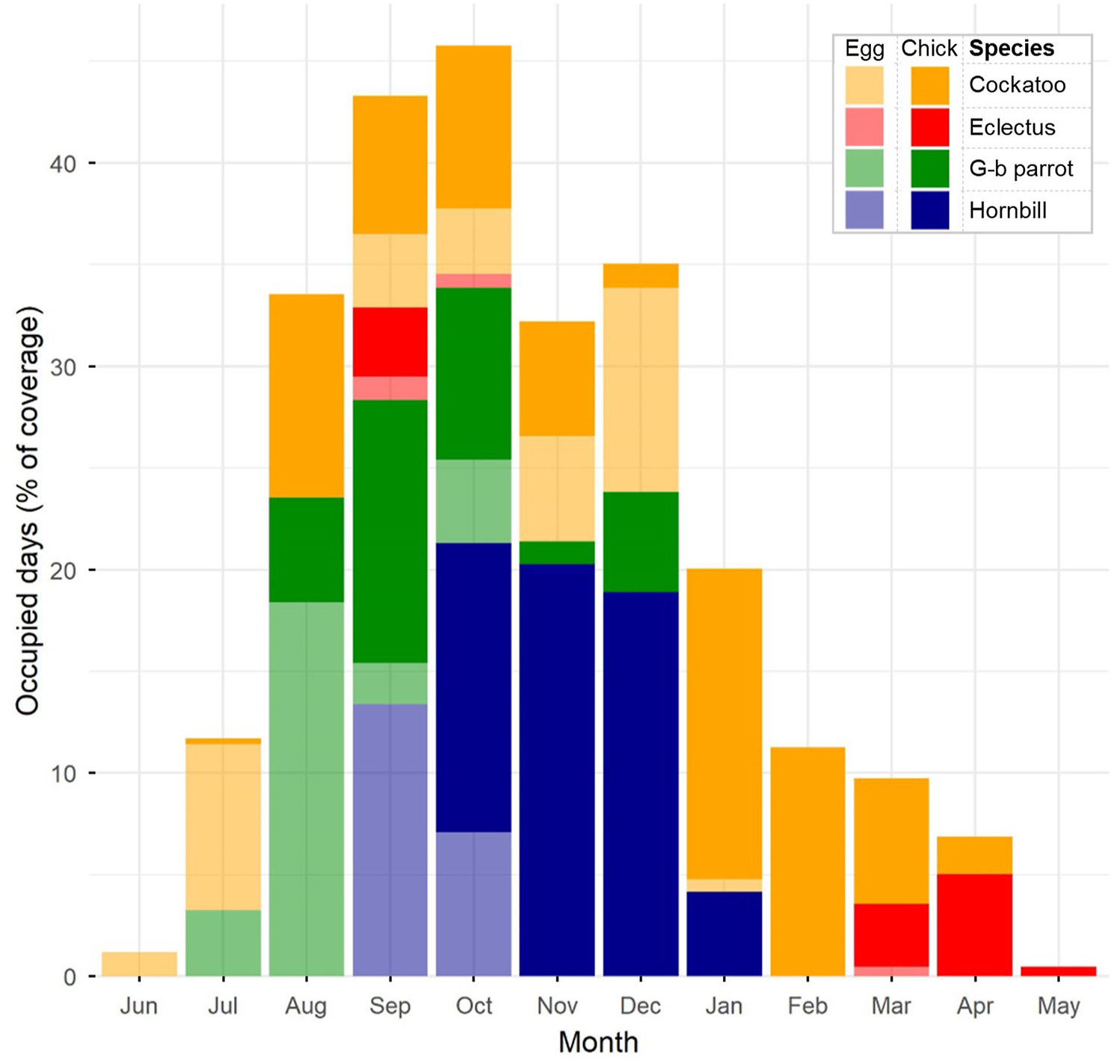
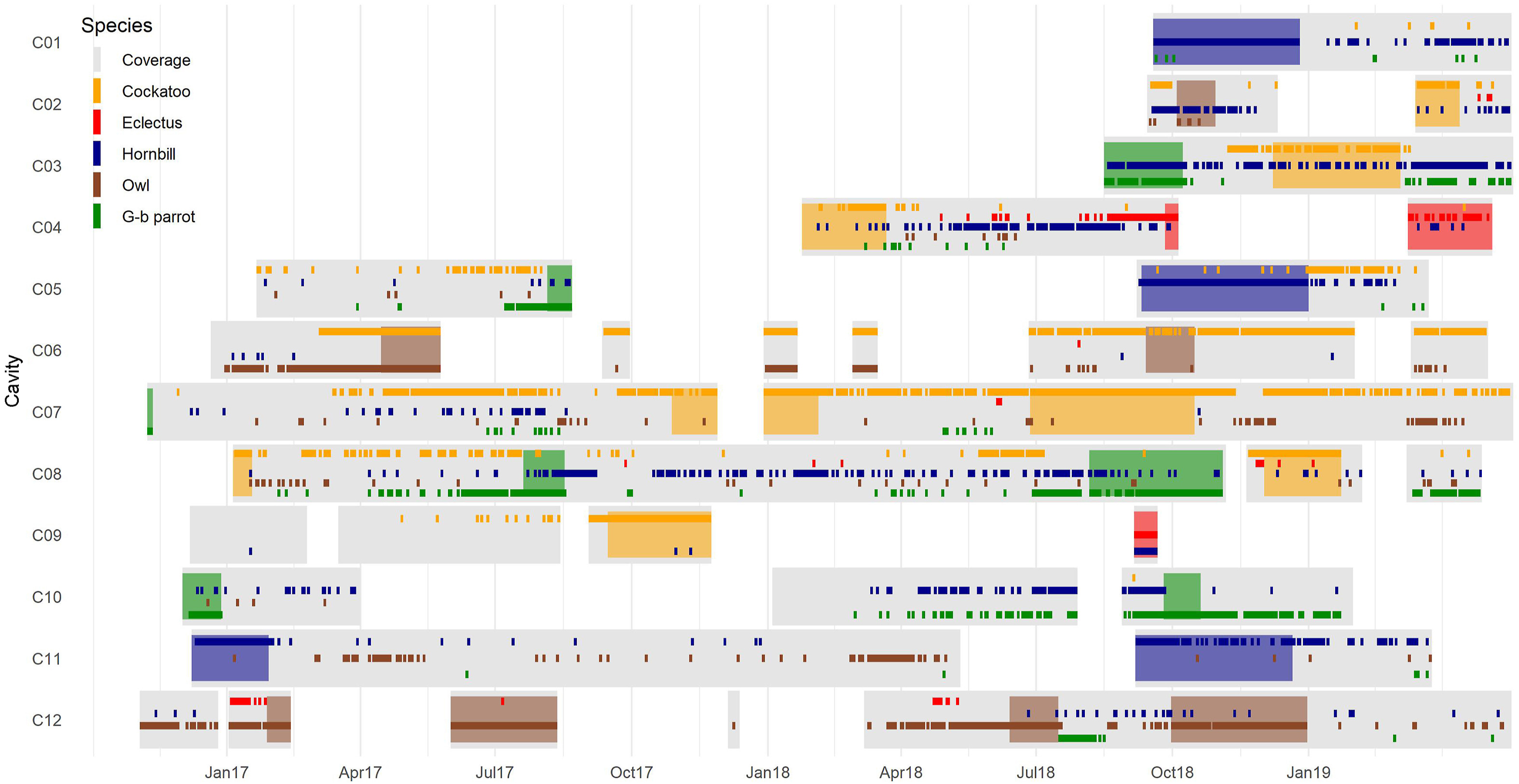
|
Agista, D., Rubyanto, D., 2001. Telaah awal status, penyebaran, dan populasi kakatua kecil jambul kuning, Cacatua sulphurea parvula: di Taman Nasional Komodo, Nusa Tenggara Timur. Birdlife International Indonesia Programme, Bogor, Indonesia.
|
Altamirano, T.A., Ibarra, J.T., Martin, K., Bonacic, C., 2017. The conservation value of tree decay processes as a key driver structuring tree cavity nest webs in South American temperate rainforests. Biodivers. Conserv. 26, 2453-2472
|
|
Arendt, W.J., 2000. Impact of nest predators, competitors, and ectoparasites on Pearly-eyed Thrashers, with comments on the potential implications for Puerto Rican Parrot recovery. Ornitol. Neotrop. 11, 13-63
|
Badan Pusat Statistik, 2016. Statistical Yearbook of Indonesia. . (Accessed 17 October 2017).
|
Badan Pusat Statistik, 2021. Statistical Yearbook of Indonesia. . (Accessed 20 December 2021).
|
|
van Balen, J.H., Booy, C.J.H., van Franeker, J.A., Osieck, E.R., 1982. Studies on hole-nesting birds in natural nest sites. Ardea 70, 1-24
|
|
Bates, D., Machler, M., Bolker, B., Walker, S., 2015. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1-48
|
BirdLife International, 2021. Datazone. . (Accessed 11 March 2021).
|
Bonaparte, E.B., Cockle, K.L., 2017. Nest niche overlap among the endangered Vinaceous-breasted Parrot (Amazona vinacea) and sympatric cavity-using birds, mammals, and social insects in the subtropical Atlantic Forest, Argentina. Condor 119, 58-72
|
Brightsmith, D., Bravo, A., 2006. Ecology and management of nesting Blue-and-Yellow Macaws (Ara ararauna) in Mauritia Palm Swamps. Biodivers. Conserv. 15, 4271-4287
|
Brightsmith, D.J., 2005. Competition, predation and nest niche shifts among tropical cavity nesters: phylogeny and natural history evolution of parrots (Psittaciformes) and trogons (Trogoniformes). J. Avian Biol. 36, 64-73
|
Britt, C.R., Garcia Anleu, R., Desmond, M.J., 2014. Nest survival of a long-lived psittacid: Scarlet Macaws (Ara macao cyanoptera) in the Maya Biosphere Reserve of Guatemala and Chiquibul Forest of Belize. Condor 116, 265-276
|
Cahill, A.J., Walker, J.S.J.S., Marsden, S.J., 2006. Recovery within a population of the Critically Endangered citron-crested cockatoo Cacatua sulphurea citrinocristata in Indonesia after 10 years of international trade control. Oryx 40, 161-167
|
Chen, J.-N., Liu, N.-F., Yan, C., An, B., 2011. Plasticity in nest site selection of Black Redstart (Phoenicurus ochruros): a response to human disturbance. J Ornithol. 152, 603-608
|
Clout, M.N., Merton, D.V., 1998. Saving the kakapo: the conservation of the world’s most peculiar parrot. Bird. Conserv. Int. 8, 281-296
|
Cockle, K.L., Martin, K., Drever, M.C., 2010. Supply of tree-holes limits nest density of cavity-nesting birds in primary and logged subtropical Atlantic forest. Biol. Conserv. 143, 2851-2857
|
|
Collar, N.J., Eaton, J.A., Sykes, B., 2017. Conservation and the redaction of locality data. BirdingASIA 28, 3-4
|
|
Collar, N.J, Marsden, S.J., 2014. The subspecies of Yellow-crested Cockatoo Cacatua sulphurea. Forktail 23, 7
|
Collias, N.E., 1964. The evolution of nests and nest-building in birds. Am. Zool. 4, 175-190
|
Danchin, E., Wagner, R.H., 1997. The evolution of coloniality: the emergence of new perspectives. Trends Ecol. Evol. 12, 342-347
|
|
Delany, M.E., Tell, L.A., Millam, J.R., Preisler, D.M., 1999. Photographic candling analysis of the embryonic development of orange-winged amazon parrots (Amazona amazonica). J. Avian Med. Surg. 13, 116-123
|
|
Djawarai, Y., Bashari, H., Siregar, B., Widyanto, A., 2014. The breeding ecology of Sumba Cockatoo Cacatua Sulphurea Citrinocristata in Manupeu Tanadaru National Park and surrounding, Sumba. Burung Indonesia. Bogor, Indonesia.
|
|
Eaton, J.A., van Balen, S.B., Brickle, N.W., Rheindt, F.E., 2016. Birds of the Indonesian Archipelago Greater Sundas and Wallacea. Lynx Ediciones, Barcelona.
|
|
Eggers, S., Griesser, M., Nystrand, M., Ekman, J., 2005. Predation risk induces changes in nest-site selection and clutch size in the Siberian jay. Proc. R. Soc. B. Biol. Sci. 273, 701-706
|
Gonzalez, J.A., 2003. Harvesting, local trade, and conservation of parrots in the Northeastern Peruvian Amazon. Biol. Conserv. 114, 437-446
|
Gutzat. F., Dormann, C.F., 2018. Decaying trees improve nesting opportunities for cavity-nesting birds in temperate and boreal forests: A meta-analysis and implications for retention forestry. Ecol. Evol. 8, 8616-8626
|
Harper, G.A., Bunbury, N., 2015. Invasive rats on tropical islands: Their population biology and impacts on native species. Glob. Ecol. Conserv. 3, 607-627
|
Heinsohn, R., Legge, S., 2003. Breeding biology of the reverse-dichromatic, co-operative parrot Eclectus roratus. J. Zool. 259, 197-208
|
Heinsohn, R., Murphy, S., Legge, S., 2003. Overlap and competition for nest holes among eclectus parrots, palm cockatoos and sulphur-crested cockatoos. Aust. J. Zool. 51, 81-94
|
Heinsohn, R., Zeriga, T., Murphy, S., Igag, P., Legge, S., Mack, A.L., 2009. Do Palm Cockatoos (Probosciger aterrimus) have long enough lifespans to support their low reproductive success? Emu 109, 183-191
|
|
Heinsohn, T.E., 2001. Human influences on vertebrate zoogeography: animal translocation and biological invasions across and to the east of Wallace’s Line. In: Metcalfe, I., Smith, J.M.B., Morwood, M., Davidson, I. (Eds.), Faunal and Floral Migrations and Evolution in SE Asia-Australasia. CRC Press, Boca Raton, US, pp. 153–170.
|
Heinsohn, T.E., 2003. Animal translocation: long-term human influences on the vertebrate zoogeography of Australasia (natural dispersal versus ethnophoresy). Zoologist 32, 351-376
|
|
Hidayat, O., 2012. Lubang masa depan. Biodivers. Indon. 2, 110-118
|
Hidayat, O., Kayat, K., 2020. Pendekatan preferensi habitat dalam penyusunan strategi konservasi in situ Kakatua Sumba (Cacatua sulphurea citrinocristata, Fraser 1844). J. Penelit. Hutan dan Konserv. Alam.;17, 113-126
|
van der Hoek, Y., Gaona, G.V., Martin, K., 2017. The diversity, distribution and conservation status of the tree-cavity-nesting birds of the world. Divers. Distrib. 23,1120-1131
|
Howald, G., Donlan, C.J., Galvan, J.P., Russell, J.C., Parkes, J., Samaniego, A., et al., 2007. Invasive rodent eradication on islands. Conserv. Biol. 21, 1258-1268
|
Igag, P., Mack, A.L., Legge, S., Heinsohn, R., 2019. Breeding biology of three large, sympatric rainforest parrots in New Guinea: Palm Cockatoo, Pesquet’s Parrot and Eclectus Parrot. Emu 119, 196-204
|
|
Ihsannudin, I., Hidayat, K., Sukesi, K., Yuliati, Y., 2020. Perception of the local community toward the yellow-crested cockatoo (Cacatua sulphurea abbotti), a critically endangered species in Masakambing Island, Indonesia. Geografia 16, 1-14
|
|
Imansyah, M.J., Purwandana, D., Ariefiandy, A., Benu, Y.J., Jessop, T.S., Trainor, C.R., 2016. Valley-floor censuses of the Critically Endangered Yellow-crested Cockatoo Cacatua sulphurea occidentalis on Komodo Island, East Nusa Tenggara province, Indonesia, point to a steep population decline over a six-year period. Forktail 32, 66-72
|
Indraswari, K., Friedman, R.S., Noske, R., Shepherd, C.R., Biggs, D., Susilawati C., et al., 2020. It’s in the news: Characterising Indonesia’s wild bird trade network from media-reported seizure incidents. Biol. Conserv. 243, 108431
|
Jones, C.G., 1980. Parrot on the Way to Extinction. Oryx 15, 350-354
|
|
Jones, C.G., 1987. The larger native landbirds of Mauritius. In: Diamon, A.W. (Ed.), Studies of Mascarene Island Birds. Cambridge University Press, Cambridge, UK, pp. 271–300.
|
|
Jones, C.G., Malham, J., Reuleaux, A., Richards, H.A., Raisin, C., Tollington, S., et al., 2013. Echo parakeet Psittacula eques perruche de Maurice. In: Safford, R., Hawkins, F. (Eds.), The Birds of Africa. Vol. VIII. Birds of the Malagasy Region. A&C Black, Madagascar, pp. 433–438.
|
Jones, M.J., Linsley, M.D., Marsden, S.J., 1995. Population sizes, status and habitat associations of the restricted-range bird species of Sumba, Indonesia. Bird Conserv. Int. 5, 21-52
|
Kaisin, O., Gazagne, E., Savini, T., Huynen, M.-C., Brotcorne, F., 2018. Foraging strategies underlying bird egg predation by macaques: A study using artificial nests. Am. J. Primatol. 80, e22916
|
Karande, A.A., 1967. Timber, Tetrameles nudiflora R., resistant to teredid borers in Bombay Harbour. Nature 213, 105
|
Koenig, S.E., Wunderle, J.M., Enkerlin-Hoeflich, E.C., 2007. Vines and canopy contact: A route for snake predation on parrot nests. Bird Conserv. Int. 17, 79-91
|
|
Lack, D., 1968. Ecological Adaptations for Breeding in Birds. Methuen & Co Ltd, London, UK.
|
Linhart, P., Fuchs, R., Polakova, S., Slabbekoorn, H., 2012. Once bitten twice shy: long-term behavioural changes caused by trapping experience in willow warblers Phylloscopus trochilus. J. Avian Biol. 43, 186-192
|
|
Loong, S., Sin, Y.C.K., Johns, P., Plowden, T., Yong, D.L., Lee, J., et al., 2021. Nest predation by Oriental Pied Hornbills Anthracoceros albirostris in urban Singapore. BirdingASIA 35, 86-91
|
Marini, M.A., Melo, C., 1998. Predators of quail eggs, and the evidence of the remains: Implications for nest predation studies. Condor 100, 395-399
|
|
Marsden, S.J., 1995. The Ecology and Conservation of the Parrots of Sumba, Buru and Seram, Indonesia. Ph.D thesis. Manchester Metropolitan University, Manchester, UK.
|
Marsden, S.J., Jones, M.J., 1997. The nesting requirements of the parrots and hornbill of Sumba, Indonesia. Biol. Conserv. 82, 279-287
|
Martin, R.O., Perrin, M.R., Boyes, R.S., Abebe, Y.D., Annorbah, N.D., Asamoah, A., et al., 2014. Research and conservation of the larger parrots of Africa and Madagascar: a review of knowledge gaps and opportunities. Ostrich 85, 205-233
|
Masello, J.F., Pagnossin, M.L., Sommer, C., Quillfeldt, P., 2006. Population size, provisioning frequency, flock size and foraging range at the largest known colony of Psittaciformes: The Burrowing Parrots of the north-eastern Patagonian coastal cliffs. Emu 106, 69-79
|
Masello, J.F., Quillfeldt, P., 2002. Chick growth and breeding success of the Burrowing Parrot. Condor 104, 574-586
|
Matuzak, G.D., Brightsmith, D.J., 2007. Roosting of Yellow-naped Parrots in Costa Rica: estimating the size and recruitment of threatened populations. J. Field Ornithol. 78, 159-169
|
|
Monk, K., De Fretes, Y., Reksodiharjo-Lilley, G., 1997. Ecology of Nusa Tenggara and Maluku. Oxford University Press, Oxford, UK.
|
Monterrubio, T., Enkerlin-Hoeflich, E., Hamilton, R.B., 2002. Productivity and nesting success of Thick-billed Parrots. Condor 104, 788-794
|
Moorhouse, R., Greene, T., Dilks, P., Powlesland, R., Moran, L., Taylor, G., et al., 2003. Control of introduced mammalian predators improves kaka Nestor meridionalis breeding success: reversing the decline of a threatened New Zealand parrot. Biol. Conserv. 110, 33-44
|
Murphy, S., Legge, S., Heinsohn, R., 2003. The breeding biology of palm cockatoos (Probosciger aterrimus): A case of a slow life history. J. Zool. 261, 327-339
|
|
Nandika, D., Agustina, D., 2012. Few and far between. Saving the Yellow-crested Cockatoos. PsittaScene 24, 130-134
|
|
Nandika, D., Mulyani, Y.A., Prawiradilaga, D.M., Agustina, D., 2020. Monitoring of Cacatua sulphurea abbotti in Masakambing Island, Indonesia. Biotropia Southeast Asian J. Trop. Biol. 27, 271-281
|
Newton, I., 1994. The role of nest sites in limiting the numbers of hole-nesting birds: A review. Biol. Conserv. 70, 265-276
|
|
Nilsson, S.G., 1986. Evolution of hole-nesting in birds: on balancing selection pressures. Auk 103, 432-435
|
Olah, G., Butchart, S.H.M., Symes, A., Guzman, I.M., Cunningham, R., Brightsmith, D.J., et al., 2016. Ecological and socio-economic factors affecting extinction risk in parrots. Biodivers. Conserv. 25, 205-223
|
|
Persulessy, Y.E., Djawarai, Y.B., Marut, R., 2003. Survei populasi dan distribusi kakatuakecil jambul-kuning Cacatua sulphurea citrinocristata dan empat jenis paruh bengkok lain di Pulau Sumba. BirdLife Indonesia/ZGAP, Bogor, Indonesia.
|
|
PHPA, L.I.P.I., BirdLife International-IP, 1998. Species Recovery Plan Yellow-Crested Cockatoo. PHPA/LIPI/BirdLife International-Indonesia Programme, Bogor,
Indonesia.
|
Pierce, A.J., Pobprasert, K., 2013. Nest predators of southeast Asian evergreen forest birds identified through continuous video recording. Ibis 155, 419-423
|
Pires, S.F., 2012. The illegal parrot trade: a literature review. Glob. Crime. 13, 176-190
|
Pires, S.F., Olah, G., Nandika, D., Agustina, D., Heinsohn, R., 2021. What drives the illegal parrot trade? Applying a criminological model to market and seizure data in Indonesia. Biol. Conserv. 257, 109098
|
|
R Core Team, 2021. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria.
|
Renton, K., 2004. Agonistic interactions of nesting and nonbreeding macaws. Condor 106, 354-362
|
Reuleaux, A., Richards, H., Payet, T., Villard, P., Waltert, M., Bunbury, N., 2014. Breeding ecology of the Seychelles Black Parrot Coracopsis barklyi. Ostrich 85, 255-265
|
Reuleaux, A., Siregar, B.A., Collar, N.J., Panggur, M.R., Mardiastuti, A., Jones, M.J., et al., 2020. Protected by dragons: Density surface modeling confirms large population of the critically endangered Yellow-crested Cockatoo on Komodo Island. Condor 122, duaa042
|
|
Ribeiro-Silva, L., Perrella, D.F., Biagolini, Jr., C.H., Zima, P.V.Q., Piratelli, A.J., Schlindwein, M.N., et al., 2018. Testing camera traps as a potential tool for detecting nest predation of birds in a tropical rainforest environment. Zoologia 35, 1-8
|
Richardson, T.W., Gardali, T., Jenkins, S.H., 2009. Review and meta-analysis of camera effects on avian nest success. J. Wildl. Manage. 73, 287-293
|
|
Rocamora, G., Laboudallon, V., 2013. Seychelles black parrot Coracopsis barklyi. In: Safford, R., Hawkins, F. (Eds.), The Birds of Africa. Vol. VIII. Birds of the Malagasy Region. Madagascar, Seychelles, Comoros, Mascarenes. A&C Black, Madagascar, pp. 445–447.
|
Rolland, C., Danchin, E., de Fraipont, M., 1998. The evolution of coloniality in birds in relation to food, habitat, predation, and life-history traits: A comparative analysis. Am. Nat. 151, 514-529
|
|
Rowley, I., 1990. Behavioural Ecology of the Galah, Eolophus roseicapillus. Surrey Beatty & Sons, New South Wales, Australia.
|
Rowley, I., Chapman, G., 1991. The breeding biology, food, social organisation, demography and conservation of the Major Mitchell or Pink Cockatoo, Cacatua leadbeateri, on the margin of the Western Australian Wheatbelt. Aust. J. Zool. 39, 131-142
|
Sanchez-Mercado, A., Ferrer-Paris, J.R., Rodriguez, J.P., Tella, J.L., 2021. A literature synthesis of actions to tackle illegal parrot trade. Diversity 13, 191
|
Sanders, M.D., Maloney, R.F., 2002. Causes of mortality at nests of ground-nesting birds in the Upper Waitaki Basin, South Island, New Zealand: a 5-year video study. Biol. Conserv. 106, 225-236
|
Saunders, D.A., Dawson, R., Mawson, P.R., Cunningham, R.B., 2020. Artificial hollows provide an effective short-term solution to the loss of natural nesting hollows for Carnaby’s Cockatoo Calyptorhynchus latirostris. Biol. Conserv. 245, 108556
|
Saunders, D.A., Smith, G.T., Rowley, I., 1982. The availability and dimensions of tree hollows that provide nest sites for cockatoos (Psittaciformes) in Western Australia. Wildl. Res. 9, 541-556
|
|
Schaaf, A.A., Garcia, C.G., Ruggera, R.A., Tallei, E., Vivanco, C.G., Rivera, L., et al., 2021. Influence of logging on nest density and nesting microsites of cavity-nesting birds in the subtropical forests of the Andes. Forestry 95, 73-82
|
|
Schaaf, A.A., Ruggera, R.A., Vivanco, C.G., Tallei, E., Benavidez, A., Albanesi, S., et al., 2020. Tree use, niche breadth and overlap for excavation by woodpeckers in subtropical piedmont forests of northwestern Argentina. Acta Ornithol. 55, 111-119
|
|
Setiawan, I., 1996. The Status of Cacatua Sulphurea Parvula in Nusa Penida, Bali, and Sumbawa. PHPA/BirdLife International, Bogor, Indonesia.
|
Shoemaker, K.T., Breisch, A.R., Jaycox, J.W., Gibbs, J.P., 2013. Reexamining the minimum viable population concept for long-lived species. Conserv. Biol. 27, 542-551
|
|
Snyder, N.F.R., McGowan, P., Gilardi, J., Grajal, A., 2000. Parrots. Status Survey and Conservation Action Plan 2000-2004. IUCN, Oxford, UK.
|
|
Spoon, T.R., 2006. Parrot reproductive behavior, or who associates, who mates, and who cares. In: Luescher, A. (Ed.), Manual of Parrot Behavior. Blackwell Publishing Ltd, Ames, Iowa US, pp. 63–78.
|
|
Stattersfield, A.J., Crosby, M.J., Long, A.J., Wege, D.C., 1998. Endemic bird areas of the world: priorities for biodiversity conservation. BirdLife Conservation Series No. 7, BirdLife International
|
|
Stojanovic, D., Alves, F., Cook, H., Crates, R., Heinsohn, R., Peters, A., et al., 2017. Further knowledge and urgent action required to save Orange-bellied Parrots from extinction. Emu 118, 126-134
|
|
Tatayah, R.V.V., Malham, J., Haverson, P., Reuleaux, A., Van de Wetering, J., 2007. Design and provision of nest boxes for echo parakeets Psittacula eques in Black River Gorges National Park, Mauritius. Conserv. Evid. 4, 16-19
|
|
Trainor, C.R., Verbelen, P., Johnstone, R.E., 2012. The avifauna of Alor and Pantar, Lesser Sundas, Indonesia. Forktail 28, 77-92
|
Valle, S., Collar, N.J., Harris, W.E., Marsden, S.J., 2018. Trapping method and quota observance are pivotal to population stability in a harvested parrot. Biol Conserv. 217, 428-436
|
Vigo-Trauco, G., Garcia-Anleu, R., Brightsmith, D.J., 2021. Increasing survival of wild macaw chicks using foster parents and supplemental feeding. Diversity 13, 121
|
|
Walker, J.S., Cahill, A.J., Marsden, S.J., 2001. The nesting ecology of Yellow-crested Cockatoo Cacatua Sulphurea on Sumba and the potential for using artificial nest sites to increase recruitment. Preliminary Report 2001. Manchester Metropolitan University, Manchester, UK.
|
Walker, J.S., Cahill, A.J., Marsden, S.J., 2005. Factors influencing nest-site occupancy and low reproductive output in the Critically Endangered Yellow-crested Cockatoo Cacatua sulphurea on Sumba, Indonesia. Bird Conserv. Int. 15, 347-359
|
White, K.L., Eason, D.K., Jamieson, I.G., Robertson, B.C., 2015. Evidence of inbreeding depression in the critically endangered parrot, the kakapo. Anim. Conserv. 18, 341-347
|
|
White, T.H.J., Brown, G.G., Collazo, J.A., 2006. Artificial cavities and nest site selection by Puerto Rican Parrots: a multiscale assessment. Avian Conserv. Ecol. 1, 5
|
|
Widmann, P., Lacerna-Widmann, I.D., 2008. The cockatoo and the community: ten years of Philippine Cockatoo Conservation Programme. BirdingASIA 10, 23-29
|
|
Wungo, E., 2011. Laporan survei populasi paruh bengkok terancam punah Cacatua sulphurea citrinocristata, Sumba, Nusa Tenggara Timur. Burung Indonesia, Bogor, Indonesia.
|

 DownLoad:
DownLoad:














 Email Alerts
Email Alerts RSS Feeds
RSS Feeds