Figure
1.
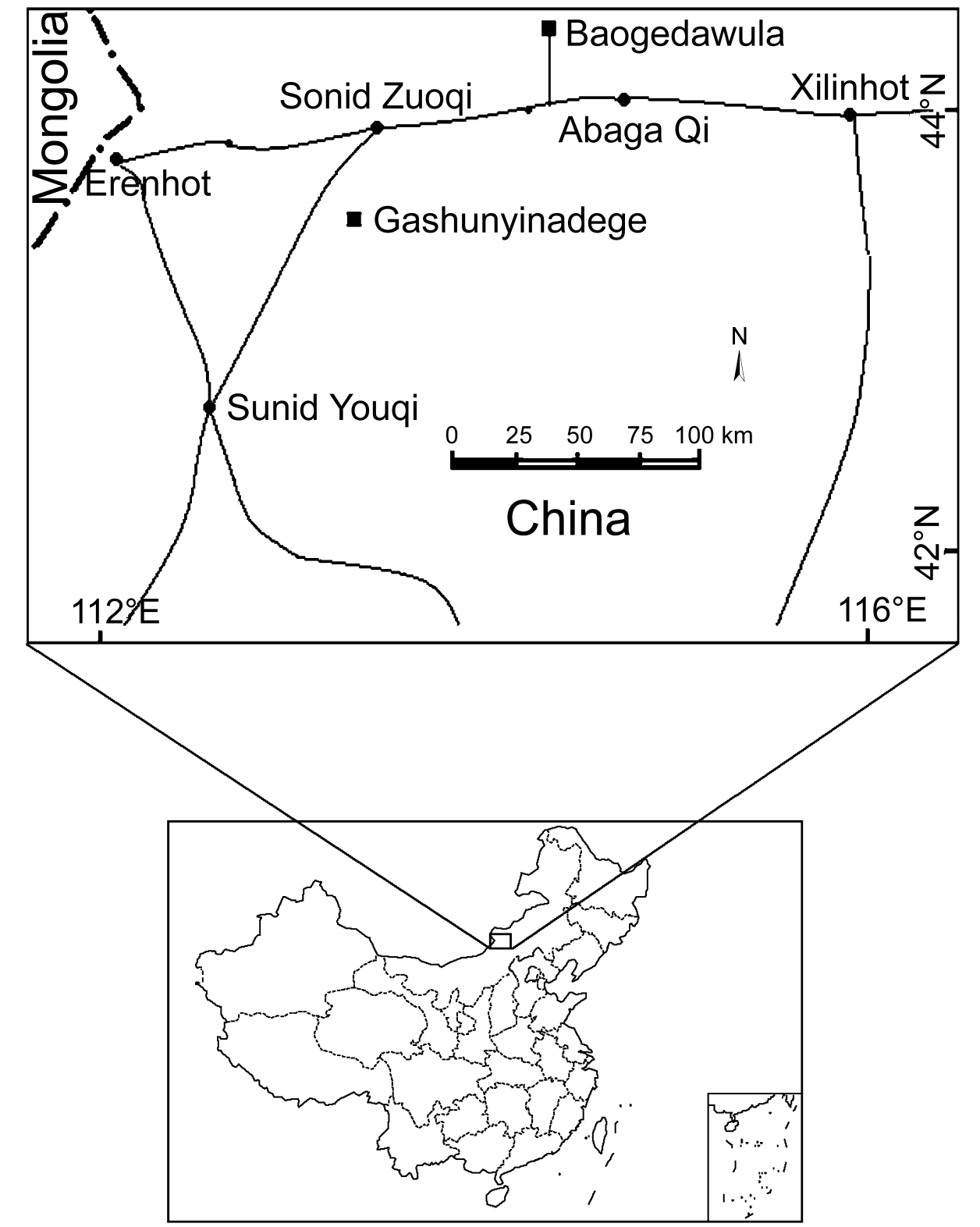
Geographic locations of fossil localities in Inner Mongolia, China
| Citation: |
Shuo WANG, Ying HU, Lu WANG. 2011: New ratite eggshell material from the Miocene of Inner Mongolia, China. Avian Research, 2(1): 18-26. DOI: 10.5122/cbirds.2011.0002

|
Some Miocene ratite eggshell fragments from Gashunyinadege (about 17.5 Ma) and Baogedawula (7.11 ± 0.48 Ma), Inner Mongolia, China, are described. The outer surfaces of eggshell fragments from Gashunyinadege are smooth with a sting pore pattern, while their entire thickness is around 2.12 mm. In a cross-sectional view, the surface crystal layer is very thin and the ratio of cone layer to the columnar layer is about 1:2.4. The specimens from Baogedawula present similar microscopic characters as those from Gashunyinadege, but seem much thinner and have a lower pore density. Both characteristics fit aepyornithid-type eggshells, implying that the distribution of aepyornithid-type eggshells can be extended to East Asia. The sediments at Gashunyinadege date to approximately 17.5 Ma, implying that the specimens from this locality are relatively early examples of aepyornithid-type eggshells. Aepyornithid-type eggshells seem to have appeared in Eurasia much earlier than struthionid-type eggshells. The faunal compositions in the two localities reflect gradual drying of the Mongolian plateau during the Miocene. A reduction in the number of pores 10 Ma ago, probably occurred in response to this drying process.
Ratites, a clade of birds lacking the ability to fly, most likely originated in Gondwana (Patnaik et al., 2009). Large ratite eggs are frequently found preserved and recorded in Tertiary and Quaternary sediments. Ratite skeletons, however, are rarely preserved, so that fossil eggshells constitute a richer and more widely used source of information on ratite evolution. The distribution of extant ratites is essentially restricted to the Southern Hemisphere (Cooper et al., 2009), but several fossils have been reported from the Northern Hemisphere (Mayr, 2009). Fossils have been found in Eurasia (Lowe, 1933; Burchak-Abramovich, 1962; Sauer and Rothe, 1972; Hou et al., 2005; Bibi et al., 2006; Wang, 2008; Patnaik et al., 2009), northern Africa (Sauer, 1972; Mikhailov, 1997) and even in North America (Houde and Olson, 1981). The biogeographic history of ratites through geological time remains an active and contentious area of research.
The first description of a fossil ratite from Asia was based on some bones collected in the Siwaliks, India (Davies, 1880); this material was subsequently named Struthio asiaticus by Milne-Edwards (Burchak-Abramovich, 1962). In 1874, Brandt (1874) described a ratite eggshell fossil from the Ukraine; since then, a great many reports concerning Eurasian fossil ratites have been published (Burchak-Abramovich, 1962; Sauer, 1972; Mikhailov, 1988; Mikhailov and Kurochkin, 1988). In China, ratite research has a long history. Lowe reported Struthio anderssoni, S. mongolicus and S. wimani from Plio-Pleistocene sediments in China (Lowe, 1931), Yang described some additional specimens from Shanxi and Hebei provinces and the Zhoukoudian locality near Beijing, and discussed their biostratigraphic significance (Yang, 1933, 1937, 1947, 1952; Yang and Sun, 1960). Zhao and his colleagues discussed the microstructure and amino acid composition of ratite eggshell fossils from the Zhoukoudian locality (Zhao et al., 1981). Hou reported two pelvic skeletons from the Miocene sediments of Hezheng (Hou et al., 2005) and more recently, fossil eggshells from the same site have been described by Wang(2007, 2008).
During the past two decades, several ratite eggshell fossils have been collected and reported from other continents, particularly from Africa (Pickford and Dauphin, 1993; Pickford et al., 1995; Dauphin et al., 1996; Harrison and Msuya, 2005). They have been used to analyze ratite evolutionary relationships, discuss changes in their geographic distribution and establish biostratigraphic divisions. These recent studies indicate that the fossil eggshell record is potentially far more informative than previously suspected.
Recently, a special field project carried out by the IVPP (Institute of Vertebrate Paleontology and Paleoanthropology, Chineae Academy of Sciences) in Inner Mongolia, China focused on the study of Miocene mammals. This expedition acquired a large collection of fossil micromammals in addition to some other specimens, including ratite eggshells. We briefly describe some new fossil eggshell material from the Gashunyinadege and Baogedawula localities in Inner Mongolia. These specimens provide important new data on the distribution of fossil ratites in China.
Ten eggshell fragments were recovered from two localities, six from Gashunyinadege and four from Baogedawula (Fig. 1). Gashunyinadege is about 40 km south of Sonid Zuoqi (Sunitezuoqi), Inner Mongolia (43°33′39.2″N, 113°32′51.9″E) and has been studied in detail by various paleontologists after being initially reported by Meng and others (Meng et al., 1996; Qiu and Wang, 1999; Qiu et al., 2006; Wang et al., 2009). The Gashunyinadege fauna was initially assigned a Late Oligocene-Early Miocene age (Meng et al., 1996), but was later reassessed as an Early Miocene deposit (about 17.5 Ma) on the basis of micromammals (Qiu et al., 2006). Baogedawula, however, is some 3 km northeast of Baogeawula Sumu, Abaga Qi (44°08′33.8″N, 114°35′41.6″E), Inner Mongolia (Qiu et al., 2006; Wang et al., 2009). The deposits consist of grayish and yellow sandy mudstones and sandstones dating to 7.11 ± 0.48 Ma. For more information about these two localities, see Qiu et al. (2006).
Ratite eggshells can be identified using both macroscopic and microscopic approaches (She, 1995). The macroscopic approach is to observe or measure characters including pore complex structure, pore density and shell thickness using a low power hand lens and a set of calipers (Pickford et al., 1995). The microscopic approach is to polish the eggshells and then section them for scanning electron microscopy (SEM) and optical microscopy in order to observe their internal layering, crystal arrangement and other detailed features. The present study utilizes both approaches to analyze the new Inner Mongolian eggshell fragments and try to arrive at a comprehensive description.
The thickness of each eggshell fragment was measured using calipers; subsequently, we selected three well-preserved eggshell fragments from each fossil site and used them to prepare microscopic samples. SEM samples were used for observing the radial section of each selected fragment as well as its inner and outer surfaces. Each sample was first polished with sandpaper, then etched with 1% hydrochloric acid (5 s for the inner surface, 18 s for the outer surface and 45 s for the radial section), which was then washed off and coated with gold in a vacuum environment (Becking, 1975; Pooley, 1979; Mikhailov, 1987; She, 1995; Mikhailov, 1997). Finally, scanning electron microscopy (JSM model No. 6100) was used to examine each prepared sample and make measurements of pore diameters. We also made thin sections in order to observe each sample using optical microscopy.
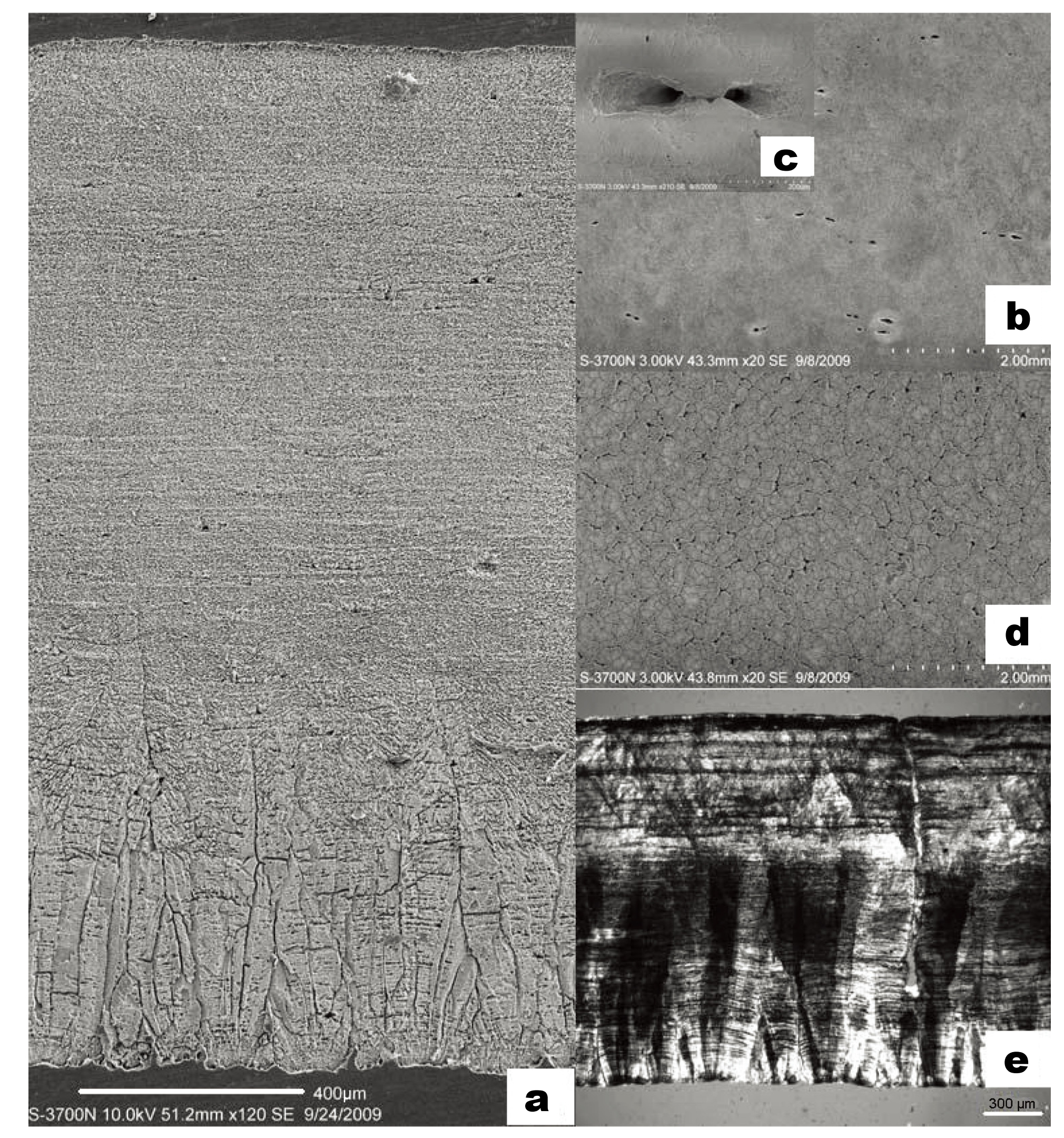
The eggshell samples collected from Gashunyinadege are smooth, with sting-point pores distributed on their outer surfaces, producing no depressions or pore complexes. The pores can be seen with the naked eye (Fig. 2, left). Through a low power hand lens, most pores can be seen to occur in pairs and to range in diameter from 0.1–0.2 mm; however, some of the pores are isolated. These features are consistent with the normal characteristics of sting pores (Sauer, 1972). As to the pore density, more than 70 pores per cm2 can be seen with the naked eye, compared to 130 pores per cm2 under SEM examination. Their entire thickness averages around 2.12 mm (from 1.7 to 2.2 mm; n = 6 measured in 36 places). SEM examination (Fig. 3) shows that most of the pores are elliptical or in sting shape (Fig. 3b, c). In a cross-sectional view (Fig. 3a), the surface crystal layer is very thin, the outer portion of the column layer is slightly thicker than in Orientornis linxiaensis (Fig. 3e) and the ratio of the thickness of the cone layer to that of the entire column layer is about 1:2.4. The cunei run from the cone layer to the inner column layer. The pore canals are non-branching and straight (Fig. 3e).
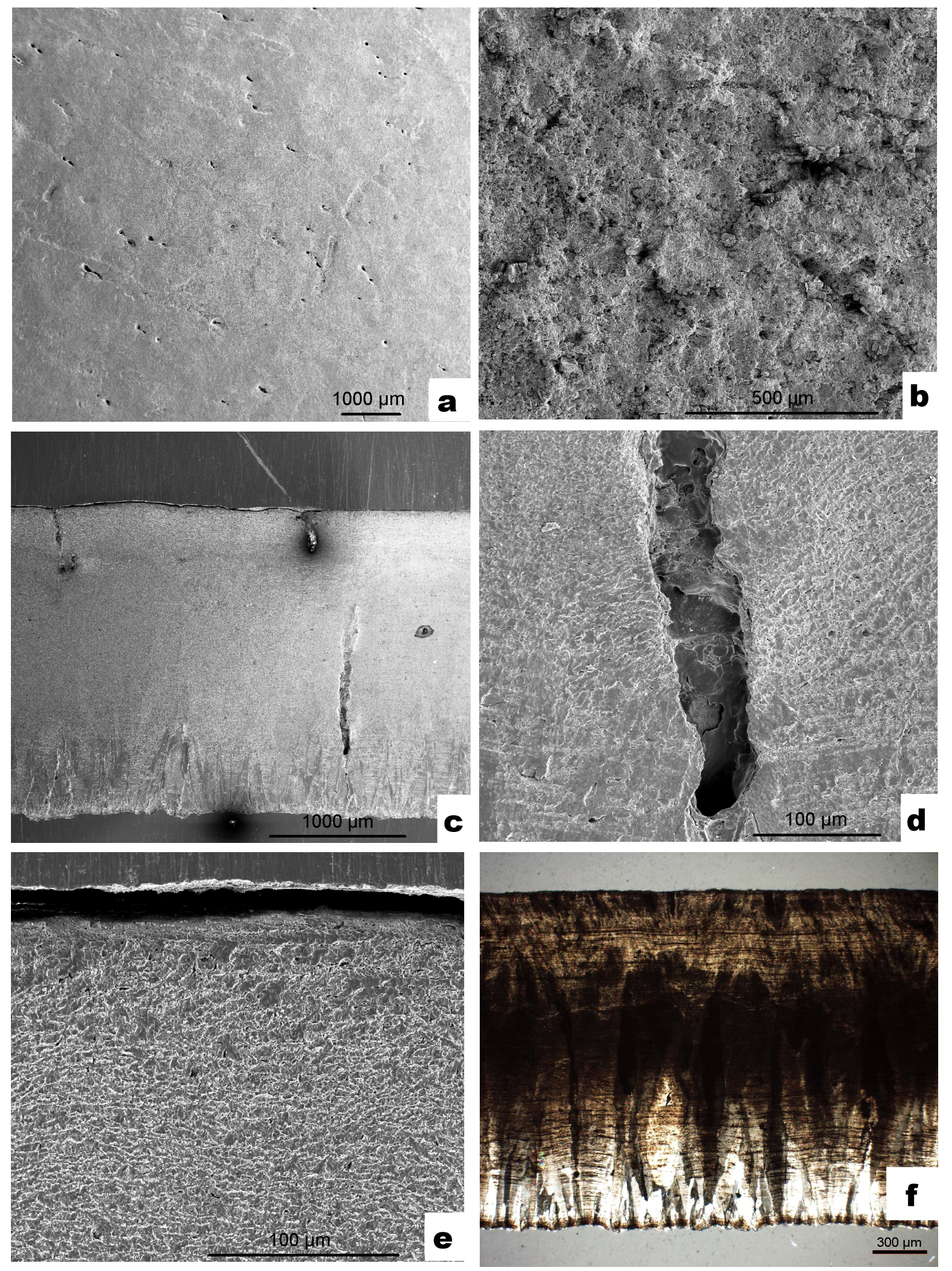
Samples from Baogedawula are similar in their pore pattern to those from Gashunyinadege. It is clear that small pore openings are irregularly distributed on the smooth outer surface, producing no depressions or pore complexes (Fig. 2, right). They show a relatively low pore density, about 8–15 pores per cm2 can be seen with the naked eye, but under the SEM, more tiny pores appear in the field (Fig. 4a) and the density rises to about 50 pores per cm2. From the SEM results (Fig. 4), the pore diameter ranges from 0.05–0.2 mm and the pores are round or elliptical and uniformly distributed on the outer surface. The pore distribution seems to display some regularity, in that the elliptical pores are always in pairs and the round ones are single. Details of the inner surface, however, cannot be observed clearly because they are obscured by sediment (Fig. 4b). As the cross-sectional view shows, the samples range in thickness from 1.7 to 2.1 mm, with a mean of 2.02 mm (n = 4 measured in 23 places); the pore canals are single and straight (Fig. 4c), with rough inner surfaces (Fig. 4d); the columnar layer is compact (Fig. 4e); the ratio of the thickness of the cone layer to that of the columnar layer is about 1:1.4. The cone layer is thinner, representing about one third of the entire shell thickness, the cones are slender and compact (Fig. 4f).
Obviously, the eggshells from Gashunyinadege and Baogedawula share a smooth outer surface; although the Baogedawula samples show considerable wear (Fig. 4a) and are close in thickness to those from Gashunyinadege. However, there are also some significant differences between the eggshell fragments from Gashunyinadege and those from Baogedawula. Pore density in the Gashunyinadege samples is more than 70 per cm2, slightly higher than in the Baogedawula samples. In cross-sectional view, the cone density of the Gashunyinadege samples is somewhat higher than in the Baogedawula samples; furthermore, the column layer is much thicker in the former than in the latter and the Gashunyinadege samples also differ from the Baogedawula samples in having a relatively thicker outer column layer.
According to this description, these new eggshell fragments differ from Aepyornis eggshells in having a generally lower thickness and lacking pore grooves (Bibi et al., 2006), differ from Diamantornis eggshells in having a lower thickness and in lacking pore complexes (Pickford and Dauphin, 1993). Compared with Struthio camelus, the pore density of these Inner Mongolian eggshell fossils is higher; they also differ from S. camelus in lacking pore complexes. Their thickness is slightly less than in Struthio chrosonensis, S. kakesiensis, S. karingarabensis (Harrison and Msuya, 2005) and even in Orientornis linxiaensis (Wang, 2007). Also, pores of Struthio daberasensis clump together and produce "a reticulation of minuscule flat islands" (Sauer, 1972) while the pores of the new specimens are paired or single. The new specimens also differ from S. anderssoni in lacking pore pits. Sauer (1972) generalized the characters of the struthionid pore pattern, emphasizing that most of the clustered pores and grooves are located in pits and that there is no pore alignment in parallel with the axes of the egg; besides, the pore canals seen in struthionid eggshell are divergent. However, the new Inner Mongolian specimens lack these critical characters, implying that they do not represent typical struthionid eggshell. Previously, almost all ostrich eggshell fossils in China were described as S. anderssoni (Lowe, 1931), but recent studies suggest that additional types of eggshell fossils exist in China. The new specimens represent one such type, but we consider the material too fragmentary to justify the formal erection of a new taxon.
Both similarities and differences between these Inner Mongolian fossils and Orientornis linxiaensis are obvious. The eggshell fossils from these two independent localities share characters including a smooth outer surface and similar pore shape. The pores of the new specimens are larger and more densely distributed than in O. linxiaensis. Furthermore, the column layer is proportionally thinner in the new material than in O. linxiaensis.
Ratite eggshells are generally classified as either struthionid-type or aepyornithid-type (Patnaik et al., 2009). Sauer (1972) first described ratite eggshells with smooth outer surfaces and a sting-point pore pattern. According to his description, the pore pattern is the key distinguishing character of this type of eggshell. Pores on these eggshells are always linearly aligned long, and form short grooves with a dagger point, tiny circular to ovoid needle point pore shapes. Because these characters are quite similar to those of Aepyornis maximus in Madagascar, many authors have designated smooth eggshells of this kind as aepyornithid-type eggshells (Sauer and Sauer, 1978; Bibi et al., 2006). In gross morphology, the new Inner Mongolian fossils are similar to aepyornithid-type eggshells: their sting-point pores occur in rows that are conspicuously oriented parallel to one another very close in thickness to aepyornithid-type eggshells from Lanzarote (Sauer and Rothe, 1972). The only difference is that the new specimens lack pore grooves. Hence we are inclined to believe that these Inner Mongolian fossil materials may belong to aepyornithid-type eggs. Unfortunately, however, most studies on aepyornithid-type eggshell have not presented cross-sectional features, so that the new specimens cannot be readily compared to previously described aepyornithid-type material with respect to cross-sectional morphology.
Most previously collected aepyornithid-type eggshell material has come from the Miocene-Pliocene sediments of western Europe and Asia (Sauer, 1972; Sauer and Rothe, 1972; Sauer, 1976; Sauer and Sauer, 1978; Stern et al., 1994; Harris and Leakey, 2003); however, Sauer (1972) reported some aepyornithid-like eggshell fragments from the Eocene Irdin Manha Formation. At Camp Margetts, near Iren Dabasu, Inner Mongolia, China (Sauer, 1972), and Bibi et al. (2006) reported aepyornithid-type eggshells from the Oligocene sediments of Mongolia. If these occurrences can be confirmed, the stratigraphic range of aepyornithid-type eggshells in East Asia would extend down into the Eocene (Bibi et al., 2006). It appears to be true because the age of the Gashunyinadege eggshells were dated at about 17.5 Ma, close to the Oligocene-Miocene boundary. It is quite possible that even earlier eggshells, from the Oligocene or Eocene, do in fact exist.
Patnaik et al. (2009) suggested that ratites dispersed into Eurasia from Africa via an India-Madagascar biogeographic link. The Indian subcontinent was connected with the African plate during the Cretaceous. Subsequently, the India subcontinent drifted northward, and collided with Asia in a process that started at about 65 Ma and was completed by the end of Eocene (Zhu et al., 2005; Mo et al., 2007). If this biogeographic hypothesis is true, early ratites would be expected to have spread into Eurasia not earlier than the Eocene. However, more fossil evidence from the Indian subcontinent is needed in order to confirm this view. Another important aspect of ratite paleobiogeography is that both struthionid-type and aepyornithid-type eggshells have been found in Lanzarote, Canary Islands (Sauer and Rothe, 1972), in Siwalik (Sauer, 1972; Stern et al., 1994; Patnaik et al., 2009) and in Inner Mongolia (Sauer, 1972). In other words, the two types occur together at sites located from western Europe to East Asia. However, the aepyornithid-type eggshells from these three localities seem much older than the associated struthionid-type eggshells (Sauer, 1972; Sauer and Rothe, 1972; Patnaik et al., 2009). From the perspective of biochronology, the eggshells collected from the Irdin Manha Formation (Sauer, 1972) and Gashunyinadege might be considered the earliest presently known aepyornithid-type eggshell fossils (respectively Eocene, dated to 17.5 Ma), whereas struthionid-type eggshells occur in the Pliocene of this area according to Sauer (1972). Stern et al. (1994) identified some aepyornithid-type eggshells from Siwalik (about 11 Ma, whereas struthionid-type eggshell fragments from the same area range in age from 2.2 to 0.5 Ma). Orientornis linxiaensis (9.6 Ma) eggshells have a smooth surface, but are characterized by pores that are different from those of the Gashunyinadege specimens (Wang, 2007). Finally, the Baogedawula (7.11 Ma) samples again display a smooth outer surface and aepyornithid-like pore shape. The record suggests that ratites that laid aepyornithid-type eggs were present in Eurasia well before those that produced struthionid-type eggs. We are not sure whether this phenomenon simply implies two independent dispersal events of ratites, an early dispersal involving aepyornithid-laying birds and a later one involving struthionid-laying birds, into Eurasia.
Mikhailov and Kurochkin reviewed the morphology and distribution of ratite eggshell fossils from Eurasia based on a large amount of material (Mikhailov, 1988; Mikhailov and Kurochkin, 1988); they concluded that aepyornithid-type eggshells appeared and disappeared periodically during the Neogene, alternating with struthionid-type eggshells. They considered both types of eggshell to belong to the genus Struthio. The characteristics that define the two types, i.e., pore structure and shell thickness, varied in response to the alternation of humid and dry conditions, or cold and warm ones (Mikhailov, 1988; Mikhailov and Kurochkin, 1988). This conclusion is still controversial because their results were largely based on morphological characters and lack supporting geochemical data. Since geochemical data would better reflect changes in climate, Stern et al. (1994) examined stable carbon and oxygen isotopes in ratite eggshell fossils from the Siwaliks, focusing on determining the nature and timing of environmental change in the Himalayan foothills. Stern et al. (1994) claimed that no differences in mineral content exist among aepyornithid-type, struthionid-type and extant ostrich eggshells, whereas the isotope value of aepyornithid-type eggshells is quite similar to that of struthionid-type eggshells at the specific time point of 2.24 Ma. This suggests that both aepyornithid-type and struthionid-type eggshells occurred at the same time period, sharing a similar environment. In contrast, although the δ18O values of aepyornithid-type eggshells from 5.26 to 2.34 Ma are quite different and the δ13C values also indicate their dietary sources had not changed during these 3 Ma (Stern et al., 1994), it has been clearly shown that aepyornithid-type eggshell characteristics were stable during this time and that struthionid-type eggshells did not appear until 2.24 Ma. Therefore, Mikhailov's conclusions might, to some extent, be problematic.
Gashunyinadege and Baogedawula are both important Miocene sites in China; they have been repeatedly mentioned in the literature (Meng et al., 1996; Qiu and Wang, 1999; Qiu et al., 2006; Wang et al., 2009). The Gashunyinadege fauna contains at least 40 taxa of mammals and the Baogedawula fauna 22. Most of the mammals, especially micromammals, are ones that dispersed wilely within Eurasia (Qiu et al., 2006). This suggests that the central Inner Mongolian localities had good possibilities for interchange with other faunas. The faunal compositions of these two localities reflect a gradual drying of the Mongolia plateau during Miocene (Dong et al., 2006; Qiu et al., 2006; Liu et al., 2009). The reduction of the number of pores from the Gashunyinadege sample to the Baogedawula sample, some 10 Ma later, may reflect a response to this drying process.
We thank Dr. Zhuding Qiu, Dr. Qiang Li, Dr. Xijun Ni (IVPP) and Yuri Kimura (National Science Museum, Japan) for preparing specimens; Yannicke Dauphin (University of Paris XI-Orsay, Franch), Dr. E.N. Kurochkin (Paleontological Institute of the Russian Academy of Sciences), Dr. Yaroslav Kuzmin (Institute of Geology & Mineralogy, Russia) and Dr. Rejeev Patnaik (Panjab University, India) for providing important references; Wending Zhang and Shukang Zhang (IVPP) for technical expertise and photography and Corwin Sullivan and Rejeev Patnaik for reviewing the manuscript. This study was supported by the Knowledge Innovation Program of the Chinese Academy of Sciences (KZCX2-YW-120) and the Key Laboratory of Evolutionary Systematics of Vertebrates, Chinese Academy of Sciences (2010LESV007).
|
Becking JH. 1975. The ultrastructure of avian eggshell. Ibis, 117: 143–151.
|
|
Bibi F, Shabel AB, Kraatz BP, Stidham TA. 2006. New fossil Ratite (Aves: Palaeognathae) eggshell discoveries from the late Miocene Baynunah formation of the United Arab Emirates, Arabian Peninsula. Paleontol Electr, 9: 1–3.
|
|
Brandt A. 1874. On a large fossil egg from the neighborhood of Cherson. Ibis, 16: 4–7.
|
|
Burchak-Abramovich. 1962. On the review of fossil ostrich from the old continent. Vertebrat PalAsiatic, 6: 299–309.
|
|
Dauphin Y, Pickford M, Senut B. 1996. Microstructures des coquilles d'oeufs d'oiseaux fossils de Namibie. Revue de Paléobiol, 15: 225–241. (in French)
|
|
Dong MX, Li FL, Gao DZ, Geng MS, Li LY. 2006. A study of the late Miocene dry event based on sporo-pollen variations in Sunitezuoqi, Inner Mongolia. Acta Geosci Sinica, 27: 207–212. (in Chinese)
|
|
Harris JM, Leakey MG. 2003. Lothagam birds. In: Leakey MG, Harris JM (eds) Lothagam: the Dawn of Humanity in Eastern Africa. Columbia University Press, New York, pp 161–166.
|
|
Lowe PR. 1931. Struthious remains from China and Mongolia with description of Struthio wimani, Struthio anderssoni and Struthio mongolicus spp. nov. Palaeontol Sinica, 6: 1–46.
|
|
Lowe PR. 1933. On some Struthious remains: 1. Description of some pelvic remains of a large fossil ostrich, Struthio oldwayi, sp. n. from the Lower Pleistocene of Oldaway (Tanganyika Territorory); 2. Eggshell fragments referable to Psammornis and other Struthiones collected by Mr. St. John Philby in South Arabia. Ibis, 3: 652–658.
|
|
Mayr G. 2009. Paleogene Fossil Birds. Springer, Berlin.
|
|
Meng J, Wang BY, Bai ZQ. 1996. A new Middle Tertiary mammalian locality from Sunitezuoqi, Nei Mongol. Vertebrat PalAsiatic, 34: 297–304.
|
|
Mikhailov KE, Kurochkin EN. 1988. The eggshells of Struthioniformes from the Palearctic and its position in the system of views on Ratite evolution. In: The Joint Soviet-Mongolian Palaeontological Expedition (ed) Fossil Reptiles and Birds of Mongolia, Transactions, vol. 34. Nauka, Moscow, pp 43–64. (in Russian)
|
|
Mikhailov KE. 1987. The principle structure of the avian eggshell; data of SEM studies. Acta Zool Cracovien, 30: 53–70.
|
|
Mikhailov KE. 1988. The comparison of East European and Asian ostriches Pliocene eggshells. In: The Joint Soviet-Mongolian Palaeontological Expedition (ed) Fossil Reptiles and Birds of Mongolia, Transactions, vol. 34. Nauka, Moscow, pp 65–72. (in Russian)
|
|
Mikhailov KE. 1997. Fossil and recent eggshells in amniotic vertebrates: Fine structure, comparative morphology and classification. Spec Pap Palaeontol, 56: 1–80.
|
|
Mo XX, Zhao ZD, Zhou S, Dong GC, Liao ZL. 2007. On the timing of India-Asia continental collision. Geol Bull China, 26: 1240–1244. (in Chinese with English abstract)
|
|
Patnaik R, Sahni A, Cameron D, Pillans B, Chatrath P, Simons E, Williams M, Bibi F. 2009. Ostrich-like eggshells from a 10.1 million-yr-old Miocene ape locality, Haritalyangar, Himachal Pradesh, India. Curr Sci, 96: 1485–1495.
|
|
Pickford M, Dauphin Y. 1993. Diamantornis wardi nov. gen. nov. sp., giant extinct bird from Roilpel, Lower Miocene, Namibia. Comptes Rendus de l'Académie, 316: 1643–1650.
|
|
Pickford M, Senut B, Dauphin Y. 1995. Biostratigraphy of the Tsondab Sandstone (Namibia) based on gigantic avian eggshells. Geobios, 28: 85–98.
|
|
Pooley AS. 1979. Ultrastructural relationships of mineral and organic matter in avian eggshe. Scan Electron Micros, 2: 457–482.
|
|
Qiu ZD, Wang XM, Li Q. 2006. Faunal succession and biochronology of the Miocene through Pliocene in Nei Mongol (Inner Mongolia). Vertebrat PalAsiatic, 44: 164–181. (in Chinese with English abstract)
|
|
Qiu ZD, Wang XM. 1999. Small mammal faunas and their ages in Miocene of central Nei Mongolia (Inner Mongolia). Vertebrat PalAsiatic, 37: 120–139. (in Chinese with English abstract)
|
|
Sauer EGF, Sauer EM. 1978. Ratite eggshell fragments from Mio-Pliocene-continental sediments in the district of Ourzazate, Morocco. Palaeontogr Abt A, 161: 1–54.
|
|
Sauer EGF. 1972. Ratite eggshells and phylogenetic questions. Bonner Zool Beitr, 23: 3–48.
|
|
Sauer EGF. 1976. Aepyornithid eggshell fragments from the Miocene and Pliocene of Anatolia, Turkey. Palaeontogr Abt A, 153: 62–115.
|
|
She DW. 1995. A study of eggshells with scanning electron microscopy. Acta Zool Sin, 41: 243–255. (in Chinese with English abstract)
|
|
Stern LA, Johnson GD, Chamberlain CP. 1994. Carbon isotope signature of environmental change found in fossil ratite eggshells from a South Asian Neogene sequence. Geology, 22: 419–422.
|
|
Wang S. 2008. Rediscussion in the taxonomic assignment of "Struthio linxiaensis Hou, et al., 2005. Acta Paleotologica Sinica, 47: 362–368.
|
|
Wang XM, Qiu ZD, Li Q, Tomida Y, Kimura Y. 2009. A new early to late Miocene fossilferous region in central Nei Mongol: lithostratigraphy and biostratigraphy in Aoerban strata. Vertebrat PalAsiatic, 47: 111–134.
|
|
Yang ZJ, Sun AL. 1960. New material of ostrich eggs and its stratigraphic significance. Vertebrat PalAsiatic, 2: 115–119.
|
|
Yang ZJ. 1933. On the new find of fossil eggs of Struthio anderssoni Lowe in North China, with remarks on the egg remains found in Shansi, Shensi and in Choukoutien China. Bull Geol Soc, 12: 145–152.
|
|
Yang ZJ. 1937. Fossil ostrich egg in China. Geol Rev, 2: 267–272. (in Chinese)
|
|
Yang ZJ. 1947. On the new discovery of Struthio anderssoni. Geol Rev, 12: 159–161. (in Chinese)
|
|
Yang ZJ. 1952. On the fossil ostrich egg from Huixian, Pingyuan Province. Chinese Sci Bull, 3: 404–405. (in Chinese)
|
|
Zhao ZK, Yuan JQ, Wang K. 1981. The microstructure and amino acid analysis of the ostrich eggshell from Zhou Koudian. Vertebrat PalAsiatic, 19: 327–335. (in Chinese with English abstract)
|
|
Zhu B, Kidd WSF, Rowley DB, Currie BS, Shafique N. 2005. Age of Initiation of the India-Asia Collision in the East-Central Himalaya. J Geol, 113: 265–285. (in Chinese with English abstract)
|
Figures(4)





 DownLoad:
DownLoad:

 Email Alerts
Email Alerts RSS Feeds
RSS Feeds