiny Cowbirds (i.e., < 32 g) and, except for 2 of 42 cases in House Wrens and 3 of 11 in Rufous-collared Sparrows (Zonotrichia capensis), they did not desert the nest in association with experimental parasitism.
Screaming Cowbird nestlings and fledglings look identical to those of Baywings (Fig. 1A, B) and it has been proposed that such a resemblance is an adaptation in response to host discrimination against odd-looking young (Fraga, 1998). De Mársico et al. (2012) tested this hypothesis by comparing the survival rates of young Screaming Cowbird and non-mimetic Shiny Cowbirds cross-fostered to Baywing nests. They found that Shiny Cowbirds suffered higher post-fledging mortality rates than Screaming Cowbirds due to host rejection. These authors also quantified the similarity in plumage color and begging calls between host and cowbird fledglings and found that only Screaming were indistinguishable from host young in plumage color and begging calls, supporting the hypothesis that visual and vocal similarity between Screaming Cowbirds and Baywings was a counterdefense driven by host discrimination against non-mimetic juveniles.
Figure
1.
Photographs of (A) nestlings (7 days of age) and (B) fledglings (3 weeks after fledging) of Baywing (Agelaioides badius, host) and Screaming Cowbird (Molothrus rufoaxillaris, parasite). (C) Nest of House Wren (Troglodytes aedon) parasitized with two Shiny Cowbird (M. bonariensis) eggs, one white immaculate and the other spotted. (D) Nest of Chalk-browed Mockingbird (Mimus saturninus) parasitized with 4 Shiny Cowbird spotted eggs. Photographs were taken by (A) María C. De Mársico, (B) Cynthia Ursino, (C) Diego T. Tuero and (D) Vanina D. Fiorini.
Lichtenstein (2001) reported that Shiny Cowbird chicks were fed significantly less frequently than host chicks in nests of the Rufous-bellied Thrush (Turdus rufiventris), and suggested that parents avoided feeding them. However, in her study and in another study of the same host (Sackmann and Reboreda, 2003), most Shiny Cowbird chicks survived, which indicates that even if hosts discriminate between their own and parasitic chicks, such discrimination apparently is not effective enough to preclude successful parasitism. More recently (Delhey et al., 2011), a putative case of early discrimination of cowbird nestlings was described in the Firewood-gatherer (Anumbius annumbi), a host known to accept nonmimetic, spotted Shiny Cowbird eggs (Mason, 1986). These authors reported that, although cowbird eggs usually hatched 4–5 days before host eggs, all parasitic nestlings died within 48 h, whereas hosts continued with their breeding attempts. They proposed that cowbird deaths were likely due to neglect because little food was found in stomachs of dead nestlings and this could be due to differences in visual or acoustic appearance between host and parasitic hatchlings.
Other antiparasitic defenses
De Mársico and Reboreda (2008b) reported that in Baywings the length of the prelaying period (time since nest lining was completed until the host laid its first egg) ranged from one to 19 days. Latency of Screaming Cowbird parasitism (time since nest lining was completed until laying of the first parasite egg) was 1–3 days. As a result, more than 30% of parasitic eggs were laid before hosts had started laying and were ejected. These authors proposed that Baywings' prelaying behavior precludes an accurate synchronization of parasitism with the host's laying and, therefore, may act as an antiparasitic defense, as it decreases the incidence of successful parasitism.
Discussion
Most hosts of South American cowbirds have evolved some type of defense regardless of the presumed time of interaction with its brood parasite. Although egg ejection was the most common defense, there is evidence of defensive behavior against cowbirds at nests of hosts of the three parasites, and chick discrimination in the main host of the Screaming Cowbird. There also is indirect evidence of chick discrimination in two hosts of the Shiny Cowbird, but further studies are necessary to confirm whether this behavior is truly an antiparasitic defense. Most species that eject cowbird eggs were grasp-ejectors with only one documented case of puncture-ejection in a host of the Shiny Cowbird (the Crested Cardinal) and one case of egg ejection using the feet in the main host of the Screaming Cowbird (the Baywing). In response to host's defenses, the more ancestral Screaming Cowbird and the more derived Shiny Cowbird have evolved some type of counterdefenses, such as laying a larger egg size by Shiny Cowbirds parasitizing Rufous Horneros, and mimicking plumage color and begging calls of fledglings when Screaming Cowbirds parasitize Baywings. Thus, for most hosts analyzed the time they have coevolved with its brood parasite has been enough to favor the evolution of at least one line of defense against parasitism and in a few cases, the evolution of a reciprocal counterdefense in the parasite. However, some hosts apparently do not exhibit antiparasitic defenses. Below, we discuss some of these cases and provide possible explanations.
Lack of egg rejection in small hosts of the shiny Cowbird
Peer and Sealy (2004) analyzed correlates of egg rejection in hosts of the Brown-headed Cowbird using the pairwise comparative method and found that rejecter species are larger and have larger nests and bills than accepter species. They also found that species with longer periods of contact with the parasite show greater rejection behavior than those with shorter periods and proposed that this result supports evolutionary lag hypothesis (Rothstein, 1975).
Similarly, all small hosts of the Shiny Cowbird that have been tested for egg rejection accept parasite eggs and, in most cases, do not abandon parasitized nests although the degree of mismatching in size and coloration between host and parasite eggs is remarkable (Fig. 1C). One possible explanation for the absence of egg rejection in these hosts would be that there has been not enough time for those behaviors to evolve and spread within the host population (evolutionary lag hypothesis). However, at least one of the hosts (the House Wren) increases nest defensive behaviors when there is a Shiny Cowbird model close to its nest (De Mársico and Reboreda, 2008a), which suggests that it recognizes the female cowbird as a threat to its nest, thus, indicating that the time of interaction with the parasite has been long enough for the evolution of this type of antiparasitic defense. In addition, Shiny Cowbird hosts larger in body mass than the parasite have likely similar histories of sympatry than smaller hosts (i.e., they also nest in relatively open habitats and their ranges overlap the historic range of the parasite), but in most cases are egg ejectors. One explanation consistent with this pattern is that small hosts cannot puncture eject Shiny Cowbird eggs and because they have small bills they cannot grasp eject the eggs either. All cowbird species have thicker eggshells than nonparasitic icterids (Spaw and Rower, 1987; Rahn et al., 1988; Mermoz and Ornelas, 2004). The thicker eggshells of cowbirds may increase the costs of puncture-ejecting parasite eggs, forcing small-billed hosts to accept them (Rohwer and Spaw, 1988; Picman, 1989; Rohwer et al., 1989; Røskaft et al., 1993). Accordingly, the only host that puncture-ejects Shiny Cowbird eggs is the Crested Cardinal (Segura and Reboreda, 2012), which has a larger body mass than nonejector species (Table 3), but a bill not large enough to grasp eject cowbird eggs (but see Underwood and Sealy[2006] for a case of a small host in which bill size does not constrain grasp-ejection). If this interpretation if correct, we expect medium sized hosts of Shiny Cowbirds (i.e., 35–45 g) will be puncture-ejectors.
Although thicker eggshells in Shiny Cowbirds may preclude small hosts from puncturing parasite eggs, it is not clear why they do not abandon the nest or bury the parasitized clutch, as it happens in some hosts of the Brown-headed Cowbirds (Goguen and Mathews, 1996; Guigueno and Sealy, 2011). This behavior would be particularly advantageous for small hosts, as Shiny Cowbird parasitism reduces severely the hatching success of eggs and the survival of chicks (Fraga, 1978;
Cruz and Andrews, 1997; Kattan, 1998; Tuero et al., 2007). Possible explanations for not abandoning the nest would be high costs of delaying reproduction and low probability to escape parasitism or depredation in future reproductive attempts (Kattan, 1998). Accordingly, Krüger (2011) found that Cape Bulbuls (Pycnonotus capensis), a small host of the Jacobin Cuckoo (Clamator jacobinus), do not eject parasitic eggs or desert the nest after being parasitized. Jacobin Cuckoos have large eggs with thick shells, making it almost impossible for Cape Bulbuls to puncture-eject them. In addition, higher predation and parasitism risks later in the season make nest desertion more costly for hosts than to accept the cuckoo egg (Krüger, 2011). Some of the small hosts of the Shiny Cowbird could have experienced a similar evolutionary scenario, which may have precluded the evolution of nest desertion. Interestingly, Kattan (1998) reported that House Wrens abandoned most nests that received more than two Shiny Cowbird eggs, but continued incubation in nests with one or two parasite eggs. This suggests that the decision of abandoning the nest by House Wrens is flexible and would depend on the relative costs of deserting the nest vs. accepting parasitism. Further studies analyzing the costs associated with abandoning the nest vs. accepting parasite eggs are necessary to understand the lack of defenses in most small hosts of Shiny Cowbirds.
Is there any benefit in accepting parasite eggs?
Most hosts larger than the Shiny Cowbird grasp-eject parasite eggs of the white morph, but accept those of the spotted morph (Fig. 1D). These hosts lay spotted eggs, which suggests that they use the level of mismatching in shell coloration between their own and the parasitic eggs as a cue for egg-rejection (i.e., Peer et al., 2002; Moskát et al., 2008). In agreement with this explanation, de la Colina et al. (2012) found that the presence of spotting in Shiny Cowbird eggs significantly decreased the probability of ejection by Chalk-browed Mockingbirds, whereas increments in brightness significantly increased rejection frequencies. These authors proposed that the cognitive rules underlying mockingbird rejection can be explained by a decision-making model that predicts changes in the levels of rejection in direct relation to the number of relevant attributes shared between host and parasite eggs. However, in a study conducted in the same host,
Gloag et al. (2012) reported a potential benefit of accepting parasite eggs, as these eggs dilute the probability of host egg losses in subsequent parasite attacks and may favor egg acceptance provided that parasitism does not affect host's offspring survival. Mockingbird nests with manipulated clutch compositions were more likely to escape puncture by female cowbirds as the number of cowbird eggs in the host's clutch increased. They also showed through a Monte Carlo simulation that acceptors enjoy higher egg survivorship than rejecters in host populations where multiple parasitism occurs. This "dilution effect" had been previously proposed by Sato et al. (2010b) to explain why Large-billed Gerygones (Gerygone magnirostris) sometimes reject parasite nestlings but not eggs. Results of Gloag et al.'s (2012) study raises the question of why do mockingbirds eject white immaculate Shiny Cowbird eggs if they can bear a net benefit to host's reproductive success. Recent experiments conducted in this host (De Mársico MC, unpublished results) indicate that the presence of white immaculate parasite eggs, but not spotted parasite eggs, increases the probability of depredation of mockingbird nests and suggest that the ejection of white immaculate Shiny Cowbird eggs in this host may have been driven by other selective pressures. Studies analyzing whether this "dilution effect" is present in multiple parasitized hosts of parasitic species that puncture or remove eggs may also help us to understand why some hosts do not eject parasite eggs.
Concluding remarks
South American cowbirds provide a very fruitful model for studying the evolution of antiparasitic defenses. This group includes parasites with different degrees of specialization in host use, from the extreme specialist Screaming Cowbird, to the extreme generalist Shiny Cowbird. This allows us to study processes of pairwise as well as diffuse coevolution (Rothstein, 1990). Besides, Giant and Shiny cowbirds use hosts that differ markedly in body size, with some hosts much smaller and others much larger than the parasite (Ortega, 1998;
Lowther, 2011). Costs of parasitism to host's fitness vary considerably according to the differences in body mass between parasites and hosts. For hosts larger than the parasite the main cost is the ruin or removal of their eggs (Sackmann and Reboreda, 2003; Astié and Reboreda, 2006), whereas hosts smaller than the parasites pay additional costs as parasitism lowers the hatchability of their eggs and survival of their chicks (Payne and Payne, 1998; Hoover, 2003; Tuero et al., 2007). Thus, different lines of antiparasitic defense are expected depending on the differences in size between hosts and parasites. Finally, the Shiny Cowbird has expanded considerably its range over the past century (Cruz et al., 1985; 1999) and during this expansion has included new hosts species that lack a history of sympatry with the parasite (Post and Wiley, 1977a, 1977b; Briskie et al., 1992). These new hosts provide a natural experiment to study the evolution of antiparasitic defenses (i.e. Cruz and Wiley, 1989;
Post et al., 1990; Briskie et al., 1992; Robert and Sorci, 1999; Cruz et al., 2008). Most hosts of Giant and Shiny cowbirds remain unstudied and may provide very useful insights to unravel the evolutionary mechanisms underlying the development of host's defenses against parasitism.
Acknowledgements
We thank "Fundación Elsa Shaw de Pearson" for allowing us to conduct part of this study at "Reserva de Flora y Fauna El Destino". We greatly appreciate comments on a previous version of this manuscript by Bruce Lyon, Anders Møller and Spencer Sealy. MCDM, VDF and JCR are Research Fellows of Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). This work was supported by grants of Agencia Nacional de Promoción Científica y Tecnológica and Universidad de Buenos Aires.
Arora, S., Bhaukhandi, K.D., Mishra, P.K., 2020. Coronavirus lockdown helped the environment to bounce back. Sci. Total Environ. 742, 140573. .
Bar, H., 2021. COVID-19 lockdown: animal life, ecosystem and atmospheric environment. Environ. Dev. Sustain. 23, 8161-8178. .
Bates, A.E., Primack, R.B., Moraga, P., Duarte, C.M., 2020. COVID-19 pandemic and associated lockdown as a "Global Human Confinement Experiment" to investigate biodiversity conservation. Biol. Conserv. 248, 6. .
Bates, A.E., Primack, R.B., Biggar, B.S., Bird, T.J., Clinton, M.E., Command, R.J., et al., 2021. Global COVID-19 lockdown highlights humans as both threats and custodians of the environment. Biol. Conserv. 263, 109175. .
Behera, A.K., Kumar, P.R., Priya, M.M., Ramesh, T., Kalle, R., 2022. The impacts of COVID-19 lockdown on wildlife in Deccan Plateau, India. Sci. Total Environ. 822, 153268. .
Bischof, R., Ali, H., Kabir, M., Hameed, S., Nawaz, M.A., 2014. Being the underdog: an elusive small carnivore uses space with prey and time without enemies. J. Zool. 293, 40-48. .
Bogdan, V., Junek, T., Vymyslicka, P.J., 2016. Temporal overlaps of feral cats with prey and competitors in primary and human-altered habitats on Bohol Island, Philippines. PeerJ 4, e2288. .
Budiman, I., Kusumaratna, R.K., 2021. Human-nature interactions through the lens of global pandemics: a review. Hum. Ecol. 28, 15-24. .
Chen, X., Fu, F., 2022. Highly coordinated nationwide massive travel restrictions are central to effective mitigation and control of COVID-19 outbreaks in China. Proc. R. Soc. A 478, 20220040. .
China Meteorological Administration, 2020. China Meteorological Administration Land Surface Data Assimilation System (CLDAS-V2.0) real-time product dataset. .
Chowdhury, R.B., Khan, A., Mahiat, T., Dutta, H., Tasmeea, T., Arman, A.B.B., et al., 2021. Environmental externalities of the COVID-19 lockdown: Insights for sustainability planning in the Anthropocene. Sci. Total Environ. 783, 147015. .
Cooke, S.J., Cramp, R.L., Madliger, C.L., Bergman, J.N., Reeve, C., Rummer, J.L., et al., 2021. Conservation physiology and the COVID-19 pandemic. Conserv. Physiol. 9, coaa139. .
Corradini, A., Peters, W., Pedrotti, L., Hebblewhite, M., Bragalanti, N., Tattoni, C., et al., 2021. Animal movements occurring during COVID-19 lockdown were predicted by connectivity models. Glob. Ecol. Conserv. 32, e01895. .
Cusack, J.J., Dickman, A.J., Kalyahe, M., Rowcliffe, J.M., Carbone, C., MacDonald, D.W., et al., 2017. Revealing kleptoparasitic and predatory tendencies in an African mammal community using camera traps: a comparison of spatiotemporal approaches. Oikos 126, 812-822. .
Derryberry, E.P., Phillips, J.N., Derryberry, G.E., Blum, M.J., Luther, D., 2021. Singing in a silent spring: birds respond to a half-century soundscape reversion during the COVID-19 shutdown. Integer. Comp. Biol. 61, E197-E198.
Di Bitetti, M.S., Iezzi, M.E., Cruz, P., Varela, D., De Angelo, C., 2020. Effects of cattle on habitat use and diel activity of large native herbivores in a South American rangeland. J. Nat. Conserv. 58, 125900. .
Dirzo, R., Young, H.S., Galetti, M., Ceballos, G., Isaac, N.J.B., Collen, B., 2014. Defaunation in the Anthropocene. Science 345, 401-406. .
Dormann, C.F., Elith, J., Bacher, S., Buchmann, C., Carl, G., Carre, G., et al., 2013. Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36, 27-46. .
.
Gaynor, K.M., Brown, J.S., Middleton, A.D., Power, M.E., Brashares, J.S., 2019. Landscapes of fear: spatial patterns of risk perception and response. Trends Ecol. Evol. 34, 355-368. .
Geldmann, J., Joppa, L.N., Burgess, N.D., 2014. Mapping change in human pressure globally on land and within protected areas. Conserv. Biol. 28, 1604-1616. .
Gil, M.A., Baskett, M.L., Munch, S.B., Hein, A.M., 2020. Fast behavioral feedbacks make ecosystems sensitive to pace and not just magnitude of anthropogenic environmental change. P. Natl. Acad. Sci. USA 117, 25580-25589. .
Gomez, H., Wallace, R.B., Ayala, G., Tejada, R., 2005. Dry season activity periods of some Amazonian mammals. Stud. Neotrop. Fauna Environ. 40, 91-95. .
Hua, J.Q., Lu, S., Song, K., Wang, J.Y., Wang, J.F., Xu, J.L., 2022. Effects of livestock grazing on spatio-temporal patterns and behaviour of Reeves’s Pheasant Syrmaticus reevesii. Animals 12, 2968. .
Hughes, I., 2000. Biological consequences of global warming: is the signal already apparent? Trends Ecol. Evol. 15, 56-61. .
Jiménez-Valverde, A., 2012. Insights into the area under the receiver operating characteristic curve (AUC) as a discrimination measure in species distribution modelling. Global Ecol. Biogeogr. 21, 498-507. .
Johann, F., Handschuh, M., Linderoth, P., Dormann, C.F., Arnold, J., 2020. Adaptation of wild boar (Sus scrofa) activity in a human-dominated landscape. BMC Ecol. 20, 4. .
Jones, K.R., Venter, O., Fuller, R.A., Allan, J.R., Maxwell, S.L., Negret, P.J., et al., 2018. One-third of global protected land is under intense human pressure. Science 360, 788-791. .
Karanth, K.U., Srivathsa, A., Vasudev, D., Puri, M., Parameshwaran, R., Kumar, N.S., 2017. Spatio-temporal interactions facilitate large carnivore sympatry across a resource gradient. Proc. R. Soc. B 284, 20161860. .
Koju, N.P., Kandel, R.C., Acharya, H.B., Dhakal, B.K., Bhuju, D.R., 2021. COVID-19 lockdown frees wildlife to roam but increases poaching threats in Nepal. Ecol. Evol. 11, 9198-9205. .
Kraemer, M.U.G., Yang, C.H., Gutierrez, B., Wu, C.H., Klein, B., Pigott, D.M., et al., 2020. The effect of human mobility and control measures on the COVID-19 epidemic in China. Science 368, 493-497. .
Kupferschmidt, K., Cohen, J., 2020. China's aggressive measures have slowed the coronavirus. They may not work in other countries. Science 2. .
Lecchini, D., Brooker, R.M., Waqalevu, V., Gairin, E., Minier, L., Berthe, C., et al., 2021. Effects of COVID-19 pandemic restrictions on coral reef fishes at eco-tourism sites in Bora-Bora, French Polynesia. Mar. Environ. Res. 170, 105451. .
Li, P., 2021. Conceptualizing China's spatial lockdown during the COVID-19 pandemic: a neo-liberal society or a pre-liberal one? Soc. Transf. Chin. Soc. 17, 101-108. .
Lindsey, P., Allan, J., Brehony, P., Dickman, A., Robson, A., Begg, C., et al., 2020. Conserving Africa's wildlife and wildlands through the COVID-19 crisis and beyond. Nat. Ecol. Evol. 4, 1300-1310. .
Liu, L., Lin, Q.H., Liang, Z.R., Du, R.G., Zhang, G.Z., Zhu, Y.H., et al., 2021. Variations in concentration and solubility of iron in atmospheric fine particles during the COVID-19 pandemic: An example from China. Gondwana Res. 97, 138-144. .
Loh, H.C., Looi, I., Ch'ng, A.S.H., Goh, K.W., Ming, L.C., Ang, K.H., 2021. Positive global environmental impacts of the COVID-19 pandemic lockdown: a review. GeoJournal 87, 4425-4437. .
Lu, S., Liu, Z.X., Tian, S., Song, K., Hu, Q., Xu, J.L., 2022. Sex-specific movement responses of Reeves’s Pheasant to human disturbance: Importance of body characteristics and reproductive behavior. Animals 12, 1619. .
Madhusudan, M.D., 2004. Recovery of wild large herbivores following livestock decline in a tropical Indian wildlife reserve. J. Appl. Ecol. 41, 858-869. .
Manenti, R., Mori, E., Di Canio, V., Mercurio, S., Picone, M., Caffi, M., et al., 2020. The good, the bad and the ugly of COVID-19 lockdown effects on wildlife conservation: Insights from the first European locked down country. Biol. Conserv. 249, 108728. .
Markov, N., Pankova, N., Morelle, K., 2019. Where winter rules: Modeling wild boar distribution in its north-eastern range. Sci. Total Environ. 687, 1055-1064. .
Massei, G., Genov, P.V., 2004. The environmental impact of wild boar. Galemys 16, 135-145. .
McGinlay, J., Gkoumas, V., Holtvoeth, J., Fuertes, R.F.A., Bazhenova, E., Benzoni, A., et al., 2020. The impact of COVID-19 on the management of European protected areas and policy implications. Forests 11, 1214. .
Mishra, C., Wieren, S., Heitkonig, I., Prins, H., 2002. A theoretical analysis of competitive exclusion in a Trans-Himalayan large-herbivore assemblage. Anim. Conserv. 5, 251-258. .
Meredith, M., Ridout, M., 2021. Overlap: Estimates of coefficient of overlapping for animal activity patterns. R package version 0.3.4. .
Mishra, C., Van Wieren, S.E., Ketner, P., Heitkonig, I.M.A., Prins, H.H.T., 2004. Competition between domestic livestock and wild bharal Pseudois nayaur in the Indian Trans-Himalaya. J. Appl. Ecol. 41, 344-354. .
Muche, M., Yemata, G., Molla, E., Muasya, A.M., Tsegay, B.A., 2022. COVID-19 lockdown and natural resources: a global assessment on the challenges, opportunities, and the way forward. Bull. Natl. Res. Cent. 46, 20. .
Neupane, D., 2020. How conservation will be impacted in the COVID-19 pandemic. Wildl. Biol., wlb.00727 , 2020.
Niedballa, J., Wilting, A., Sollmann, R., Hofer, H., Courtiol, A., 2019. Assessing analytical methods for detecting spatiotemporal interactions between species from camera trapping data. Remote Sens. Ecol. Con. 5, 272–285. .
O'Brien, T.G., Kinnaird, M.F., Wibisono, H.T., 2003. Crouching tigers, hidden prey: Sumatran tiger and prey populations in a tropical forest landscape. Anim. Conserv. 6, 131-139. .
Paital, B., 2020. Nurture to nature via COVID-19, a self-regenerating environmental strategy of environment in global context. Sci. Total Environ. 729, 139088. .
Parsons, A.W., Bland, C., Forrester, T., Baker-Whatton, M.C., Schuttler, S.G., McShea, W.J., et al., 2016. The ecological impact of humans and dogs on wildlife in protected areas in eastern North America. Biol. Conserv. 203, 75–88. .
Pine, M.K., Wilson, L., Jeffs, A.G., McWhinnie, L., Juanes, F., Sceuderi, A., et al., 2021. A Gulf in lockdown: How an enforced ban on recreational vessels increased dolphin and fish communication ranges. Global Change Biol. 27, 4839-4848. .
Pudyatmoko, S., 2017. Free-ranging livestock influence species richness, occupancy, and daily behaviour of wild mammalian species in Baluran National Park, Indonesia. Mamm. Biol. 86, 33-41. .
R Core Team, 2021. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. .
Ridout, M.S., Linkie, M., 2009. Estimating overlap of daily activity patterns from camera trap data. J. Agric. Biol. Environ. Stat. 14, 322-337. .
Royle, J.A., Link, W.A., 2006. Generalized site occupancy models allowing for false positive and false negative errors. Ecology 87, 835-841. .
Rutz, C., Loretto, M.C., Bates, A.E., Davidson, S.C., Duarte, C.M., Jetz, W., et al., 2020. COVID-19 lockdown allows researchers to quantify the effects of human activity on wildlife. Nat. Ecol. Evol. 4, 1156-1159. .
Smith, A.T., Yan, X., Robert, S.H., Darrin, L., John, M., Don, E.W., et al., 2010. A Guide to the Mammals of China. Princeton University Press, Princeton.
Smith, M.K.S., Smit, I.P.J., Swemmer, L.K., Mokhatla, M.M., Freitag, S., Roux, D.J., et al., 2021. Sustainability of protected areas: Vulnerabilities and opportunities as revealed by COVID-19 in a national park management agency. Biol. Conserv. 255. 108985, .
Sollmann, R., 2018. A gentle introduction to camera-trap data analysis. Afr. J. Ecol. 56, 740-749. .
Sun, Z.B., Zhang, H., Yang, Y.F., Wan, H., Wang, Y.X., 2020. Impacts of geographic factors and population density on the COVID-19 spreading under the lockdown policies of China. Sci. Total Environ. 746, 141347. .
Tomasz, P., Grzegorz, B., Bogumila, J., Leif, S., Stanislaw, Ś., Włodzimierz, J., et al., 2013. Spatiotemporal behavioral plasticity of wild boar (Sus scrofa) under contrasting conditions of human pressure: primeval forest and metropolitan area. J. Mammal. 94, 109-119. .
Tucker, M.A., Bohning-Gaese, K., Fagan, W.F., Fryxell, J.M., Van Moorter, B., Alberts, S.C., et al., 2018. Moving in the Anthropocene: Global reductions in terrestrial mammalian movements. Science 359, 466-469. .
Vimal, R., 2022. The impact of the Covid-19 lockdown on the human experience of nature. Sci. Total Environ. 803, 149571. .
Waddle, J.H., Dorazio, R.M., Walls, S.C., Rice, K.G., Beauchamp, J., Schuman, M.J., et al., 2010. A new parameterization for estimating co-occurrence of interacting species. Ecol. Appl. 20, 1467-1475. .
Wagenmakers, E.J., 2003. Model selection and multimodel inference: a practical information-theoretic approach. J. Math Psychol. .
Walther, G-R., Post, E., Convey, P., Menzel, A., Parmesan, C., Beebee, T.J.C., et al., 2002. Ecological responses to recent climate change. Nature 416, 389-395. .
Wang, Q.Y., Zhao, Y.Z., Luo X., Hua, J.Q., Xu, J.L., 2016. Potential nest predators of Syrmaticus reevesii based on camera traps and artificial nests. Chin. J. Appl. Ecol. 27, 1968-1974. . (in Chinese)
Whitworth, A., Beirne, C., Huarcaya, R.P., Whittaker, L., Rojas, S.J.S., Tobler, M.W., et al., 2019. Human disturbance impacts on rainforest mammals are most notable in the canopy, especially for larger-bodied species. Divers. Distrib. 25, 1166-1178. .
Xu, J.L., Zhang, X.H., Sun, Q.H., Zheng, G.M., Wang, Y., Zhang, Z.W., 2009. Home range, daily movements and site fidelity of male Reeves’s Pheasants Syrmaticus reevesii in the Dabie Mountains, central China. Wildlife Biol. 15, 338-344. .
Zellmer, A.J., Wood, E.M., Surasinghe, T., Putman, B.J., Pauly, G.B., Magle, S.B., et al., 2020. What can we learn from wildlife sightings during the COVID-19 global shutdown? Ecosphere 11, e03215. .
Zhang, J.D., Hull, V., Ouyang, Z.Y., Li, R.G., Connor, T., Yang, H.B., et al., 2017. Divergent responses of sympatric species to livestock encroachment at fine spatiotemporal scales. Biol. Conserv. 209, 119–129. .
Table
1.
Summary of main antiparasitic defenses shown by hosts, costs avoided by the host as a result of the defense, costs already paid by the host at the time the defense acted and potential costs associated with the defense
Host defense
Costs avoided by the defense
Costs incurred before the defense has acted
Costs associated with the defense
Nest defense and aggression towards the parasite
Egg losses, lower hatchability, lower chick and young survival, lower future reproductive success of parents
None
Time, increased detectability of the nest by other brood parasites or predators
Egg ejection
Lower hatchability, lower chick and young survival, lower future reproductive success of parents
Egg losses
Recognition errors, breakage of own eggs, retaliation by the parasite ("mafia" behavior)
Nest abandonment
Lower hatchability, lower chick and young survival, lower future reproductive success of parents
Egg losses
Time and energy for renesting, lower reproductive success with time of breeding
Chick discrimination
Lower chick and young survival, lower future reproductive success of parents
Egg losses, lower hatchability
Recognition errors
Young discrimination
Lower young survival, lower future reproductive success of parents

 DownLoad:
DownLoad:






 Email Alerts
Email Alerts RSS Feeds
RSS Feeds