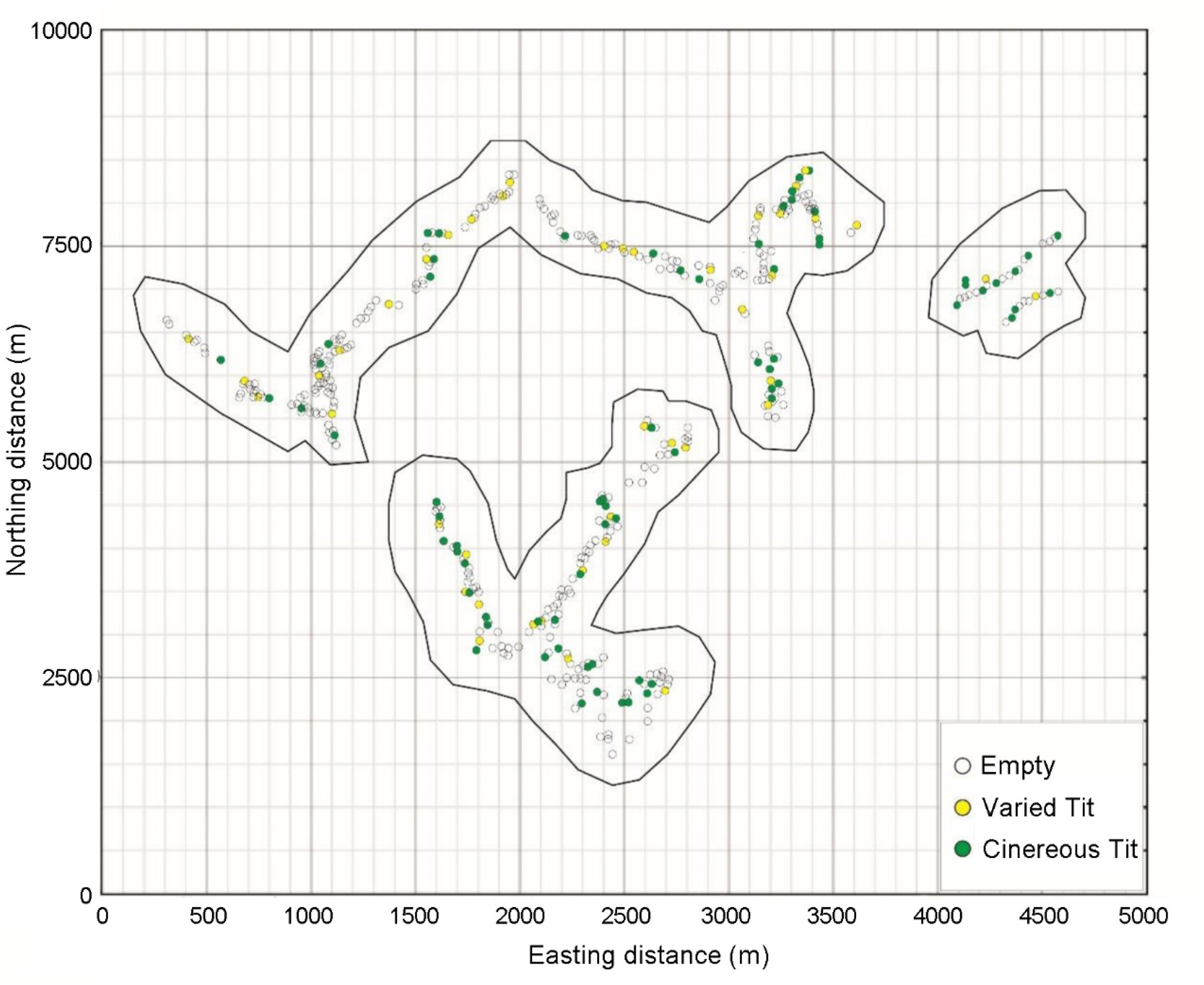
The spatial distribution and breeding behavior of neighboring birds (conspecific and heterospecific) may influence reproduction and the effects differ across species. In this study, we investigated intra- and inter-specific spatial distribution of breeding via artificial nestboxes and examined whether overlap with neighbors affected reproductive success of the focal breeding pair. Regarding spatial distribution, Varied Tits (Sittiparus varius) and Cinereous Tits (Parus cinereus) showed similar pattern: distances of nestboxes containing conspecific tits were greater than those with heterospecific ones. In terms of breeding behavior, reproductive success was not significantly related to the distance from their neighbor’s nest. The reproductive success of Varied Tits was significantly related to that of their neighbors, while in Cinereous Tits, no strong link was found to any recorded reproductive behavior of the neighbors. These findings explain spatial variation in nest site selection of Varied Tits and Cinereous Tits. Reproductive success rate of Varied Tits is affected by that of the neighbors while that of Cinereous Tits is not. The results highlight that there are differences in reproductive outcome among related species and offer suggestions on the use of artificial nestboxes in experiment and conservation programs.
Endemic species have long been a key focus in conservation efforts (Myers et al., 2000), given that the level of endemics might be positively correlated with species richness (Lamoreux et al., 2006). Besides, endemic species with limited dispersal capacity might be sensitive to changes in local climate, or vulnerable to invasive species (Ohlemüller et al., 2008). A good understanding of evolutionary processes such as population subdivisions, changes of effective population size and genetic connectivity of endemic species would shed light on evolutionary processes as well as on conservation management.
The Blue Eared Pheasant (Crossoptilon auritum), belonging to Phasianidae, Galliformes, is a rare and endemic pheasant species in western China (Lei and Lu, 2006). Its wild populations are found at Helan Mountain, as well as along the eastern edge of the Qinghai-Tibetan Plateau (QTP), covering Qinghai, Gansu and Sichuan provinces (Lei and Lu, 2006). Although previous studies have been carried out on the biology and ecology of C. auritum (Sun et al., 2005; Li et al., 2009; Wu and Liu, 2010), a thorough assessment of genetic diversity is urgently needed to assess the population viability of this species. Furthermore, apart from C. auritum, the genus Crossoptilon includes three other species, i.e., C. mantchuricum, C. harmani and C. crossoptilon. These species are endemic to China (Zheng, 2011) and all are listed on the IUCN Red List of Threatened Species (IUCN 2011) because of a rapid decline in the size of their population, caused by habitat fragmentation and hunting (Lei and Lu, 2006). Given these concerns, obtaining molecular markers is a prerequisite in understanding the genetic background of C. auritum and might be useful for population genetic studies in Crossoptilon species.
Microsatellites are powerful tools for conservation genetic studies such as population genetics, mating systems and investigations into kinship (Primmer et al., 2005; Karl et al., 2011). Compared with isolated novel microsatellite markers, cross-species microsatellite amplification from closely related species is cost-effective (Zane et al., 2002). More importantly, it has been suggested that this method has successfully worked among species belonging to the same genera, different genera and even different families (Barbará et al., 2007; Huang and Liao, 2010). Given that numerous microsatellite markers have been developed for various Phasianidae species (Cheng et al., 1995; Wang et al., 2009; Zhou and Zhang, 2009), we attempted to establish microsatellite markers for C. auritum through cross-species amplification from a large number of marker candidates. In order to test the effectiveness of these markers, we also carried out preliminary parentage analysis among captive individuals of known pedigree.
Materials and methods
A total of 20 C. auritum blood samples from brachial veins were collected to develop microsatellite loci by cross-species amplification, nine of which were from the Linxia Zoo in Gansu Province and the other 11 from Huzhu in Qinghai Province, China. Additionally, nine individual birds from two families of the Beijing Wildlife Park in Daxing and Beijing Zoo were used to conduct parentage analysis. We have detailed the known pedigree of these two families of birds.
Genomic DNA was extracted using DNA extraction kits (Tian Gen Biotech, Beijing, China). The cross-species microsatellite markers (Table 1) came from various Galliforme species, including 16 loci from Meleagris gallopavo (Burt et al., 2003), 30 from Tragopan temminckii (Zhou and Zhang, 2009), 20 from Syrmaticus reevesii (Wang et al., 2009), 41 from Gallus gallus (Cheng et al., 1995; Dawson et al., 2010), four from C. mantchuricum (Zhao et al., Beijing Normal University, unpublished results) and one from Syrmaticus mikado (S.H. Li, Taiwan Normal University, unpublished results). After filtering out those loci with poor cross-amplification, the polymorphism of the remaining pairs were tested with either 6-FAM or HEX fluorescent dyelabeled on 5′ of a single forward primer. Polymerase chain reaction (PCR) was carried out in a 10 µL reaction system containing 100 ng DNA, 0.25 µL of each primer, 1 µL of a 10 × PCR buffer, 1.5 mM MgCl2, 0.2 mM dNTP mix and 0.75 U Taq polymerase (Takara, Japan). The reaction was denatured at 94℃ for 5 min, followed by 40 cycles at 94℃ for 30 s, a touch-down annealing process from 58–47℃, reducing in steps of 0.5℃ per cycle and another 20 cycles annealing at 47℃ and then 72℃ for 50 s, with a final extension at 72℃ for 5 min. Fragment analysis was conducted on an ABI PRISM 3100 Genetic Analyzer using the GeneMapper software (Applied Biosystems) with ROX-500 as the standard for size. We conducted PCR amplification and fragment analysis at least twice to ensure the accuracy of individual genotypes. Sequencing of selected homozygotes was also conducted in both directions with amplification primers (BGI Bio Tech, Beijing, China) to ensure that the products were genuine microsatellites.
Table
1.
Characterization of 11 microsatellite loci
Locus
Primer sequences (5′–3′)
Accession No.
Repeat motifs
n
NA
Size range (bp)
HO
HE
Species origin
Chromosome
2580
F: TTAACCTATCAGGTCGTTGCG
AL592580
(CA)n
20
8
191–213
1.00
0.80
M. gallopavo
21
R: CAGTGCACATGCAGGCAG
3D2
F: TCTCTGACGTATCGCATCT
FJ221373
(GT)n
19
6
286–304
0.47
0.58
S. reevesii
4
R: ACTTCCCCTGGTAAACT
1H4
F: TGAACAAGTGAGGCGGAGC
/
(TG)n
20
10
127–161
0.65
0.81
S. reevesii
/
R: CTGCACACAGCCCGAAGC
2420
F: CATCATCTGCCAATGCAGAGG
/
(TTTA)n
20
4
118–142
0.55
0.54
S. mikado
/
R: AAGCCCATATATGCTTCCTGG
4H1
F: TATGAAACAGACTTAATCC
FJ221388
(GTTT)n
20
4
203–211
0.85
0.67
S. reevesii
1
R: TGCAGCATTTGAGTAAC
5C9
F: TATGGGAAATGTGTACCTTTA
GQ184557
(CA)n
20
10
221–259
0.95
0.89
C. mantchuricum
10
R: TCCAGGCAACACGTAACA
TT06
F: TGAGAGATTTTGACCCA
GQ181183
(CA)n
20
7
225–237
0.85
0.83
T. temminckii
6
R: CAAGACTTCACCCTACAGATA
4F8
F: GTGGCATGCCTAGTAGATGTT
/
(AC)n
20
11
186–214
0.75
0.88
C. mantchuricum
/
R: CCCTGTGGTACGAACTGTC
SR11
F: ATCAATATGGACTGCTCCGT
FJ221381
(TG)n
20
5
210–248
0.55
0.58
S. reevesii
17
R: TCCTTCAAGGCCAAGTG
5H7
F: CCAAGAGGGAGGCACACGTTC
U60782
(TG)n
20
3
186–194
0.55
0.42
G. gallus
8
R: AGCCATAAATAAGCAAACGC
4C12
F: ATAGGCGGACAGAGGATAGA
FJ221385
(CA)n
20
4
160–170
0.30
0.59
S. reevesii
Z
R: CCCCGCATCGAGGTG
Notes: n is the number of successfully genotyped individuals and NA the number of alleles. HO is observed heterozygosity and HE expected heterozygosity.
We blast the acquired microsatellite sequences from C. auritum on the chicken genome in GenBank to find their locations and to investigate the potential linkages among the loci. If some loci were mapped in sex chromosome, we carried out sexing identification with our sample set by using primer sex1/sex2 (Wang and Zhang, 2009) to cross-validate the sex-linkage loci.
We tested observed heterozygosity (HO), expected heterozygosity (HE), the number of allele per locus, the Hardy-Weinberg equilibrium (HWE) and linkage-disequilibrium in each population using Arlequinv.3.11 (Excoffier et al., 2005). We calculated the frequencies of null alleles by using FreeNA (Chapuis and Estoup, 2007). High polymorphic markers with HWE were used for paternity tests in CERVUS 3.0 with 100000 times simulations to estimate their resolving power. Significant levels were recorded after application of the sequential Bonferroni correction (Rice, 1989; Excoffier et al., 2005).
Results
Sixty-two (55.4%) of the 112 cross-species markers could be efficiently amplified in C. auritum, while 11 (17.7%) of the 62 markers had a moderate to high level of polymorphism (3–11 loci), with the expected heterozygosities ranging from 0.42 to 0.89 (Table 1). Locus 4C12 was found to be homozygous in heterogametic females and heterozygous in 56.2% of the males (n = 28) and thus most likely to link with the Z chromosome. Three loci (1H4, 2420 and 4F8) were not targeted on the chicken genome. All loci were at the HWE; neither linkage disequilibrium among pairs of loci nor null alleles was found. The parentage analysis results showed that if neither parent was known, the successful assignment rates were 98% at a 95% confidence level and 98% at an 80% confidence level. Paternity test results are consistent with known pedigrees.
Discussion
The phylogenetic relationship between the original and target species seems to affect the success of cross-species amplification (Primmer et al., 1996). In our analyses, cross-species amplification from Crossoptilon was the most effective (2/4 = 50%), followed by those from Syrmaticus (6/21 = 28.6%), Meleagris (1/16 = 6.25%), Tragopan (1/30 = 3.0%) and Gallus (1/41 = 2.4%). Phylogenetic analysis showed a successive relationship between Crossoptilon and the genera mentioned earlier (Kimball et al., 2011), indicating that evolutionary relationships may play an important role in cross-species amplification of microsatellite markers within Galliformes.
Although large numbers of microsatellite markers are found in autosomes, Z-linked microsatellites markers are still rarely available, even in the well-characterized chicken genome (Groenen et al., 2000). One Z-linked marker TUT, originally isolated from the Tetrao urogallus (Segelbacher et al., 2000, Wang et al., 2011), failed to be amplified in chickens and might be specific for Tetraoninae grouse. In the present study, the Z-linked polymorphic locus 4C12 seems to have general application in different Galliforme species and might yet provide a valuable tool in kinship and demographic analyses combined with other loci.
Birkhead TR, Johnson SD, Nettleship DN. Extra-pair matings and mate guarding in the Common Murre Uria Aalge. Anim Behav. 1985; 33: 608-619
Boucher-Lalonde V, Morin A, Currie DJ. A consistent occupancy-climate relationship across birds and mammals of the Americas. Oikos. 2014; 123:1029-1036
David M, Auclair Y, Cezilly F. Personality predicts social dominance in Female Zebra Finches, Taeniopygia guttata, in a feeding context. Anim Behav. 2011; 81:219-224
Deeming DC, Biddle LE, Christopher RF. Interspecific and intraspecific spatial separation by birds breeding in nest boxes. Avian Conserv Ecol. 2017; 12:1
Dhondt, A.A., 1977. Interspecific competition between Great and Blue Tit. Nature 268, 521–523
Dhondt AA. Effects of competition on great and blue tit reproduction: Intensity and importance in relation to habitat quality. J Anim Ecol. 2010; 79: 257-265
Dhondt, A.A., 2012. Interspecific competition in birds. In: Birkhead, T.R. (Ed.), Oxford Avian Biology Series. Oxford University Press, Oxford, UK.
Evans MR, Lank DB, Boyd WS, Cooke FA. Comparison of the characteristics and fate of Barrow’s Goldeneye and Bufflehead nests in nest boxes and natural cavities. The Condor. 2002; 104:610-619
Finch DM. Effects of predation and competitor interference on nesting success of house wrens and tree swallows. The Condor. 1990; 92:674-687
Fowler, J., Cohen, L., Jarvis, P., 1998. Practical Statics for Field Biology, second ed. Wiley, Chichester, UK.
Gurevitch J, Morrow LL, Wallace A, Walsh JS. A meta-analysis of competition in field experiments. Am Nat. 1992; 140:539-572
Kruger O, Chakarov N, Nielsen JT, Looft V, Grunkorn T, Struwe-Juhl B, et al. Population regulation by habitat heterogeneity or individual adjustment? J Anim Ecol. 2012; 81:330-340
Laube, I., 2011. Avian Range Dynamics: Traits, Biotic Interactions and Niches. Ph.D thesis. Goethe University Frankfurt, Frankfurt.
Mand RV, Tilgar A, Lohmus, Leivits A. Providing nest boxes for hole-nesting birds - does habitat matter? Biodiver Conserv. 2005; 14:1823-1840
Martin TE. Nest predation and nest sites new perspectives on old patterns. BioScience. 1993; 43:523-532
Martin TE. Food as a limit on breeding birds: A life-history perspective. Ann Rev Ecol System. 1987; 18:453-487
Martinez-Rivera CC, Gerhardt HC. Advertisement-call modification, male competition, and female preference in the bird-voiced treefrog Hla avivoca. Behav Ecol Sociobiol. 2008; 63:195-208
Minot EO, Perrins CM . Interspecific interference competition - nest sites for blue and great tits. J Anim Ecol. 1986; 55: 331-350
Moeller AP, Adriaensen F, Artemyev A, Banbura J, Barba E, Biard C. Clutch-size variation in western Palaearctic secondary hole-nesting passerine birds in relation to nest box design. Methods Ecol Evol. 2014; 5:353-362
Moeller AP, Balbontin J, Dhondt AA, Remes V, Adriaensen F, Biard C. Effects of interspecific coexistence on laying date and clutch size in two closely related species of hole-nesting birds. J Anim Ecol. 2018; 87:1738-1748
Schoener TW. Field experiments on interspecific competition. Am Nat. 1983; 122:240-285
Szabo ND, Algar AC, Kerr JT. Reconciling topographic and climatic effects on widespread and range-restricted species richness. Glob Ecol Biogeogr. 2009; 18:735-744
Tarjuelo R, Morales MB, Arroyo B, Manosa S, Bota G, Casas F. Intraspecific and interspecific competition induces dependent habitat niche shifts in an endangered steppe bird. Ecol Evol. 2017; 7:9720-9730
Teather KL. An experimental study of competition for food between male and female nestlings of the red-winged blackbird. Behav Ecol Sociobiol. 1992; 31:81-87
Zhang L, Wang J, Wan DM, Bi YJ, Zhang L. Narrow or wide distribution? A comparison of behavioral traits between the Varied Tit (Sittiparus varius) and Great Tit (Parus cinereus). Chin J Zool. 2019; 54:471-477
Ibrahim M. Ahmad, Dongming Li. More than a simple egg: Underlying mechanisms of cold tolerance in avian embryos. Avian Research, 2023, 14: 100104.
DOI:10.1016/j.avrs.2023.100104
2.
Ava-Rose F Beech, Mattheus C Santos, Emily B Smith, et al. High within-clutch repeatability of eggshell phenotype in Barn Swallows despite less maculated last-laid eggs. Ornithology, 2022, 139(4)
DOI:10.1093/ornithology/ukac024
3.
Joanna Rosenbeger, Kamil Pytlak, Ewa Łukaszewicz, et al. Variation in Bird Eggs—Does Female Factor, Season, and Laying Order Impact the Egg Size, Pigmentation, and Eggshell Thickness of the Eggs of Capercaillie?. Animals, 2021, 11(12): 3454.
DOI:10.3390/ani11123454
4.
Longwu Wang, Gangbin He, Yuhan Zhang, et al. Cryptic eggs are rejected less frequently by a cuckoo host. Animal Cognition, 2021, 24(6): 1171.
DOI:10.1007/s10071-021-01507-2
5.
Wei-Guo Du, Richard Shine, Liang Ma, et al. Adaptive responses of the embryos of birds and reptiles to spatial and temporal variations in nest temperatures. Proceedings of the Royal Society B: Biological Sciences, 2019, 286(1915): 20192078.
DOI:10.1098/rspb.2019.2078
6.
Ping Ye, Canchao Yang, Wei Liang. Nest site availability and niche differentiation between two cavity‐nesting birds in time and space. Ecology and Evolution, 2019, 9(20): 11904.
DOI:10.1002/ece3.5698
Table
1.
Characterization of 11 microsatellite loci
Locus
Primer sequences (5′–3′)
Accession No.
Repeat motifs
n
NA
Size range (bp)
HO
HE
Species origin
Chromosome
2580
F: TTAACCTATCAGGTCGTTGCG
AL592580
(CA)n
20
8
191–213
1.00
0.80
M. gallopavo
21
R: CAGTGCACATGCAGGCAG
3D2
F: TCTCTGACGTATCGCATCT
FJ221373
(GT)n
19
6
286–304
0.47
0.58
S. reevesii
4
R: ACTTCCCCTGGTAAACT
1H4
F: TGAACAAGTGAGGCGGAGC
/
(TG)n
20
10
127–161
0.65
0.81
S. reevesii
/
R: CTGCACACAGCCCGAAGC
2420
F: CATCATCTGCCAATGCAGAGG
/
(TTTA)n
20
4
118–142
0.55
0.54
S. mikado
/
R: AAGCCCATATATGCTTCCTGG
4H1
F: TATGAAACAGACTTAATCC
FJ221388
(GTTT)n
20
4
203–211
0.85
0.67
S. reevesii
1
R: TGCAGCATTTGAGTAAC
5C9
F: TATGGGAAATGTGTACCTTTA
GQ184557
(CA)n
20
10
221–259
0.95
0.89
C. mantchuricum
10
R: TCCAGGCAACACGTAACA
TT06
F: TGAGAGATTTTGACCCA
GQ181183
(CA)n
20
7
225–237
0.85
0.83
T. temminckii
6
R: CAAGACTTCACCCTACAGATA
4F8
F: GTGGCATGCCTAGTAGATGTT
/
(AC)n
20
11
186–214
0.75
0.88
C. mantchuricum
/
R: CCCTGTGGTACGAACTGTC
SR11
F: ATCAATATGGACTGCTCCGT
FJ221381
(TG)n
20
5
210–248
0.55
0.58
S. reevesii
17
R: TCCTTCAAGGCCAAGTG
5H7
F: CCAAGAGGGAGGCACACGTTC
U60782
(TG)n
20
3
186–194
0.55
0.42
G. gallus
8
R: AGCCATAAATAAGCAAACGC
4C12
F: ATAGGCGGACAGAGGATAGA
FJ221385
(CA)n
20
4
160–170
0.30
0.59
S. reevesii
Z
R: CCCCGCATCGAGGTG
Notes: n is the number of successfully genotyped individuals and NA the number of alleles. HO is observed heterozygosity and HE expected heterozygosity.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds