Eun-Jeong Kim, Wee-Heang Hur, Hwa-Jung Kim, Yu-Seong Choi, Dongwon Kim, Who-Seung Lee, Sejeong Han, Hyerin Joo, Chang-Yong Choi. 2024: Population trend and spatio-temporal distribution of Greater White-fronted (Anser albifrons) and Bean Geese (Anser fabalis) in Korea. Avian Research, 15(1): 100214. DOI: 10.1016/j.avrs.2024.100214
Citation:
Eun-Jeong Kim, Wee-Heang Hur, Hwa-Jung Kim, Yu-Seong Choi, Dongwon Kim, Who-Seung Lee, Sejeong Han, Hyerin Joo, Chang-Yong Choi. 2024: Population trend and spatio-temporal distribution of Greater White-fronted (Anser albifrons) and Bean Geese (Anser fabalis) in Korea. Avian Research, 15(1): 100214. DOI: 10.1016/j.avrs.2024.100214
Eun-Jeong Kim, Wee-Heang Hur, Hwa-Jung Kim, Yu-Seong Choi, Dongwon Kim, Who-Seung Lee, Sejeong Han, Hyerin Joo, Chang-Yong Choi. 2024: Population trend and spatio-temporal distribution of Greater White-fronted (Anser albifrons) and Bean Geese (Anser fabalis) in Korea. Avian Research, 15(1): 100214. DOI: 10.1016/j.avrs.2024.100214
Citation:
Eun-Jeong Kim, Wee-Heang Hur, Hwa-Jung Kim, Yu-Seong Choi, Dongwon Kim, Who-Seung Lee, Sejeong Han, Hyerin Joo, Chang-Yong Choi. 2024: Population trend and spatio-temporal distribution of Greater White-fronted (Anser albifrons) and Bean Geese (Anser fabalis) in Korea. Avian Research, 15(1): 100214. DOI: 10.1016/j.avrs.2024.100214
Department of Agriculture, Forestry, and Bioresources, Seoul National University, Seoul, 08826, Republic of Korea
b.
Research Institute of Agriculture and Life Science, Seoul National University, Seoul, 08826, Republic of Korea
c.
National Migratory Birds Center, National Institute of Biological Resources, Incheon, 22689, Republic of Korea
d.
Division for Natural Environment, Korea Environment Institute, Sejong, 30147, Republic of Korea
Funds:
“The analysis of biogeographical and phenological migration pattern of birds as vectors of avian influenza” of the Korea Environment Institute0525–202300058
“Long-term change of waterbird population in climate change” of the National Institute of Biological Resources in South KoreaNIBR202318101
“Long-term change of waterbird population in climate change” of the National Institute of Biological Resources in South KoreaNIBR202416101
Geese have undergone significant changes in their distribution and population size due to human-induced impacts. To improve our understanding of the two main geese populations in East Asia—Greater White-fronted (Anser albifrons; GWFG) and Bean Geese (Anser fabalis sensu lato; BEAG), which includes the Tundra A. f. serrirostris (TDBG) and Taiga Bean Geese A. f. middendorffii (TGBG), this study estimated their long- and short-term trends and the migration strategies based on their distribution during the migratory and wintering seasons, using nationwide census data collected over 24 years in South Korea. The TRIM (TRends and Indices for Monitoring data) analysis supported a robust long-term population increase since 1999, recently stabilizing around 150,000 individuals in GWFG and 100,000 in TDBG, respectively. But TGBG stabilized at around 7000 with no significant change of population over time. The shorter-term trends within a season suggested that wintering populations of GWFG and TDBG in China might stop over in Korea in their autumn migration before their sea-crossing, evidenced by the large congregation near the Yellow Sea coast followed by a gradual decrease. In contrast to autumn, there was no similar peak in spring migration, suggesting the potential for elliptical migration of some populations around the Yellow Sea. Our findings indicate a growing East Asian population and imply one of the migration strategies of the Greater White-fronted and Tundra Bean Geese. By collaborating with recent tracking data, long-term monitoring efforts could offer more detailed insights into population trends and migration strategies, thereby contributing to the effective management and conservation of goose species in East Asia.
Understanding these variations in population size and distribution is not only vital for developing effective management and conservation measures but is also crucial for monitoring recently emerging infectious diseases, such as highly pathogenic avian influenza (HPAI). With the advancement of technology like tracking devices (e.g., Global Positioning System, GPS; Platform Transmitter Terminal, PTT), we can readily gather valuable insights into migration characteristics (Mitchell et al., 2016; Si et al., 2018; Zheng et al., 2018), local movements (Mitchell et al., 2016; Luukkonen et al., 2022), and changes in the distribution of geese (Lameris et al., 2023). Nevertheless, individual-based data may yield incorrect species-specific information at the population level due to the small sample size and limitations in non-random deployment in specific, easily accessible areas (Hebblewhite and Haydon, 2010; Meehan et al., 2022), including the relatively high cost compared to other methods (Gregory et al., 2023).
For these reasons, large-scale monitoring data (Fuentes et al., 2023) or various methods integrating telemetry or tracking data (Meehan et al., 2022; Gregory et al., 2023) have been implemented to track population changes. In cases where tracking data are insufficient, large-scale monitoring data such as census surveys and citizen science data could be utilized to understand the population movements (Fuentes et al., 2023). Census data, which counts all birds within the entire study area (Sutherland, 1996; Gregory et al., 2004), has the advantage of providing national population information when covering extensive regions, resolving the issues related to the representativeness of individual-based data. Moreover, when conducted at regular intervals over a long period, census data could unveil the trends in population size and population-scale migration movement through changes in distribution (Walker and Taylor, 2017; Fuentes et al., 2023). There is also the possibility of discovering the migration strategies of populations not confirmed in a limited number of tracking data.
The Greater White-fronted (Anser albifrons; GWFG) and Bean Geese (A. fabalis sensu lato; BEAG) are the dominant goose species in South Korea (the Republic of Korea) and representative long-distance migrants breeding in northeastern Russia. GWFG and BEAG populations in Korea are part of the East Asian population, divided into three populations based on their wintering countries: China, Japan, and Korea (Wetlands International, 2024b). Due to their extensive breeding range (BirdLife International, 2018, 2022) and the attributes of breeding area in Arctic Russia (GWFG: Northeast Russian Arctic; BEAG: from Central to Northeast Russian Arctic), primarily in remote, inaccessible, and uninhabited areas, accurately calculating the population size and trends of East Asian populations in the breeding sites is challenging compared to those of Europe and North America. Therefore, to gain a better understanding of the East Asian population of this species, the population information from winter counts is vital (Delany, 2005; Wetlands International, 2024a).
In this study, we aimed to document information about both GWFG and BEAG populations in South Korea. We utilized national census data to identify temporal (long-term and short-term) and spatial changes in the numbers of GWFG and BEAG during non-breeding periods. Therefore, we investigated how the population size in South Korea has changed over 24 years using the TRIM (TRends and Indices for Monitoring data) model. We also understood migration patterns from monthly patterns of population sizes, predicting if both species spend their non-breeding time in Korea, a highly peaked number would be observed in the main winter month (December of November). Lastly, we checked the distribution and primary wintering regions.
2.
Materials and methods
2.1
National census data
The Nationwide Winter Waterbird Census (NWWC) data are prepared and compiled by the Korean Ministry of Environment and associated institutes, namely the National Institute of Environmental Research (1999–2007) and the National Institute of Biological Resources (2008–present). Conducted annually every January since 1999, the NWWC covers important wintering habitats of migratory waterbirds, including lakes, wetlands, bays, and seashores. Beginning with 82 sites in 1999, the survey area has consistently expanded, reaching 200 sites by 2014. In addition, to meet the growing demand for data on the distribution status of migratory waterbirds, additional surveys have been conducted from October to March of the subsequent year since 2014 (NIBR, 2024). Nationwide censuses have been synchronously carried out over two consecutive days in the middle of each month. The census team for each region was composed of two experienced researchers, such as ornithologists and bird watchers. They moved through the survey area by vehicle, boat, and foot, recording all the species and number of birds observed at the target sites.
Since NWWC was conducted without taxonomic splitting of the Bean Goose from the beginning, we tried to divide this species into two subspecies based on their main geographic distribution (Park, 2002; Jia et al., 2016). So, we considered BEAG observed in the Nakdong River watershed, Upo Wetland, and Junam Reservoir as TGBG.
2.2
Long-term population trends of GWFG, TDBG, and TGBG
In long-term monitoring, specific sites may not be counted in certain years, or new sites may be added while others vanish. One might attempt to address this issue by comparing only the sites monitored in all years, but such an approach would be inadequate for accurately identifying the population size and trends (Braak et al., 1994). The TRIM (TRends and Indices for Monitoring data) model is designed to analyze time series of counts with missing observations and is commonly used to estimate the growth and decline of animal populations (Pannekoek and van Strien, 2001). Thus, to understand the long-term population trend of GWFG and BEAG, we employed the TRIM method with winter census data collected every January from 1999 to 2023 using the rtrim package (Version 2.1.1; Bogaart et al., 2020), considering the variability in survey sites and missing values in our data.
2.3
Short-term spatial-temporal population trends
Given that the NWCC survey period included not only the main wintering season (December and January) but also migration seasons (spring: February and March; autumn: October and November), we expected that census data have sufficient potential to reflect population-scale migration information from changes in their distribution in South Korea. Therefore, we analyzed spatio-temporal population changes during winter using Generalized Additive Mixed Models (GAMMs; Wood, 2017) because the relationships between counts data and spatio-temporal variables could be nonlinear (Zhang et al., 2015; Knape, 2016).
The number of survey sites was not identical across survey months; 200 sites were surveyed during the main wintering season, January and February, while only 112 main areas were surveyed during the migration season. So, to control the effect of the varying number of survey sites, only sites surveyed consistently every month were selected for spatio-temporal analysis.
First, we estimated monthly patterns of population size for the entire wintering GWFG, TDBG, and TGBG using the following procedure. For our response variable, we filtered out the monthly count data of each survey site between 2014 and 2023, and we aggregated country-level monthly count data by merging each species’ abundances of all survey sites in each survey season; for example, the 2022–2023 season denoted the survey conducted from October 2022 to March 2023. In GAMMs, we included the intercept for survey seasons as a random effect and a smooth effect for survey months with a Poisson distribution. After comparing with the possible subset models based on the Akaike Information Criterion adjusted for small sample sizes (AICc; Burnham and Anderson, 2002), we selected the optimal model with the lowest AICc values. We assessed the significance of smooth effects for each group via p-values in the selected GAMMs and considered p < 0.05 to indicate statistically significant predictors. Predicted fitted values from the GAMMs were visualized to illustrate monthly changes in GWFG and BEAG, respectively.
Second, we examined whether monthly population changes were related to the latitudinal gradient. Because TGBG was distributed in restricted regions, we performed this analysis for BEAG (sensu lato). We used the site-level count data and calculated the centroid points to obtain the latitude of survey sites using QGIS (QGIS, 2023). Regarding site-level counts, as count data often include zero observations, a zero-inflated Poisson distribution was applied to GAMMs. The model for each species relating the site-level counts (response variables) to explanatory variables in interaction with survey sites’ latitude and month, including survey seasons as a random effect, was assessed and visualized using the same methods as above.
Population sizes were given as the means with standard deviations (SD). All analyses were conducted in R version 4.2.3 (R Core Team, 2023) to implement GAMMs using the mgcv package (Version 1.9.1; Wood, 2017), and the results were visualized through the ggplot2 package (Version 3.5.1; Wickham, 2016).
3.
Results
3.1
Long-term population trends of GWFG, TDBG, and TGBG
The TRIM model for GWFG in South Korea revealed a ‘moderate increase’ from 1999 to 2023 (p < 0.01). The GWFG population in January started at 61,328 in 1999 and rapidly increased after 2003 and 2016. In 2020, the population peaked at 182,608, three times larger than in 1999. The most recent survey in January 2023 recorded 127,843 GWFGs.
The Tundra Bean Goose's population showed a ‘moderate increase’ in the TRIM model (p < 0.01). A total of 34,177 individuals were recorded in January 1999, and the TDBG population also increased sharply around 2005. However, after that, January's TDBG population size fluctuated, reaching 95,344 in 2023. In contrast, Taiga Bean Goose's population trends fluctuated between 4000 and 10,000 over 24 years, not showing a clear increasing or decreasing trend in the TRIM Model (Fig. 1).
Figure
1.
The population trend of the wintering Greater White-fronted (GWFG, Anser albifrons; A), Tundra Anser fabalis serrirostris (TDBG; B) and Taiga Bean Geese A. f. middendorffii (TGBG; C) observed in January from 1999 to 2023 in South Korea. The red line and band represent the result of the TRIM (trends and indices for monitoring data) analysis with 95% confidence interval, and the black point shows the counted number of geese in the Nationwide Winter Waterbird Census.
We found strong spatiotemporal patterns in the monthly population trends of GWFG and BEAG based on the survey month and site's latitude, with high significance in the explanatory variables in GAMMs (p < 0.001; Table 1).
Table
1.
Results of the generalized additive mixed models (GAMMs) with temporal (month) and spatial (latitude) variables affecting Greater White-fronted (Anser albifrons; GWFG) and Bean Geese (Anser fabalis sensu lato; BEAG) population sizes in South Korea. Tundra Anser fabalis serrirostris (TDBG) and Taiga Bean Geese A. f. middendorffii, (TGBG) were estimated based on their wintering distribution, not by census data.
Species
Model
Model term
Coeff.
SE
EDFa
F
t
p
GWFG
Monthly
Intercept
116810
7115
16.42
<0.001
Month
3.57
29.79
<0.001
Seasonb
6.18
3.40
<0.001
Latitudinal
Intercept
1969.70
291.20
6.76
<0.001
Latitudec × Month
474.37
20.25
<0.001
Seasonb
7.48
3.19
<0.001
TDBG
Monthly
Intercept
61178
3003
20.37
<0.001
Month
1.00
133.75
<0.001
Seasonb
0.61
0.08
0.391
TGBG
Monthly
Intercept
4873.40
371.80
13.11
<0.001
Month
3.97
29.23
<0.001
Seasonb
4.65
1.39
<0.05
BEAG
Latitudinal
Intercept
1147.69
57.32
20.02
<0.001
Latitudec × Month
412.27
21.04
<0.001
Seasonb
1.76
0.27
0.274
a Number of the effective degrees of freedom. b Random effect. c Values from the centroid of each survey site.
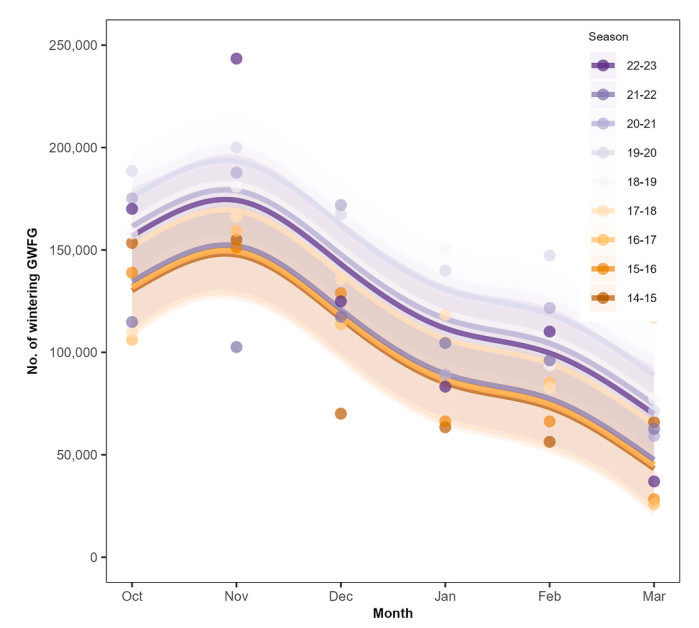
The estimated monthly population trend of GWFG was right-skewed unimodal (Fig. 2), peaking in November and continuously decreasing afterward. The average GWFG counts and standard deviations (SD) during spring migration (February and March) were 77,016 ± 31,825, which was only half of the average population size during autumn migration (October and November; 158,879 ± 36,121) and significantly lower than in autumn (Tukey test: mean difference = −81,862.50, p < 0.001). In winter (December and January), the average counts were 114,534 ± 34,656, slightly lower than autumn migration counts (Tukey test: mean difference = −44,344.78, p < 0.001).
Figure
2.
Monthly population change of Greater White-fronted Geese (Anser albifrons; GWFG) in South Korea. The numbers of counted GWFG (points) and the trend, fitted values from the generalized additive mixed models (GAMMs; lines) with 95% confidence interval (bands), are displayed. Each color represents each season, from 2014 to 2015 to 2022–2023.
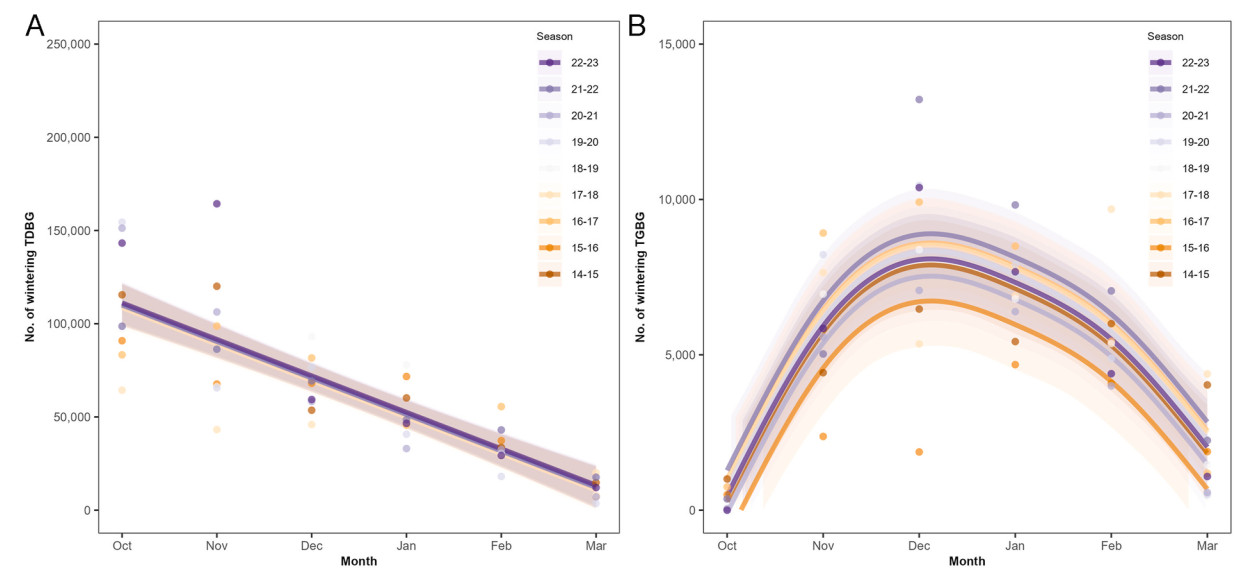
TDBG also showed the highest population counts, 100,965 ± 34,901, during the autumn migration season, steadily decreasing in winter to 59,894 ± 16,231 and in spring to 22,675 ± 14,504 (Dunn's test; autumn-spring: z = 6.22, p < 0.001; autumn-winter: z = 2.55, p < 0.05; spring-winter: z = −3.67, p < 0.001), similar to GWFG. However, the peak population of TDBG was observed in October, and the monthly trend showed a linear decrease rather than an unimodal pattern (Fig. 3A).
Figure
3.
Monthly population change of Tundra Anser fabalis serrirostris (TDBG; A) and Taiga Bean Goose A. f. middendorffii (TGBG; B) in South Korea. The numbers of counted TDBG and TGBG (points) and the trend, fitted values from the generalized additive mixed models (GAMMs; lines) with 95% confidence interval (bands), are displayed. Each color represents each season, from 2014 to 2015 to 2022–2023. Note the different scales between the two panels.
Unlike GWFG and TDBG, TGBG showed a completely different trend, displaying a bell-shaped pattern with the highest population count in winter (7,623 ± 2,595; Fig. 3B). There was no difference in the counted numbers between autumn (3,205 ± 3,316) and spring (3,792 ± 2,478) migration seasons (Dunn's test; autumn-spring: z = −0.47, p = 0.64; autumn-winter: z = −3.89, p < 0.001; spring-winter: z = −3.42, p < 0.001).
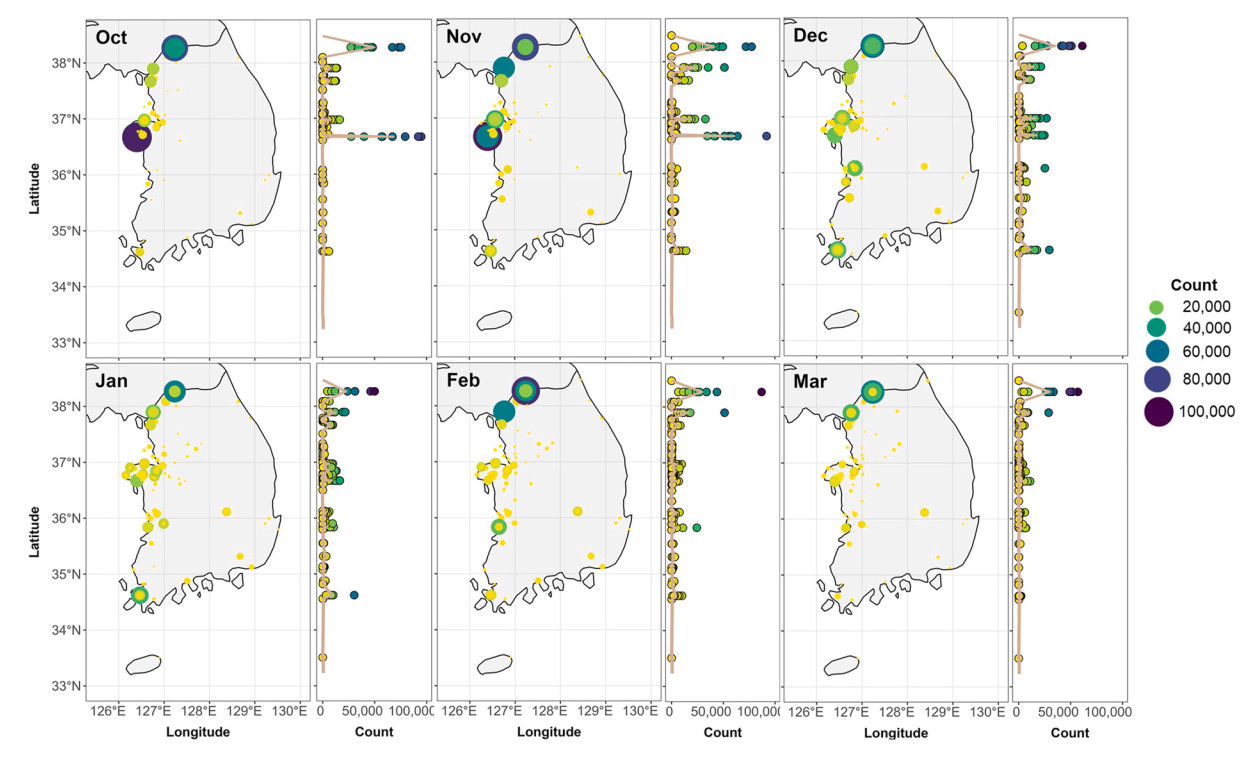
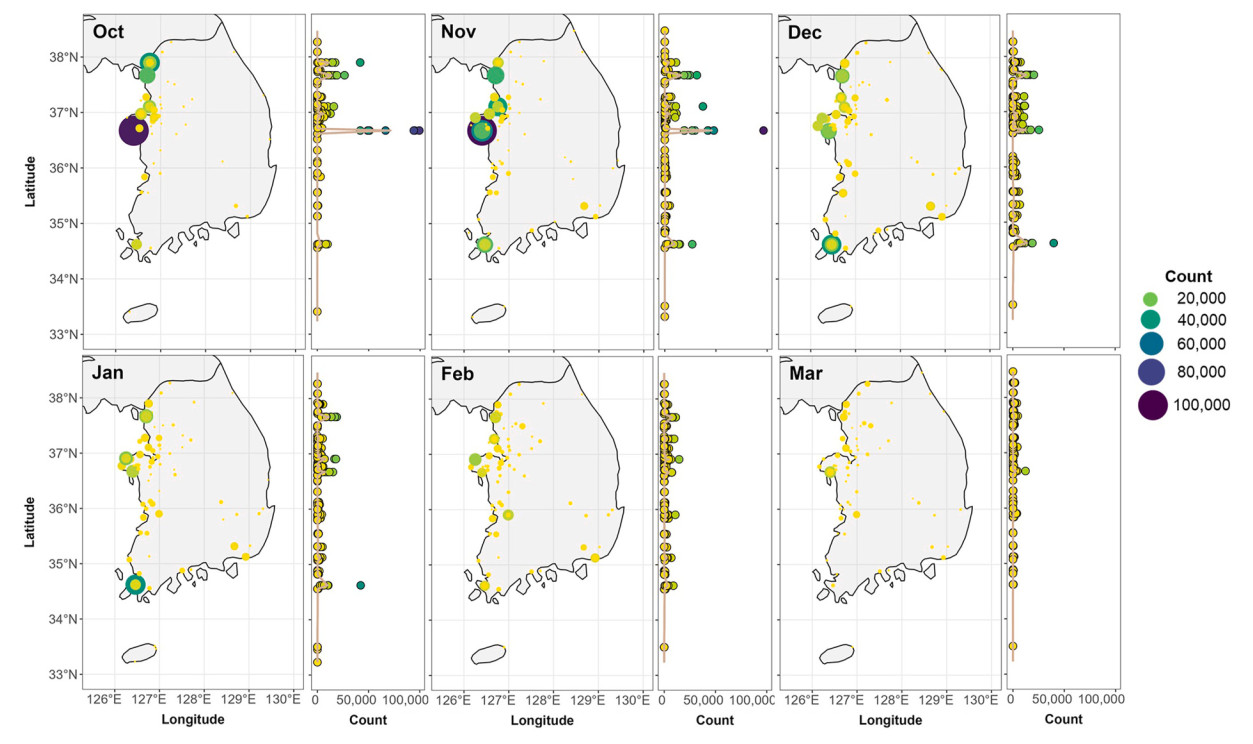
From our GAMMs, which included interactions between latitude and survey month, we identified different monthly distributions of GWFG and BEAG along the Yellow Sea coast (Figs. 4 and 5). High populations of GWFG, about 100,000 and 80,000, were observed at 36.7° N and 38.3° N, respectively, during autumn migration. Although the GWFG's high population disappeared at 36.7° N afterward, it was still present at 38.3° N (Table 1; Fig. 4). A notable number of BEAG was also observed at 36.7° N and 37.9° N during the autumn migration season. As winter approached, the significant numbers observed at this latitude vanished. There were no specific latitudes where a significant number of individuals were observed in February and March (Fig. 5).
Figure
4.
The monthly observed Greater White-fronted Geese (Anser alfibrons; GWFG) numbers of survey sites (left) and the trend of numbers according to the latitudinal gradient (right). On the right graph, the color indicates the number of observed GWFG, the same color as the left graph's legend.
Figure
5.
The monthly observed Bean Goose (Anser fabalis sensu lato; BEAG) numbers of survey sites (left) and the trend of numbers according to the latitudinal gradient (right). On the right graph, the color indicates the number of observed BEAG, the same color as the left graph's legend.
In this study, we aimed to understand trends in the population size and spatio-temporal distribution of GWFG and BEAG (TDBG and TGBG) in South Korea using the national-level census data from NWWC. We noted a substantial increase in the wintering population sizes of GWFG and TDBG in South Korea over 24 years. After the moderate increasing trends from 1999, the population has stabilized at about 150,000 for GWFG and 100,000 for TDBG over the past five years. Contrary to TDBG, TGBG in Korea has not experienced significant population size changes over time.
In Japan, Shimada et al. (2019) confirmed an increase in the Japanese GWFG population based on national-scale population data from the Japanese Ministry of Environment, indicating different population trends depending on latitude, mainly concentrated in Northern Japan (Miyagi Prefecture). On the other hand, Deng et al. (2020) reported a decrease in the GWFG population in China from 142,159 in 1992/93 to 54,223 in 2019/20, though this decrease was not statistically significant. The rise in the Korean population might imply a shift of Chinese populations to South Korea. Still, population changes in all three countries indicate an overall increase since 2014, when Korea's survey areas have not been changed. Moreover, the magnitude of the increase in South Korea (about 40,000 from 2014; NIBR, 2024) and Japan (about 70,000 from 2014; Shimada et al., 2019; Deng et al., 2020) is greater than the decrease in China's population (a reduction of 90,000 from 1993; Deng et al., 2020), suggesting the possibility of an overall growth in the East Asian GWFG population.
Li et al. (2020) confirmed the TDBG population has been increasing in all three East Asian countries (South Korea, China and Japan). While Japan showed only a slight increase (from 1500 to 8754 since the 1970s), China reported a remarkable nine-fold increase in population since the 1990s. With South Korea's current population size growing by an additional 20,000 individuals compared to Li et al. (2020), the East Asian population of TDBG will continue to grow constantly unless there are changes in population trends in other countries.
On the other hand, Li et al. (2020) described that the TGBG populations in Japan and Korea increased slightly, but considering the fluctuation range of the Japanese population over time is similar to ours and our TRIM results, it is believed that the East Asian population of TGBG has been maintained without any significant changes for a long time.
While our study relies solely on data collected in South Korea, insights into East Asian population variations can be derived from other research. Therefore, it is essential to compare not only Korea's data but also that of Japan and China to understand changes in the overall East Asian population trends comprehensively. However, the limited data on the Chinese population in the absence of regular nationwide surveys, unlike in South Korea and Japan, makes it difficult to grasp the population size trends. Therefore, for an accurate assessment of trends in the East Asian population and to compare population changes among countries, international cooperation among Korea, Japan, and China is essential to conduct census surveys with systematic methods, such as identical or similar survey periods and efforts.
4.2
Spatio-temporal distribution and migration strategy
Due to the survey intervals (once a month), it was not possible to capture detailed short-term temporal changes, but we were able to discern their migration strategies, which had not been confirmed previously. TGBG, consistent with our hypothesis, was observed in larger numbers during the main winter season, in December and January, than during the autumn and spring migration seasons, indicating that TGBG observed are true wintering individuals in Korea. However, we observed more individuals of GWFG and TDBG during the migration seasons, especially in autumn. Moreover, the high number detected near the Yellow Sea coast (36.7° N) in the early autumn migration season suggested the occurrence of a passage group that may stop over in Korea during their southward migration heading for wintering grounds in China (such as Poyang Lake and/or unconfirmed sites). Interestingly, the short-term decreasing trend in winter and the absence of a second peak in March provided a possible clue that GWFG and TDBG may use different routes, such as detouring South Korea to employ an elliptical migration pattern, during their northward spring migration.
One GWFG migration track in Movebank (https://www.movebank.org; movebank ID: 1614083125, GWFG ID: H129) displayed a southward migration from its breeding ground to China via northern South Korea and a subsequent northward migration detouring Korea, which supports the suggested migration pattern observed in our spatio-temporal data. Although the track represented only one individual, given their tendency to migrate in groups, some passage groups of GWFG in South Korea likely employ this elliptical migration strategy more broadly during their migration.
For TDBG, one individual wintering in Nantong, China, passed through Seoul, Korea, during its northward migration (movebank ID: 1614098614, BEAG ID: H1048). Additionally, among individuals tracked over multiple years, some utilized both Korea and China during the same winter season (Li et al., 2020). There was no evidence of an elliptical migration pattern proven by telemetry. However, the single peak only in autumn still suggests the pattern and the close migratory connectivity between Korea and China.
4.3
Increasing populations and pressures
The different latitudinal variations of counts in each month were linked to specific survey sites. The largest number near the Yellow Sea coast, occurring solely during the early autumn migration season, was from Cheonsu Bay (36.7° N, 126.4° E), consisting of 15,409 ha of agricultural land developed into reclaimed land as part of a national reclamation project. The northern region of South Korea includes the Cheorwon Plains (38.3° N, 127.2° E), characterized by high observations of GWFG counts during all survey periods, and the Imjin River (37.9° N, 126.8° E), where many TDBG are observed only in the autumn migration season. The variation in the geese population in wintering grounds is known to be primarily influenced by food availability (Zhao et al., 2010; Pearse et al., 2011; Cunningham et al., 2021) and mortality rates resulting from hunting (Lindberg and Malecki, 1994; Menu et al., 2002; Panov et al., 2022). With the strong regulation of wildlife hunting and poaching in South Korea since 1994 (Ministry of Government Legislation, 2011b, Ministry of Government Legislation, 1994), food availability has become the main factor for determining population size. Specifically, geese wintering in South Korea heavily depend on grains in rice paddies (Fujioka et al., 2010; Kim et al., 2016; Fox et al., 2017), making rice fields crucial habitats for sustaining goose populations as they offer a substantial amount of nutrition compared to natural wetlands and also serve as roosting and resting places (Czech and Parsons, 2002; Fujioka et al., 2010; Toral and Figuerola, 2010; Natuhara, 2013). Moreover, the conversion of saltmarshes and mudflats into rice fields, exemplified by the Saemangeum Seawall Project from 1991 to 2010 (Saemangeum Development Agency, 2024), has provided advantages for wintering geese by allowing them to forage on the remaining grain after harvest on the fields. Kim et al. (2016) confirmed that the increase in the population of geese overwintering in South Korea was closely related to the completion of the Saemangeum Project, suggesting that the geese may have utilized the newly reclaimed area near the west coast.
Nevertheless, the recent stable trends of geese in Korea may suggest that the geese populations might reach the carrying capacity for geese in Korea. Korean rice fields have been recently adopted bale silages for the collection of waste grains and straws to provide additional agricultural products and feed for livestock (Chung et al., 2021). Ongoing anthropogenic interference with urbanization continues to reduce the agricultural land, resulting in the decreased availability of grains in rice fields (Statistics Korea, 2023). Moreover, changes in agricultural policy due to reduced demand for rice (Jeong et al., 2021) and population decline in rural areas (Li et al., 2023) can make it hard to predict future changes in food availability for geese. Consequently, there could be a possibility that geese may need to move more within wintering grounds to find sufficient food or switch to less nutritious food. This could result in difficulties in obtaining enough energy for long-distance migration, negatively impacting the survival of wintering and migrating geese (Terrill, 1990; Owen et al., 1992; Harrison et al., 2013; Aharon-Rotman et al., 2017). Additionally, considering the density-dependent effects of the increasing population size, such as agricultural conflicts (Jensen et al., 2008; Mason et al., 2018) and intra- and interspecific competition for food and habitats (Schmutz and Laing, 2002; Zhao et al., 2020), appropriate management of key habitats is deemed necessary in South Korea to sustain the increasing trend or maintain the current population size.
5.
Conclusions
Our study holds great value in supporting the increase in the East Asian population of GWFG and BEAG and confirming a previously unobserved migration strategy, known as elliptical migration, through long-term census data. We believe further investigation is required to understand the reasons for using elliptical migration by linking it with external environmental factors and examining the ecological impacts of population growth along the complex migration network in the East Asian-Australasian Flyway. Additionally, collaborating with tracking data, long-term monitoring efforts could offer more detailed insights into population trends and migration strategies, contributing to the management and conservation of goose species in this region.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Béchet, A., Giroux, J., Gauthier, G., 2004. The effects of disturbance on behaviour, habitat use and energy of spring staging snow geese. J. Appl. Ecol. 41, 689-700. .
BirdLife International, 2018. Anser fabalis. The IUCN Red List of Threatened Speceis 2018: e.T22679875A132302864. .
BirdLife International, 2022. Anser albifrons. The IUCN Red List of Threatened Species 2022: e.T22679881A213839615. . RLTS.T22679881A213839615.en. (Accessed on 12 November 2024).
Bogaart, P., van der Loo, M., Pannekoek, J., 2020. rtrim: trends and indices for monitoring data. .
Braak, C.T., Van Strien, A., Meijer, R., Verstrael, T., 1994. Analysis of monitoring data with many missing values: which method? In: Hagemeijer, W., Verstrael, T. (Eds.), Bird Numbers 1992: Distribution, Monitoring and Ecological Aspects. Statistics Netherlands & SOVON, Voorburg/Heerlen & Beek-Ubbergen, pp. 663–673.
Burgers, J., Smit, J.J., van der Voet, H., 1991. Origins and systematics of two types of the Bean Goose Anser fabalis (Latham, 1787) wintering in The Netherlands. Ardea 79, 307-316.
Burnham, K.P., Anderson, D.R., 2002. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, second ed. Springer-Verlag, New York. .
Chung, O.-S., Hur, W.-H., Kim, H.-J., Lee, J.K., Park, J.-Y., 2021. Decline in the population of Korean wintering waterbirds after use of bale silage and completion of a massive river engineering project. Waterbirds 44, 185-191. .
Clausen, K.K., Clausen, P., Fælled, C.C., Mouritsen, K.N., 2012. Energetic consequences of a major change in habitat use: endangered Brent Geese Branta bernicla hrota losing their main food resource. Ibis 154, 803-814. .
Cunningham, S.A., Zhao, Q., Weegman, M.D., 2021. Increased rice flooding during winter explains the recent increase in the Pacific Flyway White‐fronted Goose Anser albifrons frontalis population in North America. Ibis 163, 231-246. .
Czech, H.A., Parsons, K.C., 2002. Agricultural wetlands and waterbirds: a review. Waterbirds 25, 56-65.
Delany, S., 2005. Guidelines for Participants in the International Waterbird Census (IWC). Wetlands International, Wageningen, The Netherlands.
Deng, X., Zhao, Q., Diana, S., Lee, H., Bysykatova-harmey, I., Xu, Z., et al., 2020. Contrasting trends in two East Asian populations of the greater white-fronted goose Anser albifrons. Wildfowl 6, 181-205.
Desmonts, D., Fritz, H., Cornulier, T., Maheo, R., 2009. Rise in human activities on the mudflats and Brent Geese (Branta bernicla) wintering distribution in relation to Zostera spp. beds: a 30-year study. J. Ornithol. 150, 733-742. .
Ely, C.R., Dzubin, A.X., Mlodinow, S.G., Kirwan, G.M., Carboneras, C., Garcia, E., 2024. Greater white-fronted goose (Anser albifrons). In: Sly, N.D., Keeney, B.K. (Eds.), Birds of the World. Cornell Lab of Ornithology. .
Fox, A.D., Madsen, J., Boyd, H., Kuijken, E., Norriss, D.W., Tombre, I.M., et al., 2005. Effects of agricultural change on abundance, fitness components and distribution of two arctic‐nesting goose populations. Glob. Chang. Biol. 11, 881-893. .
Fox, A.D., Elmberg, J., Tombre, I.M., Hessel, R., 2017. Agriculture and herbivorous waterfowl: A review of the scientific basis for improved management. Biol. Rev. 92, 854-877. .
Fox, A.D., Frederiksen, M., Heinicke, T., Clausen, K.K., van der Jeugd, H.P., 2021. Annual survival estimates of Taiga Anser fabalis and Tundra bean geese A. serrirostris wintering in The Netherlands, 1967–1987. J. Ornithol. 162, 925-929. .
Fuentes, M., van Doren, B.M., Fink, D., Sheldon, D., 2023. BirdFlow: Learning seasonal bird movements from eBird data. Methods Ecol. Evol. 14, 923-938. .
Fujioka, M., Don Lee, S., Kurechi, M., 2010. Bird use of rice fields in Korea and Japan. Waterbirds 33, 8-29. .
Gregory, R.D., Gibbons, D.W., Donald, P.F., 2004. Bird census and survey techniques. In: Bird Ecology and Conservation. Oxford University Press, pp. 17–56. .
Gregory, K.A., Francesiaz, C., Jiguet, F., Besnard, A., 2023. A synthesis of recent tools and perspectives in migratory connectivity studies. Mov. Ecol. 11, 69. .
Harrison, X.A., Hodgson, D.J., Inger, R., Colhoun, K., Gudmundsson, G.A., McElwaine, G., et al., 2013. Environmental conditions during breeding modify the strength of mass-dependent carry-over effects in a migratory bird. PLoS One 8, e77783. .
He, K., Lei, J., Jia, Y., Wu, E., Sun, G., Lu, C., et al., 2022. Temporal dynamics of the goose habitat in the middle and lower reaches of the Yangtze River. Rem. Sens. 14, 1883. .
Hebblewhite, M., Haydon, D.T., 2010. Distinguishing technology from biology: A critical review of the use of GPS telemetry data in ecology. Philos. Trans. R. Soc. B Biol. Sci. 365, 2303-2312. .
Heinicke, T., 2018. A2 western Tundra bean goose Anser fabalis rossicus. In: Fox, A.D., Leafloor, J.O. (Eds.), A Global Audit of the Status and Trends of Arctic and Northern Hemisphere Goose Populations (Component 2: Population Accounts). Conservation of Arctic Flora and Fauna (CAFF), Akureyri, Iceland, pp. 10–13.
Jensen, R.A., Wisz, M.S., Madsen, J., 2008. Prioritizing refuge sites for migratory geese to alleviate conflicts with agriculture. Biol. Conserv. 141, 1806-1818. .
Jeong, O.-Y., Park, H.-S., Baek, M.-K., Kim, W.-J., Lee, G.-M., Lee, C.-M., et al., 2021. Review of rice in Korea: Current status, future prospects, and comparisons with rice in other countries. J. Crop Sci. Biotechnol. 24, 1-11.
Jia, Q., Koyama, K., Choi, C.-Y., Kim, H.-J., Cao, L., Gao, D., et al., 2016. Population estimates and geographical distributions of swans and geese in East Asia based on counts during the non-breeding season. Bird. Conserv. Int. 26, 397-417. .
Kim, M.K., Lee, S., Lee, S.D., 2016. Habitat use and its implications for the conservation of the overwintering populations of bean goose Anser fabalis and greater white-fronted goose A. albifrons in South Korea. Ornithol. Sci. 15, 141-149. .
Kirby, J.S., Stattersfield, A.J., Butchart, S.H.M., Evans, M.I., Grimmett, R.F.A., Jones, V.R., et al., 2008. Key conservation issues for migratory land- and waterbird species on the world's major flyways. Bird. Conserv. Int. 18, S49-S73. .
Knape, J., 2016. Decomposing trends in Swedish bird populations using generalized additive mixed models. J. Appl. Ecol. 53, 1852-1861. .
Knudsen, E., Lindén, A., Both, C., Jonzén, N., Pulido, F., Saino, N., et al., 2011. Challenging claims in the study of migratory birds and climate change. Biol. Rev. 86, 928-946. .
Koons, D.N., Rockwell, R.F., Aubry, L.M., 2014. Effects of exploitation on an overabundant species: The lesser snow goose predicament. J. Anim. Ecol. 83, 365-374. .
Kurechi, M., Yokota, Y., Otsu, M., 1983. Notes on the field identification of Anser fabalis serrirostris and A. f. middendorfi. Jpn. J. Ornithol. 32, 95-108. .
Lameris, T.K., Pokrovskaya, O.B., Kondratyev, A.V., Anisimov, Y.A., Buitendijk, N.H., Glazov, P.M., et al., 2023. Barnacle geese Branta leucopsis breeding on Novaya Zemlya: Current distribution and population size estimated from tracking data. Polar Biol. 46, 67-76. .
Layton‐Matthews, K., Hansen, B.B., Grøtan, V., Fuglei, E., Loonen, M.J.J.E., 2020. Contrasting consequences of climate change for migratory geese: Predation, density dependence and carryover effects offset benefits of high‐arctic warming. Glob. Chang. Biol. 26, 642-657. .
Li, C., Zhao, Q., Solovyeve, D., Lameris, T., Batbayar, N., Bysykatova-harmey, I., et al., 2020. Population trends and migration routes of the East Asian bean goose Anser fabalis middendorffii and A. f. serrirostris. Wildfowl 6, 124-156.
Li, W., Zhang, L., Lee, I., Gkartzios, M., 2023. Overview of social policies for town and village development in response to rural shrinkage in East Asia: The cases of Japan, South Korea and China. Sustainability 15, 10781.
Lindberg, M.S., Malecki, R.A., 1994. Hunting vulnerability of local and migrant Canada Geese in Pennsylvania. J. Wildl. Manag. 58, 740. .
Luukkonen, B.Z., Klaver, R.W., Jones Ⅲ, O.E., 2022. Movement of Canada Geese in urban and rural areas of Iowa, USA. Avian Conserv. Ecol. 17, 27. .
Madsen, J., Schreven, K.H.T., Jensen, G.H., Johnson, F.A., Nilsson, L., Nolet, B.A., et al., 2023. Rapid formation of new migration route and breeding area by Arctic geese. Curr. Biol. 33, 1162-1170. .
Marjakangas, A., Alhainen, M., Fox, A.D., Heinicke, T., Madsen, J., Nilsson, L., et al., 2015. International single species action plan for the conservation of the Taiga bean goose Anser fabalis fabalis. AEWA Technical Series No. 56. African-Eurasian Migratory Waterbird Agreement, Bonn, Germany. . (Accessed 25 March 2024).
Mason, T.H.E., Keane, A., Redpath, S.M., Bunnefeld, N., 2018. The changing environment of conservation conflict: Geese and farming in Scotland. J. Appl. Ecol. 55, 651-662. .
Meehan, T.D., Saunders, S.P., DeLuca, W.V., Michel, N.L., Grand, J., Deppe, J.L., et al., 2022. Integrating data types to estimate spatial patterns of avian migration across the Western Hemisphere. Ecol. Appl. 32, e2679. .
Menu, S., Gauthier, G., Reed, A., 2002. Changes in survival rates and population dynamics of greater snow geese over a 30‐year period: Implications for hunting regulations. J. Appl. Ecol. 39, 91-102.
Ministry of Government Legislation, 1994. Birds and mammals protection and management Act. (In Korean) . (Accessed 6 January 2024).
Ministry of Government Legislation, 2011b. Wildlife protection and management Act. . (Accessed 6 January 2024).
Mitchell, C., Griffin, L., Maciver, A., Minshull, B., Makan, N., 2016. Use of GPS tags to describe the home ranges, migration routes, stop-over locations and breeding area of Taiga Bean Geese Anser fabalis fabalis wintering in central Scotland. Hous. Theor. Soc. 63, 437-446. .
National Institute of Biological Resources, 2024. Nationwide winter waterbird census. . (Accessed 26 March 2024).
Natuhara, Y., 2013. Ecosystem services by paddy fields as substitutes of natural wetlands in Japan. Ecol. Eng. 56, 97-106. .
Owen, M., Wells, R.L., Black, J.M., 1992. Energy budgets of wintering Barnacle Geese: The effects of declining food resources. Ornis Scand. 23, 451. .
Panov, I.N., Litvin, K.E., Ebbinge, B.S., Rosenfeld, S.B., 2022. Reasons for the reduction in the population of the western subspecies of the Bean Goose (Anser fabalis fabalis and Anser fabalis rossicus): What do the ringing data say? Biol. Bull. 49, 839-850. .
Pannekoek, J., van Strien, A., 2001. TRIM 3 Manual. (Trends and Indices for Monitoring Data). Research Paper No. 0102. Statistic Netherlands, Voorburd, the Netherlands.
Park, J.Y., 2002. Current Status and Distribution of Birds in Korea. Kyung Hee University, Seoul.
Pearse, A.T., Alisauskas, R.T., Krapu, G.L., Cox, R.R., 2011. Changes in nutrient dynamics of midcontinent greater white‐fronted geese during spring migration. J. Wildl. Manag. 75, 1716-1723. .
QGIS, 2023. QGIS Geographic Information System. QGIS Association. .
R Core Team, 2023. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria.
Rantanen, M., Karpechko, A.Y., Lipponen, A., Nordling, K., Hyvärinen, O., Ruosteenoja, K., et al., 2022. The Arctic has warmed nearly four times faster than the globe since 1979. Commun. Earth Environ. 3, 168. .
Rosin, Z.M., Skórka, P., Wylegała, P., Krąkowski, B., Tobolka, M., Myczko, Ł., et al., 2012. Landscape structure, human disturbance and crop management affect foraging ground selection by migrating geese. J. Ornithol. 153, 747-759. .
Rozenfeld, S.B., Volkov, S.V., Rogova, N.V., Kirtaev, G.V., Soloviev, M.Y., 2021. The impact of changes in breeding conditions in the Arctic on the expansion of the Russian population of the Barnacle Goose (Branta leucopsis). Biol. Bull. 48, 1528-1540. .
Ruokonen, M., Litvin, K., Aarvak, T., 2008. Taxonomy of the bean goose–pink-footed goose. Mol. Phylogenet. Evol. 48, 554-562. .
Saemangum Developmental Agency, 2024. History of Saemangum project. . (Accessed 1 May 2024) [in Korean].
Schmutz, J.A., Laing, K.K., 2002. Variation in foraging behavior and body mass in broods of Emperor Geese (Chen Canagica): evidence for interspecific density dependence. Auk 119, 996-1009. .
Shimada, T., Mori, A., Tajiri, H., 2019. Regional variation in long-term population trends for the Greater White-fronted Goose Anser albifrons in Japan. Wildfowl 69, 105-117.
Si, Y., Xu, Y., Xu, F., Li, X., Zhang, W., Wielstra, B., et al., 2018. Spring migration patterns, habitat use, and stopover site protection status for two declining waterfowl species wintering in China as revealed by satellite tracking. Ecol. Evol. 8, 6280-6289. .
Statistics Korea, 2023. Farmland status. . (Accessed 6 January 2024).
Sutherland, W.J., 1996. Ecological Census Techniques: A Handbook. Cambridge University Press, Cambridge, UK.
Tombre, I.M., Oudman, T., Shimmings, P., Griffin, L., Prop, J., 2019. Northward range expansion in spring‐staging barnacle geese is a response to climate change and population growth, mediated by individual experience. Glob. Chang. Biol. 25, 3680-3693. .
Toral, G.M., Figuerola, J., 2010. Unraveling the importance of rice fields for waterbird populations in Europe. Biodivers. Conserv. 19, 3459-3469. .
Terrill, S.B., 1990. Food availability, migratory behavior, and population dynamics of terrestrial birds during the nonreproductive season. Stud. Avian Biol. 13, 438-443.
Walker, J., Taylor, P.D., 2017. Using eBird data to model population change of migratory bird species. Avian Conserv. Ecol. 12, 4. .
Wetlands International, 2024a. International Waterbird Census. . (Accessed 12 July 2024).
Wetlands International, 2024b. Waterbird Population Estimates. Wetlands International, Ede, the Netherlands. . (Accessed 7 January 2024).
Wickham, H., 2016. ggplot2: Elegant Graphics for Data Analysis. Springer-Verlag, New York.
Wood, S.N., 2017. Generalized Additive Models: An Introduction with R, second ed. Chapman and Hall/CRC, New York.
Zhang, Y., Jia, Q., Prins, H.H.T., Cao, L., de Boer, W.F., 2015. Effect of conservation efforts and ecological variables on waterbird population sizes in wetlands of the Yangtze River. Sci. Rep. 5, 17136. .
Zhao, J.-M., Lyu, N., Cheng, W., Zhou, L.-Z., 2020. Cryptic competition from neighbours significantly reduces intake rate of foraging greater white-fronted geese. Anim. Behav. 167, 119-126. .
Zhao, M.J., Cao, L., Fox, A.D., 2010. Distribution and diet of wintering Tundra bean geese Anser fabalis serrirostris at Shengjin lake, Yangtze river floodplain, China. Wildfowl 60, 52-63.
Zheng, R., Smith, L., Prosser, D., Takekawa, J., Newman, S., Sullivan, J., et al., 2018. Investigating home range, movement pattern, and habitat selection of Bar-headed Geese during breeding season at Qinghai Lake, China. Animals 8, 182. .
Table
1.
Results of the generalized additive mixed models (GAMMs) with temporal (month) and spatial (latitude) variables affecting Greater White-fronted (Anser albifrons; GWFG) and Bean Geese (Anser fabalis sensu lato; BEAG) population sizes in South Korea. Tundra Anser fabalis serrirostris (TDBG) and Taiga Bean Geese A. f. middendorffii, (TGBG) were estimated based on their wintering distribution, not by census data.
Species
Model
Model term
Coeff.
SE
EDFa
F
t
p
GWFG
Monthly
Intercept
116810
7115
16.42
<0.001
Month
3.57
29.79
<0.001
Seasonb
6.18
3.40
<0.001
Latitudinal
Intercept
1969.70
291.20
6.76
<0.001
Latitudec × Month
474.37
20.25
<0.001
Seasonb
7.48
3.19
<0.001
TDBG
Monthly
Intercept
61178
3003
20.37
<0.001
Month
1.00
133.75
<0.001
Seasonb
0.61
0.08
0.391
TGBG
Monthly
Intercept
4873.40
371.80
13.11
<0.001
Month
3.97
29.23
<0.001
Seasonb
4.65
1.39
<0.05
BEAG
Latitudinal
Intercept
1147.69
57.32
20.02
<0.001
Latitudec × Month
412.27
21.04
<0.001
Seasonb
1.76
0.27
0.274
a Number of the effective degrees of freedom. b Random effect. c Values from the centroid of each survey site.


 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds