Ruben C. Fijn, Wouter Courtens, Bas W.R. Engels, Emiel E. van Loon, Judy Shamoun-Baranes. 2024: Combining video and GPS-tracking to study the spatial foraging distribution of a single-prey loading seabird. Avian Research, 15(1): 100196. DOI: 10.1016/j.avrs.2024.100196
Citation:
Ruben C. Fijn, Wouter Courtens, Bas W.R. Engels, Emiel E. van Loon, Judy Shamoun-Baranes. 2024: Combining video and GPS-tracking to study the spatial foraging distribution of a single-prey loading seabird. Avian Research, 15(1): 100196. DOI: 10.1016/j.avrs.2024.100196
Ruben C. Fijn, Wouter Courtens, Bas W.R. Engels, Emiel E. van Loon, Judy Shamoun-Baranes. 2024: Combining video and GPS-tracking to study the spatial foraging distribution of a single-prey loading seabird. Avian Research, 15(1): 100196. DOI: 10.1016/j.avrs.2024.100196
Citation:
Ruben C. Fijn, Wouter Courtens, Bas W.R. Engels, Emiel E. van Loon, Judy Shamoun-Baranes. 2024: Combining video and GPS-tracking to study the spatial foraging distribution of a single-prey loading seabird. Avian Research, 15(1): 100196. DOI: 10.1016/j.avrs.2024.100196
Waardenburg Ecology, Department of Bird Ecology, Culemborg, the Netherlands
b.
University of Amsterdam, Institute for Biodiversity and Ecosystem Dynamics, Amsterdam, the Netherlands
c.
Research Institute for Nature and Forest (INBO), Brussels, Belgium
Funds:
GPS-tracking of Sandwich Terns was part of the monitoring programme into the effects of the compensation measures designed for the construction of the seaward expansion of the Rotterdam Harbour (‘Tweede Maasvlakte’). This programme (PMR-NCV) was initiated by the Dutch Ministry of Infrastructure and the Environment and commissioned by Rijkswaterstaat WVL (M. van Eerden, K. Borst). Bureau Waardenburg (BW), the Research Institute for Nature and Forest (INBO), and Deltamilieu Projecten (DMP) carried out the research on terns within a project commissioned by Wageningen Marine Research (a.o. I. Tulp) and Deltares (a.o. T. Prins). Tracking of Sandwich Terns was performed under project licence for animal procedures AVD401002015102 of the Central Authority for Scientific Procedures on Animals. Camera monitoring was part of a research grant to DMP and BW by Vogelbescherming Nederland. Project manager was F. Arts (DMP) and he is greatly thanked for his input. Fieldwork in both projects was carried in a nature reserve of Natuurmonumenten and W. van Steenis, H. Meerman, J. de Roon and M. Broere are specifically acknowledged for their cooperation and hospitality. The authors would like to thank T. Boudewijn, J. de Jong, E. Bravo Rebolledo, A. Gyimesi, M. Poot (BW), E. Stienen, H. Verstraete, M. Van der walle and N. Vanermen (INBO) for help in the field. The INBO team is specifically thanked for their observations of prey items. R. Middelveld (BW) is thanked for providing the map with foraging locations. The comments of two anonymous referees and Giacomo Dell'Omo greatly improved this manuscript
Seabirds are valuable indicators of marine ecosystem processes and studying seabird diets can shed light on natural or human-induced variability in food-web composition. Specifically single-prey loading seabird species such as terns have the potential to act as visual sentinels of prey availability offshore. However, obtaining diet information from remote bird colonies is often challenging and time consuming. In this pilot study we present a novel approach to combine two established methods to study seabird foraging ecology, providing a powerful and cost-effective tool to study the distribution of prey items available to seabirds. We combined GPS tracking data of Sandwich Terns (Thalasseus sandvicensis) with prey-observations from a hide in 2012 and 2013, and from semi-continuously recorded camera footage in 2017. By doing so, we identified 115 approximate catch locations of prey (86 herring/sprat Clupeidae, 29 sandeel Ammodytidae). Combining GPS-data and prey observations yielded detailed knowledge on the movements and chick diets of tracked birds as well as the spatial origin and lengths of captured prey items. Further catch distances of both Clupeidae and Ammodytidae resulted in deliveries of larger prey items and thus higher energy yield per trip, but also a higher energy expenditure per trip. We discuss the limitations and potential of our methodological approach to study foraging energetics during chick-provisioning of seabirds that carry prey items visible in their beaks.
Long-term climate and short-term weather conditions changes can have an important effect on biodiversity (Urban, 2015), especially when environmental conditions are harsher, like in winter (Bélanger and Bédard, 1989; Guzmán et al., 2017). It is well known that both migratory and resident birds can be negatively affected by severe winter weather (Newton, 1998), especially during periods of continuous freezing temperatures accompanied with heavy snow and strong wind, also known as 'cold spells' (De Vries et al., 2012). In these situations, migratory and resident birds might show different responses, so while migratory birds may 'escape' from the cold (Péron et al., 2011), resident birds will endure weather conditions through the regulation of body energy reserves, which are used to evaluate the impact of cold weather on birds' survival (Sánchez-García et al., 2018). In France, Lormeé et al. (2012) evaluated the effects of a cold spell in February 2012, and found 1791 dead birds, the most important causes of death being starvation (56%), vehicle collisions (24%) and predation (8%). Similar conclusions were observed in Greater Flamingos (Phoenicopterus roseus) after several cold spells in southern France (Deville et al., 2014).
A large snowstorm named 'Filomena' affected from central to the northeast of Spain from 5 to January 10, 2021, but mainly Central Spain, outstanding the region of Madrid, which was covered with snow (reaching 50–60 cm of accumulated snow), and was affected by freezing temperatures (the minimum temperatures ranged from −12 to −4.4 ℃), with harsh conditions being extended during a fortnight (AEMET, 2021a, 2022; Smart, 2021). Filomena caused five human casualties and stopped public and private transportation, producing damages not only in buildings and infrastructure, but also to around 500,000 trees in the city of Madrid (Pérez-González et al., 2022). This cold spell was characterized by the amount of snow, being the heaviest snowfall recorded in Madrid since 1971 (Tapiador et al., 2021).
In the current climate change scenario with increased extreme weather events, management measures targeting wildlife could avoid detrimental effects. This is the case of food and water provision, which are common practice in hunting grounds of western Europe targeting small game species such as Partridges (Alectoris and Perdix genera), Wild Rabbit (Orytolagus cuniculus) and Hares (Lepus spp.) (Armenteros et al., 2021; Estrada et al., 2015) together with other measures aiming to provide shelter through land interventions (Sánchez-García et al., 2024). In the case of game species, there are national schemes to halt or restrict hunting during cold spells, mainly for waders and waterfowl (see Sánchez-García et al., 2018 and the review herein).
To the best of our knowledge, few studies have addressed the possible effects of Filomena on birds' population levels, showing displacements and significant changes on abundance depending on the species (Cano-Barbacil and Sánchez, 2022; Pérez-Granados et al., 2023). Aiming to gain knowledge on the impact of this stochastic event on birds, this study evaluated the populations of gamebirds in the region of Madrid, comparing field data gathered at the same locations after the arrival of Filomena (February–March 2021), with the previous year. Our results may help to understand the impact of severe weather conditions in gamebirds and promote management and conservation measures that would benefit gamebirds but also other wild birds (e.g. endangered steppe-bird populations) that occur at the same habitats. As the studied species are gamebirds, hunters and managers conduct management targeting these species, hence our conclusions may be applied to prevent detrimental effects of cold spells.
2.
Materials and methods
2.1
Study area
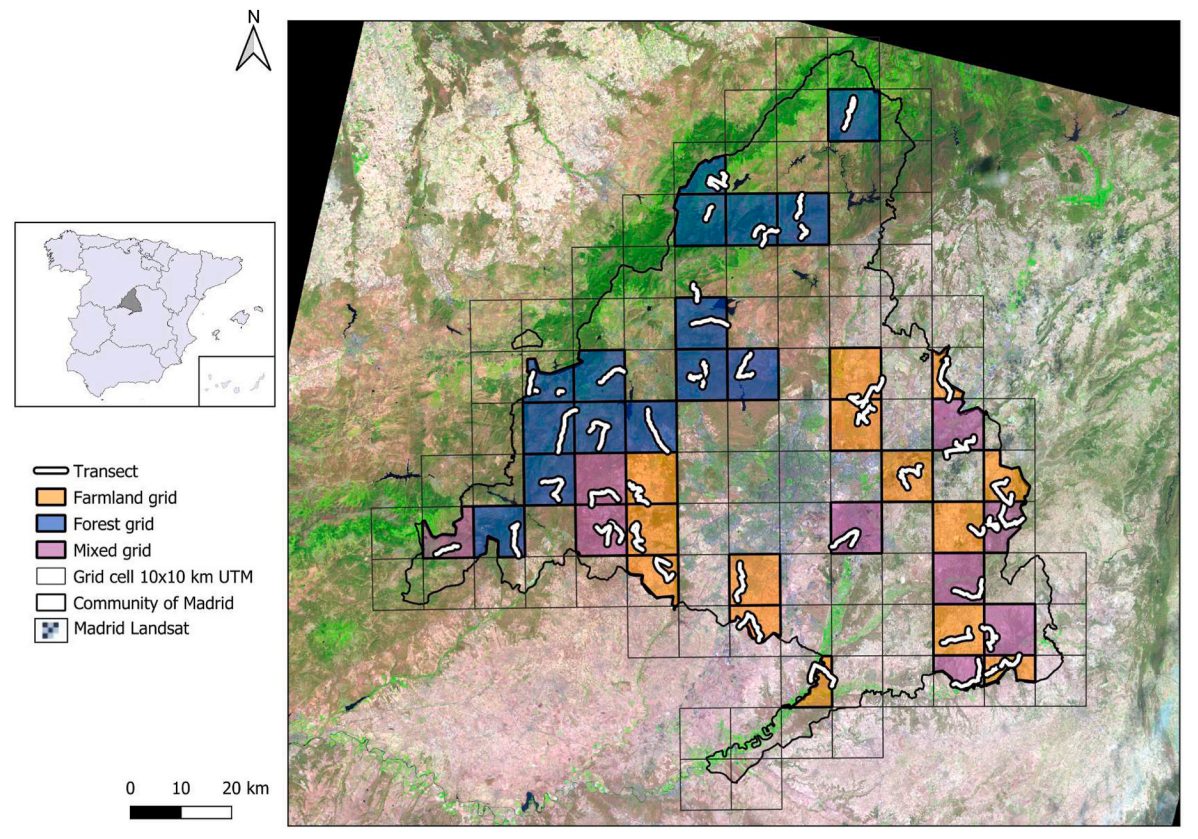
This study was conducted in the region of Madrid, which covers 8028 km2 in Central Spain (Fig. 1). The main landscapes in this region are forest and shrublands (55%), and cultivated areas (29%), mainly cereal but also olive groves. The region of Madrid has two main climates, Mediterranean continental, with hot dry summers, cold winters and a annual mean rainfall of 400–650 mm (though some areas are below 300 mm), and mountain climate restricted to areas ≥1200 m a.s.l., with cold and wet autumn-winter (~1500 mm, including snow), and mild summer (Chazarra et al., 2009).
Figure
1.
Map showing the region of Madrid and the location of the transects conducted during 2020 and 2021.
This study was part of a gamebird monitoring project developed in the region of Madrid during 2020 (year-1, before Filomena) and 2021 (year-2, after Filomena). Gamebird hunting is a legal activity of high socioeconomic importance in Spain and other European countries, with existing regulations in which the game species and hunting periods are indicated to ensure that hunting is sustainable, with active habitat management being conducted by hunters and game managers (Sánchez-García et al., 2024). Of the species hunted in the region of Madrid, three were chosen due to their wide distribution in the study area and the different seasonal movement strategies: (1) Red-legged Partridge (Alectoris rufa), a strictly sedentary gamebird species, which occurs mainly in farmland but also mixed habitats (Casas et al., 2022); (2) Wood Pigeon (Columba palumbus), a sedentary gamebird species but also migrant in Iberia, present in farmland, forest and mixed habitats, and also in urban and peri-urban areas (Purroy and Purroy, 2016; Butkauskas et al., 2019); and (3) Eurasian Magpie (Pica pica), a sedentary species occurring in a wide range of habitats, including urban and peri-urban areas (Martínez, 2011). The magpie is a gamebird, but it is mainly hunted as part of a predator removal activities through a special permission.
A grid cell of 10 × 10 km (UTM) was used only to select a uniform distribution of sampling points within the region of Madrid (Fig. 1). From a total of 113 grid cells, the ones that were covered by more than 80% of urban areas were excluded. From the remaining 83 grids, 50% were randomly selected for sampling (n = 41). The habitat type in each grid was categorized using the National Information System of Land Use (SIOSE, https://www.siose.es/). After excluding urban areas, three main habitat categories were established: 'forest' (when at least 70% of the grid surface was covered by trees and shrubland), 'farmland' (when at least 70% of the grid surface was covered by open-land mostly dedicated to farmland such as cereal fields, ploughed fields, fallow), and 'mixed' when none of the previous applied.
Field work took place in two consecutive years, 2020 and 2021, during February and early March, once the hunting season was finished. At this time of the year, the studied species are in pairs, at the beginning of the breeding season (Rocha et al., 2009; Martínez, 2011; Sánchez-García et al., 2017).
In 2020, we visited each grid in advance, and designed one transect of 10 km in length per grid. Surveys were conducted on consecutive days, in early morning (within 3 h after dawn) and evening (last 3 h before dusk), when birds are more active and detectable. Transects were covered by car at low speed (10 km/h) using local track networks and stopping every time the studied species were located. All gamebirds detected were recorded, and the distance to the observer was measured using a range finder (NUM'axes, Ltd., 6 × 25), together with the angle using a protractor set on the top of the car. Field-work was conducted in the absence of rain, snow or heavy wind which may affect birds' behavior.
2.3
Hunting data
To study the effects of hunting activity on gamebirds during the hunting season right after this extreme cold event, we used the gamebag data of the small game species (number of animals hunted/season) during the hunting seasons of 2019–2020 (before Filomena) and 2020–2021 (after Filomena). In all hunting grounds it is mandatory to report gamebag data at the end of each season to the regional administration. For this analysis, we used the data provided by the regional government of Madrid (which is compiled at national scale; MITERD, 2023).
The open season for the studied species covers two main periods: from October to February, when the three studied species can be hunted, and from mid-August to mid-September, when Wood Pigeons and Magpies can be hunted (but not Partridges). Also, as part of predator removal management, Magpies can be hunted in February, though a special permission must be given by the regional government. Red-legged Partridges are released for hunting or repopulation purposes (Sánchez-García et al., 2022), but according to official data, most of farm-reared birds are released in commercial sites, hence for the analysis only gamebag data from sites without releasing were used.
Data was made available at hunting ground scale, but taking into account that transects covered more than one hunting ground, the home range of the species (Buenestado et al., 2008; Martínez, 2011; Pérez et al., 2015), and the migratory behavior of Wood Pigeons (Schumm et al., 2022), data was merged to evaluate hunting pressure at regional level.
2.4
Data analysis
For each target species and hunting season, it was calculated the hunting rate to address whether it varied among the two consecutive hunting seasons, understood as the proportion of animals of a given species from the total gamebag.
To estimate the impact of Filomena snowstorm, a Snow Persistence Index (SPI) was computed using Sentinel-2 satellite images (UTM 30T tiling grids UL, UK, VL, VK). The images selected were those of January 13 and 18, 2021 due to the low percentage of cloud cover (< 10%) after the main snowfall on January 8 and 9, 2021. The Normalized Difference Snow Index (NDSI) was used, and the positive values were chosen to select the locations with the highest probability of snow presence. The SPI was calculated as the mean NDSI between both dates, so that values close to 1 (blue) indicated that the snow was persistent, remained in that location for the entire period, values close to 0 (red) corresponded to locations where snow did not occur, and intermediate values (yellow) meant that the snow melted during that time interval (Fig. 2).
Figure
2.
(A) Normalized Difference Snow Index (NDSI), of January 13, 2021; (B) NDSI of January 18, 2021 and (C) Snow Persistence Index (SPI) as mean for both values of NDSI. Blue colors represent high SPI (1) and warm ones low SPI (0). (For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)
Bird abundance was calculated through a Kilometric Abundance Index (KAI), defined as the ratio of the total number of individuals of a given species observed along a transect by the total transect length covered at each site (Preatoni et al., 2012). Possible differences on gamebird abundance among years, landscapes, and snow persistence were studied using generalized linear models (GLM) with negative binomial errors to control data overdispersion. Likewise, to address whether there were differences in hunting between the two hunting seasons in the region of Madrid, a GLM with negative binomial errors was applied using species and season as the explanatory variable and the number of individuals hunted as the response variable.
We assumed that the difference in the number of individuals hunted in a hunting season has an impact on the species population (KAI) in the following year. To calculate the expected abundance in the absence of Filomena (expected KAI values in 2021), we understood that the ratio of individuals hunted in both hunting seasons could be reflected in the ratio of abundance indices (KAI values in 2020 and 2021) but inversely, as the reduction of hunting in the 2020–2021 season should increase the KAI values in 2021. This estimation allowed us to analyze the effect of Filomena by comparing the value of expected KAI in 2021 due to the reduction of individuals hunted in the 2020–2021 season with the observed KAI value in 2021 after Filomena occurred. All statistical analyses were conducted using R software v.3.4 (R Core Team, 2023), specifically MASS package (Venables and Ripley, 2002). The confidence intervals (CI) were estimated using the 95% region. We also used the interval between the 25 and 75 percentiles (IQi) as a robust estimation of the variation of the values due to non-normality of the data.
3.
Results
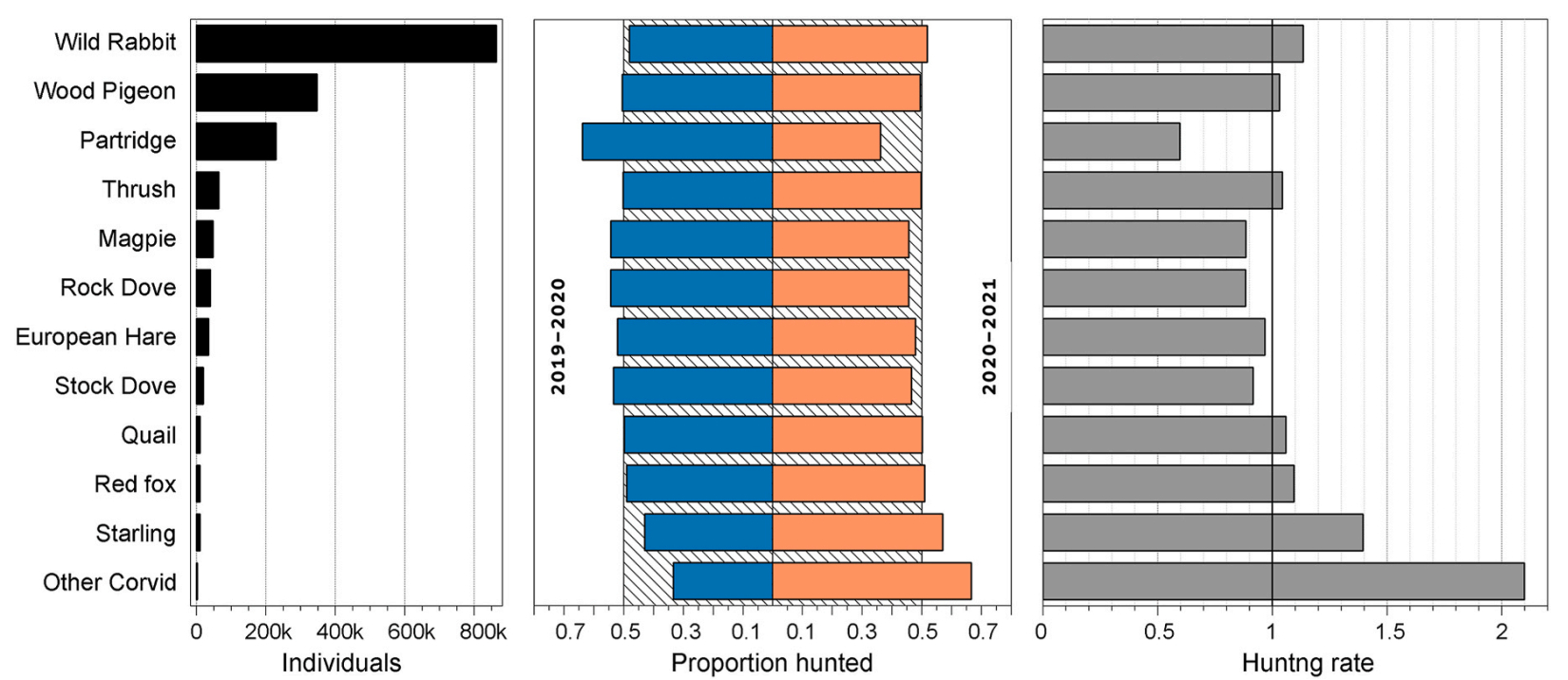
In the region of Madrid, the hunting bag of small game species in the 2020–2021 season was slightly lower when Filomena occurred than in the previous season (after Filomena, 95.1% of the 2019–2020 season). The GLM applied to study the hunting bag of the small game species in the two hunting seasons showed significant differences by species but no between hunting seasons, hence hunting was not influence by the season (P = 0.874). When looking at the studied species, the gamebag was reduced in all of them in the 2020–2021 season (Fig. 3.). The number of Partridges hunted in 2020–2021 was 56.8% of those caught in the previous season, and the hunting rate was 59.7%. In the same way, Magpie gamebag was reduced to 84.1% and the hunting rate to 88.4%. By contrast, in the case of Wood Pigeon, the gamebag (i.e. the number of individual hunted) decreased slightly to 98.1% in the 2020–2021 season, though was recorded a slight increase (3.1%) in hunting rate i.e. the number of Wood Pigeons hunted with respect to the total number of small game hunted was slightly higher.
Figure
3.
Small game bag data from the region of Madrid, for the seasons 2019–2020 and 2020–2021. (A) Number of animals hunted per species and season (gamebag); (B) proportion of animals hunted per species and season (stripped area indicates equal number of animals per season); (C) hunting rate for the two seasons (vertical line in "1" indicates equal proportion in the two seasons).
The SPI values ranged from 0.280 (low snow persistence) to 0.932 (high snow persistence) considering all sampled locations (Appendix Fig. S1). However, in farmland the mean SPI values were 0.855 (IQi: 0.865–0.917), while in forest the mean values were 0.671 (IQi: 0.596–0.810), hence snow prevailed longer in farmland.
The abundance analysis of the studied species, including year, SPI and habitat type using the GLM analysis (Table 1) showed no significant differences between years. Also, the SPI variable was not significant, and this information already provided by the habitat type variable. When considering only habitat type in the GLM analysis, the data revealed strong effect of habitat type on Wood Pigeon (P < 0.001) and Partridge abundance (P < 0.001), together with a moderate effect on Magpie abundance (P = 0.012).
Table
1.
The two GLM models applied to the data of Wood Pigeon, Red-legged Partridge, and Eurasian Magpie using the variables: habitat (forest/non-forest), year (2021/2020), and SPI.
Wood Pigeon
Red-legged Partridge
Eurasian Magpie
Estimate
S.E.
P
Estimate
S.E.
P
Estimate
S.E.
P
Intercept
1.934
1.106
0.080
−2.808
2.007
0.162
0.533
0.891
0.550
Habitat
−2.252
0.446
< 0.001
−2.101
0.742
0.005
−0.605
0.348
0.082
Year
−0.336
0.344
0.328
−0.060
0.554
0.913
−0.477
0.277
0.085
SPI
−0.948
1.255
0.450
2.592
2.289
0.257
0.963
1.010
0.341
Estimate
S.E.
P
Estimate
S.E.
P
Estimate
S.E.
P
Intercept
1.053
0.209
< 0.001
−0.604
0.059
0.059
1.148
0.173
< 0.001
Habitat
−2.284
0.387
< 0.001
−2.455
0.620
< 0.001
−0.759
0.304
0.0124
Note: the columns correspond to: coefficients (Estimate), standard error (SE), and probability (P).The coeffcient estimates with significant differences (P < 0.05) are in bold type.
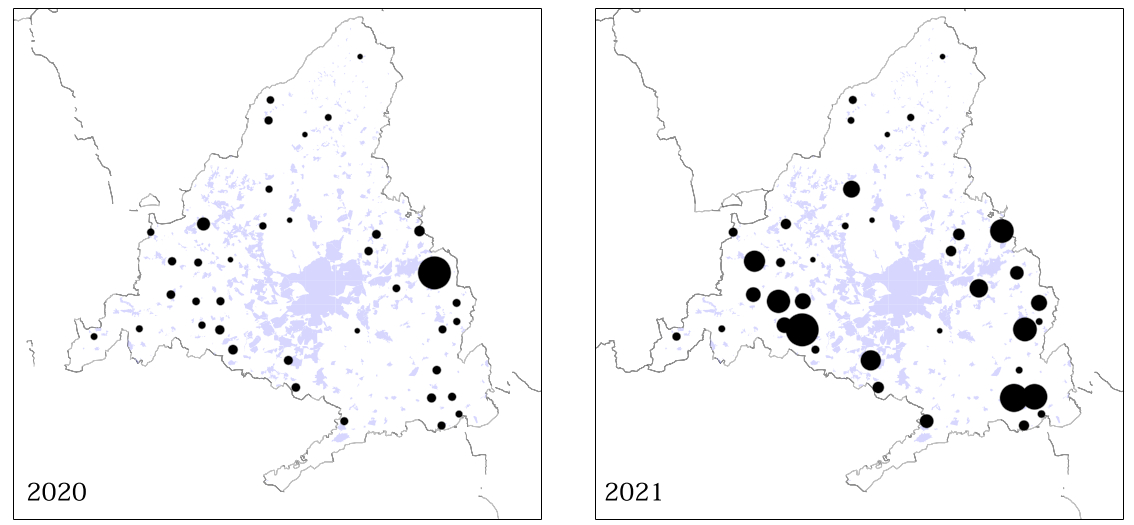
Wood Pigeons were detected in 88.6% of transects conducted in farmland and in 50% of those conducted in forest (Appendix Fig. S2). The abundance value in farmland was 4.8 times higher (median: 1.49 birds/km, IQi: 0.83–4.94 birds/km) compared to forest (median: 0.31 birds/km, IQi: 0.13–0.53 birds/km). Similarly, Partridges were observed in 47.8% of farmland transects and in 22.8% of forest transects, with 5 times higher abundance values in farmland (median: 0.55 birds/km, IQi: 0.40–1.31 birds/km) compared to forest (median: 0.11 birds/km, IQi: 0.10–0.27 birds/km). However, Magpies showed a larger distribution because they were observed in 93.2% of transects conducted in farmland and in 81.8% of those conducted in forest, although with a higher abundance (2.3 times) in farmland (median: 2.35 birds/km, IQi: 1.31–3.61 birds/km) compared to forest (median: 1.02 birds/km, IQi: 0.59–2.17 birds/km). For this species, although no significant differences in the abundance was recorded among years (P = 0.077), we found a higher variability between sampling points after Filomena (year 1, IQ: 1.62; year 2, IQ: 2.84). As shown in Fig. 4, Magpies tended to aggregate in higher numbers in the 20–21 hunting season compared to the previous.
Figure
4.
Kilometric Abundance Index (KAI) for Magpies at the sampled locations for the 2019–2020 and 2020–2021 seasons.
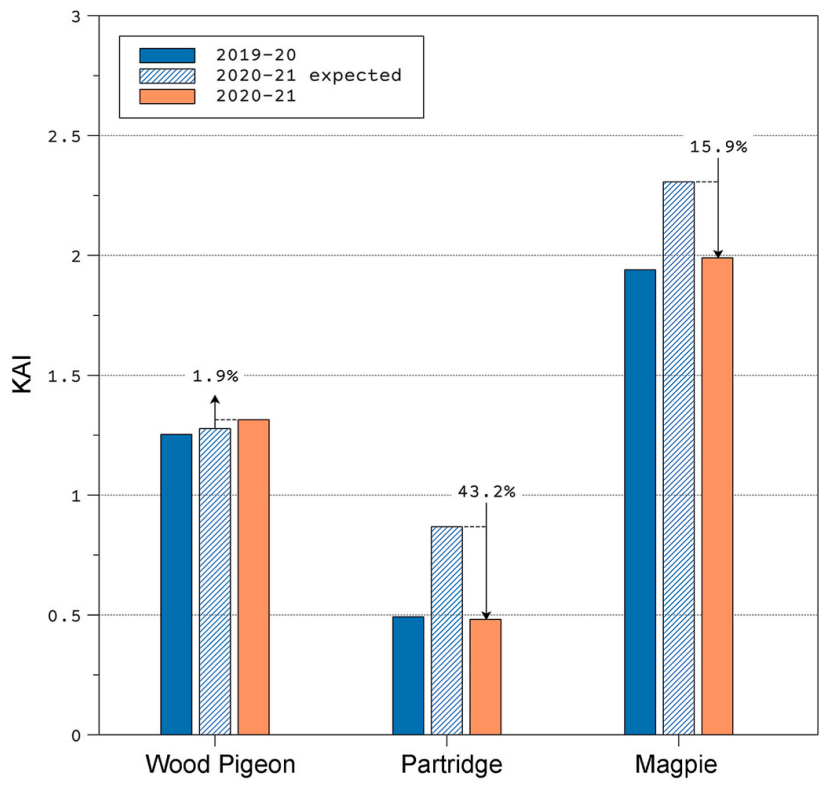
The "expected abundance" calculated in the absence of Filomena for the 2020–2021 predicted that in the absence of Filomena, KAI values would have been 1.9% higher in Pigeons, 43.2% higher in Partridges and 15.9% higher in Magpies (Fig. 5).
Figure
5.
Kilometric Abundance Index (KAI) of the three studied species for the 2019–2020 and 2020–2021 seasons, together with the expected KAI value (stripped histogram) considering the number of animals hunted in the 2020–2021 season.
This study has shown no significant differences in abundance between the two studied periods for the three species. Considering hunting rates, our results suggest that the abundance of Partridge's was negatively affected by Filomena, while no effect was found in Wood Pigeons and a moderate effect (but not negligible) was recorded in Magpie's abundance.
Owing to the mobility restrictions during and after the COVID-19 pandemics (Henríquez et al., 2020), we hypothesized that hunting could had been reduced in all species during the 2020–2021 season, but it was only significantly reduced in Partridges, which could be explained by the shorter period of time when Partridges were hunted compared to the other species, as Wood Pigeons and Magpies were also hunted from mid-August to mid-September, and the latter species can be controlled in February. It is important to note that in Madrid the hunting season finishes by the end of January, and the majority of hunting grounds decided to stop hunting after Filomena to avoid harming gamebird populations (Lara, pers. comm), reducing the hunting season (one month less than in the year before Filomena). In Spain and other European countries, small game hunting is forbidden when the snow covers the ground (Agencia Estatal Boletín Oficial del Estado, 2015). Interestingly enough, during both seasons, the Wood Pigeon gamebag was similar, but a small increase in the hunting rate was recorded during 2020–2021 season. These results were similar for other game species (Fig. 3), which suggests that reducing hunting in some species may increase the hunting rate in others.
The occurrence and abundance of the three species varied among habitats, though all species showed higher abundances in farmland compared to forest. As described by the Spanish meteorological agency, the snowfall was heaviest in lowland areas, dominated by farmland (AEMET, 2022), but no clear habitat "shifts" to forest areas were detected. Despite Wood Pigeons being associated to habitats dominated by forest (Purroy and Purroy, 2016), they reached higher densities in farmland, which could be explain by the adaptation to urban and peri-urban habitats during the last decades observed in Spain and other European areas (Purroy and Purroy, 2016). Interestingly enough, after the arrival of Filomena, Magpies tended to aggregate (Fig. 4), which could be explained by an anti-predatory strategy when the weather is harsh, as described previously in urban habitats of Italy (Ientile, 2014).
Overall, our results show that the abundance of the studied gamebirds in the region of Madrid did not change across the study periods, but the expected abundance in the absence of Filomena (Fig. 5) points towards the idea of different responses depending on the species.
In Wood Pigeons, which were subject to slightly higher hunting rate in the 2020–2021 period, it is possible that they managed to escape from the severe weather, coming back to their sites once weather conditions improved, a pattern already described in the species (Schumm et al., 2022). Magpies, which are sedentary and were subject to lower hunting rate in 2020–2021, could have merged in flocks and search of habitats with better conditions within Madrid (Ientile, 2014), showing a certain pattern of aggregation, which may had alleviated detrimental effects of food depravation from "frozen grounds", making the most of available food at urban and peri-urban habitats, such as carrion from wildlife road-kills (Martínez, 2011). In Magpies and Wood Pigeons, it is important to note that they are able to perch and roost in trees, bushes and buildings of different size, this behavior being less frequent in Red-legged Partridges (Potts, 2012).
Quite the opposite, results suggest that Partridges were negatively affected by Filomena, as no abundance difference was found between years, but in the 2020–2021 hunting season, there was a dramatic hunting reduction (56%) compared to the previous. Together with shooting restraint, works at large scale were conducted to clear roads and tracks from snow, which may had alleviated food deprivation owing to snow persistence and frost, and hunters and game managers provided food at many hunting grounds (Pompa, 2022), a very common practice for small game species in Spain (Sánchez-García et al., 2024). In other words, if Filomena had not affected Partridges with half of the hunting pressure and measures "on the ground" against snow and frost, we would have expected abundance to be higher in 2021 (Fig. 5), which was not the case. It is true that we do not know the Partridge breeding success in 2020, but in Partridge populations located at close distance, productivity between these two periods was similar (authors, data not published). Moreover, in Madrid the springtime was warm and wet (AEMET, 2021b), conditions which favor Partridges (Lucio, 1990), with no severe storms recorded in Madrid during that period that could have reduced productivity. During the first days after the arrival of Filomena, some hunters and game managers shared photographs of dead wildlife in social media (including small game species), that could have died from starvation or freezing temperatures, collisions or predation, but no systematic studies were conducted.
Our results are partially in agreement with Cano-Barbacil and Sánchez (2022), who studied the impact of Filomena in farmland birds in the region of Madrid. These authors found that the populations of sedentary species such as Thekla's Lark (Galerida theklae) and Corn Bunting (Emberiza calandra), were decreased after the snowstorm, while other migrants, such as Eurasian Lapwing (Vanellus vanellus), Eurasian Skylark (Alauda arvensis) and Meadow Pipit (Anthus pratensis), were not significantly affected owing to their migratory behavior, coming back to their territories once weather conditions improved and snow had melted. Additionally, these authors suggest that the Crested Lark (Galerida cristata), which is not a migratory species, moved to southern locations, being recorded back to their territories as the other species. In addition, Pérez-Granados et al. (2023), who studied the endangered Dupont's Lark (Chersophilus duponti) in several locations affected by Filomena, found a significant population decline in the next reproductive season.
Existing research in other bird species at other European countries has confirmed that birds are often badly affected by cold spells and snowstorms, though these studies were conducted in waterfowl and waders (shorebirds), using different methodologies, such as the collection of dead birds (Davidson and Clark, 1985; Péron et al., 2011; Lormeé et al., 2012). As waterfowl and the majority of waders occur in aquatic environments (wetlands, shorelines), they are very sensitive to severe winter weather (Lormeé et al., 2012), with a marked migratory behavior when environmental conditions get harsh, as demonstrated in species such as the Eurasian Woodcock (Scolopax rusticola) (Péron et al., 2011) and the Eurasian Golden Plover (Pluvialis apricaria) (Machín et al., 2015).
5.
Conclusions
Stochastic weather, such as the snowstorm Filomena, poses wild birds at risk, with different effects depending on the severity and the ability to endure or escape from the harsh weather from a species given. Then, three types of responses to extreme weather events may have occurred: (1) migratory species such as Pigeons responding with a temporary flight from the affected area avoiding mortality at the cost of travel expenses; (2) sedentary species with limited movement capacity remaining within their home range at the cost of high mortality as Partridges; and (3) sedentary species with certain movement capacity, regrouping in favorable areas that allows them to reduce the cost of mortality, such as Magpies.
Owing to the difficulties on predicting these events, in the case of gamebirds the two main actions to take are hunting restraint and habitat management. In Spain, around 90% of the land is declared as 'hunting ground' (Sánchez-García et al., 2021), hence the role of hunters and game managers through supplementary feeding, shelter and clearing the ground from snow, could be crucial not only for gamebirds, but also for other wildlife species. Special attention should be paid to sedentary species which may not be able to escape from harsh weather (such as Partridges), and regular game and wildlife monitoring schemes should be promoted to fully understand the effects of these events, and take management decisions accordingly (https://www.observatoriocinegetico.org).
Ethics statement
This study did not involve handling animals and monitoring was approved by the Regional Government of Madrid.
The authors declare no conflict of interest, either competing financial or personal interests that could have influenced the work reported in this paper.
Acknowledgements
We are indebted to all hunters and managers who allowed access to hunting grounds and the Hunters' Federation of Madrid for providing contacts. We thank to J.L. and L.E. for sharing from the Área de Conservación de Flora y Fauna (Comunidad de Madrid), and N.W.S. for his linguistic revision and advice. Reviewers provided useful comments which improved this manuscript.
Andrews Ⅲ, A.G., Strasburger, W.W., Farley Jr, E.V., Murphy, J.M., Coyle, K.O., 2016. Effects of warm and cold climate conditions on capelin (Mallotus villosus) and Pacific herring (Clupea pallasii) in the eastern Bering Sea. Deep Sea Res. Part Ⅱ 134, 235–246. .
Barrett, R.T., Camphuysen, K., Anker-Nilssen, T., Chardine, J.W., Furness, R.W., Garthe, S., et al., 2007. Diet studies of seabirds: a review and recommendations. ICES J. Mar. Sci. 64, 1675–1691. .
Bates, D., Mächler, M., Bolker, B., Walker, S., 2015. Fitting linear mixed-effects models using the lme4 package in R. J. Stat. Software 67, 1–48.
Beal, M., Byholm, P., Lötberg, U., Evans, T.J., Shiomi, K., Åkesson, S., 2021. Habitat selection and foraging site fidelity in Caspian Terns (Hydroprogne caspia) breeding in the Baltic Sea. Ornis Fenn. 98, 128–141.
Bennet, D.G., Horton, T.W., Goldstien, S.J., Rowe, L., Briskie, J.V., 2019. Flying south: foraging locations of the Hutton's shearwater (Puffinus huttoni) revealed by Time‐Depth Recorders and GPS tracking. Ecol. Evol. 9, 7914–7927. .
Bicknell, A.W., Oro, D., Camphuysen, K., Votier, S.C., 2013. Potential consequences of discard reform for seabird communities. J. Appl. Ecol. 50, 649–658.
Bond, A.L., Jones, I.L., 2009. A practical introduction to stable-isotope analysis for seabird biologists: approaches, cautions and caveats. Mar. Ornithol. 37, 183–188.
Brenninkmeijer, A., Stienen, E.W.M., Klaassen, M., Kersten, M., 2002a. Feeding ecology of wintering terns in Guinea Bissau. Ibis 144, 602–613. .
Brenninkmeijer, A., Doeglas, G., De Fouw, J., 2002b. Foerageergedrag van sterns in de
westelijke Westerschelde in 2002. Altenburg en Wymenga rapport 346. Feanwâlden,
Netherlands.
Buckingham, L., Daunt, F., Bogdanova, M.I., Furness, R.W., Bennett, S., Duckworth, J., et al., 2023. Energetic synchrony throughout the non‐breeding season in common guillemots from four colonies. J. Avian Biol. 2023, e03018 .
Burger, A.E., Shaffer, S.A., 2008. Perspectives in ornithology application of tracking and data-logging technology in research and conservation of seabirds. Auk 125, 253–264. .
Camphuysen, C.J., Garthe, S., 2000. Seabirds and commercial fisheries: population
trends of piscivorous seabirds explained. In: Kaiser, M.J., de Groot, S.J. (Eds.), The
Effects Offishing on Non-target Species and Habitats: Biological, Conservation and
Socio-Economic Issues. Blackwell, pp. 163–184.
Christensen-Dalgaard, S., 2018. Drivers of Seabird Spatial Ecology – Implications for
Development of Offshore Wind-Power in Norway. PhD thesis. Norwegian University
of Science and Technology, Trondheim, Norway.
Collins, P.M., Halsey, L.G., Arnould, J.P., Shaw, P.J., Dodd, S., Green, J.A., 2016. Energetic consequences of time‐activity budgets for a breeding seabird. J. Zool. 300, 153–162. .
Conners, M.G., Michelot, T., Heywood, E.I., Orben, R.A., Phillips, R.A., Vyssotski, A.L., et al., 2021. Hidden Markov models identify major movement modes in accelerometer and magnetometer data from four albatross species. Mov. Ecol. 9, 7.
Courtens, W., Verstraete, H., Vanermen, N., Stienen, E.W.M., 2017. Faecal samples reveal the diet of breeding adult Sandwich terns Thalasseus sandvicensis in Belgium and the southern part of The Netherlands. J. Sea. Res. 127, 182–193.
Edney, A.J., Wood, M.J., 2021. Applications of digital imaging and analysis in seabird monitoring and research. Ibis 163, 317–337.
Ellis, H.I., Gabrielsen, G.W., 2002. Energetics of free-ranging seabirds. In: Schreiber, B.
A., Burger, J. (Eds.), Biology of Marine Birds. CRC Press, Boca Raton, pp. 359–407.
Engvall, E., Waldenström, J., Hentati-Sundberg, J., 2022. Diet and prey size preference in Razorbills Alca torda breeding at Stora Karlsö, Sweden. Ornis Svec. 32, 87–98.
Evans, T.J., Kadin, M., Olsson, O., Åkesson, S., 2013. Foraging behaviour of common murres in the Baltic Sea, recorded by simultaneous attachment of GPS and time-depth recorder devices. Mar. Ecol. Prog. Ser. 475, 277–289.
Fijn, R.C., de Jong, J., Courtens, W., Verstraete, H., Stienen, E.W.M., Poot, M.J.M., 2017. GPS-tracking and colony observations reveal variation in offshore habitat use and foraging ecology of breeding Sandwich Terns. J. Sea Res. 127, 203–211.
Fijn, R.C., Gyimesi, A., 2018. Behaviour related flight speeds of Sandwich Terns and their implications for wind farm collision rate modelling and impact assessment. Environ. Impact Assess. Rev. 71, 12–16.
Fijn, R.C., Courtens, W., Arts, F.A., Daelemans, R., Grutters, B., et al., 2019. PMR-NCV onderzoek sterns in de Delta en Voordelta. Bureau Waardenburg Rapportnr , 17–247.
Fijn, R.C., Van Bemmelen, R.S.A., Collier, M.P., van Loon, E.E., Poot, M.J.M., Shamoun-Baranes, J., 2024. Evaluation of tag attachment techniques for plunge-diving terns. Ibis 166, 1003–1022.
Fort, J., Porter, W.P., Grémillet, D., 2011. Energetic modelling: a comparison of the different approaches used in seabirds. Comp. Biochem. Physiol. A 158, 358–365.
Furness, R.W., 1990. A preliminary assessment of the quantities of Shetland sandeels taken by seabirds, seals, predatory fish and the industrial fishery in 1981–83. Ibis 132, 205–217.
Furness, R.W., Camphuysen, K., 1997. Seabirds as monitors of the marine environment. ICES J. Mar. Sci. 54, 726–737.
Gaglio, D., Cook, T.R., Connan, M., Ryan, P.G., Sherley, R.B., 2017. Dietary studies in birds: testing a non invasive method using digital photography in seabirds. Methods Ecol. Evol. 8, 214–222.
Gaglio, D., Cook, T.R., McInnes, A., Sherley, R.B., Ryan, P.G., 2018. Foraging plasticity in seabirds: a non-invasive study of the diet of greater crested terns breeding in the Benguela region. PLoS One 13, e0190444.
Garthe, S., Montevecchi, W.A., Davoren, G.K., 2007. Flight destinations and foraging behaviour of northern gannets (Sula bassana) preying on a small forage fish in a low-Arctic ecosystem. Deep Sea Res. Part Ⅱ 54, 311–320.
Hornset, K., 2017. Monitoring Daily Activity and Parental Food Provisioning in the
Atlantic Puffin (Fratercula Arctica) by the Use of Camera Traps. MSc.-thesis.
Norwegian University of Life Sciences, Ås, Norway.
Horswill, C., Jackson, J.A., Medeiros, R., Nowell, R.W., Trathan, P.N., O'Connell, T.C., 2018. Minimising the limitations of using dietary analysis to assess foodweb changes by combining multiple techniques. Ecol. Indicat. 94, 218–225.
Huffeldt, N.P., Merkel, F.R., 2013. Remote time-lapse photography as a monitoring tool for colonial breeding seabirds: a case study using thick-billed murres (Uria lomvia). Waterbirds 36, 330–341.
Iglewicz, B., Hoaglin, D.C., 1993. How to Detect and Handle Outliers. ASQC Quality Press, Milwaukee.
Kabra, K., Xiong, A., Li, W., Luo, M., Lu, W., Garcia, R., et al., 2022. Deep object
detection for waterbird monitoring using aerial imagery. In: 2022 21st IEEE
International Conference on Machine Learning and Applications (ICMLA),
pp. 455–460. Nassau, Bahamas, 2022.
Kato, A., Naito, Y., Watanuki, Y., Shaughnessy, P.D., 1996. Diving pattern and stomach temperatures of foraging king cormorants at subantarctic Macquarie Island. Condor 98, 844–848.
Kross, S.M., Nelson, X.J., 2011. A portable low-cost remote videography system for monitoring wildlife. Methods Ecol. Evol. 2, 191–196.
Kuznetsova, A., Brockhoff, P.B., Christensen, R.H.B., 2017. lmerTest package: tests in linear mixed effects models. J. Stat. Software 82, 1–26.
Lankowicz, K.M., Bi, H., Liang, D., Fan, C., 2020. Sonar imaging surveys fill data gaps in forage fish populations in shallow estuarine tributaries. Fish. Res. 226, 105520.
Lynch, T.P., Alderman, R., Hobday, A.J., 2015. A high-resolution panorama camera system for monitoring colony-wide seabird nesting behaviour. Methods Ecol. Evol. 6, 491–499.
Ma, R., Kazama, K., Yasutake, Y.C., Kazama, M.T., Tsukamoto, S., Watanuki, Y., 2022. Dynamic body acceleration improves mapping of at‐sea foraging behavior in black‐tailed gulls Larus crassirostris. J. Avian Biol. 2022, e02869.
Masden, E.A., Haydon, D.T., Fox, A.D., Furness, R.W., 2010. Barriers to movement: modelling energetic costs of avoiding marine wind farms amongst breeding seabirds. Mar. Pollut. Bull. 60, 1085–1091.
McInnes, J.C., Jarman, S.N., Lea, M.A., Raymond, B., Deagle, B.E., Phillips, R.A., et al., 2017. DNA metabarcoding as a marine conservation and management tool: a circumpolar examination of fishery discards in the diet of threatened albatrosses. Front. Mar. Sci. 4, 277.
McLeay, L.J., Page, B., Goldsworthy, S.D., Paton, D.C., Teixeira, C., Burch, P., et al., 2010. Foraging behaviour and habitat use of a short-ranging seabird, the crested tern. Mar. Ecol.: Prog. Ser. 411, 271–283.
Michel, L., Cianchetti-Benedetti, M., Catoni, C., Dell'Omo, G., 2022. How shearwaters prey. New insights in foraging behaviour and marine foraging associations using bird-borne video cameras. Mar. Biol. 169, 7.
Montevecchi, W.A., Myers, A., 1996. Dietary changes of seabirds indicate shifts in pelagic food webs. Sarsia 80, 313–322.
Montevecchi, W.A., Benvenuti, S., Garthe, S., Davoren, G.K., Fifield, D., 2009. Flexible foraging tactics by a large opportunistic seabird preying on forage-and large pelagic fishes. Mar. Ecol.: Prog. Ser. 385, 295–306.
Mullers, R.H., Navarro, R.A., Daan, S., Tinbergen, J.M., Meijer, H.A., 2009. Energetic costs of foraging in breeding Cape gannets Morus capensis. Mar. Ecol.: Prog. Ser. 393, 161–171.
Parsons, M., Mitchell, I., Butler, A., Ratcliffe, N., Frederiksen, M., Foster, S., et al., 2008. Seabirds as indicators of the marine environment. ICES J. Mar. Sci. 65, 1520–1526.
Perrow, M., Harwood, A., Berridge, R., Skeate, E., 2017. The foraging ecology of Sandwich Terns in north Norfolk. Br. Birds 110, 249–308.
Peterson, S.H., Ackerman, J.T., Eagles-Smith, C.A., Herzog, M.P., Hartman, C.A., 2018. Prey fish returned to Forster's tern colonies suggest spatial and temporal differences in fish composition and availability. PLoS One 13, e0193430. .
Poupart, T.A., Waugh, S.M., Kato, A., Arnould, J.P., 2020. Foraging niche overlap during chick-rearing in the sexually dimorphic Westland petrel. R. Soc. Open Sci. 7, 191511.
R Core Team, 2022. R: A Language and Environment for Statistical Computing. R
Foundation for Statistical Computing, Vienna, Austria. .
Reif, V., Tornberg, R., 2006. Using time-lapse digital video recording for a nesting study of birds of prey. Eur. J. Wildl. Res. 52, 251–258.
Rodway, M.S., Montevecchi, W.A., 1996. Sampling methods for assessing the diets of Atlantic puffin chicks. Mar. Ecol.: Prog. Ser. 144, 41–55.
Rolland, V., Nepshinsky, M., Windhoffer, E.D., Liechty, J.S., Minor, A.K., Pierce, A.R., 2020. Identification of foraging areas and movements of royal tern Thalasseus maximus at the isles dernieres barrier islands refuge, Louisiana. Mar. Ornithol. 48, 163–168.
Sakamoto, K.Q., Takahashi, A., Iwata, T., Trathan, P.N., 2009. From the eye of the albatrosses: a bird-borne camera shows an association between albatrosses and a killer whale in the Southern Ocean. PLoS One 4, e7322.
Steen, R., 2009. A portable digital video surveillance system to monitor prey deliveries at raptor nests. J. Raptor Res. 43, 69–75.
Stienen, E.W.M., 2006. Living with Gulls. Trading off Food and Predation in the
Sandwich Tern Sterna sandvicensis. Ph.D Thesis. Rijksuniversiteit Groningen.
Stienen, E.W.M., van Beers, P.W., Brenninkmeijer, A., Habraken, J.M.P.M., Raaijmakers, M.H.J.E., van Tienen, P.G., 2000. Reflections of a specialist: patterns in food provisioning and foraging conditions in Sandwich Terns Sterna sandvicensis. Ardea 88, 33–49.
Stienen, E.W.M., Brenninkmeijer, A., Geschiere, C.E., 2001. Living with gulls: the consequences for Sandwich terns of breeding in association with black-headed gulls. Waterbirds 24, 68–82.
Stienen, E.W.M., Brenninkmeijer, A., 2002. Foraging decisions of Sandwich Terns in the presence of kleptoparasitising gulls. Auk 119, 473–486.
Stienen, E.W.M., Brenninkmeijer, A., Courtens, W., 2015. Intra-specific plasticity in parental investment in a long-lived single-prey loader. J. Ornithol. 156, 699–710.
Sugishita, J., Torres, L.G., Seddon, P.J., 2015. A new approach to study of seabird-fishery overlap: connecting chick feeding with parental foraging and overlap with fishing vessels. Glob. Ecol. Conserv. 4, 632–644.
Thaxter, C.B., Lascelles, B., Sugar, K., Cook, A.S.C.P., Roos, S., Bolton, M., et al., 2012. Seabird foraging ranges as a preliminary tool for identifying candidate Marine Protected Areas. Biol. Conserv. 156, 53–61.
Townsend, D.W., 1992. Ecology of larval herring in relation to the oceanography of the Gulf of Maine. J. Plankton Res. 14, 467–493.
van Donk, S., Shamoun-Baranes, J., van der Meer, J., Camphuysen, K.C., 2019. Foraging for high caloric anthropogenic prey is energetically costly. Mov. Ecol 7, 17.
van Donk, S., Shamoun‐Baranes, J., Bouten, W., van der Meer, J., Camphuysen, K.C., 2020. Individual differences in foraging site fidelity are not related to time‐activity budgets in Herring Gulls. Ibis 162, 429–445.
van Franeker, J.A., 2001. Mirrors in Ice. Fulmarine Petrels and Antartic Ecosystems. Ph.D
Thesis. University of Groningen, Groningen, Netherlands.
Votier, S.C., Bicknell, A., Cox, S.L., Scales, K.L., Patrick, S.C., 2013. A bird's eye view of discard reforms: bird-borne cameras reveal seabird/fishery interactions. PLoS One 8, e57376.
Wakefield, E.D., Phillips, R.A., Matthiopoulos, J., 2009. Quantifying habitat use and preferences of pelagic seabirds using individual movement data: a review. Mar. Ecol. Prog. Ser. 391, 165–182.
Williams, C.T., Buck, C.L., 2010. Using fatty acids as dietary tracers in seabird trophic ecology: theory, application and limitations. J. Ornithol. 151, 531–543.
Weimerskirch, H., Ancel, A., Caloin, M., Zahariev, A., Spagiari, J., Kersten, M., et al., 2003. Foraging efficiency and adjustment of energy expenditure in a pelagic seabird provisioning its chick. J. Anim. Ecol. 72, 500–508.
Welcker, J., Moe, B., Bech, C., Fyhn, M., Schultner, J., Speakman, J.R., et al., 2010. Evidence for an intrinsic energetic ceiling in free‐ranging kittiwakes Rissa tridactyla. J. Anim. Ecol. 79, 205–213.
Table
1.
The two GLM models applied to the data of Wood Pigeon, Red-legged Partridge, and Eurasian Magpie using the variables: habitat (forest/non-forest), year (2021/2020), and SPI.
Wood Pigeon
Red-legged Partridge
Eurasian Magpie
Estimate
S.E.
P
Estimate
S.E.
P
Estimate
S.E.
P
Intercept
1.934
1.106
0.080
−2.808
2.007
0.162
0.533
0.891
0.550
Habitat
−2.252
0.446
< 0.001
−2.101
0.742
0.005
−0.605
0.348
0.082
Year
−0.336
0.344
0.328
−0.060
0.554
0.913
−0.477
0.277
0.085
SPI
−0.948
1.255
0.450
2.592
2.289
0.257
0.963
1.010
0.341
Estimate
S.E.
P
Estimate
S.E.
P
Estimate
S.E.
P
Intercept
1.053
0.209
< 0.001
−0.604
0.059
0.059
1.148
0.173
< 0.001
Habitat
−2.284
0.387
< 0.001
−2.455
0.620
< 0.001
−0.759
0.304
0.0124
Note: the columns correspond to: coefficients (Estimate), standard error (SE), and probability (P).The coeffcient estimates with significant differences (P < 0.05) are in bold type.

 DownLoad:
DownLoad:







 Email Alerts
Email Alerts RSS Feeds
RSS Feeds