Johann H van Niekerk. 2014: Population structure and mating system of the Red-crested Korhaan (Lophotis ruficrista) in South Africa. Avian Research, 5(1): 9. DOI: 10.1186/s40657-014-0009-3
Citation:
Johann H van Niekerk. 2014: Population structure and mating system of the Red-crested Korhaan (Lophotis ruficrista) in South Africa. Avian Research, 5(1): 9. DOI: 10.1186/s40657-014-0009-3
Johann H van Niekerk. 2014: Population structure and mating system of the Red-crested Korhaan (Lophotis ruficrista) in South Africa. Avian Research, 5(1): 9. DOI: 10.1186/s40657-014-0009-3
Citation:
Johann H van Niekerk. 2014: Population structure and mating system of the Red-crested Korhaan (Lophotis ruficrista) in South Africa. Avian Research, 5(1): 9. DOI: 10.1186/s40657-014-0009-3
Department of Environmental Sciences, College of Agriculture and Environmental Sciences, University of South Africa, PO Box 392, Pretoria 0003, South Africa
The mating system of the korhaans and bustards in southern Africa is either based on polygyny or monogamy. The Red-crested Korhaan (Lophotis ruficrista) has been described as polygynous but otherwise very little is known about its breeding biology. The aims were to describe the population structure and male behavior during breeding.
Methods
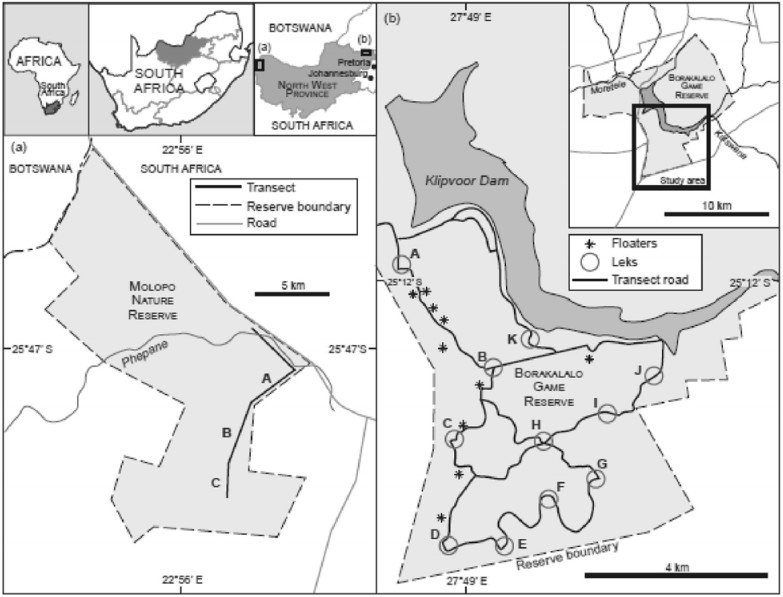
The data collected for this paper was mainly based on field transect surveys carried out in 2009, 2010 and 2011. The population structure of the Red-crested Korhaan in the temperate Borakalalo Game Reserve (BGR) was compared with that in the arid Molopo Nature Reserve (MNR) (both situated in the North West province of South Africa). The study was mainly conducted in the BGR but additional work was conducted in the MNR for comparison.
Results
The difference in mean group sizes between MNR (1.03) and BGR (1.07) was statistically not significant. Group sizes were not affected by climate despite the fact that the BGR received about 650 mm precipitation per annum and the MNR about 200 mm. In both reserves the apparent sex ratios were skewed in favor of males (1:0.29 in the BGR and 1:0.1 in the MNR). The population was dispersed in a clumped manner which is ascribed to the formation of leks. Leks were positioned in open habitat while females invariably concealed themselves under cover.
Conclusions
The leks formed the pivot of the mating system of the Red-crested Korhaan and are identifiable social structures that field ecologists can use to monitor population stability.
Breeding seasons in birds can occur at the same time each year (predictable) or at different times (opportunistic), which depends on food resources. Its variability results from the differences within and between species, latitude or the type of food. Also various local prognostic indicators, including climatic conditions, can influence the photoperiodically induced gonadal development rate and the time to start breeding (Caro et al., 2007). On the one hand, some tropical birds hardly depend on the climate or food resources during the year, and on the other hand, species breeding in the polar regions have a very predictable breeding season and are exposed to short-term unfavorable weather changes (Dawson, 2008). In all cases, there is evidence for involvement of photoperiodic control, nonphotoperiodic control, and endogenous circannual rhythmicity, but in predictable breeders (most nontropical species), photoperiod is the predominantly proximate factor which influences the behavior related to the nesting period, e.g. territory formation, courtship, pair bonding, and nest building (Dawson et al., 2001; Wingfield, 2005; Dawson, 2008).
The time of the start of breeding is one of the most important factors influencing breeding success and, in consequence, the size of the breeding population (Dunn and Møller, 2014; McLean et al., 2016). Courtship and nest construction are the first stage of the breeding performance and can be an indicator of the beginning time of the breeding season (Collias and Collias, 1984). The construction of the nest is a costly challenge for birds and is related to high energy expenditure (Hansell and Overhill, 2000; Mainwaring and Hartley, 2013). In the case of a colony nesting species, this is an effect of intragroup factors like the competition for nest sites, nests, and nest materials (aggressive behavior), the availability and location of food sources, predators pressure and weather (e.g. low temperature or strong wind) (Collias and Collias, 1984). Under temperate climate conditions the optimal period for nest construction should be in the time when the average temperature of the day is relatively high. This can be quite variable each year (Górniak, 2021); therefore, plasticity in response to changes in climate features can play an important role, and can predict population trends (Charmantier et al., 2008; Hałupka and Hałupka, 2017; Mason et al., 2019). In the Rook (Corvus frugilegus), early arrival in the breeding colony (mid-February) in Central Europe, results in quite a long time of occupation before the eggs are laid (egg laying peak is in the third decade of March and the first decade of April; Kasprzykowski, 2002; Madge, 2020). Nest is built by both sexes, male bringing in material and female undertaking most of construction work, both sexes constructing and shaping the cup, work taking 1–4 weeks (Madge, 2020). Because building a nest is time-consuming in this species (up to a month), Rooks should adjust the time of nest building to favorable temperature conditions. This behavior should be especially noticeable in early spring when air temperatures often fall below zero degrees Celsius. Moreover, rapid deterioration in weather conditions which often occurs in the early spring, like strong winds, can collapse nests, chiefly on the first stage of building. In order to avoid higher energy expenditure in these unfavorable weather conditions, rooks should extend the time of building nests, so the reaction to unfavorable weather conditions should be visible.
We hypothesize that the dynamic development of the breeding colony depends on the length of the day, because photoperiodism is an important selection factor related to the behavior of birds, especially in nontropical species (Lack, 1950; Dawson et al., 2001; Sharp, 2005). Due to the long period of the nest building phase of the Rook, we consider weather conditions such as minimum air temperature (see Busse, 1965), snowfall, rainfall or strong wind to be other factors influencing the time of nest construction. The aim of this study was to find relationships between the number of nests under construction in a colony of Rooks and weather conditions, and day length.
2.
Methods and materials
2.1
Study area
The research was conducted in a large city in north-eastern Poland (Białystok; 53°07ʹ N, 23°10ʹ E; 102 km2). It is situated on the western edge of the macroregion of the Białystok Upland, within the North Podlasie Plain, on the banks of the Biała River (Kondracki, 2013). It is characterized by a varied relief. Population density is 2.9 people per km2. The city has a warm summer continental climate, characterized by high temperatures during summer and long and frosty winters. The region is one of the coldest in Poland, with the average temperature in January being −3.4 ℃ and the average temperature over a year of 7.5 ℃. Mean annual rainfall values oscillate around 600 mm and the vegetation period lasts 203 days and snow cover stays 72 days (Górniak, 2021). Almost the entire city lies in the catchment area of the Biała River. Many semi-natural and natural ecosystems exist near urban environments inside the city limits. These natural areas occupy ca 3600 ha (about 32% of the city area), and over half of these (ca 18%) are forest complexes and parks (Kwiatkowski and Gajko, 2011).
The studied Rook colony is one of the biggest rookeries in Białystok (Zbyryt and Polakowski, 2012; Zbyryt et al., 2013) located in the central part of the city between the Biała River and one of the main roads (53°08ʹ05.2ʺ N, 23°09ʹ53.7ʺ E). The tree stand is mostly composed by alien species, Canadian poplars Populus × canadensis which are used by Rooks for nest construction. The trees grow here in a row along the river.
2.2
Fieldwork
The research was conducted in 2015–2017 from mid-February, as this is the time when first birds usually appear in the breeding colony. The nests were counted every five days between 15 February and 15 May at the same time starting between 7.30 and 8.00 a.m. In each count, the total number of nests was recorded. The old nests from previous years found in the beginning of the season were incorporated into the total number of nests in the colony. They are quite durable structures, therefore all nests from the previous breeding season are occupied and expanded by birds in the following year. A Rook nest is a fairly wide and deep cup made of sticks and lined with blades of grass, into which are laid 2–7 eggs; nests are built high on the top of trees (Cramp and Perrins, 1994; Kasprzykowski, 2008; Madge, 2020). The nest was considered to be under construction when consisting of at least several sticks and this is the first stage in the construction of the nest, which takes about 3 days.
2.3
Statistical analysis
Because the same study period was used in all years, data from all three seasons were combined. For the analysis of factors affecting the number of nests in the colony, a generalized linear model (Generalized Least Squares) was used, in which the number of nests in the pentad (five days period) was the response variable and the explanatory variables were: (1) average temperature, (2) maximum temperature (3) minimum temperature, (4) the total rainfall within five days, (5) the highest value of permanent wind recorded within five days, (6) year and (7) day-length. Parameters (1), (2) and (3) were averages over a five-day period. Due to the distribution of the data, polynomial matching was used, in which the length of the day was also used as a square and cubic factor. All continuous variables have been logarithmically transformed. To evaluate the differences between the three years of study we applied Tukey's HSD post hoc test. Meteorological data for the city of Bialystok came from www.en.tutiempo.net. Data on day-length were calculated based on the Astronomy Lab 2 program for Windows operating systems (www.ericbt.com/astronomylab2). We checked models for all combination of factors, including day length as squared and cubic factor. We selected models based of AICc, but also did check for model fit using R squared. Figure was prepared in R using ggplot.
Initially, we used the proportion of nests in the colony or the pace of nest building (calculated as the ratio of the number of nests of the next pentad to the number of nests of the previous pentad) as the dependent variable, but the results were not different from those obtained for the number of nests. The only difference was that the proportion of nests built in consecutive pentads and pace of nest building was not affected by the year. Thus considering the problems with the appropriate model when the dependent variable in a regression model is a proportion (Long, 1997) we decided to present the results only for the number of nests.
3.
Results
Nest construction started on February 20 and was finished at April 16. The maximum number of nests under construction was observed in the first half of April. The total number of nests varied significantly between successive years of research from 63 in 2016 to the highest number 104 observed in 2015. In the three years of research, the number of old nests (from previous breeding season) in the colony was, respectively, 26, 26 and 20.
The results of the analyses showed that no climatic factors affected the number of nests in the colony. The only factor responsible for the increase in the number of nests was the day length (Table 1). The increase of nests in the colony was best described by a third-degree polynomial equation (Fig. 1). The effect of year was significant (Table 1), but the significant differences were found only between 2015 and 2016 (Tukey test, z = 7.31, p < 0.001) and 2015 and 2017 (Tukey test, z = 5.39, p < 0.001), but not between 2016 and 2017 (Tukey test, z = 1.74, p = 0.08).
Table
1.
The effects of day length, year and climatic factors on the number of nests in the Rook colony.
For the sake of clarity we presented the effect of the day length on the proportion of the nests built in each pentad rather than on the absolute number of nests, as this way we removed the differences among years (Fig. 1). Thus it also shows that the pace of nest building was almost identical in all study years.
4.
Discussion
Contrary to what we assumed, only one factor, the lengthening of the day, was of key importance in the dynamics of Rook breeding colony development. This is related to photoperiodism, which regulates various periodic events in bird life. Photoperiod is the primary cue used to initiate and regulate the physiological and behavioral cascades for most seasonal events in birds (Dawson, 2008). It causes hormonal changes that contribute to the development of gonads, which facilitates further mating behavior, such as display, pairing, nesting and laying eggs (Farner, 1964; Dawson et al., 2001; Coppack and Pulido, 2004). In an example of the White Stork (Ciconia ciconia), it was shown that mean daily foraging duration increased with increasing day length in the early part of the season (Podlaszczuk et al., 2015). In the case of the Rook, this factor may also be adaptive, as it is associated with a seasonal increase in food resources, mainly associated with farm work such as plowing, sowing and mowing. Early breeding guarantees access to food resources for hatching chicks and later that the young after leaving the nest will have access to open areas with low vegetation, which facilitates foraging (Kasprzykowski, 2007), because the main source of food for this species in the growing season is invertebrates, mostly earthworms (Madge, 2020). With the passage of the growing season, the vegetation becomes progressively taller thereby reducing the feeding efficiency of fledglings, which can contribute to their lower survival. Therefore, regardless of the conditions, increasing day length can also be a strong selection factor for Rooks.
Our results differ from the earlier observations of Busse (1965), who in an example of a rookery in Warsaw, showed that the increase in the number of nests was correlated with the increasing minimum temperature of the day. The phenomenon that we observed may be a result of desynchronization of photoperiodism and temperature-dependent selective environmental factors (Coppack and Pulido, 2004). The global increase in temperature entails a number of adverse changes, which include, among others, shifts of seasons, changes in the range of species and earlier arrival and breeding (Tryjanowski et al., 2002; Charmantier et al., 2008; Rather et al., 2018). However, when we compared the average monthly temperatures from February to May for 1960–1962, the period coinciding with the Busse (1965) study, with the same period during the present study (2015–2017), the difference was not statistically significant (t = −0.0955; df = 23; p = 0.4622). The lack of significant differences between these periods seems to indicate that in the case of the Rook colony studied here temperature never played a key role in its formation and the dynamics of nest construction. This is further confirmed by the fact that the length of the day is the most important factor in this process. It seems that environmental factors, such as weather (e.g. temperature, rainfall, snowfall) or food abundance affect the timing of life-cycle stages, but provide only short-term predictive information (Coppack and Pulido, 2004). However, these factors play a minor role in the development of gonads and for fine-tuning timing of breeding with local phenological conditions. In conditions where many factors are unpredictable, such as changing spring weather in a temperate climate, the length of the day provides much more reliable information (Wingfield et al., 1992). On the other hand, in times of global warming, relying on only one factor, which is not subject to change over many generations, can be maladaptive in these circumstances. Despite the absence of significant differences in the average temperatures of the late winter and spring months compared above, it is known that in recent time spring phases have advanced up to four weeks towards an earlier start in breeding in Central Europe (Ahas et al., 2002). The latest research indicates that it is two weeks in the region where we conducted our research (Górniak, 2021). Most species of birds feed their young on specific food resources and so breeding is restricted to the time when the required food resource is sufficiently available (Dawson et al., 2001). Lack of time to fine-tune the timing of breeding to the early arrival of spring may cause peaks in some food resources to be missed or make it unavailable (e.g. by tall vegetation). In the case of the Dutch Pied Flycatcher (Ficedula hypoleuca) population, it was shown that the hatching time of the chicks was mismatched with the peaks in the number of caterpillars, which has caused a decline of about 90% over the past two decades (Both et al., 2006).
At the beginning of the breeding period, the Rooks in the present study make long flights to feeding grounds outside the city, but from the beginning of May they feed only within the city boarders (own obs.). This may suggest the lack of suitable feeding places in crop fields and meadows, where the vegetation at this time is already too high for optimal feeding (Kasprzykowski, 2003). This may be partly due to the late mowing of the meadows (after June 15th) or the complete cessation of their use (Zbyryt et al., 2013). Currently, the Rook in most areas of the country nests almost exclusively in urban areas – in western Poland, all colonies are located in highly urbanized areas, while in north-eastern Poland, where the breeding population of the Rook is the highest, only a few are located outside the cities (Zbyryt et al., 2013, 2018; Zbyryt, 2014; Jerzak et al., 2017) – which may be adaptive behavior arising from these difficulties in access to food in the critical period when chicks are hatching. Regularly mown green areas in cities may provide a substitute for mowed meadows and pastures in rural areas. In addition, people in cities feed birds all year round. However, in the case of Rook chicks this food is unsuitable as they mainly require invertebrates (Madge, 2020).
During our research, the first new nest was built at the earliest on February 20th, while in Warsaw in the mid-twentieth century the earliest date was March 8th. In both cases, after its formation, the number of nests did not usually increase for a long time. In Warsaw in the mid-twentieth century, nest numbers grew rapidly only from the second decade of March, and in Bialystok in the 21st century, each year rapid growth began at exactly the same time, at the end of the first decade of March (Busse, 1965). The peak in the number of nests in the colony in the Warsaw study occurred in the second decade of April. In our case, despite the passage of over 60 years from Busse's (1965) studies, the peak occurred in this same period. This proves that despite the clear shift in the start of spring by two weeks in recent three decades (Górniak, 2021), the Rook has not responded to this phenomenon by earlier nesting. Conservative attachment to the length of the day, which is the main factor responsible for the development of the breeding colony, makes the Rook appear to be a species with low plasticity in the face of rapid climate changes. There is evidence that some other species, even those with flexibility such as the Great Tit (Parus major), rely on photoperiod sensitivity which is not constraining the advancement of laying date under current environmental conditions (Salis et al., 2019). On the other hand, in many bird species a relationship between spring temperature and timing of breeding has been observed (Mitrus, 2003; Dunn and Winkler, 2010; Martin et al., 2019). However, in the case of the Rook, the photoperiod may play the most important role in regulation of breeding phenology, with indirect confirmation (nests must be built before egg laying) expressed in the lack of adjustment of the dynamics of breeding colony development to the current climate conditions. We suggest that this may be one of the reasons responsible for the decline in the number of breeding pairs of the Rook in Europe (BirdLife International, 2015). In Poland despite Rook protection since 1952, a decrease in the population size has been observed, especially in the last 20 years (Chylarecki et al., 2018). The fall in the Rooks population index, during the period of national monitoring in the years 2001–2016, averaged 4% per year, which qualifies it as near threatened according to IUCN criteria. In the east of the country, i.e. the region where we conducted our research, the downward trend in numbers of breeding pairs in this period was even bigger, as much as 10% per year (Chylarecki et al., 2018). Due to human-nature conflicts in many cities in Poland the Rook's colonies are destroyed after breeding season and birds are to scare away from nesting sites (Orłowski and Czapulak, 2007). However, this does not explain the decline in the breeding population of the Rook in Poland, and our research can be an important clue to solving the causes of this phenomenon. Therefore, we recommend a broader and wider research on this issue, to confirm or falsify this hypothesis.
5.
Conclusions
The photoperiod seems to be the most important factor influencing breeding behavior (among nontropical species), including nest construction in the Rook. Attachment to the length of the day makes the Rook appear to be a species with low plasticity in the face of rapid climate changes. Simultaneously, in the last 20 years a rapid decline in the number of breeding pairs of this species in Poland has been observed. Thus, we suggest that low behavioral plasticity to changes in spring temperature may indirectly influence the decrease the breeding population of the Rook. More research is needed to explain this phenomenon.
Author contributions
AZ designed the study, carried out field work and wrote the draft manuscript; KZ performed statistical analyses. CM revised and improved the manuscript. All authors read and approved the final manuscript.
Declaration of competing interest
The authors declare that they have no competing interests.
Allan DG (2005a) Northern Black Korhaan. In: Hockey PAR, Dean WJR, Ryan PG (eds) Roberts Birds of Southern Africa, 7th edn. Trustees of the John Voelcker Bird Book Fund, Cape Town, pp 298–300
Allan DG (2005b) Karoo Korhaan. In: Hockey PAR, Dean WJR, Ryan PG (eds) Roberts Birds of Southern Africa, 7th edn. Trustees of the John Voelcker Bird Book Fund, Cape Town, pp 301–302
Allan DG (2005c) Blue Korhaan. In: Hockey PAR, Dean WJR, Ryan PG (eds) Roberts Birds of Southern Africa, 7th edn. Trustees of the John Voelcker Bird Book Fund, Cape Town, pp 302–304
Allan DG (2005d) Ludwig's Bustard. In: Hockey PAR, Dean WJR, Ryan PG (eds) Roberts Birds of Southern Africa, 7th edn. Trustees of the John Voelcker Bird Book Fund, Cape Town, pp 293–294
Allan DG (2005e) Red-crested Korhaan. In: Hockey PAR, Dean WJR, Ryan PG (eds) Roberts Birds of Southern Africa, 7th edn. Trustees of the John Voelcker Bird Book Fund, Cape Town, pp 296–297
Allan DG (2005f) Kori Bustard. In: Hockey PAR, Dean WJR, Ryan PG (eds) Roberts Birds of Southern Africa, 7th edn. Trustees of the John Voelcker Bird Book Fund, Cape Town, pp 295–296
Allan DG, Anderson MD (2010) Assessment of the Threats Faced by South Africa's Bustard Species. Bustard Working Group, BirdLife South Africa
Bradbury TW (1981) The evolution of leks. In: Alexander RD, Tinkle DW (eds) Natural Selection and Social Behaviour. Chiron Press, New York and Concord, pp 138–169
Broders O, Osborne T, Wink M (2003) A mtDNA phylogeney of bustards (family Otididae) based on nucleotide sequences of the cytochrome b-gene. J Ornithol 144:176–185
Catchpole CK, Slater JB (2008) Bird Song: Biological Themes and Variation. Cambridge University Press, United Kingdom
Chase JM, Peter A, Abrams PA, Grover JP, Diehl PS, Robert C, Shane H, Richards DA, Nisbet RM, Case TJ (2002) The interaction between predation and competition: a review and synthesis. Ecol Let 5:302–315
Chittenden H, Allan D, Weiersbye I (2012) Roberts Geographic Variation of Southern African Birds. John Voelcker Bird Book Fund, Cape Town
Constantine M (2013) The Sound Approach to Birding. The Sound Approach, Dorset
Daniel JC, Blumstein DT (1998) A test of the acoustic adaptation hypothesis in four species of marmot. Anim Behav 56:1517–1528
Green RE (2004) Breeding Biology. In: Sutherland WJ, Newton I, Green RE (eds) Bird Ecology and Conservation. Oxford University Press, Oxford, pp 57–83
Hasselquist D (1994) Male Attractiveness, Mating Tactics and Realized Fitness in the Polygynous Great Reed Wabler, Thesis. Lund University, Lund, Sweden
Hasselquist D, Sherman PW (2001) Social mating systems and extrapair fertilization in passerine birds. Behav Ecol 12:457–466
Hitchcock RK, Yellen JE, Geldburd DJ, Osborn AJ, Crowell AL (1996) Subsistence hunting and resource management among the JU/HOASI of Northwestern Botswana. Afr Stud Monogr 17:153–220
Hockey PAR, Dean WJR, Ryan PG (eds) (2005) Roberts Birds of Southern Africa, 7th edn. The Trustees of the John Voelcker Bird Book Fund, Cape Town
Hӧgland J, Alatalo RV (1995) Leks. Princeton University Press, Princeton
Kemp A, Tarboton W (1976) Small South Africa bustards. Bokmakierie 28:40–43
Khan WA, Mian A (2013) Population biology of Black Francolin (Francolinus francolinus) with reference to Lal Suhanra National Park, Pakistan. Pak J Zool 45:183–191
Morales MB, Jiquet F, Arroyo B (2001) Exploded leks: what Bustards can teach us. Ardeola 48:85–98
Mucina L, Rutherford MC (eds) (2006) The vegetation of South Africa, Lesotho and Swaziland. Strelitzia 19. South African National Biodiversity Institute, Pretoria
Newton I (1998) Population Limitations in Birds. Academic, Amsterdam
Odum EP (1971) Fundamentals of Ecology, 3rd edn. WB Saunders, Philadelphia
Orians GH (1969) Age and hunting success in the Brown Pelican (Pelecanus occidenralis). Anim Behav 55:107–121
Pulliam HR (1973) On the advantages of flocking. J Theoret Biol 38:419–422
Sutherland WJ, Newton I, Green RE (2004) Bird Ecology and Conservation. Oxford University Press, New York
Tarboton WR, Kemp MI, Kemp AC (1987) Birds of the Transvaal. Transvaal Museum, Pretoria
van Niekerk JH (2014) Vocal behaviour of the red-crested Korhaan Lophotis ruficrista in South Africa. S Afr J Wildl Manage 44:24–31
Whitehead H (2008) Analyzing Animal Societies; Quantitative Methods for Vertebrate Social Analysis. University of Chicargo Press, London

 DownLoad:
DownLoad:






 Email Alerts
Email Alerts RSS Feeds
RSS Feeds