
Figure
1.
Location of Borakalalo National Park in South Africa
| Citation: |
Xin Yu, Ping Fan, Yun Wu, Yongbin Chang, Chenxi Jia, Fumin Lei. 2022: GPS tracking data reveal the annual spatiotemporal movement patterns of Bridled Terns. Avian Research, 13(1): 100065. DOI: 10.1016/j.avrs.2022.100065

|
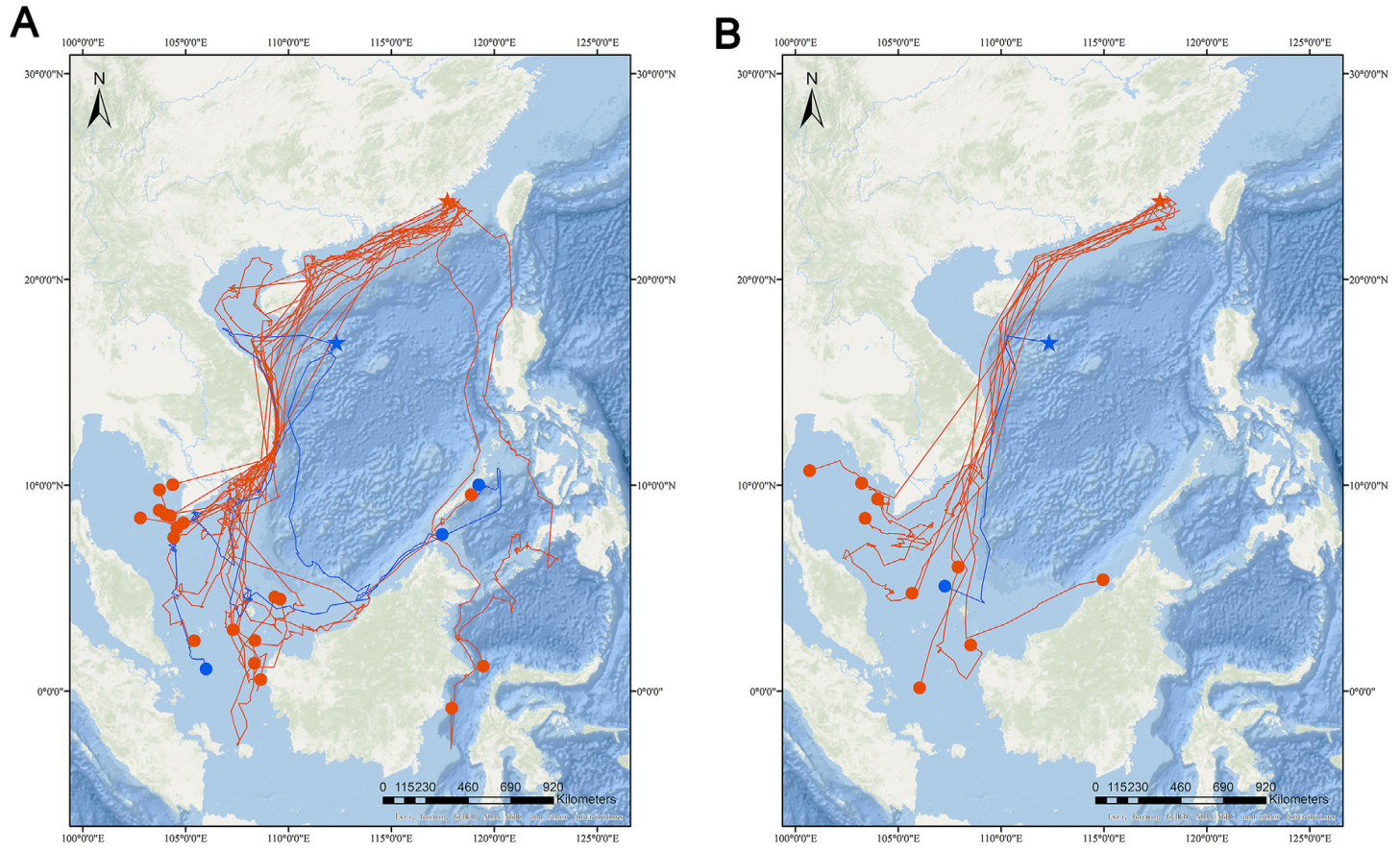
Understanding the spatiotemporal patterns of animal movement is a central theme in the growing field of movement ecology. The Bridled Tern (Onychoprion anaethetus) is widely distributed across tropical and subtropical latitudes; however, knowledge of its annual movement is based only on relatively scarce recoveries of ringed birds. Studying the annual movement of pelagic seabirds is important to identify the impact of changes of marine habitats on them. We examined fine-scaled movement patterns of the species from two colonies in southern China by using GPS-GSM transmitters in 2018–2022. Twenty-three terns bred in Xichiyu Islet and Qilianyu Islet while wintered in four different sites: Gulf of Thailand, Natuna Islands, The Sulu Sea, and Makassar Strait. Bridled Terns made small detours and employed a fly-and-forage strategy with frequent stopovers to forage during autumn migration, but took more direct routes and reduced the need for stopovers during spring migration. Distance of migration between breeding and winter sites was significantly longer in autumn (average 3635 km) when compared with spring (2777 km). Ten birds with whole-year tracking data used the same breeding sites both years and four birds with tracking data of two consecutive years returned to the same wintering area, indicating that Bridled Terns are highly faithful to their breeding and wintering grounds.
The Double-banded Sandgrouse (Pterocles bicinctus) has been hunted in the North West province of South Africa for about the last 170 years (Haagner, 1914; Delegorgue, 1997). They are still hunted today but not at the same commercial level as Burchell's P. burchelli or Namaqua P. namaqua Sandgrouse (Viljoen, 2005; Little and Crowe, 2011). Despite its sporting potential, this species has received very little scientific scrutiny and therefore very little attempt was made to assess it as a sustainable game bird (Little and Crowe, 2011).
I have visited six farms in the North West province that fall within the distribution range of Double-banded Sandgrouse on a regular basis during the past 20 years. In all the cases, group sizes were generally small (2–4 birds). The birds were sparsely distributed compared to the areas where Namaqua and Burchell's Sandgrouse occur and their visits to these farms were at irregular times of the year. Particularly, on the farms Zandfontein, about 50 km northwest of Johannesburg and La Boheme, 40 km north of Rustenburg, Double-banded Sandgrouse have been observed for 15 and six years respectively at a waterhole, with the largest group size of six, but usually more in the order of twos and fours and their visits were unpredictable in terms of annual arrival times (H. Bronkhorst personal communication and J.H. van Niekerk personal observations). Tarboton et al. (1987) described Double-banded Sandgrouse as rare vagrants in this area.
In view of the limitations of small groups and erratic movement patterns in the North West province, an effort was made to observe Double-banded Sandgrouse in Borakalalo National Park, where their visits appeared more consistent during 10 years of casual observations and where Double-banded Sandgrouse were observed in twos and fours about 100 years ago (Haagner, 1914). The main purpose of this study is to describe the intrinsic (breeding) and extrinsic factors (rainfall/vegetation) that may be responsible for the movements of Double-banded Sandgrouse. This is required to gather baseline data in order to conserve the species since it is hunted.
Borakalalo National Park (14000 ha) (25°09.262 S, 27°48.522 E) is situated 80 km north of Pretoria and 30 km north of Jericho in the North West province. Its vegetation is described as Western Sandy Bushveld including tree species such as Acacia erubescens, Combretum apiculatum, Terminalia sericea and Burkea africana (Mucina and Rutherford, 2006). Figure 1 shows the location of the park in South Africa and Fig. 2 the focal study area, which is the southern-most portion of Borakalalo National Park from the southern shoreline of the Klipvoor Dam further south. The study site was divided into a 'hinterland' section that denotes sandgrouse habitat 400–5000 m south from the shoreline of the dam and a 'dam area' where sandgrouse were observed 50–400 m from its shoreline. The veld was burnt by park officials in July 2010 (Fig. 2). Various large herbivores in herding groups occur in the park including Impala (Aepyceros melampus), White Rhinocerous (Ceratotherium simum), Kudu (Tragelaphus strepsiceros), Zebra (Equus quagga), Blue Wildebeest (Connochaetes taurinus) and Sable Antelope (Hippotragus niger).
Data collection was based on traversing the park for two days per month during 2010 along a set route (32 km) with a vehicle at 10 km·h−1. Traversing was done in the mornings from 06:30–09:30 and in the afternoons from 15:30–18:30 and on the following day another morning session, totalling 108 hours (1152 km) during 2010. In the focal study area of the park about two hours were spent in the hinterland and one hour near the dam during each trip (Fig. 2). All sightings of sandgrouse were recorded on a Garmin Colorado 300 GPS and downloaded onto a computer with MapSource. MapSource was used to determine the direct distance from a sandgrouse group to the nearest watering point, i.e., the edge of the Klipvoor Dam. For the purpose of calculating group sizes, sandgrouse recorded outside of the focal study area (Fig. 2) of the park, were also included. Upon encountering sandgrouse, group size, sex and age were recorded with the aid of binoculars. Males possess a prominent transverse black and white band across the forehead and young birds were smaller and possessed female features (Little and Crowe, 2011). At each sighting the distance was also estimated between the male and female of a pair and where applicable, the distance between two pairs. The substrate beneath every sandgrouse sighted was also noted and recorded as 1) open road, 2) short trampled grass or 3) burnt veld. The height of the observer in the vehicle above ground level during traversing was 160–180 cm. This position allowed the observer to search for sandgrouse in relatively thick grass up to 20 m away from the vehicle on either side of the road. Rainfall data was provided by the North West Parks and Tourism Board (Jericho). Statistical analysis included One-way ANOVA, Pearson correlation coefficients and chi-squares.
The mean group size of Double-banded Sandgrouse was 2.47 (n = 38, range = 1–5 and SD = 1.22). The largest group was recorded in March which was 3.7 birds (n = 7), mainly consisting of adults. Out of 36 groups recorded, 69.4% (25 groups) were pairs, 16.6% (6 groups) were family groups and 13.8% (5 groups) were single birds. Two groups were not identified. Family groups were observed during March–August 2010 and each family had only one offspring. Double-banded Sandgrouse offspring remained with parents but it is not clear at which age offspring left the natal group. The single birds might have been young birds that left the natal group. Out of 38 groups 25% were groups of four birds, always consisting of two pairs each. Pairs were males and females that foraged or sat at a mean distance of 1.93 m (SD = 196.3, n = 16, range = 0–5 m) apart while the mean distance between pairs in a group was 5.5 m (SD = 206.5, n = 7 and range = 0.2–8 m) (ANOVA: F = 15.605, df = 21, p < 0.05). The short distances between a male and female shows that they formed monogamous bonds throughout the study period. The male to female ratio in the population was 1:0.87 (males = 48, females = 42). This ratio remained more or less constant throughout all months during 2010 revealing a significant positive correlation between numbers of males and females in the population (r = 0.97, df = 11, p < 0.05) (Fig. 3). Thus, groups with one sex (e.g. males only) did not dominate in the population and neither were any groups recorded that only contained offspring.
Figure 4 shows that Double-banded Sandgrouse were mainly present in the park in two relatively dry periods, from February–March and again from July–September 2010. In other words, their movement was affected by short wet/dry spells. Their absence during the wet summer period was generally also the pattern during the previous 10 years. However, a few birds were observed in a Teminalia sericea bush in the park during December 2003, which means they are not necessarily entirely absent during wet conditions.
Table 1 and Fig. 2 shows that Double-banded Sandgrouse were more regularly observed in the hinterland in the mornings and conversely, during the late afternoons they were more regularly observed closer to water (χ2 = 7.22, df = 3, p < 0.05). The mean distance of sandgrouse to the edge of the water in the morning was 1979.1 m (SD = 1265.9, n = 11, range = 50–3500 m) and in the late afternoons 334.06 m (SD = 304.68, n = 18, range = 50–800 m) (ANOVA: F = 12.19, df = 26, p < 0.05). From here they moved closer to drink water after sunset (also see Hockey et al., 2005).
| Morning | Afternoon | ||||
| Hinterland | Dam area | Hinterland | Dam area | ||
| Hours traversing | 48 | 24 | 24 | 12 | |
| Number of sandgrouse | 16 | 4 | 2 | 39 | |
| Number of sandgrouse per hour | 0.3 | 0.16 | 0.08 | 3.25 | |
 DownLoad:
CSV
DownLoad:
CSV
Overall, Double-banded Sandgrouse were recorded in areas where open basal grass cover conditions prevailed. Table 2 shows that they were observed on the road, on short trampled/grazed grass or in burnt velds with green sprouts in a thornveld (Acacia spp. and grass). In fact, just after burning from July–September, Double-banded Sandgrouse were often recorded in burnt velds (Table 2). In previous years they have also been observed in burnt areas in other parts of the park on three occasions but also under the canopy of bushes with high trees such as Terminalia sericea and Burkea africana, where the grass and shrub cover under these trees were sparse (J.H. van Niekerk, unpublished).
| Road | Short trampled grass | Burnt grass | Chi-squares | |
| Overall | 50 (59%) | 9 (10.70%) | 25 (29.70%) | χ2 = 133.23, df = 3, p < 0.001 |
| During July–September | 15 (35.70%) | 2 (4.76%) | 25 (59.52%) | χ2 = 62.1, df = 3, p < 0.001 |
DownLoad:
CSV
The small group size of 1–5 birds that was recorded in Borakalalo National Park is similar to the small group sizes of twos and fours that were recorded in this area more than 100 years ago (Haagner, 1914). This is also consistent with the small groups generally observed in many parts of the North West province during the past 20 years. Reports of this species towards the east of South Africa, in the lowveld of Mpumalanga province and in Kruger National Park, show that they gather in larger groups of 30–50 birds to drink water (Horsbrugh, 1912; Hockey et al., 2005). Payne (1968) also reported that Double-banded Sandgrouse were recorded in twos and fours in scrubby mopane woodlands but flew in groups of dozens to water at dusk in the Hans Merensky Nature Reserve in the lowveld.
Similar to Burchell's Sandgrouse in the Molopo Game Reserve, the Double-banded Sandgrouse also sits in open roads in small groups that are closely knitted, but in Burchell's Sandgrouse these road sitters were generally in groups of 4–8 during winter and in pairs during summer, while in Double-banded Sandgrouse these groups varied between 1–5 (J.H. van Niekerk, unpublished; Little and Crowe, 2011). Unlike Burchell's and the Yellow-throated Sandgrouse (P. gutturalis) in the North West province, Double-banded Sandgrouse in Borakalalo National Park did not undertake flights in groups of 20–30 birds to watering points in the late mornings, but instead, flew closer to water in smaller groups during late afternoons and when it was dark they moved for a second time to the edge of the water (J.H. van Niekerk, unpublished). Indeed, Double-banded Sandgrouse formed larger drinking parties after sunset but in this park it is certainly less than 10 (also see Hockey et al., 2005).
This preliminary survey suggests that five factors could trigger the movement of Double-banded Sandgrouse:
1) Double-banded Sandgrouse were absent or fewer in the park during wet summer conditions; this corresponds with lower reporting rates for this species in this area during the wet season (Maclean and Herremans, 1997). This suggests that Double-banded Sandgrouse disperse to more watering points.
2) Their presence/absence in the reserve was also affected by short spells of high/low rainfall periods and not only changing seasons (Fig. 4).
3) They showed a propensity towards short trampled grass as well as recently burnt grass with green sprouts in thornvelds. Double-banded Sandgrouse were suddenly observed back in the park again just after veld burning in July 2010 and remained there for a few months (Fig. 3). Their preference for burnt grass areas was also observed in Kruger National Park (A. Kemp personal communication) which could be a matter of food availability. Their occurrence in relatively open substrates in this survey is coherent with descriptions of their habitat preferences, presented in other publications, such as in tussocky grass and on gravel plains as opposed to dense bush or thick grass (Little and Crowe, 2011). Also, at La Boheme and Magaliesberg, where searches were carried out on foot, they were not observed in thick grassy areas but on open patches (rocky outcrops) or on roads.
4) Double-banded Sandgrouse was observed in the park when they had offspring during late summer and winter, which suggests that they bred outside the park. However, although no signs of breeding was observed in the park during 2010, parents with chicks were certainly observed in April in 2007 in the hinterland of the park (n = 3, J.H. van Niekerk personal observations) which suggests that there is not necessarily a cyclic movement between breeding and foraging areas. On the other hand, where Double-banded Sandgrouse visited a farm near Magaliesberg for the last 15 years, no breeding signs were observed.
5) Finally, daily water intake certainly causes movements over a distance of at least 4 km.
Movements were not affected by demographic pressures such as juvenile flocks being rejected to move away from parents or batchelor flocks moving around while breeding takes place. The birds remained in monogamous bonds or family groups throughout. Medium grazing pressure, regular cattle/game watering points and some veld burning during late winter may favor the Double-banded Sandgrouse and are aspects that can be managed in favor of grazing and for the benefit of sandgrouse by landowners.
|
Dunlop, J.N., Rippey, E., 2006. The natural history of the bridled tern on Penguin Island, Western Australia. Naragebup-Rockingham Regional Environmental Centre, Perth.
|
|
Gochfeld, M., Burger, J., 1996. Family Sternidae (terns). In: Hoyo, J.D., Elliott, A., Sargatal, J. (Eds.), Handbook of the Birds of the World. Hoatzin to Auks. Lynx Edicions, Barcelon, pp. 624–667.
|
|
Howell, S.N., Webb, S., 1995. A Guide to the Birds of Mexico and Northern Central America. Oxford University Press, Oxford.
|
|
Mallory, M.L., Gilbert, C., 2008. Leg-loop harness design for attaching external transmitters to seabirds. Mar. Ornithol. 36, 183-188.
|
|
Perera, L., Ilangakoon, A., 2016. Results of the first systematic boat-based seabird survey in Sri Lanka. Forktail 32, 58-61.
|
Figures(4)





| Morning | Afternoon | ||||
| Hinterland | Dam area | Hinterland | Dam area | ||
| Hours traversing | 48 | 24 | 24 | 12 | |
| Number of sandgrouse | 16 | 4 | 2 | 39 | |
| Number of sandgrouse per hour | 0.3 | 0.16 | 0.08 | 3.25 | |
DownLoad:
CSV
| Road | Short trampled grass | Burnt grass | Chi-squares | |
| Overall | 50 (59%) | 9 (10.70%) | 25 (29.70%) | χ2 = 133.23, df = 3, p < 0.001 |
| During July–September | 15 (35.70%) | 2 (4.76%) | 25 (59.52%) | χ2 = 62.1, df = 3, p < 0.001 |
DownLoad:
CSV



 Email Alerts
Email Alerts RSS Feeds
RSS Feeds