Key Laboratory of Animal Physiology, Biochemistry and Molecular Biology of Hebei Province, College of Life Sciences, Hebei Normal University, Shijiazhuang 050024, China
2.
Ocean College of Hebei Agricultural University, Qinhuangdao 066003, China
Bringing free-living animals into captivity subjects them to the stress of both capture and captivity, leading to the alteration of normal physiological processes and behaviors through activation of the hypothalamic-pituitary-adrenal axis. In free-living birds, although elevated plasma corticosterone (CORT) is an important adaptation regulating physiological and behavioral responses during the process of capture and captivity stress, little information is currently available on the effects of such stress on plasma metabolite levels.
Methods
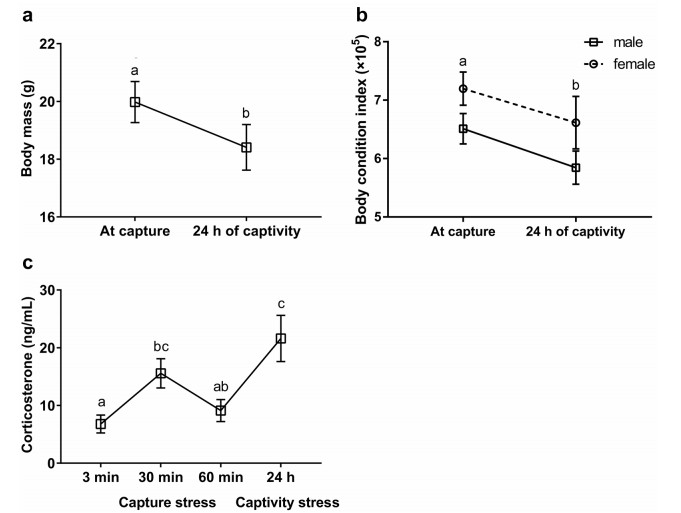
We examined the effects of immediate capture and 24-h captivity on body mass, body condition, plasma CORT, and metabolite levels including glucose (Glu), triglyceride (TG), total cholesterol (TC), uric acid (UA), in breeding Eurasian Tree Sparrows (Passer montanus).
Results
CORT and Glu levels were increased significantly by the stress of capture, whereas TC and UA levels decreased. Body mass, body condition declined notably after 24 h in captivity, but CORT, Glu, and UA levels increased. Furthermore, male sparrows had lower TG levels after both capture and captivity than those of females. The relationships between plasma CORT and metabolite levels varied between sexes.
Conclusions
Our results revealed that the metabolic status of Eurasian Tree Sparrows could be dramatically altered by capture and captivity. Monitoring the dynamic effects of both capture and captivity on plasma CORT, metabolite levels in a free-living bird contributes to a better understanding of the stress-induced pathways involved in sex-dependent energy mobilization.
Globally, the grassland biome and associated species are threatened and require management interventions to prevent further habitat loss and species extinction (With et al. 2008; Ceballos et al. 2010; Richardson et al. 2014). In southern Africa, grasslands are also one of the most threatened biomes, with 23% under cultivation, 60% irreversibly transformed, 2% formally protected and most of the remaining natural areas used as rangeland for livestock (Fairbanks et al. 2000; Reyers et al. 2001; O'Connor and Kuyler 2009). An assessment of conservation priorities in the Grassland Biome in southern Africa identified 36.7% of the biome as being important for biodiversity conservation (Egoh et al. 2011). In particular, South Africa's moist grasslands harbor globally significant biodiversity, supply essential ecosystem services, support crop and livestock, forestry and settlement, yet are poorly conserved (O'Connor and Kuyler 2009). This area also coincides with high avifaunal species richness, diversity and endemism (Maphisa et al. 2016). As a result, a number of Important Bird and Biodiversity Areas (IBAs) have been proclaimed in the area (e.g. Barnes 1998). Despite this, only a few studies (e.g. Mentis and Bigalke 1981; Jansen et al. 1999) have examined abundance or densities of bird species in relation to habitat features that can be managed. Regionally, fire is an important conservation management tool used both on private and state lands to make habitats suitable for birds (Parr and Chown 2003). In addition to heavy livestock grazing and other agricultural activities, this area is a target for energy development projects to meet growing human demand for electricity and water for consumption (Maphisa et al. 2016).
Basing conservation decisions on reliable density estimates is critical to halt current species habitat loss and species extinction (Wintle et al. 2005; Post van der Burg et al. 2011). However, estimating population densities of animals is notoriously difficult because some animals remain undetected during surveys (Diefenbach and Brauning 2003). Many grassland bird species look alike and are therefore difficult to identify in the field. Furthermore, the detection probability varies across methods and different observers, changes with habitats and weather conditions (Petitot et al. 2014). Hierarchical distance-sampling methods account for the detection process and are therefore commonly used to study animal density (Marques et al. 2007; Oedekoven et al. 2013). Improvements in distance sampling allow for the incorporation of environmental and habitat variables to fully explore variation in density of species across sites (Royle et al. 2004; Thomas et al. 2010).
Hierarchical distance sampling models consist of two components (Fiske and Chandler 2011; Sillett et al. 2012): the first component models the observation process (detection probability as a function of distance from the transect line) and the second models the biological process (density of the focal bird species). The observation model involves the choice of a distance function whose parameters will be estimated from the data. This approach has recently been used to relate density of birds to habitat variables within Fynbos Vegetation in South Africa (Lee et al. 2015) but has not been used within high-elevation grasslands of southeastern Africa. We present density estimates based on hierarchical distance sampling models for common bird species in relation to vegetation attributes at Ingula, a high-elevation grassland in eastern South Africa which will soon be declared a nature reserve (Maphisa 2012).
Few studies examine how habitat and management actions affect bird densities at high-elevation grasslands of eastern South Africa. The few studies that exist, examine the effect of burning and grazing on a single species (Parr and Chown 2003) and do not account for the observation process (e.g. Jansen et al. 1999; Little et al. 2013). Burning is the main management tool in agricultural lands and conservation areas in this region (Parr and Chown 2003). Some studies indicate that inappropriate use of fire and grazing leads to degradation of grasslands in this region (Maphisa et al. 2009, 2016; Little et al. 2015a).
We relate grass height and cover in summer to eight small grassland birds that we found to be the most common during preliminary data analysis. Summers coincide with breeding for most birds and is the period with the highest species richness in this area (Maphisa et al. 2016). We used the following eight species: Cape Longclaw (Macronyx capensis), Wing-snapping Cisticola (Cisticola ayresii), African Pipit (Anthus cinnamomeus), African Quail Finch (Ortygospiza atricollis), Yellow-breasted Pipit (Anthus chloris), Red-capped Lark (Calandrella cinerea), Zitting Cisticola (Cisticola juncidis) and Common Quail (Coturnix coturnix). Of these, only the Yellow-breasted Pipit is threatened nationally (Taylor et al. 2015). Conservation managers often rely on surrogates to estimate biodiversity to manage habitat for other species whose density cannot be estimated by distance sampling because they are rare (Nally and Fleishman 2004; Mattsson and Marshall 2009; Lewandowski 2010). Because the suite of species we chose to study represents a range of different habitat requirements in terms of their responses to grass height and cover, we suggest that they can be used as indicators to monitor habitat suitability for rare threatened species which are of conservation concern.
Methods
Study site
The study site is situated c. 23 km northeast (28°14ʹE, 29°35ʹS) of the hamlet town of Van Reenen covered by moist-high elevation grasslands typical of eastern South Africa. It is the site of the Eskom Ingula Pumped Storage Scheme (Ingula or IPSS), the main purpose of which is to augment electricity to the national grid during periods of peak consumption demand (Maphisa et al. 2016).
Ingula has an area of about 8000 ha and falls within the two provinces KwaZulu-Natal (KZN) and the Free State (FS). The average elevation on the FS site is 1700 m asl compared to 1200 m asl on the KZN site. The FS site is dominated by sweet and sour grasslands, while the lower-lying grasslands on the KZN site is dominated by Cymbopogon-Hyrpochloa-Hyperrhinia sp. Of the natural vegetation type on the FS site 39% is exposed to combined land cover threats of degradation and the effects of transformation and roads (Reyers et al. 2001; Mentis 2014), while the KZN site is degraded and mostly invested by alien tree plantations (Maphisa 2015).
The FS side of the study area is viewed as particularly important because of the presence of an Important Bird and Biodiversity Area (SA IBA 043), with the Bedford-Chatsworth Wetland at its center (Barnes 1998; Marnewick et al. 2015). Of the two sites, the lower site is therefore of reduced conservation value and has been used for above-ground infrastructures of the power station. To compensate for the part of the wetland that has been transformed into a storage dam on the FS side, additional land was bought and set aside for conservation. The overall goal of the new management is to declare this area as a nature reserve (Maphisa 2012) with conservation of birds as one of its conservation priorities.
Despite a relatively short distance separating the two sides, there are marked changes in temperature and rainfall (Maphisa et al. 2016). Most of the rainfall occurs during the summer months (October to February) with mean annual amounts of 1000 mm (Mentis 2014). The habitats at Ingula and surrounding grasslands are degraded due to heavy livestock grazing and annual fires in the past (Mentis 2006; Cauldwell 2012). Based on the perceived deterioration of the habitat resulting from years of heavy grazing and annual fires, an impact assessment report (Mentis 2006) recommended that livestock be replaced with game and recommended a minimum fire return period of 2 years. During the summer of 2005/2006, cattle that belonged to private farmers were removed so that the habitat could recover from unsuitable past farming practices. This period coincides with the construction of the Pumped Storage Scheme, the primary purpose for which the land was bought. The Ingula management set aside additional land with the primary aim of managing the area to protect surrounding ecosystems and biodiversity (Maphisa 2015). However, a relatively small number of livestock belonging to tenants, were left behind by previous land owners pending a resettlement plan. The delay to move the tenants to alternative areas has resulted in management challenges to implement burning and grazing regimes to the full as per Mentis (2006) recommendations (Maphisa et al. 2016). Although much reduced, grazing happened everywhere and anytime during the course of our study. Fires of unknown origin consumed most of the study area on an annual basis.
Bird sampling
Using 1:50, 000 topographic maps, we laid random transects of 500 m across the study area perpendicular to farm tracks, separated by 2 km, avoiding too steep, rocky or transects lying adjacent to wetlands. We found 35 transects to be suitable for bird surveys because they had similar topography and were dominantly grassland. We surveyed birds along three fixed 50 m band transects from the center of the transect on both sides of the observer (Bibby et al. 2000; Buckland et al. 2001; Thomas et al. 2010) on all the transects, once per season (winter, spring, summer and autumn) (Maphisa et al. 2016). Strip transects is one of the most commonly used methods to estimate bird abundance, species habitat preferences and species richness for large-scale monitoring programs (e.g. Carrascal et al. 2009). All transect surveys were conducted by one person (DHM), mostly during early mornings (07:00-11:00) or mid-afternoons (15:00-16:00), when birds were most likely to be active (e.g. Maphisa et al. 2009). We extended bird surveys only to afternoons when logistic factors such as unsuitable weather prevented us from completing the survey on time in the mornings. But for such later surveys we selected a narrow time frame (15:00-16:00). We carried out bird surveys under stable weather conditions with good visibility.
In this study, we made use of birds and vegetation surveys for the following three summers: 2006/2007, 2007/2008 and 2010/2011. These periods coincide with the time when most species are on site and breeding (Maphisa et al. 2016). Only seventeen of the original total transects were surveyed during the summer of 2010/2011 due to time constraints. No survey was carried out during the summer of 2009/2010 because DHM was delegated to different types of surveys still on site.
Measurements of vegetation and land cover variables
Vegetation at each transect was surveyed immediately following the bird survey (e.g. Marx et al. 2008), using a 30 cm by 30 cm light metal frame divided into nine equal squares (Maphisa et al. 2009, 2016). The grid was thrown twice (at random) every 100 m along the 500 m transects. At each throw we recorded how many of the nine squares fell on grass, bare soil, dead grass, forbs or stones. Vegetation height was recorded at every corner of the frame, at each throw point using a measuring tape. The four measurements were averaged for data analysis. Intensity of grazing along each transect was categorized independently of measured grass height and cover as none, light or heavily grazed, based on signs of habitat trampling by animals, visual bare ground cover or grass height. Given the variability in fire timing and intensity across the study area, each transect was simply classified into one of two categories: burned or unburned prior to the survey.
Data analysis and fitting the model
Distance sampling is a widely used technique for estimating the density of biological populations (e.g. Bibby et al. 2000; Buckland et al. 2001; Thomas et al. 2005, 2010). Within transect-based distance sampling, the observer records individuals at perpendicular distances away from the line of observation. One of the first steps in the analysis of distance sampling data is modelling the probability of detection (e.g. Thomas et al. 2010). Distance sampling models assume that individuals at zero distance from the line are observed with certainty (Thomas et al. 2010; Weller et al. 2012) and that the chances of detecting an individual decreases with increasing distance away from the observer (Speed et al. 2010; Weller et al. 2012). Improvements in distance sampling offer the opportunity to model both the observation process and density as a function of covariates (Marques et al. 2007; Weller et al. 2012). Rigorous statistical techniques have been developed to account for decreasing probability of detection away from the observer, allowing density or abundance to be estimated more accurately, based on the decrease in the number of observed individuals away from the transect line (Speed et al. 2010; Thomas et al. 2010).
Function 'distsamp' in a package 'unmarked' (Fiske and Chandler 2011) in R version 2.15.1 (R Development Core Team 2013) was used to fit distance sampling models and to estimate the detection and density of the eight common grassland bird species. For each species, a half-normal detection function was used (Fiske and Chandler 2012). This was chosen by visually comparing the distribution of detections against the fitted function. Other commonly used distance functions were tested with similar results. We considered a number of survey variables that could potentially influence detection (e.g. Alexander and Hepp 2014) and how quickly the detection probability declines with increasing distance away from the transect line (Broekema and Overdyck 2012; Weller et al. 2012). These variables were the intensity of grazing along each transect per survey (none, light or heavy), whether there were visible signs of burning that year prior to the survey (burned or unburned), as well as grass height and cover along each transect.
Model selection
The primary use of habitat modelling in conservation planning is to predict the spatial distribution of suitable habitats for species of interest in a landscape (Martin et al. 2010; Wintle et al. 2005). We constructed six competing detection models with the above habitat variables of our interest. The first model assumed detection was similar across all the survey transects (Null model). The second and third models tested the effect of grazing and burning individually. Next we assumed that our ability to detect species could be determined by grass height alone or grass cover alone (fourth and fifth models). Our last model combined grass height and grass cover assuming that both variables might have a combined effect on species detection or use of habitat.
We used model selection, based on Akaike's Information Criterion (AIC) (Burnham and Anderson 2002; Mellin et al. 2010), to choose the best detection model out of the list of six competing models for each species. We considered the model with the lowest AIC value as the best model describing species detection (Taillie et al. 2015; Maphisa et al. 2016). To estimate the effect of grazing and burning on density of each species we used the best detection model above (Additional file 1: Appendix S1) to create a set of eight candidate models (Additional file 2: Appendix S2).
Effect of grass height and cover on density of birds
Our major focus was on how bird densities were related to grass cover and height because these are the two key habitat variables that determine habitat use by grassland birds (e.g. Fisher and Davis 2010). We first considered linear (on a log scale) relationships between density and grass height and cover. Then, we also used regression splines with two knots to fit more flexible relationships and to provide a more accurate prediction of species response to grass height and grass cover. Our implementation of regression splines follows that of Crainiceanu et al. (2005), where a basic function is calculated for each knot; these functions are then included as additive terms in a linear model.
Results
The number of detections (shown in brackets) per species differed amongst species. We found the following species: Cape Longclaw (184), African Pipit (109), Wing-snapping Cisticola (106), Yellow-breasted Pipit (70), Zitting Cisticola (65), Common Quail (60), African Quailfinch (36) and Red-capped Lark (33). For most species, the best detection function depended on habitat variables, implying that grass height and cover affected our ability to detect individual birds (Table 1). In particular, the model that combined grass height plus cover best explained the detection of four out of eight species (Table 1). Correcting for these effects, we then examined how grazing, burning, grass height and grass cover affected the density of the eight species differently (Table 2; Figs. 1, 2, 3, 4, 5, 6).
Table
1.
Summary of the results of model selection analysis to identify the best detection model for eight grassland bird species at Ingula, South Africa
Detection models
CLC
WSC
AP
AQF
YBP
RCL
ZC
CQ
1.
dens(.)p(.)
4.08
7.35
6.00
14.41
9.71
13.36
10.26
1.49
2.
dens(.)p(Burning)
0.00
5.47
7.84
16.37
6.50
2.71
6.68
0.97
3.
dens(.)p(Grazing)
6.41
9.13
9.24
3.81
10.22
0.00
4.80
4.26
4.
dens(.)p(cover)
4.15
8.63
7.58
9.94
10.00
7.34
12.26
2.46
5.
dens(.)p(avh)
6.08
0.00
1.70
16.19
5.13
13.45
4.24
0.00
6.
dens(.)p(avh + cover)
5.37
1.58
0.00
0.00
0.00
6.08
0.00
2.64
The table shows delta AIC values, i.e., the difference in AIC between a particular model and the best. The fitted models are hierarchical distance sampling models and full model selection details are given in Additional file 1: Appendix S1. The best detection model for each species based on lowest delta AIC is highlighted in italics. Density dens(.) was held constant while considering the effects of transect habitat on the detection function p(.). The habitat variables were grass cover ('cover'), grass height ('avh') and whether the area had been grazed ('Grazing') or burned ('Burning') prior to the survey. Species are sorted from the most common (left) to least common (right); Cape Longclaw (CLC), Wing-snapping Cisticola (WSC), African Pipit (AP), African Quailfinch (AQF), Yellow-breasted Pipit (YBP), Red-capped Lark (RCL), Zitting Cisticola (ZC) and Common Quail (CQ)
Table
2.
Summary of model selection analysis (see Additional file 2: Appendix S2 for full details) used to estimate the density of eight common grassland bird species at Ingula in relation to grazing, burning, grass height and grass cover
Density models
CLC
WSC
AP
AQF
YBP
RCL
ZC
CQ
1.
dens(.)
2.72
18.36
26.61
15.93
24.24
8.81
22.78
8.03
2.
dens(Burning)
2.20
0.00
28.40
11.87
0.00
5.86
24.08
7.12
3.
dens(Grazing)
0.34
19.34
30.46
13.56
24.26
6.38
18.26
8.29
4.
dens(cover)
0.11
20.13
24.86
6.03
26.23
8.25
0.00
8.09
5.
dens(avh)
0.00
20.27
11.93
17.83
16.84
5.58
43.39
7.05
6.
dens(avh + cover)
0.55
22.09
0.00
0.00
16.55
0.00
12.60
8.29
7.
dens(s(cover))
1.71
8.24
30.53
4.82
23.17
21.58
24.94
0.00
8.
dens(s(avh))
0.73
3.59
21.01
35.61
22.31
20.08
3.93
6.95
For each species the detection function was modeled as in the best model in Table 1. The relationship between density and grass height and cover was modeled as linear (on the log scale) relationships (avh and cover) and using regression splines with two knots [s(avh) and s(cover)]. See notes to Table 1 for more detail
Figure
1.
Density of the eight most common grassland bird species at Ingula in eastern South Africa, in relation to grazing. Transects were classified as not grazed ('none'), lightly grazed ('light') and heavily grazed ('heavy') based on the three summer surveys carried out from 2006/2008 to 2010/2011. Estimates for density of each species are from Model 3, Table 2. The error bars are standard errors
Figure
2.
Density of the eight most common grassland bird species at Ingula in eastern South Africa, in relation to whether the grass along transects had been burned or not before the survey. Estimates for density of each species are from Model 2, Table 2. The error bars are standard errors
Figure
3.
The effect of average grass height and cover on the habitat suitability of Cape Longclaw and Wing-snapping Cisticola at a high-elevation grassland in eastern South Africa. Light-grey lines are 95% confidence intervals around the fitted response shape (black line) taken from models 7 and 8 in Table 2, while the blue lines are the best-fitting curves based on regression splines with two knots. Density estimates for grass height and cover are from Models 4 and 5 in Table 2
Figure
4.
Effect of average grass height and cover on the habitat suitability of African Pipit and African Quailfinch at a high-elevation grassland of eastern South Africa. See legend to Fig. 3 for details
Figure
5.
Effect of average grass height and cover on the habitat suitability of Yellow-breasted Pipit and Red-capped Lark at a high-elevation grassland in eastern South Africa. See legend to Fig. 3 for details
Figure
6.
Effect of average grass height and cover on the habitat suitability of Zitting Cisticola and Common Quail at a high-elevation grassland of eastern South Africa. See legend to Fig. 3 for details
For Zitting Cisticola, Cape Longclaw, Red-capped Lark and African Quailfinch, the model allowing for different densities between transects that differed in how they were grazed (Model 3, Table 2) was slightly better supported by the data than the constant model (Model 1, Table 2), even though it was not the best supported model for any species. Zitting Cisticolas tended to occur at higher densities on heavily grazed transects than on ungrazed ones (Fig. 1), while the Cape Longclaw occurred at its lowest density on heavily grazed transects (Fig. 1). Red-capped larks occurred in their highest densities on lightly grazed transects where African Quailfinches had the lowest densities (Fig. 1).
For the Wing-snapping Cisticola and Yellow-Breasted Pipit, the model allowing densities to vary between burned and unburned transects (Model 2, Table 2) was the best model, while for Cape Longclaw, African Quailfinch, Red-capped Lark and Common Quail, this model was better than the constant model (Model 1, Table 2). These species were more abundant on burned transects, except for Red-capped Larks, which were more abundant on unburned transects (Fig. 2).
For all species, one of the models involving grass height or cover (Models 4-8, Table 2) was better supported than the constant model (Model 1, Table 2). Model 6, allowing density to vary with both grass height and cover was clearly the best model for the African Pipit, African Quailfinch and Red-capped Lark. For Cape Longclaws Model 5, including only grass height, was the best even though several of the other models were close competitors for this species. For the Zitting Cisticola, the best model was Model 4 assuming a linear effect of grass cover on density. The Common Quail was the only species for which density was best described by a model that included a nonlinear relationship, Model 8, with a regression spline for cover. Overall, all eight species responded differently to the effect of increasing grass height and grass cover (Figs. 3, 4, 5, 6). Of these species, Cape Longclaws apparently benefitted from little grazing with their density increasing with increases in grass height and grass cover (Fig. 3). The Wing-snapping Cisticola, African Quailfinch, Yellow-breasted Pipit, Red-capped Lark and Common Quail densities decreased with increasing grass height and grass cover (Figs. 3, 4, 5, 6). African Pipit densities increased with increasing grass cover (Fig. 4). On the one hand the densities of Zitting Cisticolas increased with an increase in grass cover and Common Quail densities clearly peaked at intermediate levels of grass cover (Fig. 6).
Discussion
Our results support previous findings that sometimes grassland birds co-occur with differing habitat requirements (Maphisa et al. 2009, 2016). Fire and grazing affect aboveground biomass differently (Richardson et al. 2014). Depending on the intensity of fire and grazing, heterogeneity of micro niches is created across the landscape that affects bird species densities differently. The impact of fire and grazing on landscape and habitat attributes has been well documented (Jacobs et al. 2012). Our study is the first in the region to examine how vegetation variables affect the density of birds in a hierarchical way that incorporates detection. Grazing and burning affect species differently depending on their functional groups or the preferences of individual species (Richardson et al. 2014; Mero et al. 2015; Maphisa et al. 2016). Under managed burning and grazing (e.g. Klug et al. 2010; Richardson et al. 2014; Mero et al. 2015) we expected species that prefer short grass to be more abundant on burned transects (e.g. Apfelbaum and Haney 1981) and thereafter with little grazing as it was at Ingula the density was expected to decrease with an increase in grass height. Most of the species we studied exhibited this with only two exceptions: both Cape Longclaws and Zitting Cisticolas increased in density with an increase in grass height (Figs. 3, 6). Because burning and grazing were not managed at our study site, we therefore expected grass height and cover to be better predictors of species density because of the ambiguity of visual classification of grazing in the field. For example, higher densities of Red-capped Larks and Common Quails (Fig. 2) on burned and unburned transects contradict our knowledge of habitat preferences for these two species. The same is true for the Common Quail which occurs in similar densities across the three categories of grazing (Fig. 1).
For three of the eight species we examined, we found that a combination of grass height and cover is more important than just grass height alone or grass cover alone. This agrees with previous findings that fire and burning complement each other to create habitats for birds (e.g. Fuhlendorf et al. 2009). Our study demonstrates that grass height and cover or each one alone, depending on the habitat requirements of individual species, determines habitat use by birds (Winter et al. 2005; Devereux et al. 2006; Donald et al. 2010). Regional studies that link species abundance to habitat do not take into account detection (e.g. Jansen et al. 1999; Little et al. 2013, 2015b). The same is true about the studies from north eastern African high-elevation grasslands (Mamo et al. 2016).
Grasslands habitat in our study area is mostly in its natural state other than for degradation from annual burning and heavy grazing. A similar study from northern Uruguay found that vegetation structure has variable effects on the density of grassland birds too (Azpiroz and Blake 2016). However, this study did not account for variability in detection because of insufficient sample size. To deal with problems of insufficient data some studies estimate relative densities (Post van der Burg et al. 2011). The study by Azpiroz and Blake (2016) mostly resembles our study because it includes natural grasslands, managed with cattle, game or sheep. Overall, Azpiroz and Blake (2016) found that bird numbers were more correlated with grass cover than with grass height in contrast to observations of Henslow's Sparrow (Ammodramus henslowii) at modified Missouri grasslands (e.g. Jacobs et al. 2012). But this is because the comparison of Azpiroz and Blake (2016) was made with croplands where vegetation height tends to be higher but sparse. Also the definition of grass cover for these two studies includes forbs or shrubs. In our study we excluded forbs in our analysis because of sparse forb data.
As expected in our study area, the African Quailfinch indicated steeper density declines with increasing grass cover than with grass height. This species is associated more with croplands than with natural grasslands (Kosicki et al. 2014; Maphisa 2015). Cape Longclaws, the most common species in a group, benefitted from little grazing in our study area with their density rising with increasing grass height and grass cover. Our results contradict Mamo et al. (2014) where a similar but endemic Abyssinian Longclaw (Macronyx flavicollis) occurred with higher densities in more open habitats.
Our results for the African Pipit (Fig. 4), a grassland generalist, agree with our knowledge of how this species reacts to changes in grass height and cover. The rising density of Zitting Cisticolas with increasing grass height agrees with the observation that this species requires high grass to breed (Maphisa 2015). The lowest densities of the threatened Yellow-breasted Pipit on heavily grazed transects (Fig. 1) may explain why this species is rare, given the widespread heavy grazing in the region (Maphisa et al. 2016). On the one hand the same species occurs in much higher densities on transects that were burned (Fig. 2) indicating a need for visual inspection of habitat (e.g. Maphisa et al. 2016). This makes sense given that in the case of our study area there was little grazing. Thick tall grass is worse than heavily grazed grass because of the possible effects of visual obstruction (e.g. Devereux et al. 2006). This is important for ground nesting birds that breed and feed on the ground.
The Common Quail is a regional local migrant with birds easily identified by call and sight early in the breeding season, when birds establish their territories. Birds are easy to see in the field at this time given widespread burning. But it will be harder to see or hear later in the season even when they are present because of increase in grass height and grass cover (Fig. 2) and that bird calls are less frequent. Alternatively, our study site could be a stopover with birds moving to areas that are intensively ploughed. Agricultural fields are a more suitable habitat for this species (Kosicki et al. 2014).
For most species in our study, grazing had no strong effect on density. However, this could be because there was little grazing on site to compare amongst categories of grazing. In contrast, a study with replicated control plots from mountainous grasslands in Norway (Loe et al. 2007) managed with sheep, found that the density of birds was much higher on farms with high sheep densities. But high bird densities on heavily grazed grasslands do not necessarily indicate habitat suitability because this might simply indicate food availability (Loe et al. 2007) but may not support breeding.
Land set aside for conservation in our region is increasing in the form of private reserves and state owned lands. On these lands, we recommend that researchers implement the recommendations by Parr and Chown (2003) regarding experimental research on the effect of fire intensity, timing and variability across the landscape on fauna. A further consideration that needs to be taken into account is the number of replicates compared between sites that are managed differently (e.g. Parr and Chown 2003).
Most species of conservation concern are found on privately owned farms where application of fire and grazing is mostly variable (Howell et al. 2009; Hanley et al. 2012; Jacobs et al. 2012; Ciuzio et al. 2013). In these differently managed lands, we suggest that measurements of vegetation variables be limited to grass height and cover because these are the key habitat variables that are mostly associated with habitat use by birds (Fisher and Davis 2010; Hovick et al. 2014) and doing so also allows survey efforts to cover a large area in a short space of time. Finally we suggest that investigation of factors that affect birds density or suitability be based on surveys from inside the farms, rather than counting along roads (e.g. Wellicome et al. 2014). Habitat on road verges often does not resemble habitat within farms and birds nesting along road verges may be more exposed to predation (Wellicome et al. 2014) and may suffer more from disturbances.
Our study supports previous suggestions that worldwide fire and grazing complement each other to make habitats suitable for birds (Fuhlendorf et al. 2009). However, the most important proximate habitat variables in grasslands are grass height and cover (Fisher and Davis 2010). These two variables can be managed with fire and grazing to improve habitat suitability for grassland bird communities (Fuhlendorf et al. 2009; Maphisa et al. 2009).
Conclusions
Grass height and cover were important predictors of density for all eight common grassland species we examined. However, the species reacted to variation in grass height and cover in different ways. Our results therefore suggest that management should aim to provide a mosaic of patches with different grass height and cover to promote habitat suitability for grassland birds in general in the grasslands of eastern South Africa. Combined use of fire and grazing could be used to create such a mosaic, which is likely to benefit rare species, in addition to the common ones we studied.
Additional files
Additional file 1: Appendix S1. Negative log likelihood (-Loglike), number of parameters (K), Akaike's Information Criterion (AIC), difference in AIC (Delta AIC) and AIC model weights (AICwt). List of candidate models representing alternative ways to model the detection process for eight grassland bird species. For each species, we selected the best detection model based on lowest AIC value (in bold).
Additional file 2: Appendix S2. Negative log likelihood (-Loglike), number of parameters (K), Akaike's Information Criterion (AIC), difference in AIC (Delta AIC) and AIC model weights (AICwt). List of candidate models used to estimate the influence of burning (Burning), grazing (Grazing), grass height (avh) and grass cover (cover) as linear (on the log scale) relationships or regression splines with two knots (s(avh) and s(cover)) on eight grassland bird species. All models used the best structure for the detection function determined during the analysis presented in table in Additional file 1: Appendix S1.
Authors' contributions
DHM conceived and designed the field survey methods and carried out the fieldwork. DHM and RA analysed the data, interpreted the results and wrote the paper. HS proofread and made comments to the manuscript, LG commented on the original manuscript draft. All authors read and approved the final manuscript.
Acknowledgements
The first author was supported in the position of BirdLife South Africa Ingula Project Manager with funding by Eskom through The Ingula Partnership. The Mazda Wildlife Fund supported the first author with a vehicle for the duration of the project, while employed by BirdLife South Africa. We are greatly indebted to Malcolm Drummond for proofreading the original manuscript. RA was supported by the National Research Foundation of South Africa (Grant 85802). The NRF accepts no liability for opinions, findings and conclusions or recommendations expressed in this publication.
Competing interests
The authors declare that they have no competing interests.
Angelier F, Parenteau C, Trouvé C, Angelier N. Does the stress response predict the ability of wild birds to adjust to short-term captivity? A study of the rock pigeon (Columbia livia). R Soc Open Sci. 2016;3:160840.
Bentley PJ. Hormones that influence intermediary metabolism. In: Bentley PJ, editor. Comparative vertebrate endocrinology. Cambridge: Cambridge University Press; 1998. p. 230-8.
Braun EJ, Sweazea KL. Glucose regulation in birds. Comp Biochem Physiol B. 2008;151:1-9.
Breuner CW. Stress and reproduction in birds. In: Norris DO, Lopez KH, editors. Hormones and reproduction of vertebrates. London: Academic Press; 2011. p. 129-51.
Cohen A, Klasing K, Ricklefs R. Measuring circulating antioxidants in wild birds. Comp Biochem Physiol B. 2007;147:110-21.
Corbel H, Geiger S, Groscolas R. Preparing to fledge: the adrenocortical and metabolic responses to stress in king penguin chicks. Funct Ecol. 2010;24:82-92.
Costantini D. Oxidative stress in ecology and evolution: lessons from avian studies. Ecol Lett. 2008;11:1238-51.
Cyr NE, Dickens MJ, Romero LM. Heart rate and heart-rate variability responses to acute and chronic stress in a wild-caught passerine bird. Physiol Biochem Zool. 2009;82:332-44.
Cyr NE, Earle K, Tam C, Romero LM. The effect of chronic psychological stress on corticosterone, plasma metabolites, and immune responsiveness in European starlings. Gen Comp Endocrinol. 2007;154:59-66.
Dallman MF. Stress update: adaptation of the hypothalamic-pituitary-adrenal axis to chronic stress. Trends Endocrinol Met. 1993;4:62-9.
Davies S, Rodriguez NS, Sweazea KL, Deviche P. The effect of acute stress and long-term corticosteroid administration on plasma metabolites in an urban and desert songbird. Physiol Biochem Zool. 2013;86:47-60.
Deviche P, Beouche-Helias B, Davies S, Gao S, Lane S, Valle S. Regulation of plasma testosterone, corticosterone, and metabolites in response to stress, reproductive stage, and social challenges in a desert male songbird. Gen Comp Endocrinol. 2014;203:120-31.
Deviche P, Valle S, Gao S, Davies S, Bittner S, Carpentier E. The seasonal glucocorticoid response of male Rufous-winged Sparrows to acute stress correlates with changes in plasma uric acid, but neither glucose nor testosterone. Gen Comp Endocrinol. 2016a;235:78-88.
Deviche P, Bittner S, Davies S, Valle S, Gao S, Carpentier E. Endocrine, metabolic, and behavioral effects of and recovery from acute stress in a free-ranging bird. Gen Comp Endocrinol. 2016b;234:95-102.
Dickens MJ, Bentley GE. Stress, captivity, and reproduction in a wild bird species. Horm Behav. 2014;66:685-93.
Dickens MJ, Delehanty DJ, Romero LM. Stress: an inevitable component of animal translocation. Biol Conserv. 2010;143:1329-41.
Dickens MJ, Earle KA, Romero LM. Initial transference of wild birds to captivity alters stress physiology. Gen Comp Endocrinol. 2009;160:76-83.
Dickens MJ, Romero LM. Wild European Starlings (Sturnus vulgaris) adjust to captivity with sustained sympathetic nervous system drive and a reduced fight-or-flight response. Physiol Biochem Zool. 2009;82:603-10.
Fischer CP, Wright-Lichter J, Romero LM. Chronic stress and the introduction to captivity: how wild house sparrows (Passer domesticus) adjust to laboratory conditions. Gen Comp Endocrinol. 2018;259:85-92.
Fokidis HB, Hurley L, Rogowski C, Sweazea K, Deviche P. Effects of captivity and body condition on plasma corticosterone, locomotor behavior, and plasma metabolites in curve-billed thrashers. Physiol Biochem Zool. 2011;84:595-606.
Grégoire F, Genart C, Hauser N, Remacle C. Glucocorticoids induce a drastic inhibition of proliferation and stimulate differentiation of adult rat fat cell precursors. Exp Cell Res. 1991;196:270-8.
Guglielmo CG, Cerasale DJ, Eldermire C. A field validation of plasma metabolite profiling to assess refueling performance of migratory birds. Physiol Biochem Zool. 2005;78:116-25.
Guglielmo CG, O'Hara PD, Williams TD. Extrinsic and intrinsic sources of variation in plasma lipid metabolites of free-living western sandpipers (Calidris mauri). Auk. 2002;119:437-45.
Hazelwood RL. The intermediary metabolism of birds. In: Farner DS, King JR, editors. Avian biology. New York: Academic Press; 1972. p. 472-526.
Hershock D, Vogel WH. The effects of immobilization stress on serum triglycerides, nonesterified fatty acids, and total cholesterol in male rats after dietary modifications. Life Sci. 1989;45:157-65.
Jenni-Eiermann S, Jenni L, Kvist A, Lindström Å, Piersma T, Visser GH. Fuel use and metabolic response to endurance exercise: a wind tunnel study of a long-distance migrant shorebird. J Exp Biol. 2002;205:2453-60.
Klasing KC. Metabolism and storage of triglycerides. In: Klasing KC, editor. Comparative avian nutrition. New York: Centre for Agriculture and Bioscience International; 1998. p. 182-94.
Landys MM, Ramenofsky M, Guglielmo CG, Wingfield JC. The low-affinity glucocorticoid receptor regulates feeding and lipid breakdown in the migratory Gambel's white-crowned sparrow Zonotrichia leucophrys gambelii. J Exp Biol. 2004;207:143-54.
Landys MM, Ramenofsky M, Wingfield JC. Actions of glucocorticoids at a seasonal baseline as compared to stress-related levels in the regulation of periodic life processes. Gen Comp Endocrinol. 2006;148:132-49.
Lattin CR, Bauer CM, de Bruijn R, Romero LM. Hypothalamus-pituitary-adrenal axis activity and the subsequent response to chronic stress differ depending upon life history stage. Gen Comp Endocrinol. 2012;178:494-501.
Li D. Birds as pathology-free models of type Ⅱ diabetes. Austin Endocrinol Diabetes Case Rep. 2017;2:1007.
Li D, Wang G, Wingfield JC, Zhang Z, Ding C, Lei F. Seasonal changes in adrenocortical responses to acute stress in Eurasian tree sparrow (Passer montanus) on the Tibetan Plateau: comparison with house sparrow (P. domesticus) in North America and with the migratory P. domesticus in Qinghai Province. Gen Comp Endocrinol. 2008;158:47-53.
Li D, Wu J, Zhang X, Ma X, Wingfield JC, Lei F, Wang G, Wu Y. Comparison of adrenocortical responses to acute stress in lowland and highland Eurasian tree sparrows (Passer montanus): similar patterns during the breeding, but different during the prebasic molt. J Exp Zool. 2011;315A:512-9.
Li D, Zhang X, Li Y, Hao C, Zhang J, Wu Y. Stress responses of testosterone and corticosterone-binding globulin in a multi-brooded species, Eurasian Tree Sparrows (Passer montanus): does CBG function as a mediator? Horm Behav. 2012;61:582-9.
Li M, Sun Y, Wu J, Zhang X, Li J, Yao Y, Liu X, Li D, Wu Y. Variation in corticosterone response and corticosteroid binding-globulin during different breeding sub-stages in Eurasian tree sparrow (Passer montanus). J Exp Zool. 2016;325A:75-83.
Lynn SE, Stamplis TB, Barrington WT, Weida N, Hudak CA. Food, stress, and reproduction: short-term fasting alters endocrine physiology and reproductive behavior in the zebra finch. Horm Behav. 2010;58:214-22.
Mason G, Burn CC, Dallaire JA, Kroshko J, Kinkaid HM, Jeschke JM. Plastic animals in cages: behavioural flexibility and responses to captivity. Anim Behav. 2013;85:1113-26.
Mason GJ. Species differences in responses to captivity: stress, welfare and the comparative method. Trends Ecol Evol. 2010;25:713-21.
Morgan KN, Tromborg CT. Sources of stress in captivity. Appl Anim Behav Sci. 2007;102:262-302.
Mori JG, George JC. Seasonal changes in serum levels of certain metabolites, uric acid and calcium in the migratory Canada goose (Branta canadensis interior). Comp Biochem Physiol B. 1978;59:263-9.
Muldoon MF, Kritchevsky SB, Evans RW, Kagan VE. Serum total antioxidant activity in relative hypo-and hypercholesterolemia. Free Radic Res. 1996;25:239-45.
Orgebin-Crist MC, Tichenor PL. Effect of testosterone on sperm maturation in vitro. Nature. 1973;245:328-9.
Partonen T, Haukka J, Virtamo J, Taylor PR, Lonnqvist J. Association of low serum total cholesterol with major depression and suicide. Br J Psychiatry. 1999;175:259-62.
Rich EL, Romero LM. Exposure to chronic stress downregulates corticosterone responses to acute stressors. Am J Physiol Reg I. 2005;288:R1628-36.
Romero LM. Seasonal changes in plasma glucocorticoid concentrations in free-living vertebrates. Gen Comp Endocrinol. 2002;128:1-24.
Romero LM, Wingfield JC. Alterations in hypothalamic-pituitary-adrenal function associated with captivity in Gambel's white-crowned sparrows (Zonotrichia leucophrys gambelii). Comp Biochem Physiol B. 1999;122:13-20.
Sapolsky RM, Romero LM, Munck AU. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr Rev. 2000;21:55-89.
Sautin YY, Johnson RJ. Uric acid: the oxidant-antioxidant paradox. Nucleosides Nucleotides Nucl. 2008;27:608-19.
Schroepfer GJ. Oxysterols: modulators of cholesterol metabolism and other processes. Physiol Rev. 2000;80:361-554.
Shmueli M, Izhaki I, Zinder O, Arad Z. The physiological state of captive and migrating Great White Pelicans (Pelecanus onocrotalus) revealed by their blood chemistry. Comp Biochem Physiol A. 2000;125:25-32.
Stinefelt B, Leonard SS, Blemings KP, Shi X, Klandorf H. Free radical scavenging, DNA protection, and inhibition of lipid peroxidation mediated by uric acid. Ann Clin Lab Sci. 2005;35:37-45.
Strazzullo P, Puig JG. Uric acid and oxidative stress: relative impact on cardiovascular risk. Nutr Metab Cardiovasc. 2007;17:409-14.
Summers-Smith D. Eurasian tree sparrow (Passer montanus). In: del Hoyo J, Elliott A, Sargatal J, Christie DA, de Juana E, editors. Handbook of the birds of the world. Barcelona: Lynx Edicions; 2014.
Tift MS, Houser DS, Crocker DE. High-density lipoprotein remains elevated despite reductions in total cholesterol in fasting adult male elephant seals (Mirounga angustirostris). Comp Biochem Physiol B. 2011;159:214-9.
Tsahar E, Arad Z, Izhaki I, Guglielmo CG. The relationship between uric acid and its oxidative product allantoin: a potential indicator for the evaluation of oxidative stress in birds. J Comp Physiol B. 2006;176:653-61.
Vegiopoulos A, Herzig S. Glucocorticoids, metabolism and metabolic diseases. Mol Cell Endocrinol. 2007;275:43-61.
Wingfield JC, Kitaysky AS. Endocrine responses to unpredictable environmental events: stress or anti-stress hormones? Integr Comp Biol. 2002;42:600-9.
Wingfield JC, Maney DL, Breuner CW, Jacobs JD, Lynn S, Ramenofsky M, Richardson RD. Ecological bases of hormone-behavior interactions: the "emergency life history stage". Am Zool. 1998;38:191-206.
Wingfield JC, O'Reilly KM, Astheimer LB. Modulation of the adrenocortical responses to acute stress in arctic birds: a possible ecological basis. Am Zool. 1995;35:285-94.
Wingfield JC, Vleck CM, Moore MC. Seasonal changes of the adrenocortical response to stress in birds of the Sonoran Desert. J Exp Zool. 1992;264:419-28.
Yamada F, Inoue S, Saitoh T, Tanaka K, Satoh S, Takamura Y. Glucoregulatory hormones in the immobilization stress-induced increase of plasma glucose in fasted and fed rats. Endocrinology. 1993;132:2199-205.
Zhao Y, Gong L, Zhao B, Gao X, Wu Y, Li D. Life-history dependent relationships between plasma alkaline phosphatase activity and body condition in male Eurasian Tree Sparrows. Avian Res. 2017a;8:18.
Zhao Y, Li M, Sun Y, Wu W, Kou G, Guo L, Xing D, Wu Y, Li D, Zhao B. Life-history dependent relationships between body condition and immunity, between immunity indices in male Eurasian tree sparrows. Comp Biochem Physiol A. 2017b;210:7-13.
David J. T. Douglas, Jessica Waldinger, Zoya Buckmire, et al. A global review identifies agriculture as the main threat to declining grassland birds. Ibis, 2023.
DOI:10.1111/ibi.13223
2.
Monday V. Mdluli, Zamekile D. Bhembe, Leslie Brown, et al. The loss of vegetation cover has distinct but short-term impact on multiple vertebrate taxa in a grassland ecosystem. Global Ecology and Conservation, 2022, 38: e02198.
DOI:10.1016/j.gecco.2022.e02198
3.
Kimani Dominic, Muchane Muchai, Johnstone Kimanzi, et al. Habitat structure determines the abundance of the Endangered Sharpe’s Longclaw Macronyx sharpei (Aves: Passeriformes: Motacillidae) at Timau montane grasslands in central Kenya. Journal of Threatened Taxa, 2020, 12(5): 15565.
DOI:10.11609/jott.5366.12.5.15565-15571
4.
David H. Maphisa, Hanneline Smit-Robinson, Res Altwegg. Dynamic multi-species occupancy models reveal individualistic habitat preferences in a high-altitude grassland bird community. PeerJ, 2019, 7: e6276.
DOI:10.7717/peerj.6276
5.
Masatoshi Takeuchi. Winter bird communities in the heterogeneous farmlands of the Aso region in Japan. Journal of Asia-Pacific Biodiversity, 2019, 12(2): 152.
DOI:10.1016/j.japb.2019.01.013
6.
Sulaiman I Muhammad, Rosli Ramli, Amy Y Then. Seasonality, habitat type and locality influenced bird assemblage structure in Nigeria. Ostrich, 2018, 89(3): 221.
DOI:10.2989/00306525.2018.1425927
7.
Thomas Oliver Mérő, Antun Žuljević, Katalin Varga, et al. Reed management influences philopatry to reed habitats in the Great Reed Warbler (Acrocephalus arundinaceus). The Condor, 2018, 120(1): 94.
DOI:10.1650/CONDOR-17-114.1
Table
1.
Summary of the results of model selection analysis to identify the best detection model for eight grassland bird species at Ingula, South Africa
Detection models
CLC
WSC
AP
AQF
YBP
RCL
ZC
CQ
1.
dens(.)p(.)
4.08
7.35
6.00
14.41
9.71
13.36
10.26
1.49
2.
dens(.)p(Burning)
0.00
5.47
7.84
16.37
6.50
2.71
6.68
0.97
3.
dens(.)p(Grazing)
6.41
9.13
9.24
3.81
10.22
0.00
4.80
4.26
4.
dens(.)p(cover)
4.15
8.63
7.58
9.94
10.00
7.34
12.26
2.46
5.
dens(.)p(avh)
6.08
0.00
1.70
16.19
5.13
13.45
4.24
0.00
6.
dens(.)p(avh + cover)
5.37
1.58
0.00
0.00
0.00
6.08
0.00
2.64
The table shows delta AIC values, i.e., the difference in AIC between a particular model and the best. The fitted models are hierarchical distance sampling models and full model selection details are given in Additional file 1: Appendix S1. The best detection model for each species based on lowest delta AIC is highlighted in italics. Density dens(.) was held constant while considering the effects of transect habitat on the detection function p(.). The habitat variables were grass cover ('cover'), grass height ('avh') and whether the area had been grazed ('Grazing') or burned ('Burning') prior to the survey. Species are sorted from the most common (left) to least common (right); Cape Longclaw (CLC), Wing-snapping Cisticola (WSC), African Pipit (AP), African Quailfinch (AQF), Yellow-breasted Pipit (YBP), Red-capped Lark (RCL), Zitting Cisticola (ZC) and Common Quail (CQ)
Table
2.
Summary of model selection analysis (see Additional file 2: Appendix S2 for full details) used to estimate the density of eight common grassland bird species at Ingula in relation to grazing, burning, grass height and grass cover
Density models
CLC
WSC
AP
AQF
YBP
RCL
ZC
CQ
1.
dens(.)
2.72
18.36
26.61
15.93
24.24
8.81
22.78
8.03
2.
dens(Burning)
2.20
0.00
28.40
11.87
0.00
5.86
24.08
7.12
3.
dens(Grazing)
0.34
19.34
30.46
13.56
24.26
6.38
18.26
8.29
4.
dens(cover)
0.11
20.13
24.86
6.03
26.23
8.25
0.00
8.09
5.
dens(avh)
0.00
20.27
11.93
17.83
16.84
5.58
43.39
7.05
6.
dens(avh + cover)
0.55
22.09
0.00
0.00
16.55
0.00
12.60
8.29
7.
dens(s(cover))
1.71
8.24
30.53
4.82
23.17
21.58
24.94
0.00
8.
dens(s(avh))
0.73
3.59
21.01
35.61
22.31
20.08
3.93
6.95
For each species the detection function was modeled as in the best model in Table 1. The relationship between density and grass height and cover was modeled as linear (on the log scale) relationships (avh and cover) and using regression splines with two knots [s(avh) and s(cover)]. See notes to Table 1 for more detail

 DownLoad:
DownLoad:











 Email Alerts
Email Alerts RSS Feeds
RSS Feeds