Lirong Zuo, Ibrahim M. Ahmad, Yuanyuan Liu, Limin Wang, Shu Fang, Dongming Li. 2023: Effects of constant light and dark conditions on the locomotor activity, body mass, and body temperature rhythms of Eurasian Tree Sparrows (Passer montanus). Avian Research, 14(1): 100126. DOI: 10.1016/j.avrs.2023.100126
Citation:
Lirong Zuo, Ibrahim M. Ahmad, Yuanyuan Liu, Limin Wang, Shu Fang, Dongming Li. 2023: Effects of constant light and dark conditions on the locomotor activity, body mass, and body temperature rhythms of Eurasian Tree Sparrows (Passer montanus). Avian Research, 14(1): 100126. DOI: 10.1016/j.avrs.2023.100126
Lirong Zuo, Ibrahim M. Ahmad, Yuanyuan Liu, Limin Wang, Shu Fang, Dongming Li. 2023: Effects of constant light and dark conditions on the locomotor activity, body mass, and body temperature rhythms of Eurasian Tree Sparrows (Passer montanus). Avian Research, 14(1): 100126. DOI: 10.1016/j.avrs.2023.100126
Citation:
Lirong Zuo, Ibrahim M. Ahmad, Yuanyuan Liu, Limin Wang, Shu Fang, Dongming Li. 2023: Effects of constant light and dark conditions on the locomotor activity, body mass, and body temperature rhythms of Eurasian Tree Sparrows (Passer montanus). Avian Research, 14(1): 100126. DOI: 10.1016/j.avrs.2023.100126
Effects of constant light and dark conditions on the locomotor activity, body mass, and body temperature rhythms of Eurasian Tree Sparrows (Passer montanus)
Hebei Key Laboratory of Animal Physiology, Biochemistry and Molecular Biology, College of Life Sciences, Hebei Normal University, Shijiazhuang, 050024, Hebei Province, China
b.
Hebei Collaborative Innovation Center for Eco-Environment, Hebei Normal University, Shijiazhuang, 050024, China
Funds: This work was supported by the National Natural Science Foundation of China (NSFC, 31971413) and the Natural Science Foundation of Hebei Province (C2020205038) to Dongming Li
Hebei Key Laboratory of Animal Physiology, Biochemistry and Molecular Biology, College of Life Sciences, Hebei Normal University, Shijiazhuang, 050024, Hebei Province, China. E-mail address: lidongming@hebtu.edu.cn (D. Li)
Light is an essential environmental cue influencing the endogenous circadian clocks that regulate behavioral and physiological processes in animals. Despite extensive research on the circadian rhythms of avian behavior and physiology, the mechanisms by which they adapt and adjust to abnormal photoperiod conditions, such as artificial light, have yet to be fully elucidated. Here, the circadian rhythms of the body mass, activity, and core and peripheral body temperatures of the Eurasian Tree Sparrow (Passer montanus, ETS)—a diurnal passerine—were examined under three different lighting conditions: constant light (LL), constant darkness (DD), and normal light–dark cycles (LD). Our results showed that the activity, body mass, core, tarsometatarsus, and eye temperatures of the ETSs varied significantly with treatment, time, and the interaction between treatment and time; however, there was no significant change in body mass in response to the interaction between time and treatment. Under LD conditions, the ETSs exhibited significant circadian rhythms in activity and core, tarsometatarsus, and eye temperatures. Under LL conditions, only the core temperature exhibited significant rhythmicity. Under DD conditions, there were no significant circadian rhythms in activity, body mass, and eye temperature; however, the core and tarsometatarsus temperatures remained rhythmic. Our results indicate that the core body temperature rhythm of ETS is endogenous and can be maintained under abnormal photoperiods. In contrast, the activity and body surface temperature, uncoupled from the core body temperature under abnormal photoperiod conditions, strongly depend on light. These results differ from those of other diurnal avian species, suggesting a unique coping mechanism of ETS in response to an abnormal photoperiod.
Light is an essential environmental cue that affects the daily and seasonal activities of most free-living animals (Dixit and Singh, 2011; Dominoni, 2015; Grunst et al., 2023). Hence, the endogenous circadian clocks of animals are modulated to synchronize behavioral and physiological processes in response to exogenous diurnal cycles (Prabhat et al., 2020). However, changes in daily photoperiod regimes, such as exposure to artificial light at night, can disrupt this synchronization and lead to circadian rhythm desynchrony (Wyse et al., 2011; Spoelstra et al., 2018). This disparity between the internal physiology, behavior, and endogenous circadian clock of an animal can negatively affect its overall fitness, leading to energy loss, metabolic dysfunction, and neuroendocrinological disorders (Regas, 1998; West et al., 2017; Porcu et al., 2018). Body mass, behavioral activity, and core and peripheral body temperatures are primary indicators of energy metabolism and proper functioning of the neuroendocrinological system. By monitoring the changes in these parameters in response to abnormal photoperiod treatments, we can better understand the behavioral strategies and underlying metabolic mechanisms of animals. For example, the body temperature, body mass, and activity rhythms of birds may either be maintained or modified when the environmental photoperiod changes, especially under constant light (LL) or darkness (DD) conditions (Budki et al., 2012). However, the mechanisms by which birds adjust these parameters in response to abnormal photoperiod conditions are not fully understood.
In vertebrates, the 24-h light–dark (LD) cycle is maintained through humoral and neural loops in a cascading manner (Tosini and Menaker, 1995; Buhr and Takahashi, 2013). The retinae, suprachiasmatic nuclei (SCN) of the hypothalamus, and pineal gland of non-mammalian vertebrates possess photoreceptors and are in close cooperation to ensure well-coordinated circadian precision and amplitude (Tosini and Menaker, 1995; Cassone et al., 2009; Cassone, 2014); in contrast, the SCN solely acts as the master pacemaker in mammals (Bronstein et al., 1990). Meanwhile, the hypothalamus and pineal gland of passerines play significant roles in resynchronizing their rhythms when the timing of LD cycles suddenly changes (Binkley and Mosher, 1986). On the other hand, the retina has a significant effect on the circadian rhythm of some non-passerine birds, such as the Japanese Quail (Coturnix japonica) and Rock Pigeon (Columba livia) (Brandstaetter, 2002). Generally, by integrating external LD cycles with humoral and neural loops, animals can predict and adapt to different times of the day (24 h) and, thus, maintain the circadian rhythms regulating their body mass, body temperature, and locomotion. Previous studies reported significant circadian rhythms influencing the locomotor activity and body temperature of mammals and birds (both passerines and non-passerines). A closely coupled oscillation between the core body temperature and the activity of an animal has been detected (Refinetti and Menaker, 1992; Depres-Brummer et al., 1995; Fonken et al., 2010). For example, improving the foraging success rate and utilization efficiency of food resources is closely synchronized with metabolism (Montoya et al., 2010), and the body temperature and activity of mammals were positively correlated (van Jaarsveld et al., 2019).
However, the behavioral and physiological rhythms of animals are disrupted under LL conditions (Le Tallec et al., 2013; Fonken and Nelson, 2014). The circadian rhythms of body temperature and daily activities are two fundamental variables derived from the radical changes in neuroendocrinology, metabolism (Zhang et al., 2019), and behavior (Jiang et al., 2020). For instance, the circadian rhythm affecting the activity and core body temperature of mammals becomes arrhythmic and dissociated under LL conditions (Depres-Brummer et al., 1995; Benstaali et al., 2001; Fonken et al., 2010). In contrast, the responses of birds to LL conditions are highly phylogeny-dependent. For example, in Columbiformes, Pigeons (Aplopelia Bonaparte) maintain activity and body temperature rhythm and closely coupled vibration under LL conditions, however, their rhythmicity is lost (Yamada et al., 1988). Moreover, passerines such as House Sparrows (Passer domesticus) lose their activity and body temperature rhythms (Binkley et al., 1971); however, White-crowned Sparrows (Zonotrichia leucophrys gambelii) retain their locomotor rhythms for the first three days (Coverdill et al., 2008). Similarly, the activity rhythms of Java Sparrows (Padda oryzivora) and Indian Weaverbirds (Ploceus philippinus) persist (Ebihara and Kawamura, 1981; Pandey and Bhardwaj, 2011). These findings indicate that activity and body temperature rhythms are tightly coupled in passerines; however, such a phenomenon has not been consistently observed in different passerine species.
Under DD conditions, some mammals maintain their circadian rhythm oscillations. For example, the diurnal Desert Goat (Capra hircus) maintains its activity and body temperature rhythms but exhibits a free-running circadian rhythm (Farsi et al., 2020). Nocturnal mice and rats also maintain the circadian rhythms of activity, body temperature, melatonin, and neurons under DD conditions (Wideman and Murphy, 2009b; Stenvers et al., 2016). Galliformes species such as the diurnal Japanese Quails (Underwood et al., 1999) and laying Hens (Gallus gallus domesticus) maintain the circadian rhythm of body temperature; however, the amount of locomotor activity was almost undetectable, and the activity rhythm was not sustained (Boshouwers and Nicaise, 1987). Passeriformes species such as diurnal House Sparrows exhibit rhythmic locomotor activity and body temperature (Gaston and Menaker, 1968), and diurnal European Starlings (Sturnus vulgaris) maintain their activity, feeding and melatonin rhythms (Gwinner, 1978). Therefore, nocturnal and diurnal animals can maintain circadian rhythms under DD conditions.
The Eurasian Tree Sparrow (Passer montanus, ETS) is a representative diurnal resident bird species that is widely distributed across Eurasia (Dixit and Singh, 2012). It is commonly used as a model species to investigate the circadian rhythms of avian behavior and physiology (Ravikumar and Tewary, 1990), seasonal rhythms of baseline and stress-induced corticosterone levels (Li et al., 2008, 2011), and testosterone and gonadal hormone-releasing hormone levels (Li et al., 2012, 2017; Dixit et al., 2022; Wang et al., 2022). They also exhibit a typical circadian rhythm of hormone secretion (Jiang et al., 2020). The closely related species of ETS, the House Sparrow lost its activity and body temperature rhythms under LL conditions (Binkley et al., 1971), but could maintain those under DD conditions (Gaston and Menaker, 1968). The ETSs are therefore predicted to exhibit a similar pattern of circadian rhythms of behavioral activity and body temperature under abnormal photoperiod conditions. To validate this hypothesis, we examined the circadian rhythms of activity and core and peripheral body temperatures of ETSs under LD, LL, and DD conditions. We predicted that the rhythmicity of activity and body temperature under the DD conditions could be maintained, but their rhythms would disappear under the LL conditions.
2.
Materials and methods
2.1
Bird collection
Through the use of mist nets, 21 free-living juvenile ETSs were caught in the wild in Shijiazhuang City, Hebei Province, China (38.03° N, 114.48° E; elevation: 112 m) in June 2018. After the ETSs were captured, they were randomly assigned to three groups (n = 7/group) and individually housed in similar-sized (width × length × height: 30 cm × 40 cm × 20 cm) cages. The ETSs were then acclimatized for five days under the normal LD cycle (12 h: 12 h) in three climate-controlled chambers under uniform temperature (28 ℃) and humidity (40–60%). Food (millets) and water were provided ad libitum. All protocols in this study were approved by the Institutional Animal Care and Use Committee and the Ethics and Animal Welfare Committee of Hebei Normal University, China.
2.2
Bird treatment
After acclimatization, the light cycles of one climate-controlled chamber remained unchanged (LD group). In contrast, the other two climate-controlled chambers were adjusted to either constant bright light (LL group) or constant darkness (DD group). The light intensity was 300–400 lx (similar to sunset and sunrise intensities) in the LL and LD groups. In order to record the activity, the DD group was exposed to 850 nm wavelength infrared light. The ZT0/ZT12 (zeitgeber, lights on/off) for the LD group was 6:40 and 18:40, respectively. Food and water were provided ad libitum to the ETSs in the three groups.
2.3
Body mass, activity, and body temperature monitoring
The core and peripheral body temperatures, activity, and body mass of the ETSs were monitored for seven consecutive days. The activity in the three groups was monitored for 24 h each day using a Big Brother circadian rhythm system (COULBOURN, ACT-400, Actimetrics, Wilmette, IL, USA). In each climate room, a surveillance camera was mounted to record the activity of the ETSs under dark conditions, and infrared light was irradiated to replenish light. The bird cages were placed on three 3D shelves, and the activity of each bird was monitored using cameras for 24 h for seven consecutive days. All data were exported to the Big Brother Viewer software, and the activity rhythms were analyzed. The body mass of each bird was measured using a portable digital balance (nearest to 0.01 g) at 2:00, 6:00, 10:00, 14:00, 18:00, and 22:00 each day. The temperature of the cloaca (representing the core body temperature) was measured using a handheld thermometer (nearest to 0.01 ℃; HH-25KC; Omega Engineering, Norwalk, CT, USA) at 2:00, 6:00, 10:00, 14:00, 18:00, and 22:00 each day. The eye and tarsometatarsus surface temperatures (representing the peripheral body temperature) were measured using an infrared thermal imager (TiS20+; Fluke Corporation, Everett, WA, USA) at 2:00, 6:00, 10:00, 14:00, 18:00, and 22:00 each day. To ensure accurate temperature measurements and obtain the best image quality, we preheated the thermal imager for 10 min before placing it on the eyes and tarsometatarsus.
2.4
Statistical analysis
The effects of different light treatments and time points on the activity, body mass, core, tarsometatarsus, and eye temperatures of the ETSs were analyzed using a mixed linear model. Multiple comparisons in each treatment and six-time points were performed using Bonferroni post-hoc tests, with time points defined as repeating variables. The JTK_Cyclev.3.1 function in the R package (Hughes et al., 2010) was used to calculate the circadian rhythm parameters, including acrophase (φ), amplitude (A), and Benjamini–Hochberg Q value (BH.Q, a method for optimizing false discovery rates), for the activity, body mass, and core, eye, and tarsometatarsus temperatures. A BH.Q < 0.05 means that there is a significant circadian rhythm. All cosine curves were fitted using the formula f(t) = M + Acos (tπ/12−φ), including the mesor (M), amplitude (A), and acrophase (φ). All statistical analyses were performed in IBM SPSS Statistics v.23 (IBM Inc., NY, USA) and R 4.3.0 (Hughes et al., 2010). All figures were plotted using GraphPad Prism 8.0 (GraphPad Software, Inc., CA, USA). The data are expressed as the mean ± SEM.
3.
Results
3.1
Activity, body mass, and core and peripheral temperatures in response to abnormal photoperiod
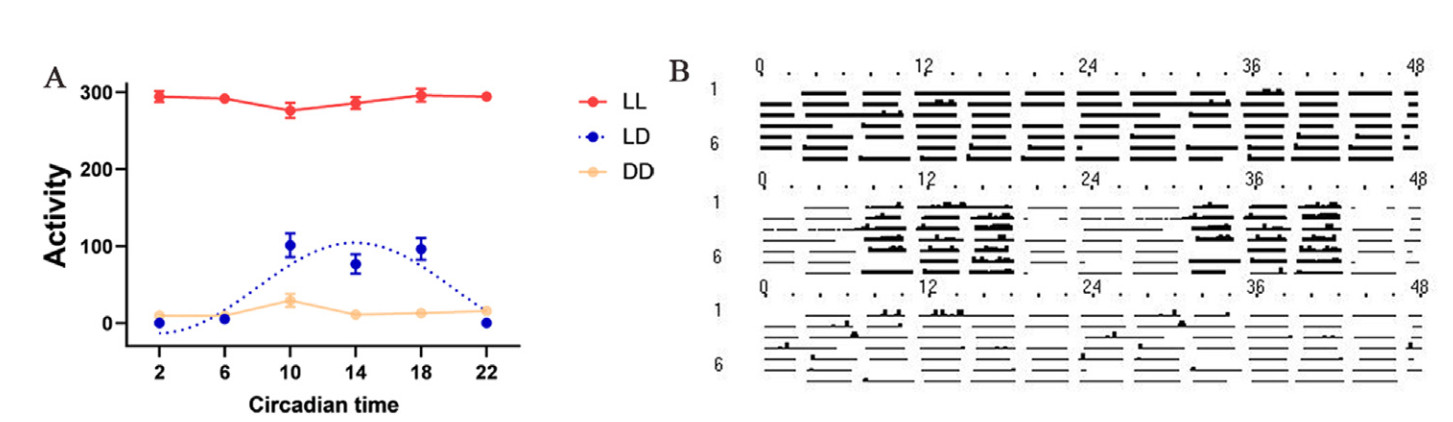
The activity, body mass, and core, tarsometatarsus, and eye temperatures of the ETSs varied significantly with treatment, time, and the interaction between treatment and time; however, there was no significant change in body mass in response to the interaction between time and treatment (Table 1; Appendix Table S1). The post-hoc results revealed that regardless of time, the birds in the LL group were significantly more active than those in the LD and DD groups (Fig. 1; Table 2; Appendix Table S2). The birds in the LD group were significantly more active during the day and less active at 22:00 than those in the DD group; however, there was no significant difference in activity between the LD and DD groups at 2:00 and 6:00 (Fig. 1; Table 2; Appendix Table S2).
Table
1.
The statistical results of treatment, time, and the interaction of treatment and time on activity, body mass, core temperature, eye temperature, and tarsometatarsus temperature of Eurasian Tree Sparrows (Passer montanus) in mixed linear models.
Figure
1.
Changes of activity of Eurasian Tree Sparrows (Passer montanus) under constant light (LL) and darkness (DD), and normal light-dark conditions (LD). (A) Average activity of each bird for eight consecutive days under a 24-h cycle (2:00, 6:00, 10:00, 14:00, 18:00, and 22:00). (B) Double-plotted actogram of daily rhythms of activity under a 48-h cycle. The cosine curve fitted in the figure represents a significant rhythm (BH.Q < 0.05). Data are shown as means ± SEM.
Table
2.
Statistical results of the average of activity, body mass, core temperature, tarsometatarsus temperature, and eye temperature of Eurasian Tree Sparrows (Passer montanus) under constant light (LL) and darkness (DD), and normal light-dark conditions (LD) at different time points (2:00, 6:00, 10:00, 14:00, 18:00, and 22:00).
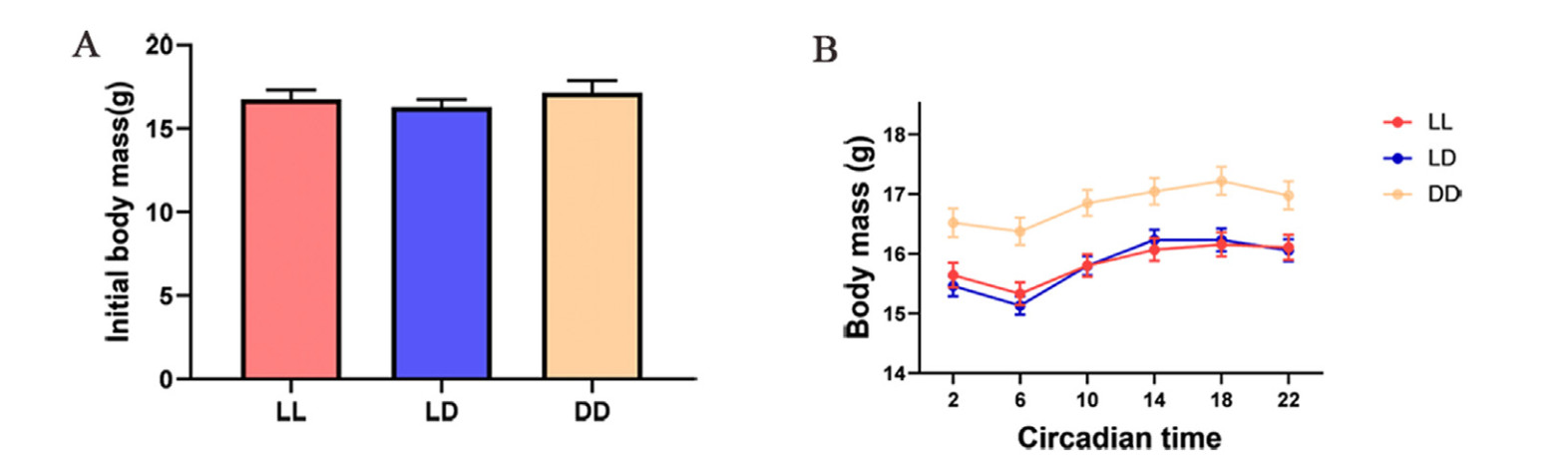
The initial body mass of the birds in the three groups did not significantly differ (F = 0.557, P = 0.583; Fig. 2). However, the birds in the DD group were significantly heavier than those in the LD and LL groups; meanwhile, the body mass of the birds in the LL and LD groups did not significantly differ at any time point (Fig. 2; Table 2; Appendix Table S2).
Figure
2.
Comparisons of initial body mass (A) and the changes of body mass (B) at six-time points (2:00, 6:00, 10:00, 14:00, 18:00, and 22:00) in a 24-h cycle under constant light (LL) and darkness (DD), and normal light-dark conditions (LD) of Eurasian Tree Sparrows (Passer montanus). Data are shown as means ± SEM.
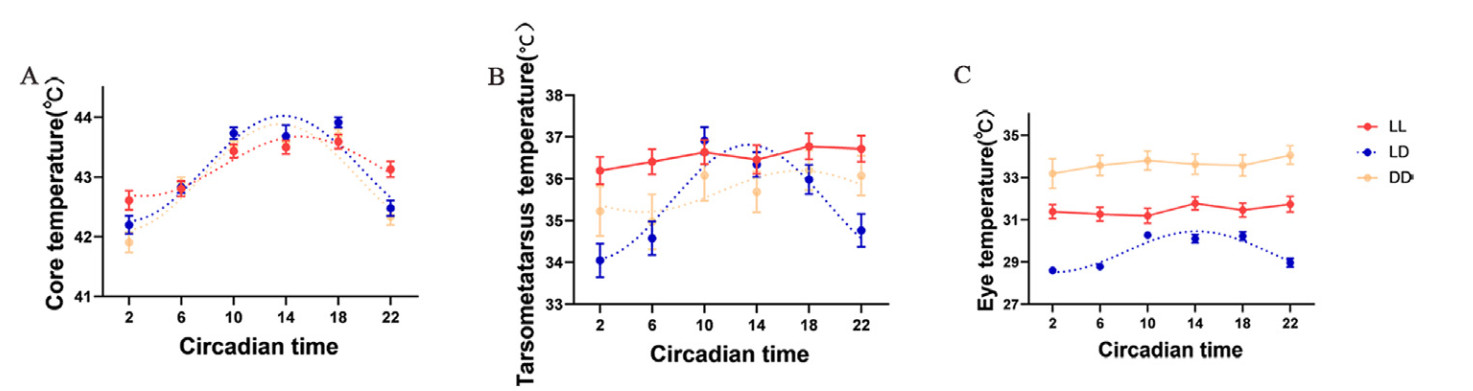
The core temperature of the LL group was significantly higher than that of the DD group at 2:00 (P = 0.002), and that of the LD (P = 0.003) and DD groups (P = 0.001) at 22:00; in contrast, it was significantly lower than that of the LD group at 10:00 (P = 0.041). The core temperature of the three groups did not significantly differ at other time points (Fig. 3; Table 2; Appendix Table S2). Although there were no significant differences in tarsometatarsus temperature across the groups at other time points, the LL group had a significantly higher tarsometatarsus temperature than the LD group at 2:00, 6:00, and 22:00. The eye temperature of the LL and LD groups was significantly lower than that of the DD group at all time points; moreover, the eye temperature of the LL group was significantly higher than that of the LD group at all time points except at 10:00 (Fig. 3; Table 2; Appendix Table S2).
Figure
3.
Daily changes of average values in core temperature (A), tarsometatarsus temperature (B), and eye temperature (C) at six-time points (2:00, 6:00, 10:00, 14:00, 18:00, and 22:00) under constant light (LL) and darkness (DD), and normal light-dark conditions (LD) of Eurasian Tree Sparrows (Passer montanus). The cosine curve fitted in the figure represents a significant rhythm (BH.Q < 0.05). Data are shown as means ± SEM.
3.2
Circadian rhythms of activity, body mass, and core and peripheral temperatures in response to constant photoperiod
Except for body mass, the ETSs in the LD group exhibited significant circadian rhythms in activity and core and tarsometatarsus temperatures, with acrophase at 12:00, and in eye temperature, with acrophase at 14:00 (Table 3). The rhythmicity of these measured parameters varied with the treatment duration. There were no significant circadian rhythms in the activity, body mass, and tarsometatarsus and eye temperatures in the LL group; however, the core temperature exhibited significant rhythmicity, with acrophase at 14:00 (delayed by 2 h relative to the LD group; Table 3). Lastly, there were no significant circadian rhythms in the activity, body mass, and eye temperature in the DD group; however, the core and tarsometatarsus temperatures exhibited significant rhythmicity, with acrophases at 12:00 and 16:00, respectively (Table 3). Compared with the LD group, the amplitudes of all measured parameters decreased in the LL and DD groups (Table 3).
Table
3.
Circadian rhythm parameters of activity, body mass, core temperature, tarsometatarsus temperature, and eye temperature of Eurasian Tree Sparrows (Passer montanus) under constant light (LL) and darkness (DD), and normal light-dark cycle (LD) conditions.
Factors
Variable
BH.Q
ADJ.P
Acrophase (φ, h)
Amplitude (A)
df
F
P
Activity
LL
1.000
1.000
22
6.393
5275
0.725
0.606
LD
< 0.05
< 0.001
12
34.850
5276
24.163
< 0.001
DD
0.072
0.035
12
5.922
5276
1.184
0.323
Body mass
LL
1.000
0.759
16
0.362
5364
3.027
0.013
LD
0.679
0.144
16
0.540
5372
8.655
< 0.001
DD
1.000
0.882
16
0.360
5401
2.089
0.071
Core temperature
LL
< 0.05
< 0.001
14
0.478
5413
7.669
< 0.001
LD
< 0.05
< 0.001
12
0.889
5422
37.124
< 0.001
DD
< 0.05
< 0.001
12
0.714
5444
32.552
< 0.001
Tarsometatarsus temperature
LL
1.000
1.000
12
0.191
5391
0.480
0.790
LD
< 0.05
< 0.001
12
1.332
5363
9.814
< 0.001
DD
0.029
0.019
16
0.470
5190
0.878
0.502
Eye temperature
LL
0.170
0.113
18
0.452
5388
0.587
0.710
LD
< 0.05
< 0.001
14
1.048
5363
24.730
< 0.001
DD
0.793
0.793
14
0.171
5190
0.243
0.941
Significant values (BH.Q < 0.05; P < 0.05) are shown in bold.
As predicted, the activity, core and peripheral body temperatures of the ETSs had significant circadian rhythms and were closely coupled under LD conditions; however, eye temperature was acrophase-delayed by approximately 2 h. In response to an abnormal photoperiod, the amplitude of concussion for each index under all treatments weakened. Under LL conditions, only the core temperature exhibited significant rhythmicity, whereas under DD conditions, the core and tarsometatarsus temperatures displayed significant rhythmicity. These results indicate that the circadian rhythm of the core body temperature of ETSs is endogenous, well-maintained, and independent of the photoperiod.
4.1
Circadian rhythms under normal conditions
Under LD conditions, the ETSs were extremely active during the day and inactive at night; in particular, locomotor activity was significantly rhythmic and acrophased at approximately 12:00. They fed during the day, significantly gained weight while fasting, and lost weight at night. Unlike nocturnal animals, diurnal animals forage, digest food, and absorb nutrients to promote anabolism during the day (Wideman and Murphy, 2009). Diurnal birds are inactive at night and rely on the mobilization of their glycogen and fat stores to enhance catabolism (Lucia et al., 2010). Our results confirm that diurnal animals are highly photosensitive: their activities rapidly increase when lights are on, but their activities end when lights are off (Dawson et al., 2001; Trivedi et al., 2005; Dawson and Sharp, 2007). Although no significant rhythmicity was detected in the body mass of the ETSs, there were significant differences in body mass at daytime and nighttime. Our results are congruent with the reports that Light-vented Bulbul (Pycnonotus sinensis), White-throated Sparrow (Z. albicollis) (Kontogiannis, 1967), and Marsh Tit (Poecile palustris) (Hurly, 1992) gained weight during the day, but lost weight at night. Such circadian rhythms in activity and the corresponding changes in body mass indicate endogenous behavioral and physiological processes of animals to adapt to the external diurnal cycles of light signals (Prabhat et al., 2020).
Meanwhile, we found that both the core and peripheral body temperatures—tarsometatarsus and eye temperatures—were remarkably rhythmic, with highly synchronized acrophases at 12:00 and 14:00, respectively. The core body temperature rhythm was synchronized with locomotor activity, indicating that they are highly coupled. The tarsometatarsus temperature had a higher amplitude and reached its peak synchronizing with the core body temperature, while the eye temperature reached its peak approximately 2 h later. Although birds can maintain an equilibrium of heat production and loss at a constant temperature in their surrounding environment, heat loss generally lags behind the core temperature (Giloh et al., 2012). Since the core body temperature is coupled with metabolic rate, the synchronicity of body temperature with locomotor activity is essential in controlling metabolic processes, ensuring that endogenous shock is synchronized with the external environment (Al-Hasani et al., 2003). All these temporal synchronizations to predictable LD cycles are essential in optimizing the fitness of organisms (Ramkisoensing and Meijer, 2015; Plano et al., 2017).
4.2
Circadian rhythms under constant light conditions
Compared with the birds under LD conditions, the birds under LL conditions lost the rhythmicity of their locomotor activity. Our results indicate that their activities are highly light signal–dependent (Dawson et al., 2001). Furthermore, our results are consistent with the findings for other avian species such as Rock Pigeons (Yamada et al., 1988), Domestic Chicken (Gallus domesticus), Ptarmigans (Lagopus muta) (Appenroth et al., 2021), and resident House Sparrows (Binkley et al., 1971). However, our results differ from the finding for migratory White-crowned Sparrows: under LL conditions, they maintain the rhythmicity of their locomotor activity for three days (Coverdill et al., 2008). Therefore, it seems that migratory and resident passerines exhibit different behavioral strategies in response to certain environmental signal changes that may be caused by different interactions between the output of the circadian oscillation and light signal.
The ETSs under LL conditions had significantly higher core and peripheral body temperatures during the subjective night than those of the LD group during the dark period. However, their body mass did not change. Increased body temperature and activity, coupled with constant body mass, denote that the ETSs increased their food intake under constant light conditions at night; however, they demonstrated a similar trend under normal conditions. In contrast, nocturnal mice increased food intake and gained weight under light-disturbed conditions (Coomans et al., 2013). Furthermore, the core body temperature of the ETSs remained rhythmic, consistent with those of Pigeons (Ebihara et al., 1984; Oshima et al., 1989). However, the peripheral body temperature was arrhythmic. These results indicate that the rhythmicity of the core body temperature is endogenous, and peripheral body temperatures and activity can be decoupled under LL conditions. Relative to normal light conditions, the core body temperature of the ETSs under LL conditions was acrophase-delayed for approximately 2 h, indicating that its rhythmicity was affected by light, and its oscillation process was under the strict control of the endogenous biological clock. The core body temperature of the ETSs in the LL group during the subjective night was higher than that in the LD group, and the trough of the core body temperature rhythm occurred during the subjective night. This downregulation of metabolism has also been observed in nocturnal rats exposed to dim light (Borniger et al., 2014). In terms of energy expenditure, constant light exposure is a costly inconvenience because core body temperature rhythms can help an animal conserve energy; for example, a rhythmic core temperature helps House Sparrows save 7% of their energy (Regas, 1998).
Generally, light signals can serve as external stimuli that induce internal physiological and behavioral changes by acting on the retinal photoreceptors of birds. When continuously exposed to constant light, the ETSs demonstrated behavioral changes and a higher level of activity, resulting in increased metabolic heat production and peripheral temperature. Unlike the core body temperature, controlled by the endogenous biological clock, the peripheral temperature strongly depends on exogenous light. Under LL conditions, the surface body temperature of the ETSs remained high, which could be induced by the increased activity at subjective night and enhanced thermogenesis through catabolism (Hohtola and Stevens, 1986). Therefore, the activity and body temperature rhythms of birds are disrupted under continuous exposure to intense light. Such a disruption may alter the homeostasis of energy metabolism and further impair the overall health of an organism. Considering the limited information on variations in peripheral body temperature, the underlying mechanisms of the decoupled relationship between peripheral and core body temperature require further investigation.
4.3
Circadian rhythms in constant dark conditions
The ETSs remained inactive at all time points and lost their activity rhythm under DD conditions. Similarly, White-crowned Sparrows lose their activity rhythm under dim light conditions; however, they maintain a high level of activity for night migration (McMillan, 1972; Coverdill et al., 2008). Our results differ from the findings that locomotor activity rhythms are maintained under DD conditions in House Sparrows (Cassone, 2014) and European Starlings (Gwinner, 1978). These results indicate that maintaining the activity rhythm under DD conditions is largely species-specific, not phylogenetic-dependent. In particular, the ETSs in the DD group were significantly heavier than those in the LD and LL groups, although they exhibited a similar trend in body mass as those in the LD group. Despite the disruption of their activity and foraging rhythms, an endogenous pattern of weight loss and gain was observed when the activities and energy consumption levels of the ETSs were low. The mechanism behind this phenomenon remains largely unknown and should be determined through ecological and physiological research.
The ETSs in the DD group had comparable core body and tarsometatarsus temperatures with those in the LD and LL groups; however, their eye temperature was significantly higher than those in the other two groups. Owing to weak vision under dark conditions, an increase in eye temperature under DD conditions is critical for better foraging, thereby ensuring adequate food intake and maintenance of energy balance (Harmening, 2017). Furthermore, the core body and tarsometatarsus temperatures exhibited a significant circadian rhythm, whereas eye temperature did not. Similarly, the circadian rhythm of the core body temperature under DD conditions has been observed in Japanese Quails and House Sparrows (Gaston and Menaker, 1968; Underwood et al., 1999). The core temperature of the ETSs with an acrophase at 12:00 indicates that constant darkness is not necessary to induce changes in the rhythmicity of the core body temperature of birds because it is controlled by an endogenous clock. However, our results showed that the acrophase of the tarsometatarsus temperature was delayed by 4 h relative to the core body temperature. Although birds can maintain an equilibrium of heat production and loss at a constant temperature in their surrounding environment, heat loss generally lags behind the core temperature (Giloh et al., 2012). In birds, the tarsometatarsus is a vital organ that regulates thermal dissipation (Mosher and White, 1978). Considering that limited information is available on the circadian rhythm of tarsometatarsus temperature under dark conditions, how birds regulate their thermal equilibrium by adjusting the surface temperature of their peripheral organs remains to be elucidated.
5.
Conclusions
In this study, we examined the circadian rhythms of activity, body mass, and core and peripheral body temperatures of ETS in response to LD, LL, and DD conditions. We found that the core body temperature maintained significant rhythmicity when the external photoperiods changed; however, the amplitude of the concussion of each index under all treatments weakened, indicating that the core body temperature is an endogenous index. The peripheral body temperature and activity decoupled when the sparrows were exposed to constant light, whereas increased eye temperature enabled adequate food intake to maintain an energy balance under DD conditions. Therefore, our results suggest that the maintenance of the core body temperature rhythm is highly rhythmic and endogenous—it is not completely dependent on light; however, but the activity and peripheral body temperature rhythms are greatly affected by photoperiod. Our results differ from the research findings for other diurnal passerines such as House Sparrows and White-throated Sparrows. These discrepancies suggest that the coping mechanisms of body condition and core and peripheral body temperatures in response to constant light signals are species-dependent, not phylogeny-dependent. Our results contribute to a better understanding of the diversity and plasticity of physiology and behavior of passerines under abnormal photoperiods. The underlying causes of such species-dependent variations in the rhythmicity of physiology and behavior in response to unfavorable photoperiod warrant further study.
CRediT authorship contribution statement
Lirong Zuo: Formal analysis, Writing original draft, Writing review & editing. Ibrahim M. Ahmad: Formal analysis, Writing original draft, Writing review & editing. Yuanyuan Liu: Investigation, Methodology. Limin Wang: Investigation, Methodology. Shu Fang: Investigation, Methodology. Dongming Li: Conceptualization, Supervision, Funding acquisition, Writing review & editing.
Ethics statements
All experimental materials were conducted in accordance with the Institutional Committee for Animal Care and Use of Hebei Normal University, China, and was carried out under the auspices of scientific collecting permits issued by the Departments of Wildlife Conservation (Forestry Bureau) of Hebei Province, China.
Al-Hasani, H., Tschop, M.H., Cushman, S.W., 2003. Two birds with one stone: novel glucokinase activator stimulates glucose-induced pancreatic insulin secretion and augments hepatic glucose metabolism. Mol. Interv. 3, 367–370. .
Appenroth, D., Nord, A., Hazlerigg, D.G., Wagner, G.C., 2021. Body temperature and activity rhythms under different photoperiods in high arctic Svalbard ptarmigan (Lagopus muta hyperborea). Front. Physiol. 12, 633866. .
Benstaali, C., Mailloux, A., Bogdan, A., Auzeby, A., Touitou, Y., 2001. Circadian rhythms of body temperature and motor activity in rodents: their relationships with the light-dark cycle. Life Sci. 68, 2645–2656. .
Binkley, S., Kluth, E., Menaker, M., 1971. Pineal function in sparrows: circadian rhythms and body temperature. Science 174, 311–314. .
Binkley, S., Mosher, K., 1986. Photoperiod modifies circadian resetting responses in sparrows. Am. J. Physiol. 251, R1156-R1162. .
Borniger, J.C., Maurya, S.K., Periasamy, M., Nelson, R.J., 2014. Acute dim light at night increases body mass, alters metabolism, and shifts core body temperature circadian rhythms. Chronobiol. Int. 31, 917–925. .
Boshouwers, F., Nicaise, E., 1987. Physical activity and energy expenditure of laying hens as affected by light intensity. Br. Poultry Sci. 28, 155–163. .
Brandstaetter, R., 2002. The circadian pacemaking system of birds. In: Kumar, V. (Ed.), Biological Rhythms. Springer, Berlin, Heidelberg, pp. 144-163. .
Budki, P., Rani, S., Kumar, V., 2012. Persistence of circannual rhythms under constant periodic and aperiodic light conditions: sex differences and relationship with the external environment. J. Exp. Biol. 215, 3774–3785. .
Buhr, E.D., Takahashi, J.S., 2013. Molecular components of the mammalian circadian clock. In: Kramer, A., Merrow, M. (Eds.), Circadian Clocks, Handbook of Experimental Pharmacology, vol. 217. Springer, Berlin, pp. 3-27. .
Cassone, V.M., 2014. Avian circadian organization: a chorus of clocks. Front. Neuroendocrinol. 35, 76–88. .
Cassone, V.M., Paulose, J.K., Whitfield-Rucker, M.G., Peters, J.L., 2009. Time's arrow flies like a bird: two paradoxes for avian circadian biology. Gen. Comp. Endocrinol. 163, 109–116. .
Coomans, C.P., van den Berg, S.A., Houben, T., van Klinken, J.B., van den Berg, R., Pronk, A.C., et al., 2013. Detrimental effects of constant light exposure and high-fat diet on circadian energy metabolism and insulin sensitivity. Faseb. J. 27, 1721–1732. .
Coverdill, A.J., Bentley, G.E., Ramenofsky, M., 2008. Circadian and masking control of migratory restlessness in Gambel's white-crowned sparrow (Zonotrichia leucophrys gambelii). J. Biol. Rhythm. 23, 59–68. .
Dawson, A., King, V.M., Bentley, G.E., Ball, G.F., 2001. Photoperiodic control of seasonality in birds. J. Biol. Rhythm. 16, 365–380. .
Dawson, A., Sharp, P.J., 2007. Photorefractoriness in birds-photoperiodic and non-photoperiodic control. Gen. Comp. Endocrinol. 153, 378–384. .
Depres-Brummer, P., Levi, F., Metzger, G., Touitou, Y., 1995. Light-induced suppression of the rat circadian system. Am. J. Physiol. 268, R1111-R1116. .
Dixit, A.S., Byrsat, S., Kataki, B., 2022. Hypothalamic expression of GnRH-Ⅰ and GnIH in the Eurasian tree sparrow over a single long day. Photochem. Photobiol. Sci. 21, 147–158. .
Dixit, A.S., Singh, N.S., 2011. Photoperiod as a proximate factor in control of seasonality in the subtropical male Tree Sparrow, Passer montanus. Front. Zool. 8, 1. .
Dixit, A.S., Singh, N.S., 2012. Seasonal variation in sensitivity of the photoperiodic response system in the subtropical tree sparrow (Passer montanus). J. Exp. Zool. A Ecol. Genet. Physiol. 317, 488–498. .
Dominoni, D.M., 2015. The effects of light pollution on biological rhythms of birds: an integrated, mechanistic perspective. J. Ornithol. 156, 409–418. .
Ebihara, S., Kawamura, H., 1981. The role of the pineal organ and the suprachiasmatic nucleus in the control of circadian locomotor rhythms in the Java sparrow, Padda oryzivora. J. Comp. Physiol. 141, 207–214. .
Ebihara, S., Uchiyama, K., Oshima, I., 1984. Circadian organization in the pigeon, Columba livia: the role of the pineal organ and the eye. J. Comp. Physiol. 154, 59–69. .
Farsi, H., Harti, D., Achaaban, M.R., Piro, M., Raverot, V., Bothorel, B., et al., 2020. Melatonin rhythm and other outputs of the master circadian clock in the desert goat (Capra hircus) are entrained by daily cycles of ambient temperature. J. Pineal Res. 68, e12634. .
Fonken, L.K., Nelson, R.J., 2014. The effects of light at night on circadian clocks and metabolism. Endocr. Rev. 35, 648–670. .
Fonken, L.K., Workman, J.L., Walton, J.C., Weil, Z.M., Morris, J.S., Haim, A., et al., 2010. Light at night increases body mass by shifting the time of food intake. Proc. Natl. Acad. Sci. USA 107, 18664–18669. .
Gaston, S., Menaker, M., 1968. Pineal function: the biological clock in the sparrow? Science 160, 1125–1127. .
Giloh, M., Shinder, D., Yahav, S., 2012. Skin surface temperature of broiler chickens is correlated to body core temperature and is indicative of their thermoregulatory status. Poultry Sci. 91, 175–188. .
Grunst, A.S., Grunst, M.L., Raap, T., Pinxten, R., Eens, M., 2023. Anthropogenic noise and light pollution additively affect sleep behaviour in free-living birds in sex-and season-dependent fashions. Environ. Pollut. 316, 120426. .
Gwinner, E., 1978. Effects of pinealectomy on circadian locomotor activity rhythms in European starlings, Sturnus vulgaris. J. Comp. Physiol. 126, 123–129. .
Harmening, W.M., 2017. Contrast sensitivity and visual acuity in animals. Ophthalmologe 114, 986–996. .
Hohtola, E., Stevens, E.D., 1986. The relationship of muscle electrical activity, tremor and heat production to shivering thermogenesis in Japanese quail. J. Exp. Biol. 125, 119–135. .
Hughes, M.E., Hogenesch, J.B., Kornacker, K., 2010. JTK_CYCLE: an efficient nonparametric algorithm for detecting rhythmic components in genome-scale data sets. J. Biol. Rhythm. 25, 372–380. .
Hurly, T.A., 1992. Energetic reserves of marsh tits (Parus palustris): food and fat storage in response to variable food supply. Behav. Ecol. 3, 181–188. .
Jiang, J.X., He, Y., Kou, H.H., Ju, Z.Q., Gao, X.B., Zhao, H.F., 2020. The effects of artificial light at night on Eurasian tree sparrow (Passer montanus): behavioral rhythm disruption, melatonin suppression and intestinal microbiota alterations. Ecol. Indicat. 108. , 105702–105702.
Kontogiannis, J.E., 1967. Day and night changes in body weight of the White-throated Sparrow, Zonotrichia albicollis. Auk 84, 390–395. .
Le Tallec, T., Perret, M., Théry, M., 2013. Light pollution modifies the expression of daily rhythms and behavior patterns in a nocturnal primate. PLoS One 8, e79250. .
Li, D., Wang, G., Wingfield, J.C., Zhang, Z., Ding, C., Lei, F., 2008. Seasonal changes in adrenocortical responses to acute stress in Eurasian tree sparrow (Passer montanus) on the Tibetan Plateau: comparison with house sparrow (P. domesticus) in North America and with the migratory P. domesticus in Qinghai Province. Gen. Comp. Endocrinol. 158, 47–53. .
Li, D., Wu, J., Zhang, X., Ma, X., Wingfield, J.C., Lei, F., et al., 2011. Comparison of adrenocortical responses to acute stress in lowland and highland Eurasian tree sparrows (Passer montanus): similar patterns during the breeding, but different during the prebasic molt. J. Exp. Zool. A Ecol. Genet. Physiol. 315, 512–519. .
Li, D., Zhang, X., Li, Y., Hao, C., Zhang, J., Wu, Y., et al., 2012. Stress responses of testosterone and corticosterone-binding globulin in a multi-brooded species, Eurasian Tree Sparrows (Passer montanus): Does CBG function as a mediator? Horm. Behav. 61, 582–589. .
Li, Y., Sun, Y., Krause, J.S., Li, M., Liu, X., Zhu, W., et al., 2017. Dynamic interactions between corticosterone, corticosteroid binding globulin and testosterone in response to capture stress in male breeding Eurasian tree sparrows. Comp. Biochem. Physiol. A 205, 41–47. .
Lucia, M., Andre, J.M., Gonzalez, P., Baudrimont, M., Bernadet, M.D., Gontier, K., et al., 2010. Effect of dietary cadmium on lipid metabolism and storage of aquatic bird Cairina moschata. Ecotoxicology 19, 163–170. .
McMillan, J., 1972. Pinealectomy abolishes the circadian rhythm of migratory restlessness. J. Comp. Physiol. 79, 105–112. .
Montoya, A., Lopez-Olmeda, J.F., Garayzar, A.B., Sanchez-Vazquez, F.J., 2010. Synchronization of daily rhythms of locomotor activity and plasma glucose, cortisol and thyroid hormones to feeding in Gilthead seabream (Sparus aurata) under a light-dark cycle. Physiol. Behav. 101, 101–107. .
Mosher, J.A., White, C.M., 1978. Falcon temperature regulation. Auk 95, 80–84. .
Oshima, I., Yamada, H., Goto, M., Sato, K., Ebihara, S., 1989. Pineal and retinal melatonin is involved in the control of circadian locomotor activity and body temperature rhythms in the pigeon. J. Comp. Physiol. 166, 217–226. .
Pandey, R.K., Bhardwaj, S.K., 2011. Circadian and seasonal responses in Indian weaver bird: subjective interpretation of day and night depends upon both light intensity and contrast between illuminations. Chronobiol. Int. 28, 758–763. .
Plano, S.A., Casiraghi, L.P., García Moro, P., Paladino, N., Golombek, D.A., Chiesa, J.J., 2017. Circadian and metabolic effects of light: implications in weight homeostasis and health. Front. Neurol. 8, 558. .
Porcu, A., Riddle, M., Dulcis, D., Welsh, D.K., 2018. Photoperiod-induced neuroplasticity in the circadian system. Neural Plast. 2018, 5147585. .
Prabhat, A., Malik, I., Jha, N.A., Bhardwaj, S.K., Kumar, V., 2020. Developmental effects of constant light on circadian behaviour and gene expressions in zebra finches: Insights into mechanisms of metabolic adaptation to aperiodic environment in diurnal animals. J. Photochem. Photobiol. B 211, 111995. .
Ramkisoensing, A., Meijer, J., 2015. Synchronization of biological clock neurons by light and peripheral feedback systems promotes circadian rhythms and health. Front. Neurol. 6, 128. .
Ravikumar, G., Tewary, P.D., 1990. Photo refractoriness and its termination in the subtropical house sparrow, Passer domesticus: involvement of circadian rhythm. Chronobiol. Int. 7, 187–191. .
Refinetti, R., Menaker, M., 1992. The circadian rhythm of body temperature. Physiol. Behav. 51, 613–637. .
Regas, P.P., 1998. Circadian rhythms of body temperature and metabolic rate in the House Sparrow (Passer domesticus). Honors Theses. St. John's University, New York. .
Spoelstra, K., Verhagen, I., Meijer, D., Visser, M.E., 2018. Artificial light at night shifts daily activity patterns but not the internal clock in the great tit (Parus major). Proc. Biol. Sci. 285, 20172751. .
Stenvers, D.J., van Dorp, R., Foppen, E., Mendoza, J., Opperhuizen, A.L., Fliers, E., et al., 2016. Dim light at night disturbs the daily sleep-wake cycle in the rat. Sci. Rep. 6, 35662. .
Tosini, G., Menaker, M., 1995. Circadian rhythm of body temperature in an ectotherm (Iguana iguana). J. Biol. Rhythm. 10, 248–255. .
Trivedi, A.K., Rani, S., Kumar, V., 2005. Differential responses of the photoperiodic clock in two passerine birds possessing a strongly self-sustained circadian system. Chronobiol. Int. 22, 801–806. .
Underwood, H., Steele, C.T., Zivkovic, B., 1999. Effects of fasting on the circadian body temperature rhythm of Japanese quail. Physiol. Behav. 66, 137–143. .
van Jaarsveld, B., Bennett, N.C., Hart, D.W., Oosthuizen, M.K., 2019. Locomotor activity and body temperature rhythms in the Mahali mole-rat (Cryptomys hottentotus mahali): The effect of light and ambient temperature variations. J. Therm. Biol. 79, 24–32. .
Wang, L., Nabi, G., Zhang, L., Liu, D., Li, M., Li, J., et al., 2022. Seasonal variations in gonad morphology and hypothalamic GnRH-Ⅰ and GnIH in Eurasian Tree Sparrow, a multi-brooded passerine. Avian Res. 13, 100037. .
West, A.C., Smith, L., Ray, D.W., Loudon, A.S.I., Brown, T.M., Bechtold, D.A., 2017. Misalignment with the external light environment drives metabolic and cardiac dysfunction. Nat. Commun. 8, 417. .
Wideman, C.H., Murphy, H.M., 2009. Constant light induces alterations in melatonin levels, food intake, feed efficiency, visceral adiposity, and circadian rhythms in rats. Nutr. Neurosci. 12, 233–240. .
Wyse, C.A., Selman, C., Page, M.M., Coogan, A.N., Hazlerigg, D.G., 2011. Circadian desynchrony and metabolic dysfunction; did light pollution make us fat? Med. Hypotheses 77, 1139–1144. .
Yamada, H., Oshima, I., Sato, K., Ebihara, S., 1988. Loss of the circadian rhythms of locomotor activity, food intake, and plasma melatonin concentration induced by constant bright light in the pigeon (Columba livia). J. Comp. Physiol. 163, 459–463. .
Zhang, X.J., Yang, W.Y., Liang, W., Wang, Y., Zhang, S.P., 2019. Intensity dependent disruptive effects of light at night on activation of the HPG axis of tree sparrows (Passer montanus). Environ. Pollut. 249, 904–909. .
Jinjie He, Chang Wang, Ying Han, et al. Waterbird community response to wetland and climate changes in the Liaohe River Estuary wetlands, China. Journal of Environmental Management, 2025, 374: 124165.
DOI:10.1016/j.jenvman.2025.124165
2.
Rong Ma, Qi Xu, Yongqian Gao, et al. Patterns and drivers of plant sexual systems in the dry-hot valley region of southwestern China. Plant Diversity, 2024, 46(2): 158.
DOI:10.1016/j.pld.2023.07.010
3.
Dawei Wu, Taiyu Chen, Wei Hu, et al. Impact of active vegetation restoration and native vegetation on breeding bird communities in coastal wetlands under disturbance by reintroduced Père David’s deer. Global Ecology and Conservation, 2024, 56: e03345.
DOI:10.1016/j.gecco.2024.e03345
4.
Lingxuan Gong, Lili Wang, Yao Yao, et al. Integrity and Size of Green Spaces Are Critical for the Functional Diversity of Birds: Evidence from the Eastern and Northwestern Regions of China. Chinese Geographical Science, 2024, 34(6): 1073.
DOI:10.1007/s11769-024-1474-1
5.
Xiangyi Li, Zhaojun Li, Yu Chen, et al. Enhancing Bird Diversity in Urban Parks: Insights from the Futian Mangrove Ecological Park, Shenzhen. Forests, 2024, 15(12): 2088.
DOI:10.3390/f15122088
6.
Binaya Adhikari, Suresh C. Subedi, Shivish Bhandari, et al. Climate‐driven decline in the habitat of the endemic spiny babbler (Turdoides nipalensis). Ecosphere, 2023, 14(6)
DOI:10.1002/ecs2.4584
7.
Jonathan O. Hernandez, Muhammad Naeem, Wajid Zaman. How Does Changing Environment Influence Plant Seed Movements as Populations of Dispersal Vectors Decline?. Plants, 2023, 12(7): 1462.
DOI:10.3390/plants12071462
8.
Lu Feng, Xiaoming Ma, Alice C. Hughes, et al. Elevation range and contemporary climate determine the taxonomic, functional and phylogenetic diversity of forest mammals. Biodiversity and Conservation, 2023, 32(14): 4651.
DOI:10.1007/s10531-023-02715-7
9.
Muhammad Nawaz Rajpar, Shahab Ali Khan, Allah Ditta, et al. Subtropical Broad-Leaved Urban Forests as the Foremost Dynamic and Complex Habitats for a Wide Range of Bird Species. Sustainability, 2021, 13(23): 13021.
DOI:10.3390/su132313021
10.
O. Serebryakov, E. Turchaninova. INFLUENCE OF RECREATIONAL LOAD ON ORNITOFAUNA OF THE CITY OF VORONEZH. Materials of the All-Russian Youth Scientific and Practical Conference "Modern problems of the ecology of the animal and plant world",
DOI:10.34220/MPEAPW2021_88-93
Table
1.
The statistical results of treatment, time, and the interaction of treatment and time on activity, body mass, core temperature, eye temperature, and tarsometatarsus temperature of Eurasian Tree Sparrows (Passer montanus) in mixed linear models.
Table
2.
Statistical results of the average of activity, body mass, core temperature, tarsometatarsus temperature, and eye temperature of Eurasian Tree Sparrows (Passer montanus) under constant light (LL) and darkness (DD), and normal light-dark conditions (LD) at different time points (2:00, 6:00, 10:00, 14:00, 18:00, and 22:00).
Table
3.
Circadian rhythm parameters of activity, body mass, core temperature, tarsometatarsus temperature, and eye temperature of Eurasian Tree Sparrows (Passer montanus) under constant light (LL) and darkness (DD), and normal light-dark cycle (LD) conditions.
Factors
Variable
BH.Q
ADJ.P
Acrophase (φ, h)
Amplitude (A)
df
F
P
Activity
LL
1.000
1.000
22
6.393
5275
0.725
0.606
LD
< 0.05
< 0.001
12
34.850
5276
24.163
< 0.001
DD
0.072
0.035
12
5.922
5276
1.184
0.323
Body mass
LL
1.000
0.759
16
0.362
5364
3.027
0.013
LD
0.679
0.144
16
0.540
5372
8.655
< 0.001
DD
1.000
0.882
16
0.360
5401
2.089
0.071
Core temperature
LL
< 0.05
< 0.001
14
0.478
5413
7.669
< 0.001
LD
< 0.05
< 0.001
12
0.889
5422
37.124
< 0.001
DD
< 0.05
< 0.001
12
0.714
5444
32.552
< 0.001
Tarsometatarsus temperature
LL
1.000
1.000
12
0.191
5391
0.480
0.790
LD
< 0.05
< 0.001
12
1.332
5363
9.814
< 0.001
DD
0.029
0.019
16
0.470
5190
0.878
0.502
Eye temperature
LL
0.170
0.113
18
0.452
5388
0.587
0.710
LD
< 0.05
< 0.001
14
1.048
5363
24.730
< 0.001
DD
0.793
0.793
14
0.171
5190
0.243
0.941
Significant values (BH.Q < 0.05; P < 0.05) are shown in bold.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds