Kai Gao, Xuemei Yang, Xi Huang, Wenhong Deng. 2024: Integrating host biological and ecological variables to predict probability of haemosporidian infection in raptors. Avian Research, 15(1): 100185. DOI: 10.1016/j.avrs.2024.100185
Citation:
Kai Gao, Xuemei Yang, Xi Huang, Wenhong Deng. 2024: Integrating host biological and ecological variables to predict probability of haemosporidian infection in raptors. Avian Research, 15(1): 100185. DOI: 10.1016/j.avrs.2024.100185
Kai Gao, Xuemei Yang, Xi Huang, Wenhong Deng. 2024: Integrating host biological and ecological variables to predict probability of haemosporidian infection in raptors. Avian Research, 15(1): 100185. DOI: 10.1016/j.avrs.2024.100185
Citation:
Kai Gao, Xuemei Yang, Xi Huang, Wenhong Deng. 2024: Integrating host biological and ecological variables to predict probability of haemosporidian infection in raptors. Avian Research, 15(1): 100185. DOI: 10.1016/j.avrs.2024.100185
Ministry of Education Key Laboratory for Biodiversity Science and Ecological Engineering, College of Life Sciences, Beijing Normal University, Beijing, 100875, China
Funds:
the National Natural Science Foundation of China210100191
Variations in host traits that influence their exposure and susceptibility may impact probability of vector-transmitted diseases. Therefore, identifying the predictors of infection probability is necessary to understand the risk of disease outbreaks during expanding environmental perturbation. Here, we conducted a large survey based on microscopic examination and molecular analysis of haemosporidian parasite infection in raptors rescued at the Beijing Raptor Rescue Centre. Combining these data with biological and ecological variables of the raptors, we determined predictors that affect the probability of haemosporidian infection using generalized linear mixed models and multimodel inference. Our results showed that infection probability exhibited considerable variation across host species in raptors, and body mass, sex, and evolutionary history played relatively weaker roles in driving infection probability. Instead, activity pattern, age, geographic range size, migration distance, and nest type were important predictors of the probability of haemosporidian infection, and the role of each predictor differed in the three main haemosporidian genera (Plasmodium, Haemoproteus, and Leucocytozoon). This macro-ecological analysis will add to our understanding of host traits that influence the probability of avian haemosporidian infection and will help inform risk of emerging diseases.
Parasitic infections can exert harmful effects on their hosts by modifying host survival or reproductive success and can even cause rapid population decline (Merino et al., 2000; Marzal et al., 2005; Knowles et al., 2010a; Asghar et al., 2011, 2015). Parasites may encounter different hosts during their lifetime, once these parasites meet new compatible hosts, they can cause emerging infectious diseases, which can be detrimental to host populations due to lack of preacquired resistance (Jones et al., 2008; Poulin, 2011; McClure et al., 2020). To predict emerging infectious diseases, identifying factors that cause variations in the probability of parasite infection among host species is essential to understand whether a parasite can survive in certain hosts or environments (Brooks et al., 2014; Johnson et al., 2015). Unfortunately, predicting risk of transmission is difficult due to an inadequate understanding of how biological and ecological variables of hosts affect the probability of parasite infection in wildlife.
Avian haemosporidian parasites (order Haemosporida; genera Plasmodium, Haemoproteus, and Leucocytozoon) are protists that infect bird hosts. These parasites are transmitted by blood-feeding insect vectors, are nearly globally distributed, and found in most bird families (Valkiūnas, 2005; Fecchio et al., 2021). Hence, they have gradually developed into an important model system in the fields of host-parasite interaction and evolution of wildlife disease (Bensch et al., 2000; Hellgren et al., 2009; Huang et al., 2018; Rivero and Gandon, 2018; Fecchio et al., 2020a). In the wild, the probability of avian haemosporidian infection is influenced by a combination of biological, ecological, and environmental factors, resulting in a complex infection pattern (Pérez-Rodríguez et al., 2013; Ellis et al., 2015; Clark et al., 2016; McNew et al., 2021). Investigating predictors of the probability of avian haemosporidian infection is crucial for gaining insights into how host traits shape the dynamics of parasite infection.
Variations in probability of haemosporidian infection across avian species may be explained by intra- and interspecific host traits that influence host exposure and susceptibility. At the intraspecific level, probability of haemosporidian infection was associated with several biological variables of the host, such as age, sex, and body mass. For instance, the adult Blue Tit (Cyanistes caeruleus) has a higher probability of Plasmodium and Haemoproteus infection compared than juveniles, because they may encounter competent vectors more frequently during their lifetime (Wood et al., 2007; Lachish et al., 2011; Podmokła et al., 2014). Similarly, the older Great Tit (Parus major) had a higher probability of Plasmodium infection (Isaksson et al., 2013). By contrast, hatch-year birds had higher probability of Plasmodium infection, for example, the Northern Saw-whet Owl (Aegolius acadicus) (Carlson et al., 2018) and younger Seychelles Warbler (Acrocephalus sechellensis) displayed higher probability of Haemoproteus infection (Hammers et al., 2016). Besides, although trading of reproduction for immunity in female birds may led a higher probability of infection (McCurdy et al., 1998), in the Blue Tit, male birds often had a higher probability of Plasmodium and Haemoproteus infection due to the immunosuppressive effects of testosterone (Wood et al., 2007; Lachish et al., 2011). By contrast, another study found no sex difference in this species (Podmokła et al., 2014). Larger body mass was associated with higher susceptibility to haemosporidian parasites possibly due to increased carbon dioxide emission and vector attraction (Gibson and Torr, 1999; Lehane, 2005). For example, in House Sparrow (Passer domesticus), higher body mass increased the probability of haemosporidian infection (Coon and Martin, 2014). However, Schultz et al. (2010) found weak support for body mass as a predictor in the Cape Weaver (Ploceus capensis). These discrepancies may result from variations in season, species, or detection method, warranting further investigation and comparison.
At the interspecific level, haemosporidian parasites tend to infect closely related species due to similarities in their immune systems (Barrow et al., 2019; Gupta et al., 2020). The ecological variables of several hosts are associated with the probability of haemosporidian infection. For example, migration increased probability of haemosporidian infection in birds, which would be exposed to pathogens when moving across a range of habitats (Koprivnikar and Leung, 2015; de Angeli Dutra et al., 2021). However, some studies found migration distance did not predict probability of haemosporidian infection (Sorensen et al., 2019). Nest type affected probability of haemosporidian infection by increasing or decreasing the frequency of contact with haemosporidian vectors, species with open nests often had a high probability of Haemoproteus and Plasmodium infection (Fecchio et al., 2011, 2022; Lutz et al., 2015; Ganser et al., 2020; Rodriguez et al., 2021; Rodríguez-Hernández et al., 2021). Nest site is an important determinant of the probability of haemosporidian infection, because vectors for Plasmodium and Haemoproteus vary based on ground vs. canopy nesting (Černý et al., 2011). For example, a higher probability of Haemoproteus infection would be expected in nests near canopies where the biting midge is more abundant (González et al., 2014; Matthews et al., 2016). In addition to these host biological and ecological variables, other predictors of the probability of haemosporidian infection at the host species level remain to be addressed, such as circadian rhythm, geographic range size, and habitat specialization. For example, although geographic range size correlated positively with the richness of helminths in birds (Gutiérrez et al., 2019), a few studies have examined the relationship between geographic range size of the host and probability of haemosporidian infection (except for Tella et al., 1999). Thus, it is necessary to concurrently measure multiple biological and ecological variables to identify which host traits serve as robust predictors of the probability of haemosporidian infection across bird hosts (Fecchio et al., 2022).
Considering that haemosporidian parasites from different genera use different arthropod vectors and show unique life-history characteristics (Valkiūnas, 2005; Santiago-Alarcon et al., 2012), comparative studies combining parasites from all three genera are needed to understand the influence and determinants of these parasites in wild species (Olsson-Pons et al., 2015; Williams et al., 2022). For example, two recent studies have suggested that the determinants were different across haemosporidian genera. For Plasmodium, a high minimum temperature was associated with increased probability of haemosporidian infection. By contrast, high minimum temperature was an important environmental barrier for Leucocytozoon distribution (Fecchio et al., 2020b; Clark et al., 2020). These studies highlight the importance of understanding how host biological and ecological variables impact the probability of haemosporidian infection from different genera amidst the backdrop of escalating climate change.
In this study, we present the first integrative analysis of predictors of probability of haemosporidian infection in raptors. For this study, we first used microscopic examination and molecular survey to detect haemosporidian parasites in raptors. We subsequently conducted a comprehensive analysis with the following objectives: (a) to analyze predictors influencing the probability of haemosporidian infection in raptor species (see Table 1 for hypotheses and predictions); (b) to assess the difference in the importance among predictors of infection probability; and (c) to conduct separate analysis for each of the three distinct haemosporidian parasite genera, considering their varying activity pattern and levels of pathogenicity.
Table
1.
Hypotheses and predictions linking host traits to probability of haemosporidian infection in raptors.
Predictors
Category
General hypotheses and predictions
Activity pattern
Biological variable
Unknown
Age
Biological variable
Adult raptors are expected to have a higher probability of haemosporidian infection than juveniles, because they might encounter more parasite species during their lifetimes (Wood et al., 2007).
Body mass
Biological variable
Body mass would either be positively associated with the probability of haemosporidian infection due to increased vector attraction (Gibson and Torr, 1999; Lehane, 2005), or negatively associated with the probability of haemosporidian infection as larger birds displayed better immune responses to constrain parasite invasion (Ruhs et al., 2020).
Dietary diversity
Biological variable
Raptors with a broader diet are expected to have a higher probability of haemosporidian infection than more specialized ones, because they might encounter more parasite species during their lifetimes (Leung and Koprivnikar, 2016).
Geographic range
Ecological variable
Raptors with larger geographic ranges are expected to have a higher probability of haemosporidian infection. Because larger geographic ranges offer more opportunities for host–parasite associations to become established and encompass a greater diversity of habitats suitable for different parasites (Tella et al., 1999).
Habitat specialization
Ecological variable
Raptors with broader habitat breadths are expected to have a higher probability of haemosporidian infection than more specialized ones, because they might encounter more parasite species during their lifetimes (Leung and Koprivnikar, 2016).
Migration distances
Ecological variable
Raptors with longer migration distances are expected to have a higher probability of haemosporidian infection, because migration may bring hosts in contact with "new" parasites (Koprivnikar and Leung, 2015; de Angeli Dutra et al., 2021).
Nest site
Ecological variable
A higher probability of haemosporidian infection would be expected in raptors that nest near canopies, where the vectors are more abundant (Černý et al., 2011).
Nest type
Ecological variable
Raptors with nests are expected to have a higher probability of haemosporidian infection, because nests increase the frequency of contact with haemosporidian vectors (Ganser et al., 2020; Fecchio et al., 2022).
Sex
Biological variable
Female raptors trading reproduction for immunity would be expected to have a higher probability of haemosporidian infection than males (McCurdy et al., 1998).
From 2007 to 2021, we collected blood samples from 1539 raptors belonging to 35 species rescued at the Beijing Raptor Rescue Centre (BRRC), Beijing, China, which is an important node in the East Asian–Australasian flyway of migratory birds (Xie et al., 2016). The BRRC originates from a conservation project of the International Fund for Animal Welfare, which provides care, medical treatment, and rehabilitation training to debilitated or injured raptors. The majority of raptors received by the BRRC were found in Beijing and adjacent areas. The raptors mainly included injured or debilitated raptors found by citizens, nestlings that fell out of nests, raptors that accidently entered human settlements, and raptors that were confiscated by enforcement actions.
2.2
Parasite detection
Blood samples were collected from the basilic veins of raptors the day they arrived, following Scott (2016). This allowed us to detect the presence of haemosporidian parasites under natural conditions. According to a study, both microscopic and molecular detection slightly underestimated overall probability of haemosporidian infection (Valkiūnas et al., 2008); therefore, we detected the status of haemosporidian parasite infection by combining microscopy and nested mitochondrial cytochrome b PCR. An individual was considered infected when the microscopy or PCR result was positive.
2.2.1
Microscopic examination
For each raptor individual, two or three blood films were prepared using freshly drawn blood right after collection. The smears were air-dried in 5–15 s, fixed in absolute methanol for 1 min, and stained with Giemsa at 18–20 ℃ for 1 h. Blood films from each raptor were examined using an Axio Scope A1 light microscope equipped with an Axiocam 506 color digital camera. A skilled veterinarian examined the blood films at both low (× 400) and high (× 1000) magnification for approximately 100 fields per slide (Valkiūnas et al., 2008). To determine the genera, the haemosporidian parasites were morphologically identified according to the literature (Valkiūnas, 2005).
2.2.2
Molecular detection
Blood samples were collected and stored in absolute ethanol at −40 ℃ until DNA extraction, which was accomplished using TIANamp Genomic DNA Kit DP304-03 (Tiangen Biotech Ltd., Beijing, China), according to the manufacturer's instructions. Haemosporidian parasites were identified using a nested PCR protocol that amplifies a 479-bp fragment of cyt b (Hellgren et al., 2004). HaemNFI and HaemNR3 primers were used to amplify Plasmodium, Haemoproteus, and Leucocytozoon DNA. Then, the product of the first PCR was used as a template for the second PCR using the primers HaemF and HaemR2 to amplify Haemoproteus and Plasmodium DNA and HaemFL and HaemR2L to amplify Leucocytozoon DNA. At least one negative control and one positive control were included in each round of reaction. The positive controls are usually selected from samples with a high intensity of infection. The negative control uses ddH2O instead of DNA. Each sample was tested three times to verify parasite identity and check for possible mixed infections. Positive samples were sequenced using the primers HaemF, and HaemR2, HaemFL and HaemR2L, after sequencing, the sequences were edited and spliced using the "Assemble" function in SeqMan software. Then unique haplotypes were assigned to identify the lineage of haemosporidian parasites using the MalAvi database (Bensch et al., 2009).
2.3
Host phylogeny and biological variables
2.3.1
Phylogenetic signal
Values for the probability of haemosporidian infection may be more similar among closely related raptor species than distantly related ones, and such a phylogenetic signal would create nonindependence among species in comparative analysis. To calculate the phylogenetic signal among the probability of haemosporidian infection estimated for raptor species in our dataset, we downloaded 1000 trees from http://birdtree.org (Jetz et al., 2012) and combined them into a maximum clade credibility tree using TreeAnnotator in the BEAST software package version 1.8.4 (Drummond et al., 2012). Then, we calculated the phylogenetic signal of the probability of haemosporidian infection haemosporidian based on Pagel's λ (Pagel, 1999) using the phytools R package (Revell, 2012). When Pagel's λ is close zero, phylogeny has negligible effect on the trait in question, whereas values close to 1 indicate strong phylogenetic signal.
2.3.2
Body mass, age, and longevity
Once the raptors were received, we used a digital scale to measure their body mass (up to 0.1 g). We included a binary variable for host age (hatch-year, HY, after-hatch year, AHY). Raptors were aged primarily by distinguishing characteristics of their feathers, which was determined according to the length of primaries and secondaries and presence of juvenile feathers. In general, primaries and secondaries are often longer in juveniles than adults, and juvenile feathers are structurally different from adult feathers (Scott, 2016). We used data on raptor maximum longevity (years) as reported in Healy et al. (2014), which excludes species with maximum longevity estimates based on fewer than 10 longevity records and provides a much-improved body longevity dataset for birds.
2.3.3
Sex determination
To determine the sex of the raptors, we performed molecular sexing using two primer sets, 2550F/2718R and P2/NP/MP, according to Yang et al. (2022). Briefly, 2550F/2718R was used for most species, and P2/NP/MP was used to identify the sex of the Eurasian Eagle-owl (Bubo bubo), Oriental Scops Owl (Otus sunia), and Japanese Scops Owl (Otus semitorques). We followed the procedures described by Fridolfsson and Ellegren (1999) and Lee et al. (2008) to amplify the CHD1W and CHD1Z genes. The PCRs were conducted using the following conditions: (1) for 2550R/2718F, initial denaturation at 94 ℃ for 5 min; 35 cycles of 94 ℃ for 30 s, 49 ℃ for 45 s, and 72 ℃ for 45 s; and final extension at 72 ℃ for 7 min; (2) for P2/NP/MP, initial denaturation at 94 ℃ for 4 min; 35 cycles of 94 ℃ for 30 s, 50 or 51 ℃ for 30 s, and 72 ℃ for 40 s; and final extension at 72 ℃ for 4 min.
2.3.4
Activity pattern
Depending on whether the main active period was in the day or night, raptors were categorized as nocturnal or diurnal species, respectively. For raptors active to some extent at both day and night, we defined their activity pattern based on foraging time, which is when they are most likely to be exposed to vectors (Gao et al., 2021).
2.4
Host biological and ecological variables
2.4.1
Nest type and nest site
We collected data on raptor nest type and nest site using information from "A dataset on the life history and ecological traits of Chinese birds" (Wang et al., 2021). We classified raptor nest type into two categories: (1) open—when the raptor species builds an open-cup nest; and (2) closed—when the raptor species builds a closed-cup nest, nest in tree hole, or cavity. We additionally sorted nest sites into three categories: (1) ground—when a raptor species builds its nest on the ground; (2) canopy—when a raptor species builds its nest on the canopy; and (3) cliff—when a raptor species builds its nest on a cliff.
2.4.2
Geographic range size and migratory distance
To extract the geographic range size of raptors, we first obtained spatial data. We used species-level geographic range size maps from BirdLife International (http://www.birdlife.org/datazone). This online database provides global distributional data in polygon format for all extant bird species. Once the data were downloaded, we converted geographic range size into a presence–absence matrix (1°-grid cell) and then summed the number of cells (area in km2) in which each species occurred using the package "letsR" (Vilela and Villalobos, 2015). Here, we focused on the correlation between probability of haemosporidian infection in raptors and the size of the raptors' entire range; therefore, we included range polygons regardless of seasonality ("resident", "breeding season", "non-breeding season", and "passage"; Fig. 1).
Figure
1.
The migration distances of the raptors in this study. The red dots represent the centroids of the species breeding and nonbreeding ranges, and the asterisks indicate that the species are residents. (For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)
To estimate the migration distances of the raptors, we extracted distribution polygons from species-level geographic range size maps, and then the migration distances (km) were measured as the distance between their breeding and non-breeding ("Seasonality" codes 2 and 3) range centroids using ArcGIS v10.1 (Gutiérrez et al., 2017, 2019).
2.4.3
Dietary diversity and habitat specialization
We extracted dietary information from the EltonTraits database (Wilman et al., 2014). This database includes semiquantitative dietary assessments for all extant bird species. Dietary breadth was defined by assigning values of 1–10 based on the number of components of the diet assigned to each species (invertebrates, endotherms, ectotherms, fish, vertebrates, scavengers, fruit, nectar, seed, and plant). Then, the dietary diversity of raptors was computed as follows: BA=(B-1)/(n-1), where n is the number of possible foods of each food category (n = 10), and B=1/∑p2j, where pj is calculated using ranked proportions (Olalla-Tárraga et al., 2016; Gutiérrez et al., 2019). For example, the Common Kestrel (Falco tinnunculus) can eat two types of food, and the ranked proportions of these two foods are 0.1 and 0.9; therefore, its food diversity is 0.02 (Appendix Table S1).
For habitat specialization of raptors, we used habitat data from the IUCN Red List (https://www.iucnredlist.org). From this data source, we extracted information on 15 breeding habitats used by raptors: deciduous forest, coniferous forest, woodland, shrub, savannah, tundra, grassland, mountain meadow, reed, swamps, semidesert, freshwater, marine, rocks, and human settlements. For each habitat, we recognized whether it was occupied by a given species and calculated habitat breadth as the number of occupied habitat classes (range = 1–10). We further calculated the raptor habitat specialization index as a coefficient of variation of the occurrence of a given species across the 15 habitats (Julliard et al., 2006). The habitat specialization index for a species present in h habitat classes among H possible habitat classes was (h/H−1)1/2. Among our species, the habitat specialization index ranged from 0.71 (equivalent to present in 10/15 habitat classes) for the Peregrine Falcon (Falco peregrinus) to 3.74 (equivalent to present in 1/15 habitat classes) for Pallas's Fish Eagle (Haliaeetus leucoryphus), which is a habitat specialist.
2.5
Statistical analyses
The sample size of each raptor species ranged from 1 to 570. Here, we only included species with ≥10 individuals sampled because a minimum sample size of 10 individuals per bird species gave largely similar infection probability results, and thus, we had high confidence in detecting haemosporidian parasites when actual infection probability was low (Ellis et al., 2020). To determine whether analyses that included all potential predictors of the probability of haemosporidian infection would be collinear, we used Pearson's correlation analysis for normally distributed variables or Spearman's correlation for nonnormally distributed variables to test the relevance between predictor variables. We retained variables with correlation coefficients < 0.5 in subsequent analyses, and this filtering resulted in identifying 10 important potential predictors. Host biological variables included age, sex, body mass, and activity pattern; host ecological variables included nest type, nest site, dietary diversity, geographic range size, habitat specialization, and migration distance. Age and sex were missing for 282 individuals and one raptor species, and we excluded them from further analysis, thus, a total of 35 species and 1257 individuals were retained in our final dataset.
We used generalized linear mixed-effects models (GLMMs) with a logit link and binomial-error distribution to identify the effect of all potential predictors on total probability of haemosporidian infection. The sampling months were included as random effects in all models. We detected negligible phylogenetic signals among raptor species for probability of haemosporidian infection (λ = 0.09), justifying the exclusion of raptor phylogeny into our models. To determine the effects of all potential predictors on raptor haemosporidian infection from different genera, three additional models were fit separately for Haemoproteus, Plasmodium, and Leucocytozoon. If the lineages in mixed infections belonged to two genera, they were included in both genus-specific analyses. We report posterior modes and 95% confidence intervals (CIs).
We employed multimodel inference and model averaging to identify the relative importance of each variable in affecting the probability of haemosporidian infection (Burnham and Anderson, 2002). The global model set had all possible combinations of the 10 predictor variables (total: 210 = 1024 models), and these models were ranked. We estimated the relative importance of each explanatory variable by calculating AIC weights across all of candidate models that included the variable under consideration. We listed the top 10 models (with the lowest AICc values) and retained models that were within 2 AIC units (ΔAIC ≤2) of the top models to be highly supported. All statistical analyses were performed in R version v4.1.1 and required the libraries lme4 and MuMIn (R Core Team, 2021).
3.
Results
In total, 443/1539 raptors sampled from 35 species (Appendix Table S1) were infected with haemosporidian parasites, with a total infection probability of 28.78%. These included 88 Plasmodium (5.72%), 245 Haemoproteus (15.92%), and 158 Leucocytozoon (10.27%) infections. A total of 177 and 387 raptors were identified by microscopic examination and PCR, respectively. Both methods slightly underestimated the overall probability of infection, which confirms the necessity to use both methods for detecting haemosporidian parasites. Of the 19 species with ≥10 individuals sampled, probability of haemosporidian infection was 0–88.89%, and probability of haemosporidian infection varied significantly across host species (χ2 = 307.01, df = 18, p < 0.001).
3.1
Predictors of the probability of haemosporidian infection
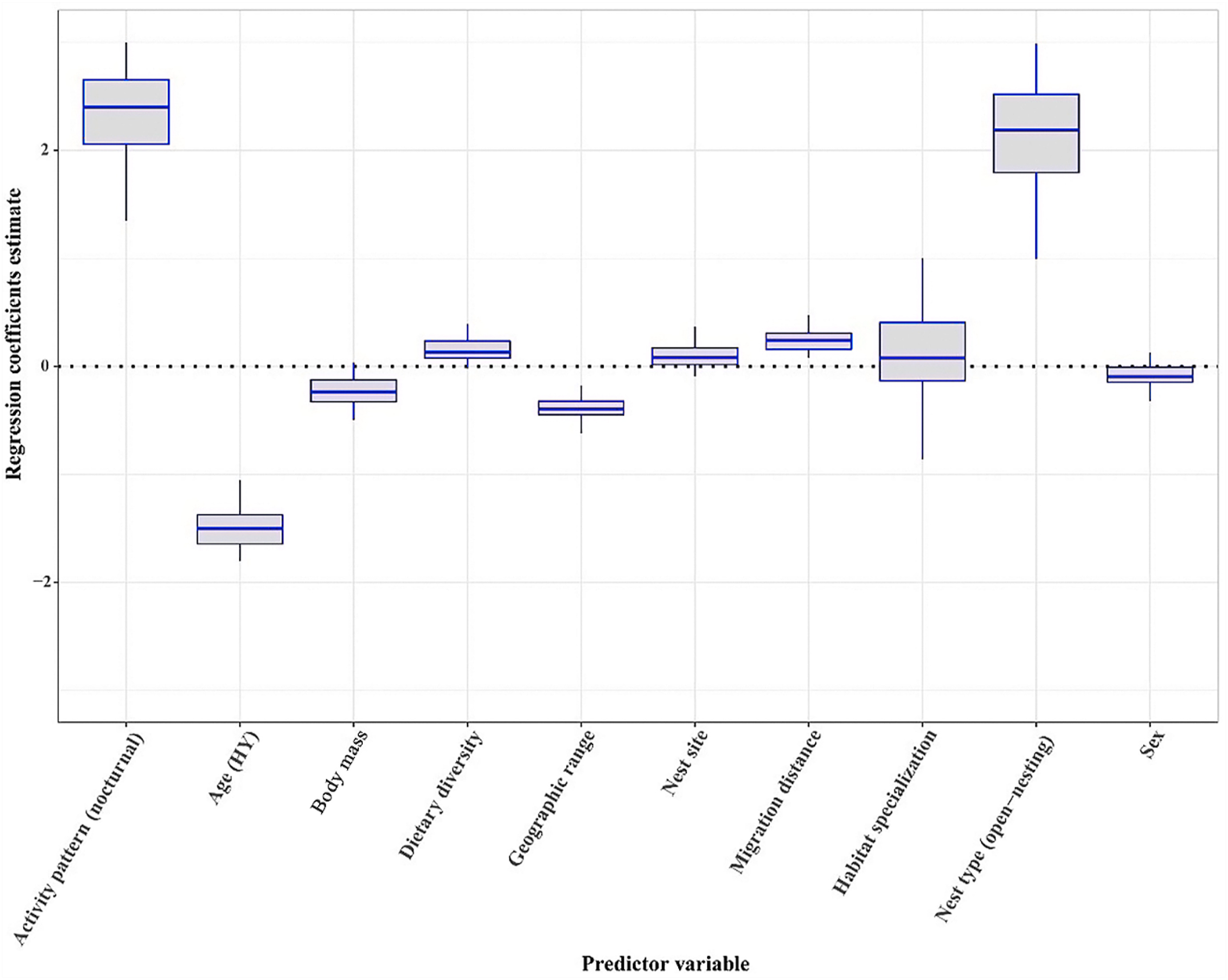
According to GLMM analysis, the best fit model included age, body mass, nest type, dietary diversity, geographic range size, migration distance, and activity pattern. Age, nest type, geographic range size, migration distance and activity pattern were statistically significant for total probability of haemosporidian infection, and the 95% CIs for these predictors only excluded zero (Fig. 2). Among these predictors, older raptors (95% CI: −1.84, −1.09), nocturnal raptors (95% CI: 1.38, 3.31), and raptors that used open nests (95% CI: 0.91, 3.19) had higher total probability of haemosporidian infection. Migration distance positively correlated with total probability of haemosporidian infection (95% CI: 0.04, 0.42), and interestingly, geographic range size negatively correlated with total probability of haemosporidian infection (95% CI: −0.57, −0.13).
Figure
2.
Estimates of the regression coefficients for the biological and ecological variables of haemosporidian parasites infection. Boxplots represent posterior modes (lines), 95% highest posterior density credible intervals (hinges) and minimums and maximums (whiskers).
Of our tested models, the top 10 included combinations of all predictor variables. However, the top five models were the most supported (i.e., ΔAIC ≤2) and contained five variables: age, nest type, geographic range size, migration distance, and activity pattern. In these five models, both fixed factors (R2 marginal) and random effects (R2 conditional) explained > 26% of the variation in probability of haemosporidian infection. There was moderate model selection fit across the multimodel set (Wi = 0.14, 0.14, 0.08, 0.06, and 0.06 for the five top models; Table 2). Age (relative importance value = 0.92), nest type (0.92), geographic range size (0.92), activity pattern (0.92), and migration distance (0.82) were the most important predictors of the probability of haemosporidian infection (Table 3).
Table
2.
The top ten generalized linear mixed-effects models (GLMMs) for the effect of biological and ecological variables on haemosporidian parasites infection of raptor species.
1
2
3
4
5
6
7
8
9
10
Activity pattern
*
*
*
*
*
*
*
*
*
*
Age
*
*
*
*
*
*
*
*
*
*
Body mass
*
*
*
*
*
*
*
*
*
*
Dietary diversity
*
*
*
*
*
*
Geographic range
*
*
*
*
*
*
*
*
*
*
Habitat specialization
*
*
*
Migration distances
*
*
*
*
*
*
*
*
*
Nest site
*
*
Nest type
*
*
*
*
*
*
*
*
*
*
Sex
*
*
*
K
7
6
8
8
7
7
6
9
7
8
AICc
1271.05
1271.14
1272.34
1272.81
1272.96
1273.07
1273.57
1274.02
1274.09
1274.4
ΔAIC
0
0.09
1.29
1.76
1.91
2.02
2.52
2.97
3.04
3.35
Wi
0.14
0.14
0.08
0.06
0.06
0.05
0.04
0.03
0.03
0.03
R2 marginal
0.27
0.26
0.27
0.27
0.26
0.26
0.27
0.27
0.27
0.27
R2 conditional
0.28
0.27
0.28
0.28
0.28
0.27
0.28
0.28
0.28
0.28
Note: *, indicates that a variable is included in the model. K, number of predictors in the model. ΔAIC, the difference between each model and the highest ranked model. AICc, Akaike's information criterion adjusted for small sample sizes. Wi (Akaike weights), the probability that a model is best given the particular set of models considered. R2 marginal, amount of variation that is explained by fixed factors (covariates). R2 conditional, amount of variation that is explained by both fixed and random factors. Models include sampling date and species as a random effect. Models are ranked in order of decreasing ΔAIC.
Table
3.
Model averaging based on GLMMs (1024 models) using biological and ecological variables, age, nest type, activity pattern, geographic range size and migration distance, to explain haemosporidian parasites infection of raptor species.
β
SE
95% CI (lower, upper)
Relative importance
Age (HY)
−1.46
0.19
−1.84, −1.09
0.92
Nest type (open-nesting)
2.05
0.58
0.91, 3.19
0.92
Activity pattern (nocturnal)
2.34
0.49
1.38, 3.31
0.92
Geographic range
−0.35
0.11
−0.57, −0.13
0.92
Migration distances
0.23
0.10
0.04, 0.42
0.82
Body mass
−0.24
0.13
−0.48, 0.01
0.81
Dietary diversity
0.16
0.10
−0.04, 0.36
0.54
Habitat specialization
0.08
0.11
−0.14, 0.30
0.29
Sex (M)
−0.07
0.14
−0.36, 0.21
0.26
Nest site (cliff)
0.11
0.42
−0.72, 0.95
0.15
Nest site (ground)
0.54
0.67
−0.77, 1.86
0.15
Note: Bold characters indicate the most important variables through GLMMs. β, model-averaged regression coefficients. HY, hatch-year. Open-nesting, raptors build an open-cup nest. Nocturnal, nocturnal raptors. M, male raptors. Cliff, raptors nest on the cliff. Ground, raptors nest on the ground.
3.3
Predictors of the probability of haemosporidian infection for different genera
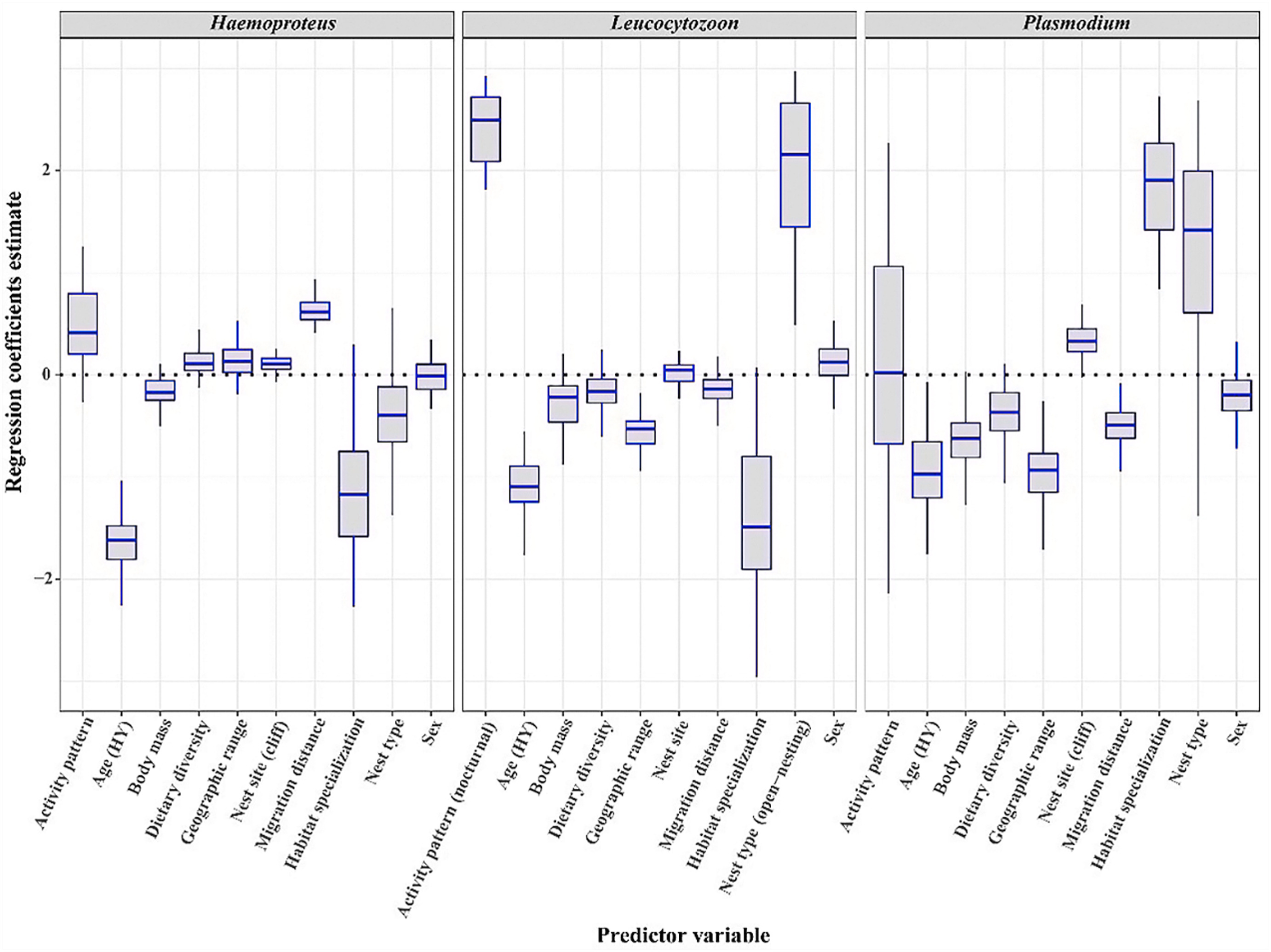
For Plasmodium, geographic range size (relative importance value = 0.89), body mass (0.89), age (0.87), nest site (0.87), migration distance (0.83), and habitat specialization (0.66) were statistically significant for probability of haemosporidian infection. Older raptors (95% CI: −1.7, −0.29) and raptors nesting on cliffs (95% CI: 0.6, 3.27) had a higher probability of Plasmodium infection. Geographic range size (95% CI: −1.52, −0.44), migration distance (95% CI: −0.88, −0.09), and body mass (95% CI: −1.16, −0.1) were negatively correlated with the probability of Plasmodium infection, and habitat specialization was positively correlated with the probability of Plasmodium infection (95% CI: 0.01, 0.62) (Fig. 3).
Figure
3.
Estimates of the regression coefficients for the predictors of infection probability for Haemoproteus, Leucocytozoon and Plasmodium. Boxplots represent posterior modes (lines), 95% highest posterior density credible intervals (hinges) and minimums and maximums (whiskers).
For Haemoproteus, age (relative importance value = 0.78) and migration distance (0.78) were statistically significant for the probability of haemosporidian infection. Raptors with older age (95% CI: −2.05, −1.09) and longer migratory distances (95% CI: 0.38, 0.83) had a higher probability of Haemoproteus infection (Fig. 3).
For Leucocytozoon, age (relative importance value = 0.91), nest type (0.91), activity pattern (0.91), and geographic range size (0.9) were statistically significant for the probability of haemosporidian infection. Raptors with older age (95% CI: −1.68, −0.5) and smaller geographic range size (95% CI: −0.89, −0.17) and raptors that used open nests (95% CI: 1.05, 3.75) had a higher probability of Leucocytozoon infection. The probability of Leucocytozoon infection was significantly higher in nocturnal raptors than diurnal raptors (95% CI: 1.76, 4.48) (Fig. 3).
4.
Discussion
Of the potential predictors of the probability of haemosporidian infection tested, biological variables of raptor species related to host immunity, such as body mass, sex, and evolutionary history, played relatively weak roles in driving total probability of haemosporidian infection. Instead, host ecological variables, such as activity pattern, age, geographic range size, migration distance, and nest type, were the best predictors of total probability of haemosporidian infection in raptor hosts. By integrating data and comparative statistical models, our study improves the understanding of how host biological and ecological variables are important determinants of probability of haemosporidian infection in raptors.
In this study, we examined the relationship between host age and probability of haemosporidian infection in raptors based on a rich dataset comprising 1257 raptors from 35 species and collected both molecular and microscopy data from these samples. Our study provides strong support for the use of host age to predict the probability of haemosporidian infection in raptors. We found that older raptors had a higher probability of haemosporidian infection, and when different parasite genera were tested separately, a similar age-related pattern was detected for Plasmodium, Haemoproteus, and Leucocytozoon. However, studies investigating the effect of age on the infection status of avian haemosporidian parasites found that age did not affect the probability of haemosporidian infection. For example, Matthews et al. (2016) sampled 329 individuals from 43 species in eastern Tennessee and found that infection status did not vary with host age. Popescu et al. (2020) used PCR to screen for infection status in 625 individuals from 35 species in western Tennessee and found that individual-level traits, such as age and sex, did not predict infection status of individuals. These contradictory results may suggest that relationships between host age and probability of haemosporidian infection are different across a broad range of host species and regions.
Regarding nest type, the observed increase in the overall probability of haemosporidian infection in raptors that build open nests is consistent with previous studies from passerines showing that open-nest hosts are more likely to be infected by haemosporidian parasites (Lutz et al., 2015; Matthews et al., 2016; Rodríguez-Hernández et al., 2021). However, when different parasite genera were tested separately, we did not observe a significant difference in the probability of Plasmodium and Haemoproteus infection across species with different nest type; however, the probability of Leucocytozoon infection was significantly higher for raptor species that had open-cup nests. The main insect vectors of Plasmodium and Haemoproteus are active at sunset or night and locate their hosts with olfactory cues (Gibson and Torr, 1999; Lehane, 2005). By contrast, the vectors of Leucocytozoon are diurnal feeders and find their hosts with visual cues (Marquardt et al., 2000; Grillet et al., 2005; Lehane, 2005). These different host-seeking behaviors may account for the higher probability of Leucocytozoon infection in raptor species with open-cup nests.
Our results suggest that longer migratory distances resulted in a higher probability of haemosporidian infection, possibly because migrants moving across a range of habitats would be exposed to many parasites, and long-distance migration may reduce their energy for immune defense (Altizer et al., 2011). Notably, a few studies have provided comparative analysis of the probability of haemosporidian infection in bird hosts with different migratory statuses, such as resident, partially migrant, and fully migrant, and reported the same pattern as our study (Koprivnikar and Leung, 2015; de Angeli Dutra et al., 2021), whereas other studies could not find evidence of such effects (Gutiérrez et al., 2017; Sorensen et al., 2019). One possible interpretation is that species with shorter migration distances invest significant resources in parasite defense (Sorensen et al., 2019). When analyzing Plasmodium, Haemoproteus, and Leucocytozoon separately, we did not observe a significant difference in the probability of Leucocytozoon infection between species with different migratory distances, but we found a positive and negative correlation between the probability of Haemoproteus and Plasmodium infection and migratory distance, respectively. Because Plasmodium is the most and Haemoproteus is the least virulent of all parasites in wild birds, we hypothesize that the migratory behavior of avian hosts reduces their probability of being infected by Plasmodium, but at the cost of higher rate of infection by Haemoproteus (Knowles et al., 2010b; Dadam et al., 2019; McClure et al., 2020).
Parasite richness increased with the size of the occupied geographic range (Kamiya et al., 2014; Gutiérrez et al., 2019). However, few studies have reported the correlation between the probability of haemosporidian infection and size of geographic range, except for Tella et al. (1999), who sampled 20 diurnal raptor species in Spain and found that infection probability was positively correlated with the host's geographic range. In this study, we found that the probability of haemosporidian infection (except for that of Haemoproteus) decreased with geographic range size. This discrepancy may result from different detection and analysis methods. Tella et al. (1999) only collected microscopy data and did not analyze infection probability in different genera separately, whereas we collected both molecular and microscopy data and analyzed infection probability in different genera. Our result is somewhat unexpected, because widespread species are expected to have higher rates of parasitic infection due to a broad distributional range and may expose hosts to a greater range of vectors and parasites. However, a study suggested that widespread species exhibited stronger adaptive immune defenses than geographically restricted species, and these immunological differences could affect the infection probability of avian malarial parasites (Lee et al., 2006). Thus, our findings are consistent with the assumption that widely distributed species are less likely to be infected with parasites due to stronger adaptive immune defense, which may contribute to host colonization over wide geographic areas.
Regarding the activity pattern, we found that nocturnal raptors had a higher probability of haemosporidian infection than diurnal raptor species. Furthermore, we observed a higher probability of Leucocytozoon infection in nocturnal raptors. The vectors of Leucocytozoon are diurnal feeders, and thus, show a unique life-history feature compared with the nocturnal vectors of Plasmodium and Haemoproteus (Marquardt et al., 2000; Grillet et al., 2005; Lehane, 2005). Because most nocturnal raptors are immobile during daytime, the diurnal feeders can bite without encountering a host defense. Thus, nocturnal raptors have a higher probability of Leucocytozoon infection.
We expected body mass would either be positively associated with the probability of haemosporidian infection due to increased vector attraction (Gibson and Torr, 1999; Lehane, 2005), or negatively associated with the probability of haemosporidian infection as larger birds displayed better immune responses to constrain parasite invasion (Ruhs et al., 2020). However, our study only found a negative correlation between body mass and probability of Plasmodium infection, one of the possible explanations is that acute infections with Plasmodium was linked to decreased body mass (Atkinson and Samuel, 2010), thus leading to a negative correlation between the probability of Plasmodium infection and body mass (Ruhs et al., 2020). In addition, we found evidence that the probability of Plasmodium infection was slightly higher in raptors nest on the cliffs than in others (ground and canopy). Because vector diversity and abundance may affect the diversity and distribution of haemosporidian parasites (Santiago-Alarcon et al., 2012), studies on vector abundance in different nest sites will help identify the reasons behind this difference.
Finally, among the remaining variables tested in our analysis, host biological variables related to sex and dietary diversity were significant determinants of the probability of haemosporidian infection (reviewed in Ellis et al., 2020). However, these traits were not identified as robust explanatory variables in our analysis. Notably, our models were designed to identify which host biological and ecological variables serve as robust predictors of raptor haemosporidian infection. Using our integrated dataset, we identified important predictors and eliminated non-statistically significant predictors. In conclusion, we found several correlates of the probability of haemosporidian infection (both for overall probability of infection and for the three genera of haemosporidian parasites). These findings will add to our understanding of host biological and ecological variables that influence the probability of avian haemosporidian infection, and help inform risk of emerging diseases. However, to identify the mechanisms driving these changes, future studies must use ecological models that clarify the distribution and diversity of vectors, along with different birds and geographic regions.
Ethics statement
All animal experiments were carried out in accordance with the guidelines and approval of the Animal Research and Ethics Committee of College of Life Science, Beijing Normal University (permissions, CLS-EAW-2021-021).
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
CRediT authorship contribution statement
Kai Gao: Writing – original draft, Visualization, Methodology, Data curation, Conceptualization. Xuemei Yang: Investigation, Data curation. Xi Huang: Writing – review & editing, Methodology, Conceptualization. Wenhong Deng: Writing – review & editing, Project administration, Funding acquisition, Conceptualization.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We gratefully acknowledge the support of the Beijing Raptor Rescue Centre (BRRC). We thank C. Dai, S. Zhang, L. Zhou, Z.S. Li and Y.G. Shang for their assistance in collecting samples.
Altizer, S., Bartel, R., Han, B.A., 2011. Animal migration and infectious disease risk. Science 331, 296-302.
Asghar, M., Hasselquist, D., Bensch, S., 2011. Are chronic avian haemosporidian infections costly in wild birds? J. Avian Biol. 42, 530-537.
Asghar, M., Hasselquist, D., Hansson, B., Zehtindjiev, P., Westerdahl, H., Bensch, S., 2015. Hidden costs of infection: chronic malaria accelerates telomere degradation and senescence in wild birds. Science 347, 436-438.
Atkinson, C.T., Samuel, M.D., 2010. Avian malaria Plasmodium relictum in native Hawaiian forest birds: epizootiology and demographic impacts on àpapane Himatione sanguinea. J. Avian Biol. 41, 357-366.
Barrow, L.N., McNew, S.M., Mitchell, N., Galen, S.C., Lutz, H.L., Skeen, H., et al., 2019. Deeply conserved susceptibility in a multi-host, multiparasite system. Ecol. Lett. 22, 987-998.
Bensch, S., Hellgren, O., Pérez-Tris, J., 2009. MalAvi: a public database of malaria parasites and related haemosporidians in avian hosts based on mitochondrial cytochrome b lineages. Mol. Ecol. Resour. 9, 1353-1358.
Bensch, S., Stjernman, M., Hasselquist, D., Örjan, Ö., Hannson, B., Westerdahl, H., et al., 2000. Host specificity in avian blood parasites: a study of Plasmodium and Haemoproteus mitochondrial DNA amplified from birds. Proc. R. Soc. A B 267, 1583-1589.
Brooks, D.R., Hoberg, E.P., Boeger, W.A., Gardner, S.L., Galbreath, K.E., Herczeg, D., et al., 2014. Finding them before they find us: informatics, parasites, and environments in accelerating climate change. Comp. Parasitol. 81, 155-164.
Burnham, K.P., Anderson, D.R., 2002. Model Selection and Inference: A Practical
Information-Theoretic Approach, second ed. Springer-Verlag, New York.
Carlson, M.L., Proudfoot, G.A., Gentile, K., Dispoto, J., WecksteinM, J.D., 2018 Haemosporidian prevalence in Northern Saw-whet Owls Aegolius acadicus is predicted by host age and average annual temperature at breeding grounds. J. Avian Biol. 49, e01817.
Černý, O., Votýpka, J., Svobodová M., 2011. Spatial feeding preferences of ornithophilic mosquitoes, blackflies and biting midges. Med. Vet. Entomol. 25, 104-108.
Clark, N., Wells, K., Dimitrov, D., Clegg, S., 2016. Co-infections and environmental conditions drive the distributions of blood parasites in wild birds. J. Anim. Ecol. 85, 1461-1470.
Clark, N.J., Drovetski, S.V., Voelker, G., 2020. Robust geographical determinants of infection prevalence and a contrasting latitudinal diversity gradient for haemosporidian parasites in Western Palearctic birds. Mol. Ecol. 29, 3131-3143.
Coon, C.A.C., Martin, L.B., 2014. Patterns of haemosporidian prevalence along a range expansion in introduced Kenyan House Sparrows Passer domesticus. J. Avian Biol. 45, 34-42.
Dadam, D., Robinson, R.A., Clements, A., Peach, W.J., Bennett, M., Rowcliffe, J.M., et al., 2019. Avian malaria-mediated population decline of a widespread iconic bird species. R. Soc. Open Sci. 6, 182197.
de Angeli Dutra, D., Fecchio, A., Martins Braga, É., Poulin, R., 2021. Migratory birds have higher prevalence and richness of avian haemosporidian parasites than residents. Int. J. Parasitol. 51, 877-882.
Drummond, A.J., Suchard, M.A., Xie, D., Rambaut, A., 2012. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 29, 1969-1973.
Ellis, V.A., Collins, M.D., Medeiros, M.C.I., Sari, E.H.R., Coffey, E.D., Dickerson R.C., et al., 2015. Local host specialization, host-switching, and dispersal shape the regional distributions of avian haemosporidian parasites. Proc. Natl. Acad. Sci. U.S.A. 112, 11294-11299.
Ellis, V.A., Fecchio, A., Ricklefs, R.E., 2020. Haemosporidian parasites of Neotropical birds: causes and consequences of infection. Auk 137, 365-378.
Fecchio, A., Bell, J.A., Bosholn, M., Vaughan, J.A., Tkach, V.V., Lutz, H.L., et al., 2020b. An inverse latitudinal gradient in infection probability and phylogenetic diversity for Leucocytozoon blood parasites in New World birds. J. Anim. Ecol. 89, 423-435.
Fecchio, A., Chagas, C.R.F., Bell, J.A., Kirchgatter, K., 2020a. Evolutionary ecology, taxonomy, and systematics of avian malaria and related parasites. Acta Trop. 204, 105364.
Fecchio, A., Clark, N.J., Bell, J.A., Skeen, H.R., Lutz, H.L., De La Torre, G.M., et al., 2021. Global drivers of avian haemosporidian infections vary across zoogeographical regions. Global Ecol. Biogeogr. 30, 2393-2406.
Fecchio, A., Dias, R.I., Ferreira, T.V., Reyes, A.O., Dispoto, J.H., Weckstein, J.D., et al., 2022. Host foraging behavior and nest type influence prevalence of avian haemosporidian parasites in the Pantanal. Parasitol. Res. 121, 1407-1417.
Fecchio, A., Lima, M.R., Silveira, R., Braga, E.M., Marini, M.A., 2011. High prevalence of blood parasites in social birds from a Neotropical savanna in Brazil. Emu 111, 132-138.
Fridolfsson, A.K., Ellegren, H., 1999. A simple and universal method for molecular sexing of non-ratite birds. J. Avian Biol. 30, 116-121.
Ganser, C., Monadjem, A., McCleery, R.A., Ndlela, T., Wisely, S.M., 2020. Is it best on the nest? Effects of avian life-history on haemosporidian parasitism. Int. J. Parasitol. Parasites Wildl. 13, 62-71.
Gao, K., Zhou, B., Yang, L.X., Dong, L., Huang, X., Deng, W.H., 2021. How does circadian rhythm shape host-parasite associations? A comparative study on infection patterns in diurnal and nocturnal raptors. Diversity 13, 338.
Gibson, G., Torr, S.J., 1999. Visual and olfactory responses of haematophagous Diptera to host stimuli. Med. Vet. Entomol. 13, 2-23.
González, A.D., Matta, N.E., Ellis, V.A., Miller, E.T., Ricklefs, R.E., Gutiérrez, H.R., 2014. Mixed species flock, nest height, and elevation partially explain avian haemoparasite prevalence in Colombia. PLoS One 9, e100695.
Grillet, M-E., Villamizar, N.J., Cortez, J., Frontado, H.L., Escalona, M., Vivas-Martínez, S., et al., 2005. Diurnal biting periodicity of parous Simulium (Diptera: Simuliidae) vectors in the onchocerciasis Amazonian focus. Acta Trop. 94, 139-158.
Gupta, P., Vishnudas, C.K., Robin, V.V., Dharmarajan, G., 2020. Host phylogeny matters: examining sources of variation in infection risk by blood parasites across a tropical montane bird community in India. Parasites Vectors 13, 536.
Gutiérrez, J.S., Piersma, T., Thieltges, D.W., 2019. Micro- and macroparasite species richness in birds: the role of host life history and ecology. J. Anim. Ecol. 88, 1226-1239.
Gutiérrez, J.S., Rakhimberdiev, E., Piersma, T., Thieltges, D.W., 2017. Migration and parasitism: habitat use, not migration distance, influences helminth species richness in Charadriiform birds. J. Biogeogr. 44, 1137-1147.
Hammers, M., Komdeur, J., Kingma, S.A., Hutchings, K., Fairfield, E.A., Gilroy, D.L., et al., 2016. Age-specific haemosporidian infection dynamics and survival in Seychelles Warblers. Sci. Rep. 6, 29720.
Healy, K., Guillerme, T., Finlay, S., Kane, A., Kelly, S.B.A., McClean, D., et al., 2014. Ecology and mode-of-life explain lifespan variation in birds and mammals. Proc. R. Soc. A B 281, 20140298.
Hellgren, O., Pérez-Tris, J., Bensch, S., 2009. A jack-of-all-trades and still a master of some: prevalence and host range in avian malaria and related blood parasites. Ecology 90, 2840-2849.
Hellgren, O., Waldenström, J., Bensch, S., 2004. A new PCR assay for simultaneous studies of Leucocytozoon, Plasmodium, and Haemoproteus from avian blood. J. Parasitol. 90, 797-802.
Huang, X., Ellis, V.A., Jönsson, J., Bensch, S., 2018. Generalist haemosporidian parasites are better adapted to a subset of host species in a multiple host community. Mol. Ecol. 27, 4336-4346.
Isaksson, C., Sepil, I., Baramidze, V., Sheldon, B.C., 2013. Explaining variance of avian malaria infection in the wild: the importance of host density, habitat, individual life-history and oxidative stress. BMC Ecol. 13, 15.
Jetz, W., Thomas, G., Joy, J., Hartmann, K., Mooers, A., 2012. The global diversity of birds in space and time. Nature 491, 444-448.
Johnson, P.T.J., de Roode, J.C., Fenton, A., 2015. Why infectious disease research needs community ecology. Science 349, 1259504.
Jones, K.E., Patel, N.G., Levy, M.A., Storeygard, A., Balk, D., Gittleman, J.L., et al., 2008. Global trends in emerging infectious diseases. Nature 451, 990-993.
Julliard, R., Clavel, J., Devictor, V., Jiguet, F., Couvet, D., 2006. Spatial segregation of specialists and generalists in bird communities. Ecol. Lett. 9, 1237-1244.
Kamiya, T., O'Dwyer, K., Nakagawa, S., Poulin, R., 2014. What determines species richness of parasitic organisms? A meta-analysis across animal, plant and fungal hosts. Biol. Rev. 89, 123-134.
Knowles, S.C.L., Palinauskas, V., Sheldon, B.C., 2010a. Chronic malaria infections increase family inequalities and reduce parental fitness: experimental evidence from a wild bird population. J. Evol. Biol. 23, 557-569.
Knowles, S.C.L., Wood, M.J., Alves, R., Wilkin, T.A., Bensch, S., Sheldon, B.C., 2010b. Molecular epidemiology of malaria prevalence and parasitaemia in a wild bird population. Mol. Ecol. 20, 1062-1076.
Koprivnikar, J., Leung, T.L.F., 2015. Flying with diverse passengers: greater richness of parasitic nematodes in migratory birds. Oikos 124, 399-405.
Lachish, S., Knowles, S.C.L., Alves, R., Wood, M.J., Sheldon, B.C., 2011. Infection dynamics of endemic malaria in a wild bird population: parasite species-dependent drivers of spatial and temporal variation in transmission rates. J. Anim. Ecol. 80, 1207-1216.
Lee, K., Martin, L., Hasselquist, D., Ricklefs, R., Wikelski, M., 2006. Contrasting adaptive immune defenses and blood parasite prevalence in closely related Passer sparrows. Oecologia 150, 383-392.
Lee, M.Y., Hong, Y.J., Park, S.K., Kim, Y.J., Choi, T.Y., Lee, H., et al., 2008. Application of two complementary molecular sexing methods for East Asian bird species. Genes Genomics 30, 365-372.
Lehane, M., 2005. The Biology of Blood-Sucking in Insects, second ed. Cambridge
University Press, Cambridge.
Leung, T.L.F., Koprivnikar, J., 2016. Nematode parasite diversity in birds: the role of host ecology, life history, and migration. J. Anim. Ecol. 85, 1471-1480.
Lutz, H.L., Hochachka, W.M., Engel, J.I., Bell, J.A., Tkach, V.V., Bates, J.M., et al., 2015. Parasite prevalence corresponds to host life history in a diverse assemblage of Afrotropical bids and haemosporidian parasites. PLoS One 10, e0121254.
Marquardt, W.C., Demaree, R.S., Grieve, R.B., 2000. Parasitology and Vector Biology,
second ed. Academic Press, Cambridge.
Marzal, A., de Lope, F., Navarro, C., Møller, A.P., 2005. Malarial parasites decrease reproductive success: an experimental study in a passerine bird. Oecologia 142, 541-545.
Matthews, A.E., Ellis, V.A., Hanson, A.A., Roberts, J.R., Ricklefs, R.E., Collins, M.D., 2016. Avian haemosporidian prevalence and its relationship to host life histories in eastern Tennessee. J. Ornithol. 157, 533-548.
McClure, K.M., Fleischer, R.C., Kilpatrick, A.M., 2020. The role of native and introduced birds in transmission of avian malaria in Hawaii. Ecology 101, e03038.
McCurdy, D.G., Shutler, D., Mullie, A., Forbes, M.R., 1998. Sex-biased parasitism of avian hosts: relations to blood parasite taxon and mating system. Oikos 82, 303-312.
McNew, S.M., Barrow, L.N., Williamson, J.L., Galen, S.C., Skeen, H.R., DuBay, S.G., et al., 2021. Contrasting drivers of diversity in hosts and parasites across the tropical Andes. Proc. Natl. Acad. Sci. U.S.A. 118, e2010714118.
Merino, S., Moreno, J., Sanz, J.J., Arriero, E., 2000. Are avian blood parasites pathogenic in the wild? A medication experiment in Blue Tits (Parus caeruleus). Proc. R. Soc. B 267, 2507-2510.
Olalla‐Tárraga, M. Á., González-Suárez, M., Bernardo-Madrid, R., Revilla, E., Villalobos, F., 2016. Contrasting evidence of phylogenetic trophic niche conservatism in mammals worldwide. J. Biogeogr. 44, 99-110.
Olsson-Pons, S., Clark, N.J., Ishtiaq, F., Clegg, S.M., 2015. Differences in host species relationships and biogeographic influences produce contrasting patterns of prevalence, community composition and genetic structure in two genera of avian malaria parasites in southern Melanesia. J. Anim. Ecol. 84, 985-998.
Pagel, M., 1999. Inferring the historical patterns of biological evolution. Nature 401, 877-884.
Pérez-Rodríguez, A., Fernández-González, S., de la Hera, I., Pérez-Tris, J., 2013. Finding the appropriate variables to model the distribution of vector-borne parasites with different environmental preferences: climate is not enough. Global Change Biol. 19, 3245-3253.
Podmokła, E., Dubiec, A., Drobniak, S.M., Arct, A., Gustafsson, L., Cichoń, M., 2014. Determinants of prevalence and intensity of infection with malaria parasites in the Blue Tit. J. Ornithol. 155, 721-727.
Popescu, M., Trychta, M., Jackson, E., Selman, J., Houston, A., Collins, M., 2020. Avian haemosporidian prevalence and its relationship to host traits in Western Tennessee. J. Ornithol. 161, 995-1010.
Poulin, R., 2011. Evolutionary Ecology of Parasites. Princeton University Press,
Princeton.
R Core Team, 2021. R: A Language and Environment for Statistical Computing. R
Foundation for Statistical Computing, Vienna.
Revell, L.J., 2012. phytools: an R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 3, 217-223.
Rivero, A., Gandon, S., 2018. Evolutionary ecology of avian malaria: past to present. Trends Parasitol. 34, 712-726.
Rodriguez, M.D., Doherty, P.F., Piaggio, A.J., Huyvaert, K.P., 2021. Sex and nest type influence avian blood parasite prevalence in a high-elevation bird community. Parasites Vectors 14, 145.
Rodríguez-Hernández, K., Álvarez-Mendizábal, P., Chapa-Vargas, L., Escobar, F., GonzálezGarcía, F., Santiago-Alarcon, D., 2021. Haemosporidian prevalence, parasitaemia and aggregation in relation to avian assemblage life history traits at different elevations. Int. J. Parasitol. 51, 365-378.
Ruhs, E.C., Martin, L.B., Downs, C.J., 2020. The impacts of body mass on immune cell concentrations in birds. Proc. R. Soc. B 287, 20200655.
Santiago-Alarcon, D., Palinauskas, V., Schaefer, H.M., 2012. Diptera vectors of avian haemosporidian parasites: untangling parasite life cycles and their taxonomy. Biol. Rev. 87, 928-964.
Schultz, A., Underhill, L.G., Earlé, R.A., Underhill, G., 2010. Infection prevalence and absence of positive correlation between avian haemosporidian parasites, mass and body condition in the Cape Weaver Ploceus capensis. Ostrich 81, 69-76.
Scott, D.E., 2016. Raptor Medicine, Surgery and Rehabilitation, second ed. CABI Press,
Wallingford.
Sorensen, M.C., Dixit, T., Kardynal, K.J., Newton, J., Hobson, K.A., Bensch, S., et al., 2019. Migration distance does not predict blood parasitism in a migratory songbird. Ecol. Evol. 9, 8294-8304.
Tella, J.L., Blanco, G., Forero, M.G., Gajón, Á., Donázar, J.A., Hiraldo, F., 1999. Habitat, world geographic range, and embryonic development of hosts explain the prevalence of avian hematozoa at small spatial and phylogenetic scales. Proc. Natl. Acad. Sci. U.S.A. 96, 1785-1789.
Valkiūnas, G., 2005. Avian Malaria Parasites and Other Haemosporidian, second ed. CRC
Press, Boca Raton.
Valkiūnas, G., Iezhova, T.A., Križanauskienė, A., Palinauskas, V., Sehgal, R.N.M., Bensch, S., 2008. A comparative analysis of microscopy and PCR-based detection methods for blood parasites. J. Parasitol. 94, 1395-1401.
Vilela, B., Villalobos, F., 2015. LetsR: A new R package for data handling and analysis in macroecology. Methods Ecol. Evol. 6, 1229-1234.
Wang, Y.P., Song, Y.F., Zhong, Y.Q., Chen, C.W., Zhao, Y.H., Zeng, D., et al., 2021. A dataset on the life-history and ecological traits of Chinese birds. Biodivers. Sci. 29, 1149-1153.
Williams, M.A., Faiad, S., Claar, D.C., French, B., Leslie, K.L., Oven, E., et al., 2022. Life history mediates the association between parasite abundance and geographic features. J. Anim. Ecol. 91, 996-1009.
Wilman, H., Belmaker, J., Simpson, J., de la Rosa, C., Rivadeneira, M.M., Jetz, W., 2014. EltonTraits 1.0: species-level foraging attributes of the world's birds and mammals. Ecology 95, 2027.
Wood, M.J., Cosgrove, C.L., Wilkin, T.A., Knowles, S.C.L., Day, K.P., Sheldon, B.C., 2007. Within-population variation in prevalence and lineage distribution of avian malaria in Blue Tits, Cyanistes caeruleus. Mol. Ecol. 16, 3263-3273.
Xie, S.L., Lu, F., Cao, L., Zhou, W.Q., Ouyang, Z.Y., 2016. Multi-scale factors influencing the characteristics of avian communities in urban parks across Beijing during the breeding season. Sci. Rep. 6, 29350.
Yang, L.X., Zhou, B., Huang, X., Gao, K., Chen, Z.R., Dai, C., et al., 2022. A survey of sex ratios of raptors at a rescue center in China. J. Raptor Res. 56, 237-244.
Table
1.
Hypotheses and predictions linking host traits to probability of haemosporidian infection in raptors.
Predictors
Category
General hypotheses and predictions
Activity pattern
Biological variable
Unknown
Age
Biological variable
Adult raptors are expected to have a higher probability of haemosporidian infection than juveniles, because they might encounter more parasite species during their lifetimes (Wood et al., 2007).
Body mass
Biological variable
Body mass would either be positively associated with the probability of haemosporidian infection due to increased vector attraction (Gibson and Torr, 1999; Lehane, 2005), or negatively associated with the probability of haemosporidian infection as larger birds displayed better immune responses to constrain parasite invasion (Ruhs et al., 2020).
Dietary diversity
Biological variable
Raptors with a broader diet are expected to have a higher probability of haemosporidian infection than more specialized ones, because they might encounter more parasite species during their lifetimes (Leung and Koprivnikar, 2016).
Geographic range
Ecological variable
Raptors with larger geographic ranges are expected to have a higher probability of haemosporidian infection. Because larger geographic ranges offer more opportunities for host–parasite associations to become established and encompass a greater diversity of habitats suitable for different parasites (Tella et al., 1999).
Habitat specialization
Ecological variable
Raptors with broader habitat breadths are expected to have a higher probability of haemosporidian infection than more specialized ones, because they might encounter more parasite species during their lifetimes (Leung and Koprivnikar, 2016).
Migration distances
Ecological variable
Raptors with longer migration distances are expected to have a higher probability of haemosporidian infection, because migration may bring hosts in contact with "new" parasites (Koprivnikar and Leung, 2015; de Angeli Dutra et al., 2021).
Nest site
Ecological variable
A higher probability of haemosporidian infection would be expected in raptors that nest near canopies, where the vectors are more abundant (Černý et al., 2011).
Nest type
Ecological variable
Raptors with nests are expected to have a higher probability of haemosporidian infection, because nests increase the frequency of contact with haemosporidian vectors (Ganser et al., 2020; Fecchio et al., 2022).
Sex
Biological variable
Female raptors trading reproduction for immunity would be expected to have a higher probability of haemosporidian infection than males (McCurdy et al., 1998).
Table
2.
The top ten generalized linear mixed-effects models (GLMMs) for the effect of biological and ecological variables on haemosporidian parasites infection of raptor species.
1
2
3
4
5
6
7
8
9
10
Activity pattern
*
*
*
*
*
*
*
*
*
*
Age
*
*
*
*
*
*
*
*
*
*
Body mass
*
*
*
*
*
*
*
*
*
*
Dietary diversity
*
*
*
*
*
*
Geographic range
*
*
*
*
*
*
*
*
*
*
Habitat specialization
*
*
*
Migration distances
*
*
*
*
*
*
*
*
*
Nest site
*
*
Nest type
*
*
*
*
*
*
*
*
*
*
Sex
*
*
*
K
7
6
8
8
7
7
6
9
7
8
AICc
1271.05
1271.14
1272.34
1272.81
1272.96
1273.07
1273.57
1274.02
1274.09
1274.4
ΔAIC
0
0.09
1.29
1.76
1.91
2.02
2.52
2.97
3.04
3.35
Wi
0.14
0.14
0.08
0.06
0.06
0.05
0.04
0.03
0.03
0.03
R2 marginal
0.27
0.26
0.27
0.27
0.26
0.26
0.27
0.27
0.27
0.27
R2 conditional
0.28
0.27
0.28
0.28
0.28
0.27
0.28
0.28
0.28
0.28
Note: *, indicates that a variable is included in the model. K, number of predictors in the model. ΔAIC, the difference between each model and the highest ranked model. AICc, Akaike's information criterion adjusted for small sample sizes. Wi (Akaike weights), the probability that a model is best given the particular set of models considered. R2 marginal, amount of variation that is explained by fixed factors (covariates). R2 conditional, amount of variation that is explained by both fixed and random factors. Models include sampling date and species as a random effect. Models are ranked in order of decreasing ΔAIC.
Table
3.
Model averaging based on GLMMs (1024 models) using biological and ecological variables, age, nest type, activity pattern, geographic range size and migration distance, to explain haemosporidian parasites infection of raptor species.
β
SE
95% CI (lower, upper)
Relative importance
Age (HY)
−1.46
0.19
−1.84, −1.09
0.92
Nest type (open-nesting)
2.05
0.58
0.91, 3.19
0.92
Activity pattern (nocturnal)
2.34
0.49
1.38, 3.31
0.92
Geographic range
−0.35
0.11
−0.57, −0.13
0.92
Migration distances
0.23
0.10
0.04, 0.42
0.82
Body mass
−0.24
0.13
−0.48, 0.01
0.81
Dietary diversity
0.16
0.10
−0.04, 0.36
0.54
Habitat specialization
0.08
0.11
−0.14, 0.30
0.29
Sex (M)
−0.07
0.14
−0.36, 0.21
0.26
Nest site (cliff)
0.11
0.42
−0.72, 0.95
0.15
Nest site (ground)
0.54
0.67
−0.77, 1.86
0.15
Note: Bold characters indicate the most important variables through GLMMs. β, model-averaged regression coefficients. HY, hatch-year. Open-nesting, raptors build an open-cup nest. Nocturnal, nocturnal raptors. M, male raptors. Cliff, raptors nest on the cliff. Ground, raptors nest on the ground.

 DownLoad:
DownLoad:






 Email Alerts
Email Alerts RSS Feeds
RSS Feeds